

Automatic Control of Chemolithotrophic Cultivation of Cupriavidus necator: Optimization of Oxygen Supply for Enhanced Bioplastic Production

 , , ,
, , ,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals, Assays and Organisms
2.2. Media and Inoculum
2.3. Safety Considerations
2.3.1. Room and Ventilated Hood
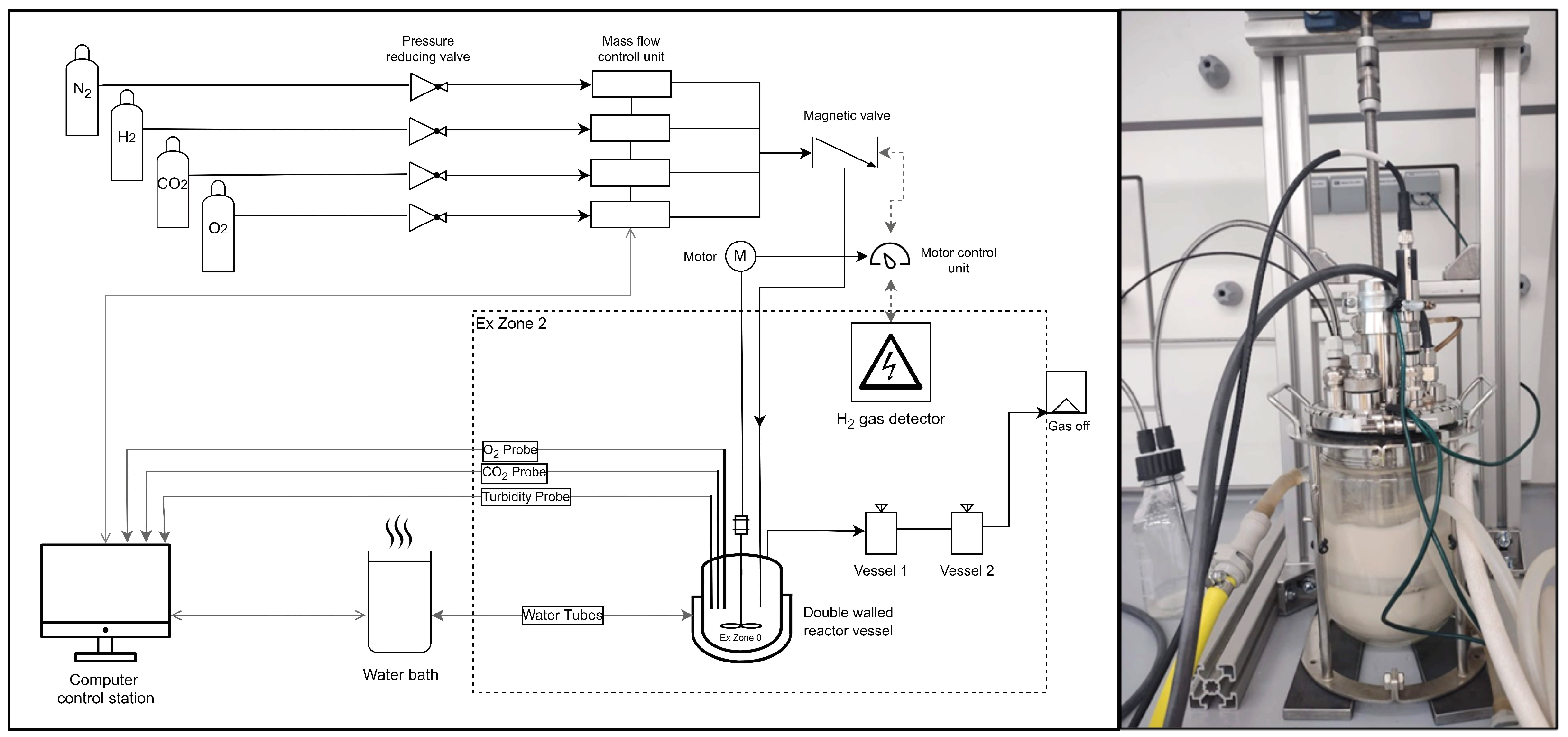
2.3.2. Bioreactor Setup: Pipe and Flow Diagram of the Lab
2.3.3. Personal Protective Measures
2.4. Operation of the Gas Cultivation System
2.5. Data Acquisition and Processing
2.6. Analytics
2.6.1. Optical Density and Cell Dry Mass
2.6.2. PHA Determination
3. Theory/Calculation
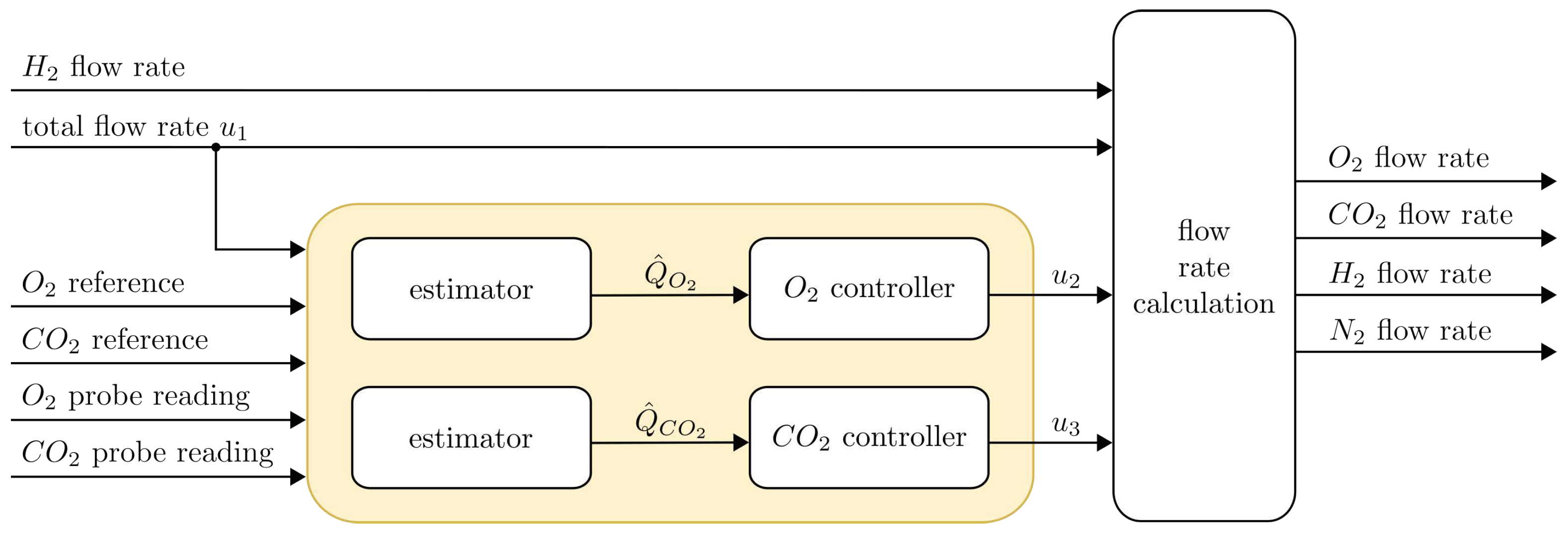
3.1. Mathematical Modelling and Automatic Control Problem Formulation
3.2. Controller Design
3.2.1. O2 Controller
3.2.2. CO2 Controller
3.2.3. Flow Rate Calculation
3.2.4. Estimator Design
3.2.5. Parameter Identification
3.2.6. Controller Parameters
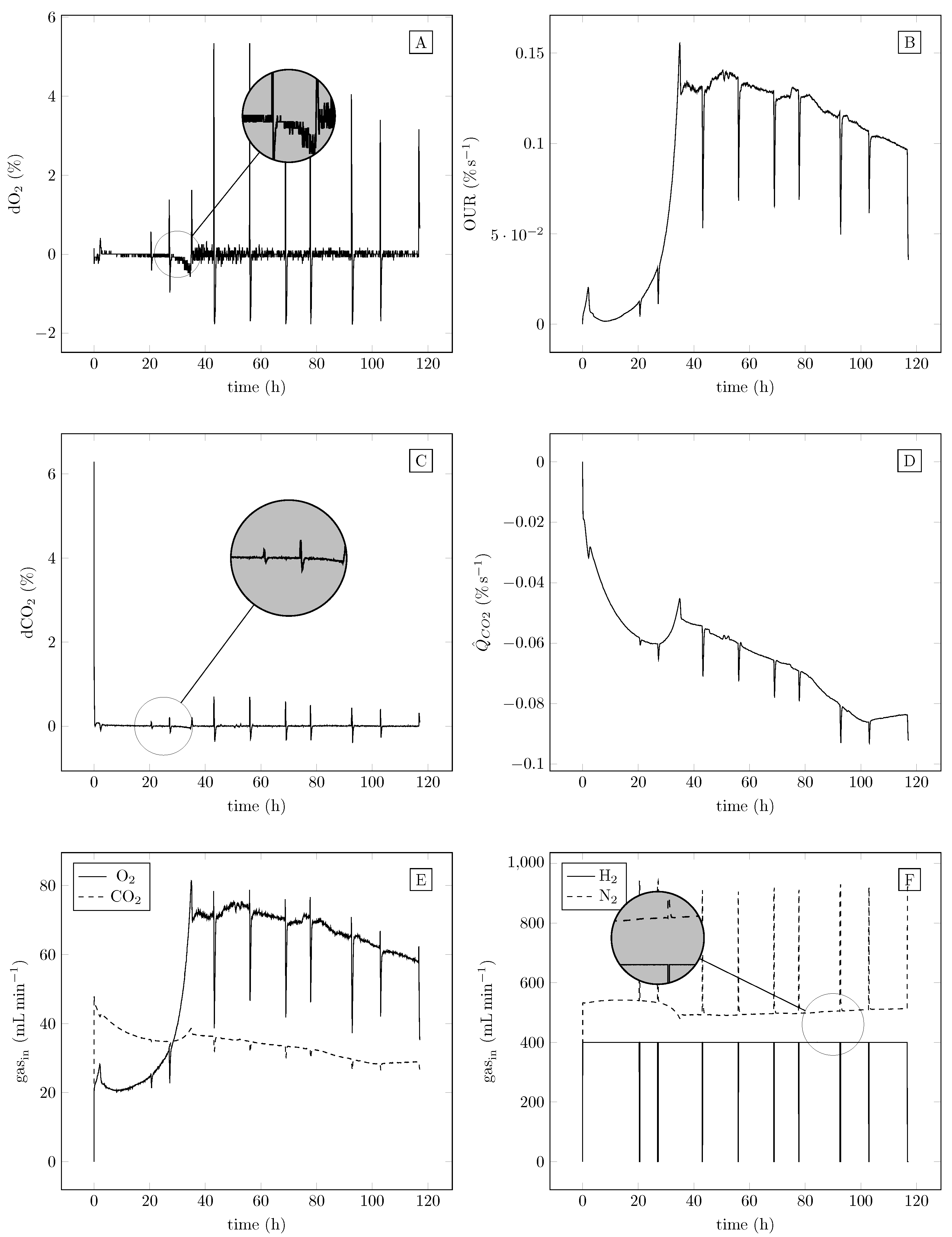
4. Results
4.1. Parameter Identification
4.2. Experimental Evaluation
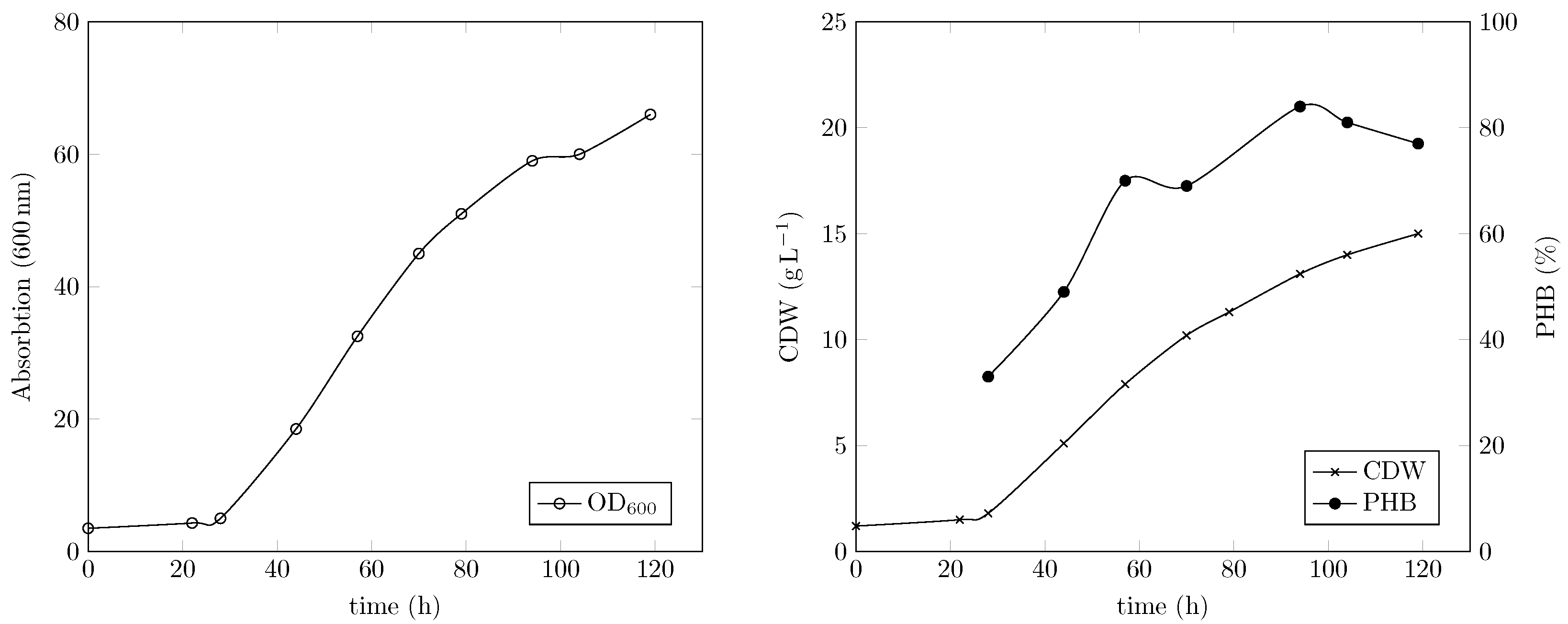
4.2.1. Fermentation Results and PHB Output
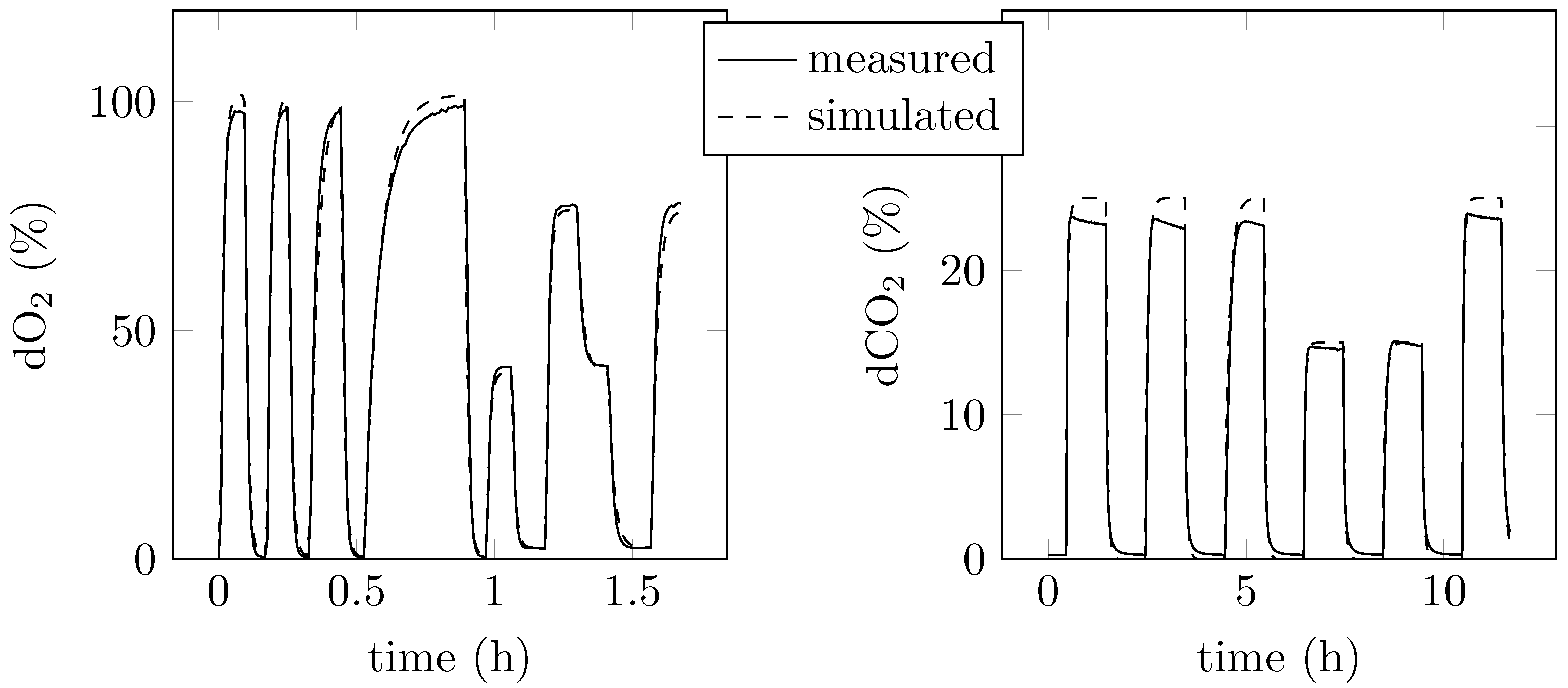
4.2.2. Data and Controller Performance
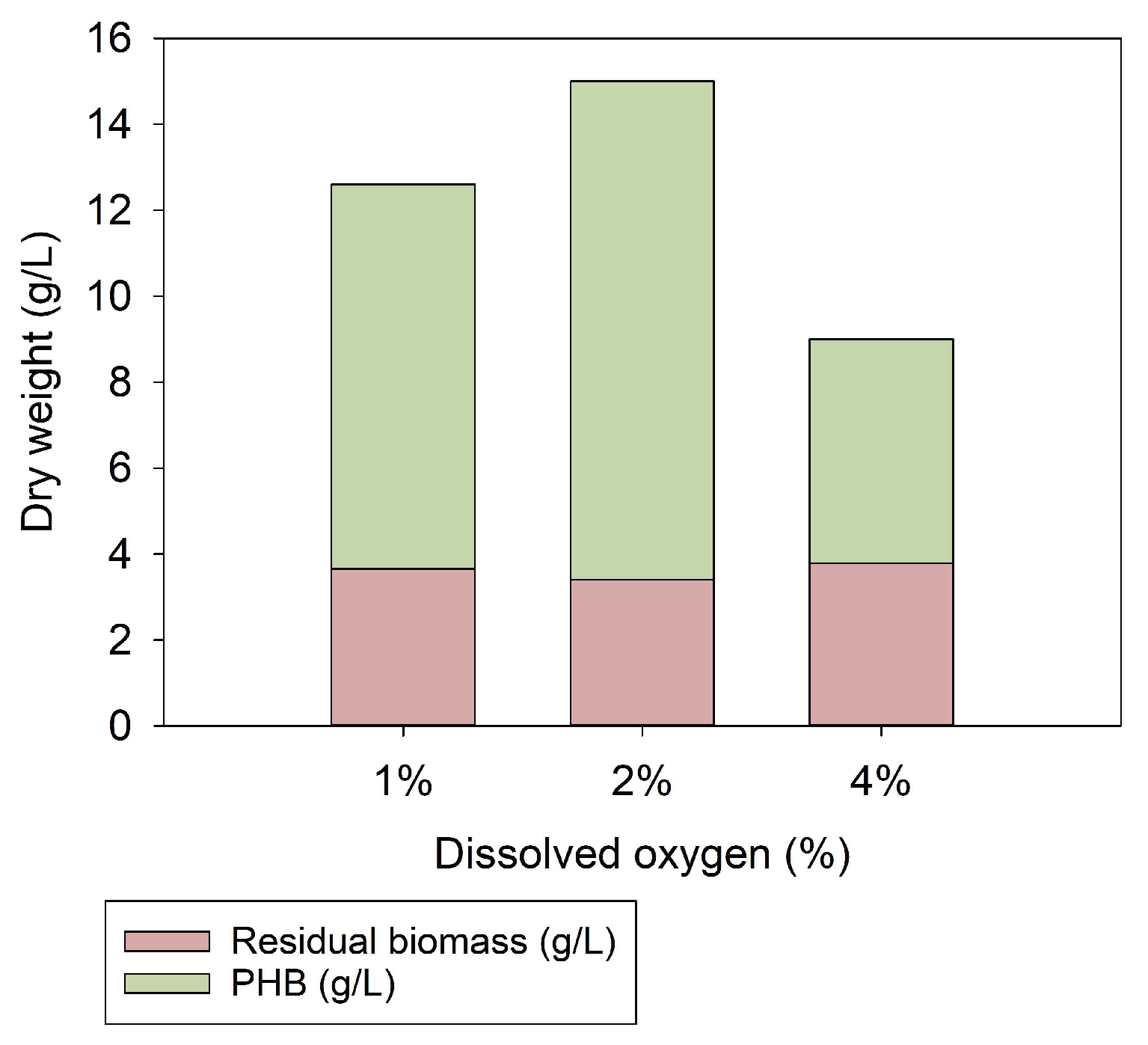
4.2.3. Gas Fermentation at Different dO2 Concentrations
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CDW | Cell dry weight (g L−1) |
| OD600 | Optical density at 600 nm |
| PHB | Poly-(R)-3-hydroxybutyrate |
| PHA | Polyhydroxyalkanoates |
| ATEX | Abbreviation from french “atmosphères explosibles” = explosive atmosphere |
