

The Application of Moringa oleifera Leaf Meal and Its Fermentation Products in the Diet of Megalobrama amblycephala Juveniles


Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Experimental Diets
2.3. Experimental Fish
2.4. Sample Collection
2.5. Laboratory Analysis
2.5.1. Growth Performance
2.5.2. Determination of Feed and Muscle Composition
2.5.3. Determination of Plasma Biochemical Parameters
2.5.4. Determination of Hepatic Antioxidant Parameters
2.5.5. Real-Time PCR (qRT-PCR) Analysis on Genes Relative Expressions
2.6. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Muscle Nutrient Composition
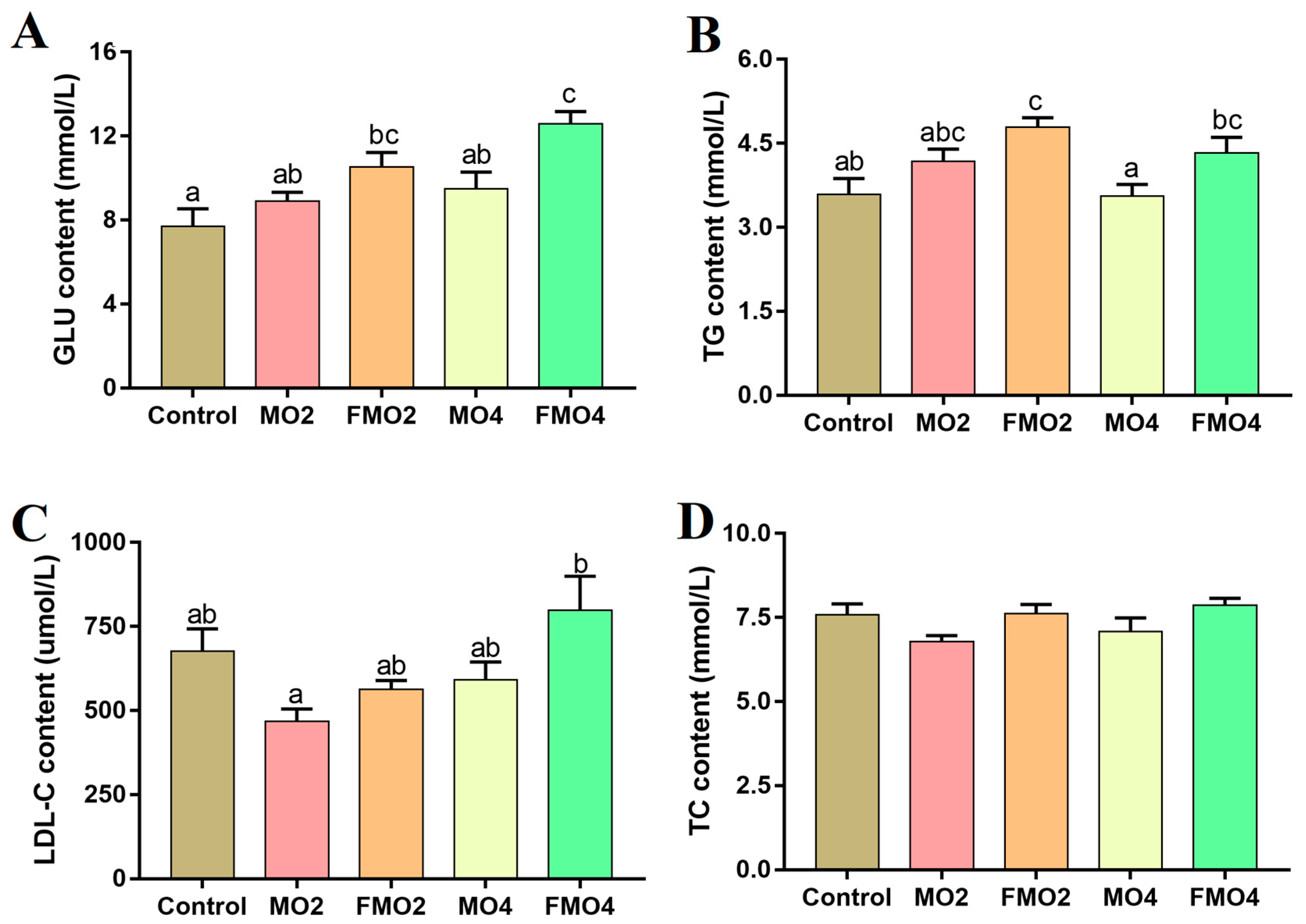
3.3. Plasma Biochemical Indices
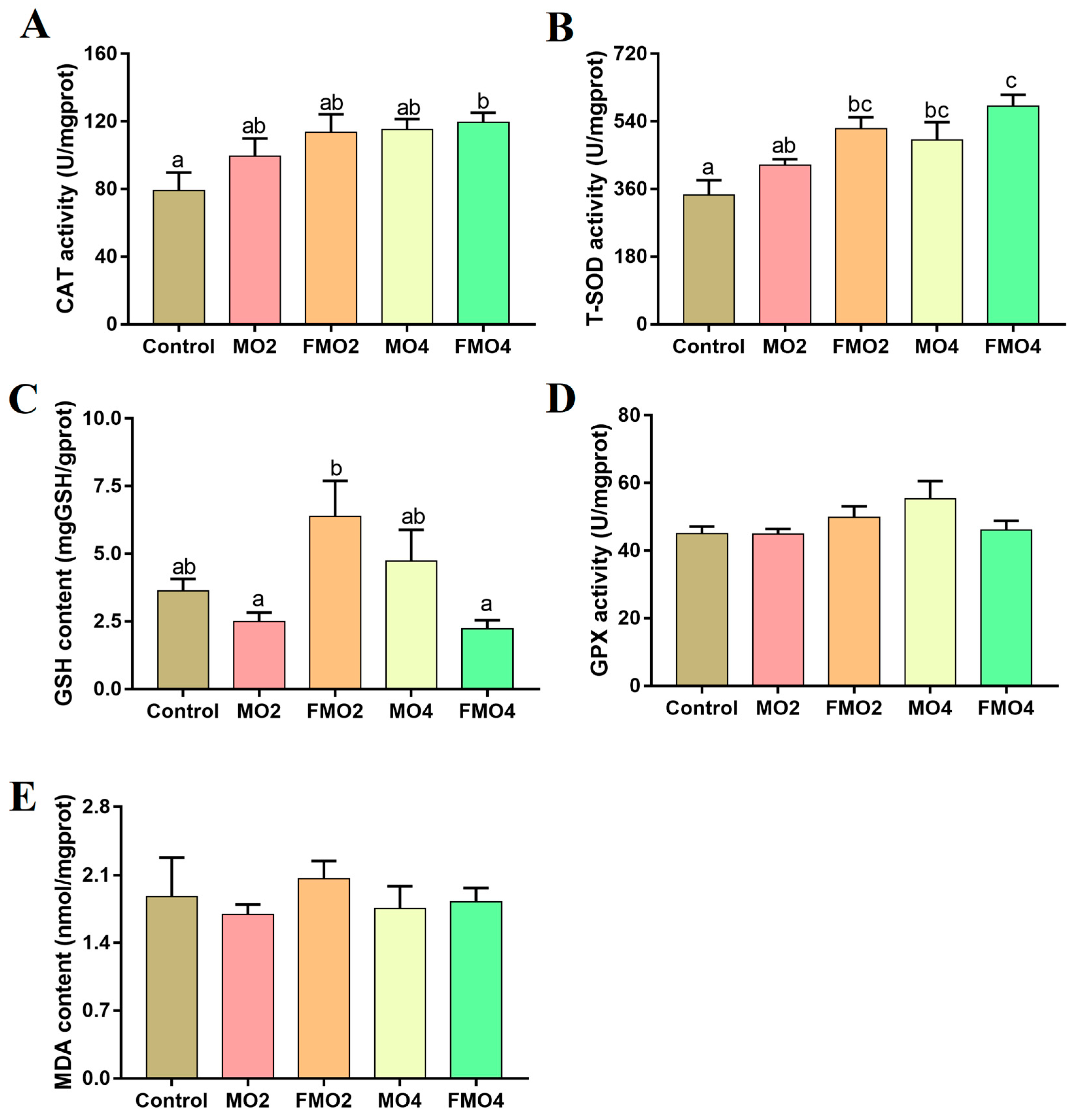
3.4. Hepatic Antioxidant Parameters
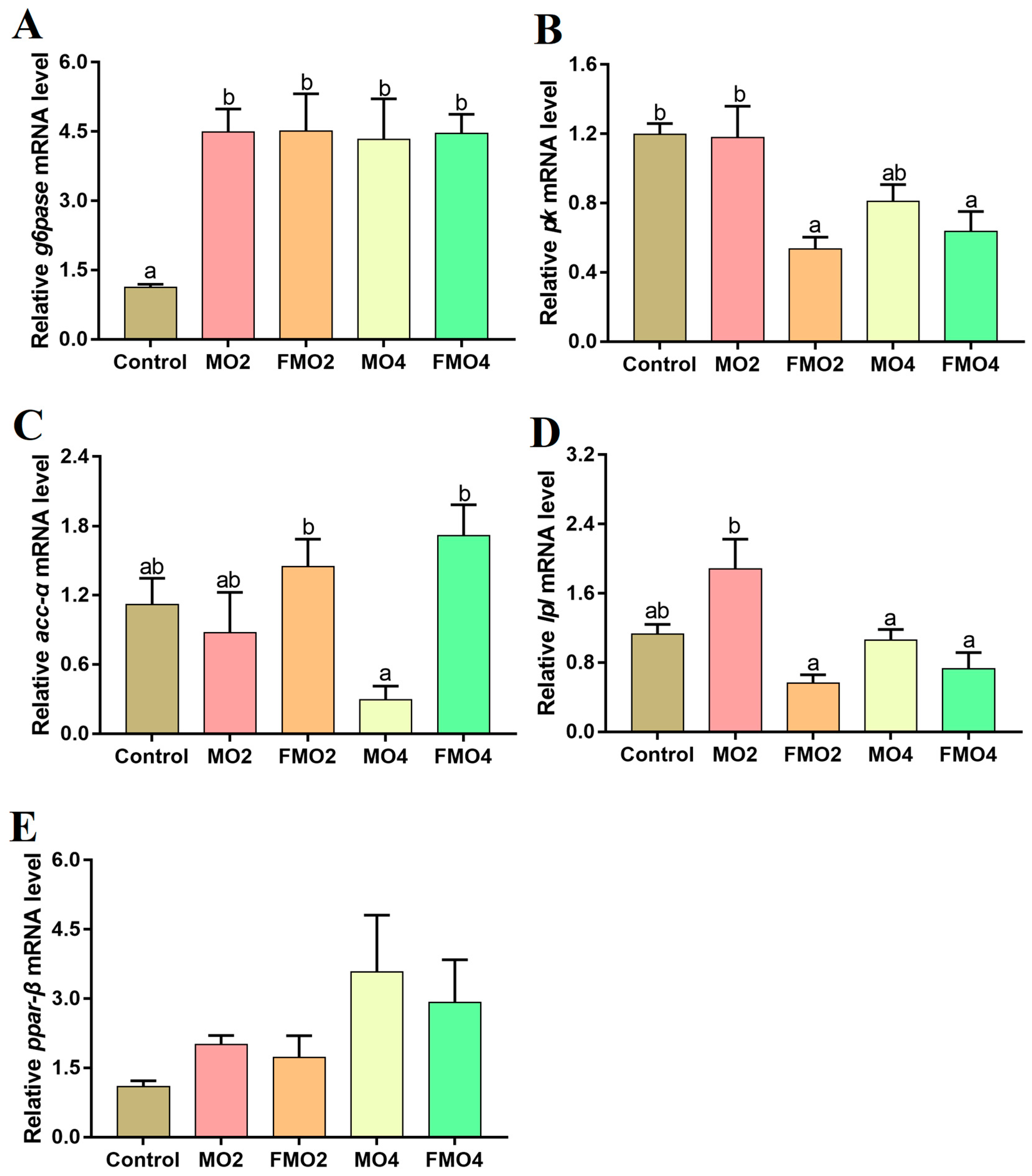
3.5. Gene Expressions Related to Glucose and Lipid Metabolism in the Liver
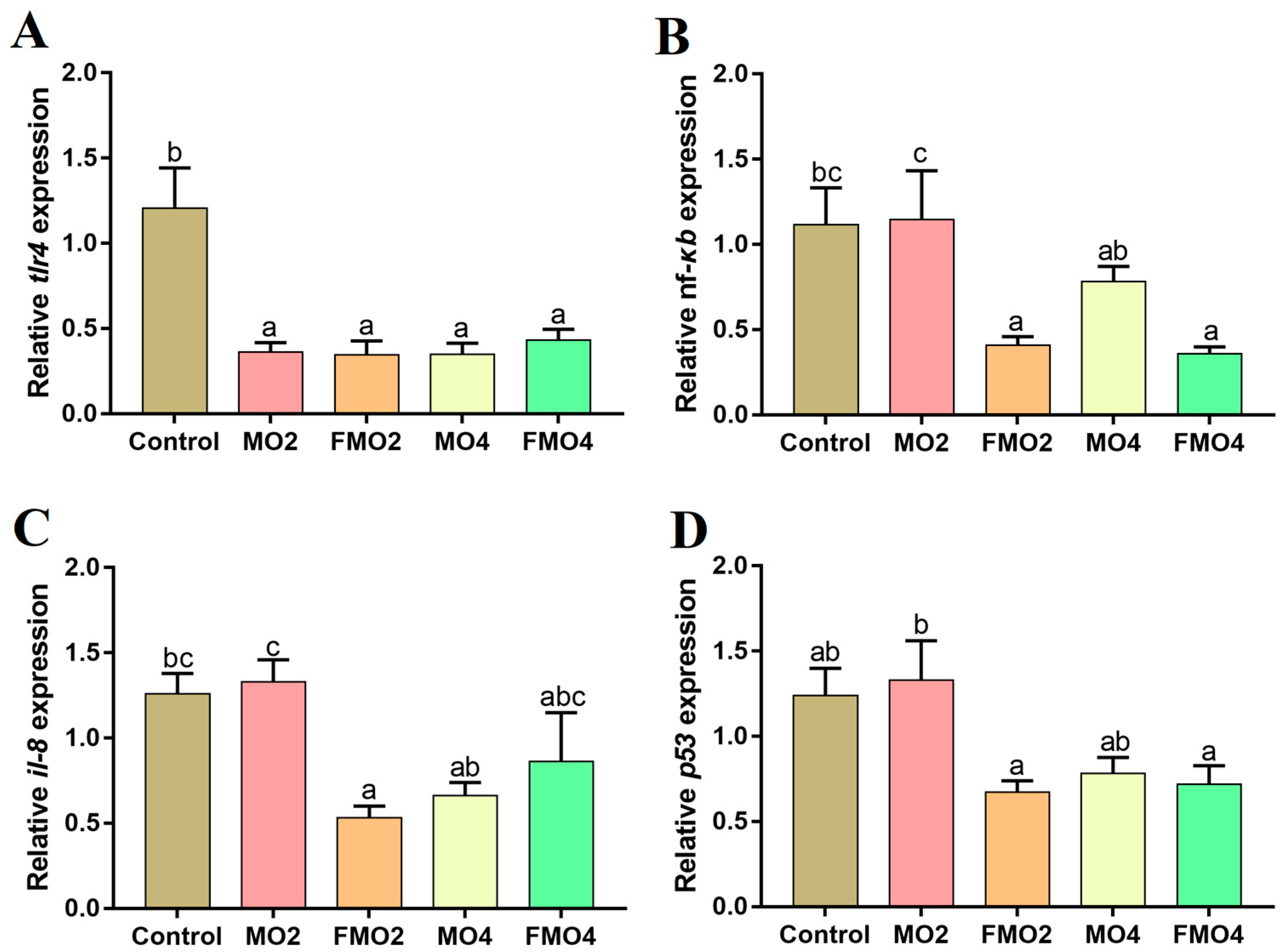
3.6. Gene mRNA Expressions Related to the Inflammatory Response in the Liver
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAO fisheries and aquaculture information and statistics services. In Aquaculture Production: Quantities; FAO: Rome, Italy, 2020; pp. 1950–2016. [Google Scholar]
- Dai, J.; Chen, T.; Guo, X.; Dai, Z.; He, Z.; Hu, Y. Evaluation of fish meal replacement by Clostridium autoethanogenum protein in diets for juvenile red swamp crayfish (Procambarus clarkii). Aquaculture 2023, 570, 739379. [Google Scholar] [CrossRef]
- Yuan, X.Y.; Liu, W.B.; Liang, C.; Sun, C.X.; Xue, Y.F.; Wan, Z.D.; Jiang, G.Z. Effects of partial replacement of fish meal by yeast hydrolysate on complement system and stress resistance in juvenile Jian carp (Cyprinus carpio var. Jian). Fish Shellfish Immunol. 2017, 67, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hu, Z.; Liu, S.; Sun, J.; Ji, H. Influence of dietary soybean meal replacement with yellow mealworm (Tenebrio molitor) on growth performance, antioxidant capacity, skin color, and flesh quality of mirror carp (Cyprinus carpio var. specularis). Aquaculture 2022, 561, 738686. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, S.; Dong, X.; Chi, S.; Yang, Q.; Liu, H.; Xie, S. Effects of fishmeal replacement by black soldier fly on growth performance, digestive enzyme activity, intestine morphology, intestinal flora and immune response of pearl gentian grouper (Epinephelus fuscoguttatus♀× Epinephelus lanceolatus♂). Fish Shellfish Immunol. 2022, 120, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Lin, Y.; Qian, L.; Miao, L.; Liu, B.; Ge, X.; Shen, H. Mulberry leaf meal: A potential feed supplement for juvenile Megalobrama amblycephala “Huahai No. 1”. Fish Shellfish Immunol. 2022, 128, 279–287. [Google Scholar] [CrossRef]
- Maiti, M.K.; Sahu, N.P.; Sardar, P.; Shamna, N.; Deo, A.D.; Gopan, A.; Sahoo, S. Optimum utilization of Hygrophila spinosa leaf meal in the diet of Labeo rohita (Hamilton, 1822) fingerlings. Aquac. Rep. 2019, 15, 100213. [Google Scholar] [CrossRef]
- Poolsawat, L.; Yang, H.; Sun, Y.F.; Li, X.Q.; Liang, G.Y.; Leng, X.J. Effect of replacing fish meal with enzymatic feather meal on growth and feed utilization of tilapia (Oreochromis niloticus× O. aureus). Anim. Feed Sci. Technol. 2021, 274, 114895. [Google Scholar] [CrossRef]
