


Enhancing the Activity of a Self-Inducible Promoter in Escherichia coli through Saturation Mutation and High-Throughput Screening


 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, Competent Cells, and Reagents
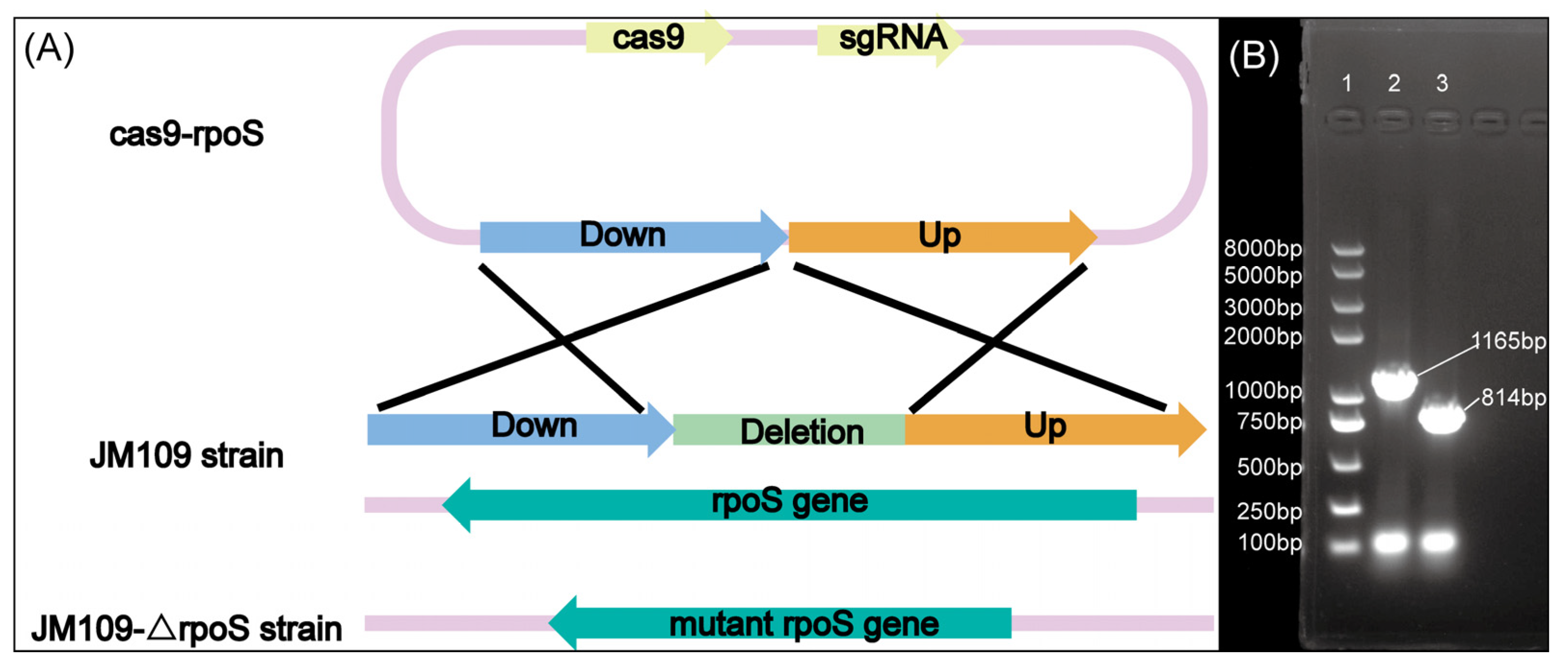
2.2. Construction of the JM109-ΔrpoS Strain
2.3. Construction of the Expression Vector
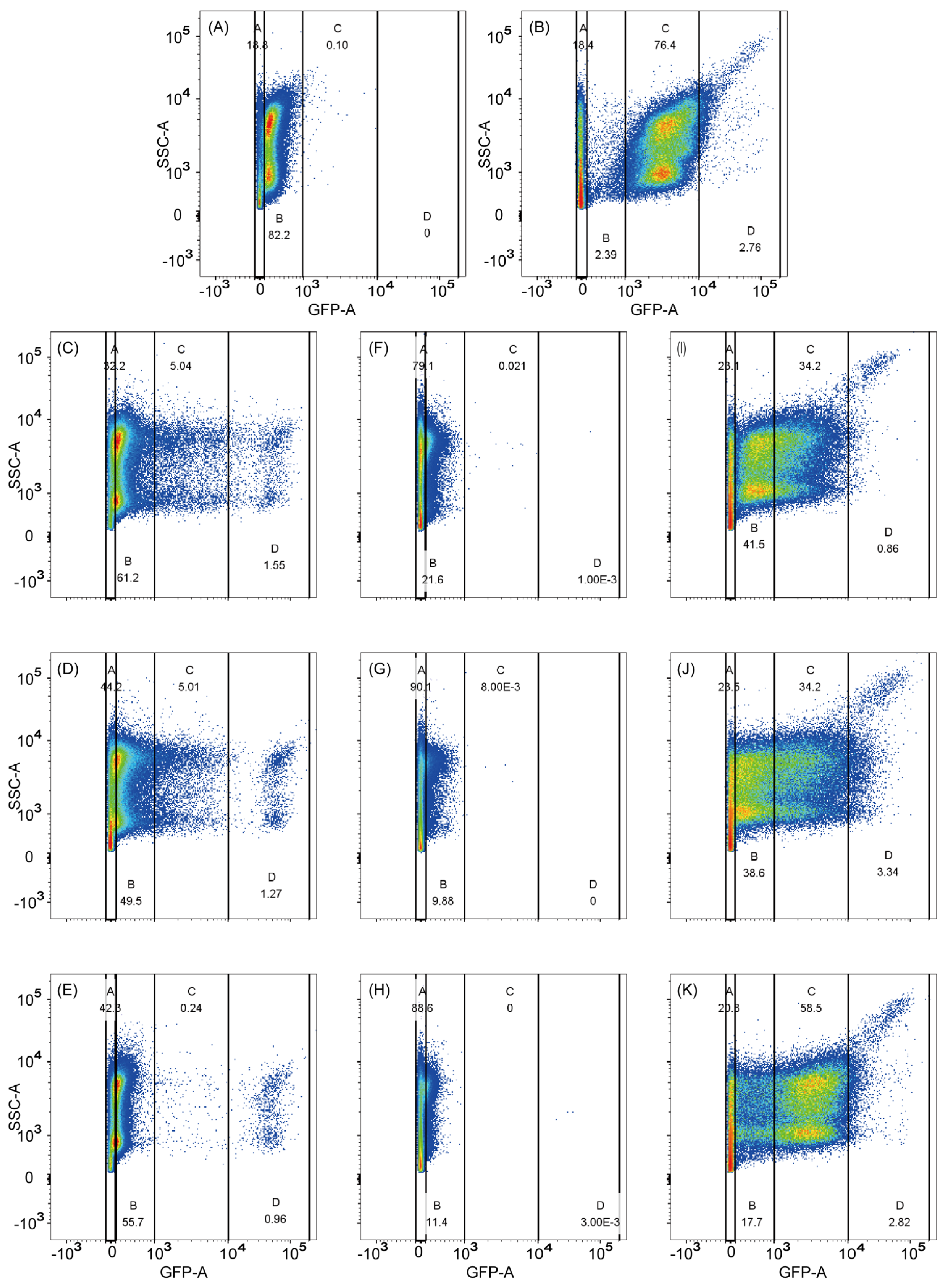
2.4. Cultivation and Screening
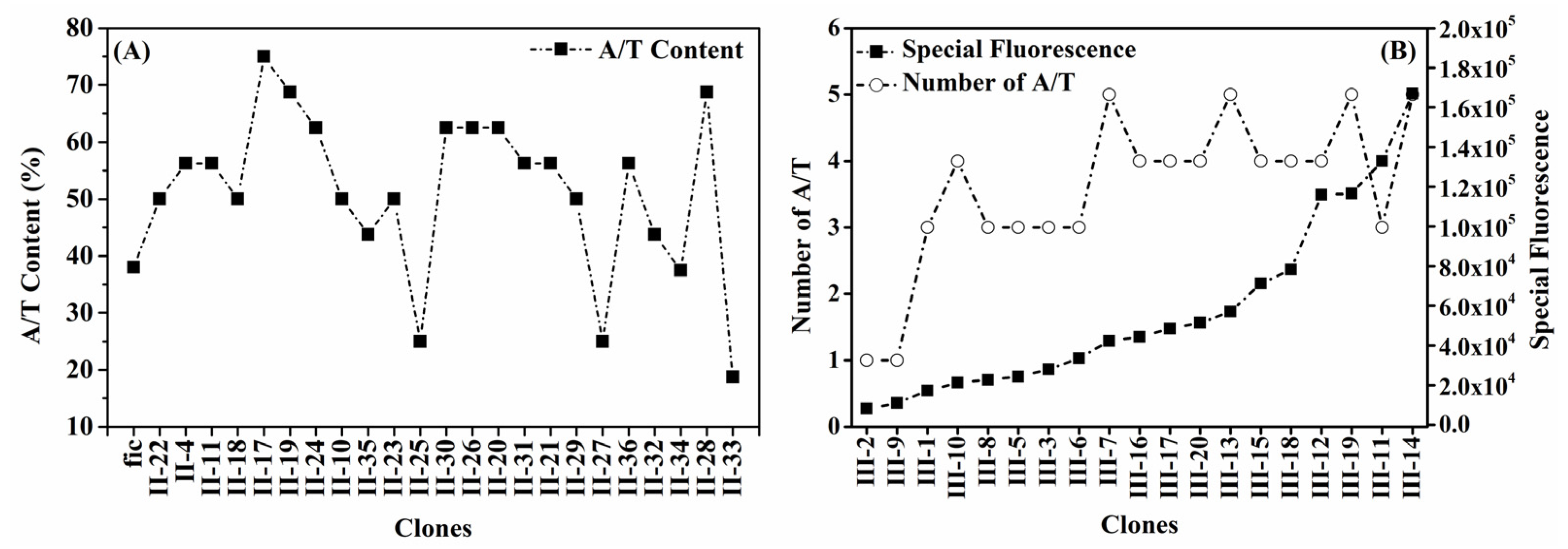