| HOBs | Hydrogen-oxidizing bacteria |
| MFCs | Mass flow controllers (mL min−1) |
| dO2 | Dissolved oxygen concentration (%) |
| dCO2 | Dissolved carbon dioxide concentration (%) |
| LEL | Lower explosion limit |
References
- Honegger, M.; Michaelowa, A.; Roy, J. Potential implications of carbon dioxide removal for the sustainable development goals. Clim. Policy 2021, 21, 678–698. [Google Scholar] [CrossRef]
- Hilaire, J.; Minx, J.C.; Callaghan, M.W.; Edmonds, J.; Luderer, G.; Nemet, G.F.; Rogelj, J.; del Mar Zamora, M. Negative emissions and international climate goals-learning from and about mitigation scenarios. Clim. Chang. 2019, 157, 189–219. [Google Scholar] [CrossRef]
- Bistline, J.E.T.; Blanford, G.J. Impact of carbon dioxide removal technologies on deep decarbonization of the electric power sector. Nat. Commun. 2021, 12, 3732. [Google Scholar] [CrossRef]
- Ghiat, I.; Mahmood, F.; Govindan, R.; Al-Ansari, T. CO2 utilisation in agricultural greenhouses: A novel ‘plant to plant’ approach driven by bioenergy with carbon capture systems within the energy, water and food Nexus. Energy Convers. Manag. 2021, 228, 113668. [Google Scholar] [CrossRef]
- Anwar, M.; Fayyaz, A.; Sohail, N.; Khokhar, M.; Baqar, M.; Yasar, A.; Rasool, K.; Nazir, A.; Raja, M.; Rehan, M.; et al. CO2 utilization: Turning greenhouse gas into fuels and valuable products. J. Environ. Manag. 2020, 260, 110059. [Google Scholar] [CrossRef]
- Lu, Y.; Yu, J. Comparison analysis on the energy efficiencies and biomass yields in microbial CO2 fixation. Process. Biochem. 2017, 62, 151–160. [Google Scholar] [CrossRef]