- Zhang, X.H.; Sun, Z.Y.; Cai, J.F.; Wang, J.H.; Wang, G.B.; Zhu, Z.L.; Cao, F.L. Effects of dietary fish meal replacement by fermented moringa (Moringa oleifera Lam.) leaves on growth performance, nonspecific immunity and disease resistance against Aeromonas hydrophila in juvenile gibel carp (Carassius auratus gibelio var. CAS III). Fish Shellfish Immunol. 2020, 102, 430–439. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Abdel-Daim, M.M.; Shukry, M.; Nowosad, J.; Kucharczyk, D. Benefits and applications of Moringa oleifera as a plant protein source in Aquafeed: A review. Aquaculture 2022, 547, 737369. [Google Scholar] [CrossRef]
- Giuberti, G.; Rocchetti, G.; Montesano, D.; Lucini, L. The potential of Moringa oleifera in food formulation: A promising source of functional compounds with health-promoting properties. Curr. Opin. Food Sci. 2021, 42, 257–269. [Google Scholar] [CrossRef]
- Singh, A.K.; Rana, H.K.; Tshabalala, T.; Kumar, R.; Gupta, A.; Ndhlala, A.R.; Pandey, A.K. Phytochemical, nutraceutical and pharmacological attributes of a functional crop Moringa oleifera Lam: An overview. S. Afr. J. Bot. 2020, 129, 209–220. [Google Scholar] [CrossRef]
- Abd El-Gawad, E.A.; El Asely, A.M.; Soror, E.I.; Abbass, A.A.; Austin, B. Effect of dietary Moringa oleifera leaf on the immune response and control of Aeromonas hydrophila infection in Nile tilapia (Oreochromis niloticus) fry. Aquac. Int. 2020, 28, 389–402. [Google Scholar] [CrossRef]
- Nepolean, P.; Anitha, J.; Emilin, R.R. Isolation, analysis and identification of phytochemicals of antimicrobial activity of Moringa oleifera Lam. Curr. Biotechnol. 2009, 3, 33–37. [Google Scholar]
- Toppo, R.; Roy, B.K.; Gora, R.H.; Baxla, S.L.; Kumar, P. Hepatoprotective activity of Moringa oleifera against cadmium toxicity in rats. Vet. World 2015, 8, 537–540. [Google Scholar] [CrossRef]
- Elabd, H.; Soror, E.; El-Asely, A.; El-Gawad, E.A.; Abbass, A. Dietary supplementation of Moringa leaf meal for Nile tilapia Oreochromis niloticus: Effect on growth and stress indices. Egypt. J. Aquat. Res. 2019, 45, 265–271. [Google Scholar] [CrossRef]
- Tageldein, M.A.; Liang, M.; Cristóbal, E.; María, G.B.J.; Ceballos, F.; Ángeles, E.M. Effects of dietary inclusion of Moringa oleifera leaves on growth and some systemic and mucosal immune parameters of seabream. Fish Physiol. Biochem. 2018, 44, 1223–1240. [Google Scholar]
- Emam, M.A.; Shourbela, R.M.; El-Hawarry, W.N.; Abo-Kora, S.Y.; Gad, F.A.M.; Abd El-latif, A.M.; Dawood, M.A. Effects of Moringa oleifera aqueous extract on the growth performance, blood characteristics, and histological features of gills and livers in Nile tilapia. Aquac. Fish. 2021, 126, 326–334. [Google Scholar] [CrossRef]
- Monir, W.; Abdel-Rahman, M.A.; El-Din, H.S.; Mansour, E.S.; Awad, S.M.M. Pomegranate peel and moringa-based diets enhanced biochemical and immune parameters of Nile tilapia against bacterial infection by Aeromonas hydrophila. Microb. Pathog. 2020, 145, 104202. [Google Scholar] [CrossRef] [PubMed]
- Kaleo, I.V.; Gao, Q.; Liu, B.; Sun, C.; Zhou, Q.; Zhang, H.; Shan, F.; Xiong, Z.; Bo, L.; Song, C. Effects of Moringa oleifera leaf extract on growth performance, physiological and immune response, and related immune gene expression of Macrobrachium rosenbergii with Vibrio anguillarum and ammonia stress. Fish Shellfish Immunol. 2019, 89, 603–613. [Google Scholar] [CrossRef]