2.5. Sequence and Sequence Logo
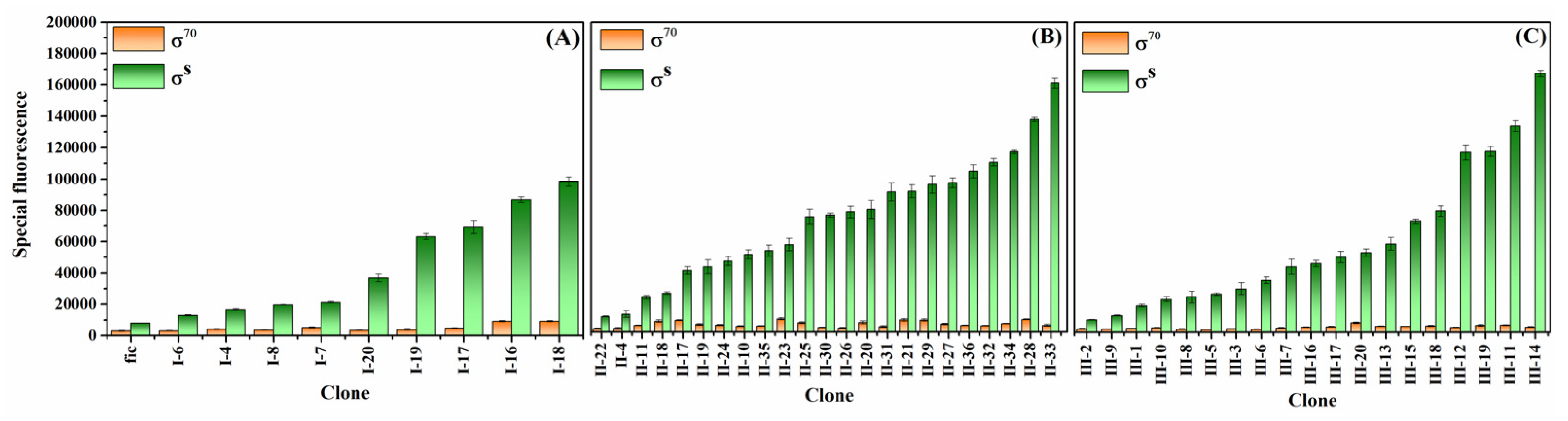
2.6. Fluorescence Assay
2.7. Dynamics under Different Fermentation Patterns
2.8. Validation of the Promoters
3. Results and Discussion