- Köpke, M.; Simpson, S.D. Pollution to products: Recycling of ‘above ground’ carbon by gas fermentation. Curr. Opin. Biotechnol. 2020, 65, 180–189. [Google Scholar] [CrossRef]
- Fackler, N.; Heijstra, B.D.; Rasor, B.J.; Brown, H.; Martin, J.; Ni, Z.; Shebek, K.M.; Rosin, R.R.; Simpson, S.D.; Tyo, K.E.; et al. Stepping on the gas to a circular economy: Accelerating development of carbon-negative chemical production from gas fermentation. Annu. Rev. Chem. Biomol. Eng. 2021, 12, 439–470. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.V.; Moutinho, L.F.; Romão-Dumaresq, A.S. Gas fermentation of C1 feedstocks: Commercialization status and future prospects. Biofuels Bioprod. Biorefining 2018, 12, 1103–1117. [Google Scholar] [CrossRef]
- Kusian, B.; Bowien, B. Organization and regulation of cbb CO2 assimilation genes in autotrophic bacteria. FEMS Microbiol. Rev. 1997, 21, 135–155. [Google Scholar] [CrossRef]
- Pohlmann, A.; Fricke, W.F.; Reinecke, F.; Kusian, B.; Liesegang, H.; Cramm, R.; Eitinger, T.; Ewering, C.; Pötter, M.; Schwartz, E.; et al. Genome sequence of the bioplastic-producing “Knallgas” bacterium Ralstonia eutropha H16. Nat. Biotechnol. 2006, 24, 1257–1262. [Google Scholar] [CrossRef]
- Sohn, Y.J.; Son, J.; Jo, S.Y.; Park, S.Y.; Yoo, J.I.; Baritugo, K.A.; Na, J.G.; il Choi, J.; Kim, H.T.; Joo, J.C.; et al. Chemoautotroph Cupriavidus necator as a potential game-changer for global warming and plastic waste problem: A review. Bioresour. Technol. 2021, 340, 125693. [Google Scholar] [CrossRef] [PubMed]