- Stevens, C.G.; Ugese, F.D.; Otitoju, G.T.; Baiyeri, K.P. Proximate and anti-nutritional composition of leaves and seeds of Moringa oleifera in Nigeria: A comparative study. Agro-Sci. 2016, 14, 9. [Google Scholar] [CrossRef]
- Puycha, K.; Yuangsoi, B.; Charoenwattanasak, S.; Wongmaneeprateep, S.; Niamphithak, P.; Wiriyapattanasub, P. Effect of moringa (Moringa oleifera) leaf supplementation on growth performance and feed utilization of Bocourti’s catfish (Pangasius bocourti). Agric. Nat. Resour. 2017, 51, 286–291. [Google Scholar] [CrossRef]
- Hlophe, S.N.; Moyo, N. A comparative study on the use of Pennisetum clandestinum and Moringa oleifera as protein sources in the diet of the herbivorous Tilapia rendalli. Aquac. Int. 2014, 22, 1245–1262. [Google Scholar] [CrossRef]
- Richter, N.; Siddhuraju, P.; Becker, K. Evaluation of nutritional quality of moringa (Moringa oleifera Lam.) leaves as an alternative protein source for Nile tilapia (Oreochromis niloticus L.). Aquaculture 2003, 217, 599–611. [Google Scholar] [CrossRef]
- Urlings, H.A.P.; Bijker, P.G.H.; Van Logtestijn, J.G. Fermentation of raw poultry byproducts for animal nutrition. J. Anim. Sci. 1993, 71, 2420–2426. [Google Scholar] [CrossRef]
- Kari, Z.A.; Kabir, M.A.; Dawood, M.A.; Razab, M.K.A.A.; Ariff, N.S.N.A.; Sarkar, T.; Pati, S.; Edinur, H.A.; Mat, K.; Ismail, T.A.; et al. Effect of fish meal substitution with fermented soy pulp on growth performance, digestive enzyme, amino acid profile, and immune-related gene expression of African catfish (Clarias gariepinus). Aquaculture 2022, 546, 737418. [Google Scholar] [CrossRef]
- He, Y.F.; Guo, X.W.; Tan, B.P.; Dong, X.H.; Yang, Q.H.; Liu, H.Y.; Zhang, S.; Chi, S. Replacing fish meal with fermented rice protein in diets for hybrid groupers (Epinephelus fuscoguttatus♀× Epinephelus lanceolatus♂): Effects on growth, digestive and absorption capacities, inflammatory-related gene expression, and intestinal microbiota. Aquac. Rep. 2021, 19, 100603. [Google Scholar] [CrossRef]
- El-Kassas, S.; Abdo, S.E.; Abosheashaa, W.; Mohamed, R.; El-Naggar, K. Growth performance, serum lipid profile, intestinal morphometry, and growth and lipid indicator gene expression analysis of mono-sex Nile tilapia fed Moringa oleifera leaf powder. Aquac. Rep. 2020, 18, 100422. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, H.S. Screening Moringa species focused on development of locally available sustainable nutritional supplements. Nutr. Res. Pract. 2019, 13, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Saleem, H.; Al-Dujaily, A.N.G.; Al-murshidi, M.H.H. Effect of Methanolic Leaf Extract of Moringa oleifera on some Biochemical Markers in obesity induced rats. Res. J. Life Sci. Bioinform. Pharm. Chem. Sci. 2016, 7, 2222–2232. [Google Scholar]
- Amad, A.A.; Zentek, J. Moringa (M. oleifera) leaf meal in diets for broilers and laying hens: A review. J. Agric. Sci. 2022, 14, 12. [Google Scholar] [CrossRef]
- Qwele, K.; Hugo, A.; Oyedemi, S.O.; Moyo, B.; Masika, P.J.; Muchenje, V. Chemical composition, fatty acid content and antioxidant potential of meat from goats supplemented with Moringa (Moringa oleifera) leaves, sunflower cake and grass hay. Meat Sci. 2013, 93, 455–462. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemistry, 15th ed.; Association of Official Analytical Chemists Inc.: Arlington, VA, USA, 2003. [Google Scholar]
- Zeng, X.F.; Xia, W.S.; Jiang, Q.X.; Yang, F. Effect of autochthonous starter cultures on microbiological and physico-chemical characteristics of Suan yu, a traditional Chinese low salt fermented fish. Food Control 2013, 33, 344–351. [Google Scholar] [CrossRef]
- Khalil, F.; Korni, F.M.M. Evaluation of Moringa oleifera leaves and their aqueous extract in improving growth, immunity and mitigating effect of stress on common carp (Cyprinus carpio) fingerlings. Aquat. Sci. Eng. 2017, 32, 170–177. [Google Scholar] [CrossRef]
- Bisht, M.; Kumar, A.; Shah, T.K. Effect of Moringa oleifera leaf powder on skin mucosal immune responses and growth performance of guppy, Poecilia reticulata (Peter, 1860). Aquac. Res. 2020, 51, 4984–4990. [Google Scholar] [CrossRef]
- Bbole, I.; Mumba, C.; Mupenda, N.; Kefi, A.S. Analysis of growth performance and haematological parameters of Oreochromis niloticus fed on a varying diet of Moringa oleifera Lam. leaf meal as an additive protein source. Int. J. Aquac. Fish Sci. 2016, 8, 105–111. [Google Scholar]
- He, M.; Yu, Y.; Li, X.; Poolsawat, L.; Yang, P.; Bian, Y.; Leng, X. An evaluation of replacing fish meal with fermented soybean meal in the diets of largemouth bass (Micropterus salmoides): Growth, nutrition utilization and intestinal histology. Aquac. Res. 2020, 51, 4302–4314. [Google Scholar] [CrossRef]
- Arsalan, M.Z.H.; Hussain, S.M.; Asrar, M.; Anwar, H.; Rehan, M.M.H.; Shahzad, M.M.; Riaz, D.; Ahmad, N.; Wahab, N. Effects of Moringa oleifera leaf meal (MOLM) based diets on carcass composition and hematology of Labeo rohita fingerlings. J. Biodivers. Environ. Sci. 2016, 9, 214–223. [Google Scholar]
- Karina, S.; Akbar, M.; Supriatna, A.; Muchlisin, Z.A. Replacement of soybean meal with moringa oleifera leaf meal in the formulated diets of tilapia (Oreochromis niloticus) fingerlings. Aquac. Aquar. Conserv. Legis. 2015, 8, 790–805. [Google Scholar]
- Shahzad, M.M.; Hussain, S.M.; Jabeen, F.; Hussain, A.I.; Arsalan, M.Z.H.; Ahmad, N. Carcass composition and hematological study of Catla catla fingerlings fed on phytase supplemented Moringa oleifera leaf meal (MOLM) based diet. J. Biodivers. Environ. Sci. 2016, 9, 57–68. [Google Scholar]
- Francesco, M.; Parisi, G.; Médale, F.; Lupi, P.; Kaushik, S.J.; Poli, B.M. Effect of long-term feeding with a plant protein mixture-based diet on growth and body/fillet quality traits of large rainbow trout (Oncorhynchus mykiss). Aquaculture 2004, 236, 413–429. [Google Scholar] [CrossRef]
- Kato, H.; Rhue, M.R.; Nishimura, T. Role of Free Amino Acids and Peptides in Food Taste; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1989. [Google Scholar]
- Liu, X.W.; Lin, F.; Jiang, W.D.; Wu, P.; Jiang, J.; Yang, D.M.; Tang, L.; Kuang, S.Y.; Shi, H.Q.; Zhou, X.Q.; et al. (2-Carboxyethyl) dimethylsulfonium Bromide (Br-DMPT) improves muscle flesh quality and antioxidant status of on-growing grass carp (Ctenopharyngodon idella) fed non-fish meal diets. Aquaculture 2020, 521, 735065. [Google Scholar] [CrossRef]