- Markl, E. PHB - Bio based and biodegradable replacement for PP: A review. Nov. Tech. Nutr. Food Sci. 2018, 2, 206–209. [Google Scholar] [CrossRef]
- Lambauer, V.; Kratzer, R. Lab-scale cultivation of Cupriavidus necator on explosive gas mixtures: Carbon dioxide fixation into polyhydroxybutyrate. Bioengineering 2022, 9, 204. [Google Scholar] [CrossRef]
- Schröder, V. Explosionsgrenzen von Wasserstoff und Wasserstoff/Methan-Gemischen; Technical Report; Wirtschaftsverlag NW: Bremerhaven, Germany, 2003. [Google Scholar]
- Tanaka, K.; Ishizaki, A.; Kanamaru, T.; Kawano, T. Production of poly(D-3-hydroxybutyrate) from CO2, H2, and O2 by high cell density autotrophic cultivation of Alcaligenes eutrophus. Biotechnol. Bioeng. 1995, 45, 268–275. [Google Scholar] [CrossRef]
- Garrigues, L.; Maignien, L.; Lombard, E.; Singh, J.; Guillouet, S.E. Isopropanol production from carbon dioxide in Cupriavidus necator in a pressurized bioreactor. New Biotechnol. 2020, 56, 16–20. [Google Scholar] [CrossRef]
- Sonnleitner, B.; Heinzle, E.; Braunegg, G.; Lafferty, R.M. Formal kinetics of poly-β-hydroxybutyric acid (PHB) production in Alcaligenes eutrophus H 16 and Mycoplana rubra R 14 with respect to the dissolved oxygen tension in ammonium-limited batch cultures. Eur. J. Appl. Microbiol. Biotechnol. 1979, 7, 1–10. [Google Scholar] [CrossRef]
- Islam Mozumder, M.S.; Garcia-Gonzalez, L.; Wever, H.D.; Volcke, E.I. Poly(3-hydroxybutyrate) (PHB) production from CO2: Model development and process optimization. Biochem. Eng. J. 2015, 98, 107–116. [Google Scholar] [CrossRef]
- Juengert, J.; Bresan, S.; Jendrossek, D. Determination of polyhydroxybutyrate (PHB) content in Ralstonia eutropha using gas chromatography and Nile red staining. Bio Protoc. 2018, 8, e2748. [Google Scholar] [CrossRef]
- Lewis, W.K.; Whitman, W.G. Principles of gas absorption. Ind. Eng. Chem. 1924, 16, 1215–1220. [Google Scholar] [CrossRef]
- Whitman, W.G. The two film theory of gas absorption. Int. J. Heat Mass Transf. 1962, 5, 429–433. [Google Scholar] [CrossRef]
- Lagneau, V.; Pipart, A.; Catalette, H. Reactive transportmodelling and long term behaviour of CO2 sequestration in saline aquifers. Oil Gas Sci. Technol.—Rev. IFP 2005, 60, 231–247. [Google Scholar] [CrossRef]
- Pedersen, O.; Colmer, T.; Sand-Jensen, K. Underwater photosynthesis of submerged plants—Recent advances and methods. Front. Plant Sci. 2013, 4, 140. [Google Scholar] [CrossRef] [PubMed]
- Doran, P.M. Chapter 10—Mass Transfer. In Bioprocess Engineering Principles, 2nd ed.; Doran, P.M., Ed.; Academic Press: London, UK, 2013; pp. 379–444. [Google Scholar] [CrossRef]
- Åström, K.J.; Hägglund, T.; Astrom, K.J. Advanced PID Control; ISA—The Instrumentation, Systems, and Automation Society: Research Triangle Park, NC, USA, 2006; Volume 461. [Google Scholar]
- Hippe, P. Windup in Control: Its Effects and Their Prevention; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Simon, D. Optimal State Estimation: Kalman, H Infinity, and Nonlinear Approaches; Wiley-Interscience: Hoboken, NJ, USA, 2006. [Google Scholar] [CrossRef]
- Lu, Y.; Yu, J. Gas mass transfer with microbial CO2 fixation and poly(3-hydroxybutyrate) synthesis in a packed bed bioreactor. Biochem. Eng. J. 2017, 122, 13–21. [Google Scholar] [CrossRef]