- Yang, L.L.; Lin, F.; Wu, P.; Jiang, W.D.; Liu, Y.; Kuang, S.Y.; Tang, L.; Zhou, X.Q. Guanidinoacetic acid supplementation totally based on vegetable meal diet improved the growth performance, muscle flavor components and sensory characteristics of on-growing grass carp (Ctenopharygodon idella). Aquaculture 2021, 531, 735841. [Google Scholar] [CrossRef]
- Odedeyi, D.O. Nutritive values of the carcass of African catfish (Clarias gariepinus) fingerlings fed raw and steam-heated Moringa oleifera diets. In Proceedings of the International Conference on Advances in Environment, Agriculture & Medical Sciences (ICAEAM’14), Kuala Lumpur, Malaysia, 16–17 November 2014; pp. 16–27. [Google Scholar]
- Cai, Z.; Mai, K.; Ai, Q. Regulation of hepatic lipid deposition by phospholipid in large yellow croaker. Br. J. Nutr. 2017, 118, 999–1009. [Google Scholar] [CrossRef]
- Lara-Castro, C.; Garvey, W.T. Intracellular lipid accumulation in liver and muscle and the insulin resistance syndrome. Endocrinol. Metab. Clin. N. Am. 2008, 37, 841–856. [Google Scholar] [CrossRef]
- Song, F.; Qin, Y.; He, C.; Geng, H.; Yang, P.; Wang, W.; Chen, Y. Transcriptome analysis reveals the effects of dietary lipid level on growth performance and immune response in golden pompano (Trachinotus ovatus). Aquaculture 2023, 563, 738959. [Google Scholar] [CrossRef]
- Guo, J.; Fu, Y.; Wu, Z.; Yu, X.; Guo, Y.; Liu, J.; Zhang, W.; Mai, K. Effects of dietary carbohydrate levels on growth performance, body composition, glucose/lipid metabolism and insulin signaling pathway in abalone Haliotis discus hannai. Aquaculture 2022, 557, 738284. [Google Scholar] [CrossRef]
- Ncha, O.S.; Michael, P.B.; Nnabuchi, U.O.; Alex, E. Effect of diets with moringa leaf meal on growth, carcass composition and haematology of Clarias gariepinus. Fish Aquat. Sci. 2015, 3, 397–401. [Google Scholar]
- Sugita, H.; Miyajima, C.; Deguchi, Y. The vitamin B12-producing ability of the intestinal microflora of freshwater fish. Aquaculture 1991, 92, 267–276. [Google Scholar] [CrossRef]
- Vine, N.G.; Leukes, W.D.; Kaiser, H. Probiotics in marine larviculture. FEMS Microbiol. Rev. 2006, 30, 404–427. [Google Scholar] [CrossRef]
- Dyck, J.; Berthiaume, L.G.; Thomas, P.D.; Kantor, P.F.; Barr, A.J.; Barr, R.; Singh, D.; Hopkins, T.A.; Voilley, N.; Prentki, M.; et al. Characterization of rat liver malonyl-CoA decarboxylase and the study of its role in regulating fatty acid metabolism. Biochem. J. 2000, 350, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liang, H.; Maulu, S.; Ge, X.; Xi, B. Dietary phosphorus affects growth, glucolipid metabolism, antioxidant activity and immune status of juvenile blunt snout bream (Megalobrama amblycephala). Anim. Feed Sci. Technol. 2021, 274, 114896. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, W.B.; Brown, P.B.; Xu, C.; Li, X.F. Utilization of raw and gelatinized starch by blunt snout bream Megalobrama amblycephala as evidenced by the glycolipid metabolism, glucose tolerance and mitochondrial function. Aquaculture 2020, 529, 735603. [Google Scholar] [CrossRef]
- Zhao, T.; Yang, S.; Chen, G.; Xu, Y.; Luo, Z. Dietary glucose increases glucose absorption and lipid deposition via SGLT1/2 signaling and acetylated ChREBP in the intestine and isolated intestinal epithelial cells of yellow catfish. J. Nutr. 2020, 150, 1790–1798. [Google Scholar] [CrossRef]