- Milder, R.; Kramer, W.; Doblhoff-Dier, O.; Bayer, K. Strategies for optimal dissolved oxygen (DO) control. IFAC Proc. Vol. 1995, 28, 292–295, In Proceedings of the 6th International Conference on Computer Applications in Biotechnology (CAB 6), Garmisch-Partenkirchen, Germany, 14–17 May 1995. [Google Scholar] [CrossRef]
- Atlić, A.; Koller, M.; Scherzer, D.; Kutschera, C.; Grillo-Fernandes, E.; Horvat, P.; Chiellini, E.; Braunegg, G. Continuous production of poly([R]-3-hydroxybutyrate) by Cupriavidus necator in a multistage bioreactor cascade. Appl. Microbiol. Biotechnol. 2011, 91, 295–304. [Google Scholar] [CrossRef]
- Lukey, M.J.; Roessler, M.M.; Parkin, A.; Evans, R.M.; Davies, R.A.; Lenz, O.; Friedrich, B.; Sargent, F.; Armstrong, F.A. Oxygen-tolerant [NiFe]-hydrogenases: The individual and collective importance of supernumerary cysteines at the proximal Fe-S cluster. J. Am. Chem. Soc. 2011, 133, 16881–16892. [Google Scholar] [CrossRef]
- Milker, S.; Sydow, A.; Torres-Monroy, I.; Jach, G.; Faust, F.; Kranz, L.; Tkatschuk, L.; Holtmann, D. Gram-scale production of the sesquiterpene α-humulene with Cupriavidus necator. Biotechnol. Bioeng. 2021, 118, 2694–2702. [Google Scholar] [CrossRef]
- Nyyssölä, A.; Ojala, L.S.; Wuokko, M.; Peddinti, G.; Tamminen, A.; Tsitko, I.; Nordlund, E.; Lienemann, M. Production of endotoxin-free microbial biomass for food applications by gas fermentation of Gram positive H2-oxidizing bacteria. ACS Food Sci. Technol. 2021, 1, 470–479. [Google Scholar] [CrossRef]