- Zhao, T.; Wu, K.; Hogstrand, C.; Xu, Y.; Chen, G.; Wei, C.; Luo, Z. Lipophagy mediated carbohydrate-induced changes of lipid metabolism via oxidative stress, endoplasmic reticulum (ER) stress and ChREBP/PPARγ pathways. Cell. Mol. Life Sci. 2019, 77, 1987–2003. [Google Scholar] [CrossRef]
- Iizuka, K.; Bruick, R.K.; Liang, G.; Horton, J.D.; Uyeda, K. Deficiency of carbohydrate response element-binding protein (ChREBP) reduces lipogenesis as well as glycolysis. Proc. Natl. Acad. Sci. USA 2004, 101, 7281–7296. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.T.; Lerin, C.; Haas, W.; Gygi, S.P.; Spiegelman, B.M.; Puigserver, P. Nutrient control of glucose homeostasis through a complex of PGC-1α and SIRT1. Nature 2005, 434, 113–128. [Google Scholar] [CrossRef]
- Michael, M.D.; Kulkarni, R.N.; Postic, C. Loss of Insulin Signaling in Hepatocytes Leads to Severe Insulin Resistance and Progressive Hepatic Dysfunction. Mol. Cell 2000, 6, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Massa, M.L.; Gagliardino, J.J.; Francini, F. Liver glucokinase: An overview on the regulatorymechanisms of its activity. IUBMB Life 2011, 63, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Liang, H.; Ren, M.; Ge, X.; Ji, K.; Yu, H.; Maulu, S. Effects of dietary lysine levels on growth performance, whole body composition and gene expression related to glycometabolism and lipid metabolism in grass carp, Ctenopharyngodon idellus fry. Aquaculture 2021, 530, 735806. [Google Scholar] [CrossRef]
- Abou-Zeid, S.M.; Ahmed, A.I.; Awad, A.; Mohammed, W.A.; Metwally, M.M.M.; Almeer, R.; Abdel-Daim, M.M.; Khalil, S.R. Moringa oleifera ethanolic extract attenuates tilmicosin-induced renal damage in male rats via suppression of oxidative stress, inflammatory injury, and intermediate filament proteins mRNA expression. Biomed. Pharmacother. 2021, 133, 110997. [Google Scholar] [CrossRef] [PubMed]
- Mabe, L.T.; Su, S.; Tang, D.; Zhu, W.; Wang, S.; Dong, Z. The effect of dietary bamboo charcoal supplementation on growth and serum biochemical parameters of juvenile common carp (Cyprinus carpio L.). Aquac. Res. 2018, 49, 1142–1152. [Google Scholar] [CrossRef]
- Kongchum, P.; Palti, Y.; Hallerman, E.M.; Hulata, G.; David, L. SNP discovery and development of genetic markers for mapping innate immune response genes in common carp (Cyprinus carpio). Fish Shellfish Immunol. 2010, 29, 356–361. [Google Scholar] [CrossRef]
- Swanson, L.; Katkar, G.D.; Tam, J.; Pranadinata, R.F.; Chareddy, Y.; Coates, J. TLR4 signaling and macrophage inflammatory responses are dampened by GIV/Girdin. Proc. Natl. Acad. Sci. USA 2020, 117, 26895–26906. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, S.; Voulgarelis, M. Toll-Like Receptors, Tissue Injury, and Tumourigenesis. Mediat. Inflamm. 2010, 2010, 581837. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, G.; Karimi, A.A.; Hafezieh, M.; Dawood, M.A.O.; Abo-Al-Ela, H.G. Pistachio hulls polysaccharide protects Nile tilapia against LPS-induced excessive inflammatory responses and oxidative stress possibly via TLR2 and Nrf2 signaling pathways. Fish Shellfish Immunol. 2021, 27, 254–268. [Google Scholar] [CrossRef]
- Zhe, Y.; Zhao, L.; Zhao, J.L.; Xu, W.X.; Guo, Z.H.; Zhang, A.Z. Dietary Taraxacum mongolicum polysaccharide ameliorates the growth, immune response, and antioxidant status in association with NF-κB, Nrf2 and TOR in Jian carp (Cyprinus carpio var. Jian). Aquaculture 2022, 547, 737522. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Control | MO2 | MO4 | FMO2 | FMO4 |