- Lai, Y.H.; Lan, J.C.W. Enhanced polyhydroxybutyrate production through incorporation of a hydrogen fuel cell and electro-fermentation system. Int. J. Hydrog. Energy 2021, 46, 16787–16800, In Proceedings of the SEGT-2019, Bangkok, Thailand, 11–14 December 2019. [Google Scholar] [CrossRef]
- Nangle, S.N.; Ziesack, M.; Buckley, S.; Trivedi, D.; Loh, D.M.; Nocera, D.G.; Silver, P.A. Valorization of CO2 through lithoautotrophic production of sustainable chemicals in Cupriavidus necator. Metab. Eng. 2020, 62, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Yoshida, K.; Orita, I.; Fukui, T. Biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from CO2 by a recombinant Cupriavidus necator. Bioengineering 2021, 8, 179. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Yu, R.; Xu, G.; Li, X.; Qiu, Y.; Wang, D.; Li, B.; Xie, L. Theoretical and experimental investigation of explosion characteristics of hydrogen explosion in a closed vessel. Energies 2022, 15, 8630. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Explanation | |
|---|---|---|
| States | O2 dissolved concentration | |
| CO2 dissolved concentration | ||
| HCO3− concentration | ||
| CO2 probe reading | ||
| Inputs | Total inlet gas flow | |
| O2 concentration in the inlet gas | ||
| CO2 concentration in the inlet gas | ||
| Parameters | O2 or CO2 mass transfer rate coefficient as a function of | |
| , | O2 or CO2 model parameters | |
| Reaction rate from CO2 to HCO3− | ||
| Reverse reaction rate from HCO3− to CO2 | ||
| pH-dependent constant | ||
| Reciprocal of the CO2 probe reading time constant | ||