|---|---|---|---|---|---|
| MO 1 | 2.2 | 4.4 | |||
| FMO 2 | 2.2 | 4.4 | |||
| Fish meal 3 | 4.3 | 4.3 | 4.3 | 4.3 | 4.3 |
| Soybean meal 3 | 21.6 | 21.6 | 21.6 | 21.6 | 21.6 |
| Rapeseed meal 3 | 23.8 | 23.8 | 23.8 | 23.8 | 23.8 |
| Cottonseed meal 3 | 16.4 | 15.3 | 14.2 | 15.3 | 14.2 |
| Wheat meal 3 | 12.2 | 12.2 | 12.2 | 12.2 | 12.2 |
| Rice bran 3 | 10.9 | 12.0 | 13.1 | 12.0 | 13.1 |
| Wheat bran 3 | 4.4 | 2.2 | 0.3 | 2.2 | 0.3 |
| Soybean oil | 2.5 | 2.5 | 2.2 | 2.5 | 2.2 |
| Calcium dihydrogen phosphate 4 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Vitamin mix and Mineral mix 5 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Vitamin C 4 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Choline chloride 4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| Microcrystalline cellulose 4 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Bentonite 4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| Nutrient composition (determination of dry basis) | |||||
| Crude protein % | 35.71 | 35.18 | 35.73 | 35.84 | 35.91 |
| Crude lipid % | 10.42 | 10.43 | 10.23 | 10.75 | 10.79 |
| Gross energy MJ·kg−1 | 19.87 | 19.96 | 19.63 | 19.38 | 19.66 |
| Genes | Primer Sequence (5′–3′) | Accession No. | Product Length (bps) | |
|---|---|---|---|---|
| tlr4 | Forward | TAATGGGCAGCCGTAAAGTC | XM_048204247.1 | 114 |
| Reverse | TGGCATTGCGTTCCATAATA | |||
| nf-κb | Forward | AGTCCGATCCATCCGCACTA | XM_048176853.1 | 85 |
| Reverse | ACTGGAGCCGGTCATTTCAG | |||
| il-8 | Forward | CAGAGAGTCGACGCATTGGT | XM_048197357.1 | 184 |
| Reverse | ATTCACGGTGCTTTGTTGGC | |||
| p53 | Forward | CCATCCTCACAATCATCAC | XM_048187452.1 | 114 |
| Reverse | TGCTCTCCTCAGTTTTCCT | |||
| g6pase | Forward | TTCAGTGTCACGCTGTTCCT | XM_048171060.1 | 119 |
| Reverse | TCTGGACTGACGCACCATTT | |||
| pk | Forward | GCCGAGAAAGTCTTCATCGCACAG | XM_048152870.1 | 157 |
| Reverse | CGTCCAGAACCGCATTAGCCAC | |||
| ppar-β | Forward | CATCCTCACGGGCAAGAC | XM_048209548.1 | 153 |
| Reverse | CACTGGCAGCGGTAGAAG | |||
| acc-α | Forward | TCTGCCCTCTATCTGTCT | XM_048189972.1 | 162 |
| Reverse | ATGCCAATCTCATTTCCT | |||
| lpl | Forward | GCCACGAGTGTTGGTGTGAA | XM_048164066.1 | 91 |
| Reverse | TGGCCCTAGCTTTGAGTACG | |||
| β-actin | Forward | TCGTCCACCGCAAATGCTTCTA | AY170122.2 | 190 |
| Reverse | CCGTCACCTTCACCGTTCCAGT |
| Parameters | IBW (g) | FBW (g) | WGR (%) | SGR (% day−1) | FCR | PER (%) | CF (g/cm3) |
|---|---|---|---|---|---|---|---|
| Control | 20.37 ± 0.07 | 48.60 ± 0.69 | 138.56 ± 3.08 | 1.64 ± 0.03 | 1.51 ± 0.03 | 1.32 ± 0.04 | 2.27 ± 0.06 b |
| MO2 | 20.15 ± 0.11 | 43.26 ± 1.70 | 114.74 ± 9.19 | 1.44 ± 0.08 | 1.79 ± 0.08 | 1.40 ± 0.01 | 2.14 ± 0.05 ab |
| FMO2 | 20.25 ± 0.07 | 45.06 ± 2.33 | 126.46 ± 14.11 | 1.53 ± 0.12 | 1.59 ± 0.18 | 1.60 ± 0.10 | 2.10 ± 0.05 ab |
| MO4 | 20.20 ± 0.15 | 46.12 ± 2.20 | 128.33 ± 10.51 | 1.55 ± 0.09 | 1.58 ± 0.12 | 1.66 ± 0.13 | 2.12 ± 0.06 ab |
| FMO4 | 20.36 ± 0.16 | 47.10 ± 0.53 | 131.42 ± 3.76 | 1.58 ± 0.03 | 1.54 ± 0.09 | 1.63 ± 0.26 | 2.05 ± 0.02 a |
| p value | 0.267 | 0.310 | 0.498 | 0.499 | 0.479 | 0.369 | 0.042 |
| Muscle Composition | Moisture % | Crude Protein % | Crude Lipid % | Ash % |
|---|---|---|---|---|
| Control | 77.89 ± 0.10 ab | 19.52 ± 0.12 bc | 2.18 ± 0.14 ab | 1.27 ± 0.02 |
| MO2 | 77.62 ± 0.21 a | 19.10 ± 0.11 ab | 1.99 ± 0.07 a | 1.29 ± 0.03 |
| FMO2 | 78.65 ± 0.10 b | 18.89 ± 0.15 a | 2.16 ± 0.14 ab | 1.28 ± 0.01 |
| MO4 | 78.46 ± 0.21 b | 19.77 ± 0.11 c | 2.15 ± 0.11 ab | 1.31 ± 0.05 |
| FMO4 | 77.34 ± 0.27 a | 19.71 ± 0.09 c | 2.60 ± 0.07 b | 1.25 ± 0.03 |
| p value | <0.001 | <0.001 | 0.008 | 0.812 |
| Content mg/100 g | Control | MO4 | FMO4 | p Value |
|---|---|---|---|---|
| Aspartic acid | 3.31 ± 0.05 b | 3.07 ± 0.04 a | 3.08 ± 0.05 a | 0.018 |
| Glutamic acid | 7.48 ± 0.42 | 7.54 ± 0.39 | 6.75 ± 0.04 | 0.166 |
| Total umami amino acids (TUAA) | 10.79 ± 0.15 | 10.61 ± 0.43 | 9.83 ± 0.15 | 0.094 |
| Threonine | 31.28 ± 0.32 b | 28.78 ± 0.53 a | 27.65 ± 0.14 a | 0.048 |
| Glycine | 105.30 ± 1.32 a | 122.77 ± 0.79 b | 105.51 ± 0.21 a | 0.006 |
| Alanine | 27.56 ± 0.23 b | 22.97 ± 0.68 a | 23.43 ± 0.02 a | 0.004 |
| Proline | 14.65 ± 0.34 ab | 9.55 ± 1.13 a | 17.57 ± 2.65 b | 0.038 |
| Serine | 1.88 ± 0.72 | 3.06 ± 0.18 | 2.41 ± 0.21 | 0.124 |
| Total sweetish amino acids (TSAA) | 180.67 ± 0.85 ab | 187.13 ± 1.75 b | 176.57 ± 2.83 a | 0.018 |
| Valine | 4.06 ± 0.06 b | 3.60 ± 0.09 a | 3.91 ± 0.13 ab | 0.037 |
| Methionine | 2.06 ± 0.04 a | 2.64 ± 0.17 b | 2.80 ± 0.03 b | 0.005 |
| Isoleucine | 2.40 ± 0.08 | 2.09 ± 0.13 | 2.48 ± 0.03 | 0.057 |
| Leucine | 4.90 ± 0.05 b | 3.83 ± 0.22 a | 4.33 ± 0.08 ab | 0.005 |
| Phenylalanine | 2.09 ± 0.36 a | 3.44 ± 0.78 ab | 5.05 ± 0.07 b | 0.007 |
| Histidine | 555.23 ± 6.91 | 569.57 ± 13.25 | 584.75 ± 7.99 | 0.184 |
| Lysine | 20.64 ± 0.24 b | 11.82 ± 1.18 a | 17.59 ± 0.22 b | <0.001 |
| Argnine | 7.76 ± 0.07 c | 4.78 ± 0.39 a | 6.67 ± 0.02 b | <0.001 |
| Tyrosine | 4.50 ± 0.10 a | 5.74 ± 0.43 b | 4.88 ± 0.21 ab | 0.025 |
| Cysteine | 11.04 ± 0.36 b | 8.68 ± 0.66 b | 5.32 ± 1.03 a | 0.004 |
| Total bitter amino acids (TBAA) | 614.68 ± 6.48 | 616.20 ± 14.60 | 637.78 ± 7.89 | 0.142 |
| Total free amino acids (TFAA) | 807.06 ± 7.69 a | 813.94 ± 15.53 a | 824.18 ± 5.12 b | 0.029 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, W.; Qian, L.; Zhao, Y.; Lin, Y.; Yang, Y.; Shen, H.; Ge, X.; Miao, L. The Application of Moringa oleifera Leaf Meal and Its Fermentation Products in the Diet of Megalobrama amblycephala Juveniles. Fermentation 2023, 9, 577. https://doi.org/10.3390/fermentation9060577
Jiang W, Qian L, Zhao Y, Lin Y, Yang Y, Shen H, Ge X, Miao L. The Application of Moringa oleifera Leaf Meal and Its Fermentation Products in the Diet of Megalobrama amblycephala Juveniles. Fermentation. 2023; 9(6):577. https://doi.org/10.3390/fermentation9060577
Chicago/Turabian StyleJiang, Wenqiang, Linjie Qian, Yongfeng Zhao, Yan Lin, Yang Yang, Huaishun Shen, Xianping Ge, and Linghong Miao. 2023. "The Application of Moringa oleifera Leaf Meal and Its Fermentation Products in the Diet of Megalobrama amblycephala Juveniles" Fermentation 9, no. 6: 577. https://doi.org/10.3390/fermentation9060577
APA StyleJiang, W., Qian, L., Zhao, Y., Lin, Y., Yang, Y., Shen, H., Ge, X., & Miao, L. (2023). The Application of Moringa oleifera Leaf Meal and Its Fermentation Products in the Diet of Megalobrama amblycephala Juveniles. Fermentation, 9(6), 577. https://doi.org/10.3390/fermentation9060577