| Dis * | O2 estimator | |
| CO2 estimator |
| O2 model | |||||
| 0.0822 | 9.4149 | ||||
| CO2 model | |||||
| 0.1440 | −1.2616 | 0.0185 | 6.4943 · 104 | 0.0167 |
| 0.02 | 0 | 0.0001 | 0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambauer, V.; Permann, A.; Petrášek, Z.; Subotić, V.; Hochenauer, C.; Kratzer, R.; Reichhartinger, M. Automatic Control of Chemolithotrophic Cultivation of Cupriavidus necator: Optimization of Oxygen Supply for Enhanced Bioplastic Production. Fermentation 2023, 9, 619. https://doi.org/10.3390/fermentation9070619
Lambauer V, Permann A, Petrášek Z, Subotić V, Hochenauer C, Kratzer R, Reichhartinger M. Automatic Control of Chemolithotrophic Cultivation of Cupriavidus necator: Optimization of Oxygen Supply for Enhanced Bioplastic Production. Fermentation. 2023; 9(7):619. https://doi.org/10.3390/fermentation9070619
Chicago/Turabian StyleLambauer, Vera, Alexander Permann, Zdeněk Petrášek, Vanja Subotić, Christoph Hochenauer, Regina Kratzer, and Markus Reichhartinger. 2023. "Automatic Control of Chemolithotrophic Cultivation of Cupriavidus necator: Optimization of Oxygen Supply for Enhanced Bioplastic Production" Fermentation 9, no. 7: 619. https://doi.org/10.3390/fermentation9070619
APA StyleLambauer, V., Permann, A., Petrášek, Z., Subotić, V., Hochenauer, C., Kratzer, R., & Reichhartinger, M. (2023). Automatic Control of Chemolithotrophic Cultivation of Cupriavidus necator: Optimization of Oxygen Supply for Enhanced Bioplastic Production. Fermentation, 9(7), 619. https://doi.org/10.3390/fermentation9070619