3.1. Construction of the JM109-ΔrpoS Strain
3.2. Screening of Mutant Promoters
3.3. Generation of Enhanced Self-Inducible Promoter Libraries
3.4. Features of Enhanced Self-Inducible Promoter Sequences
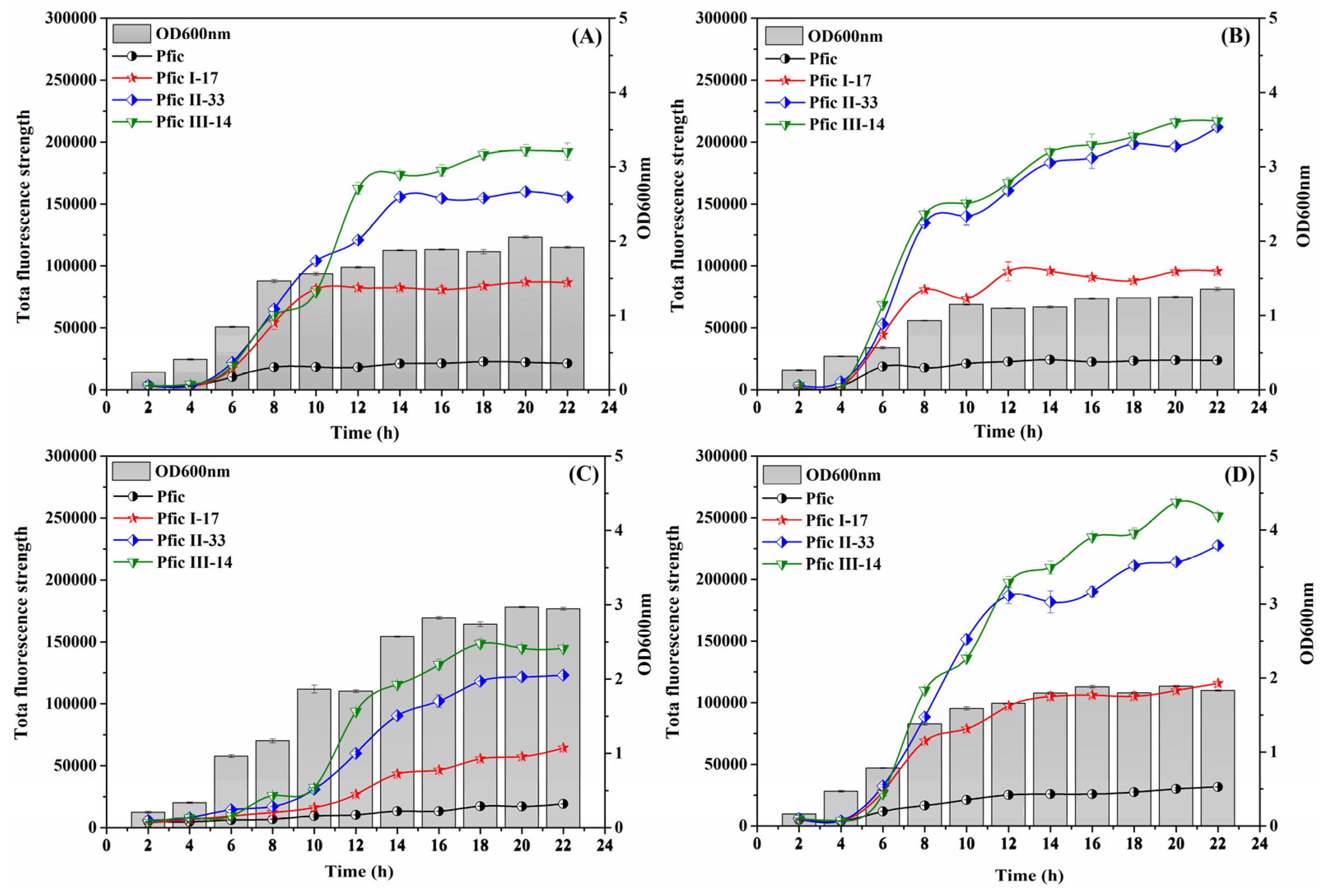
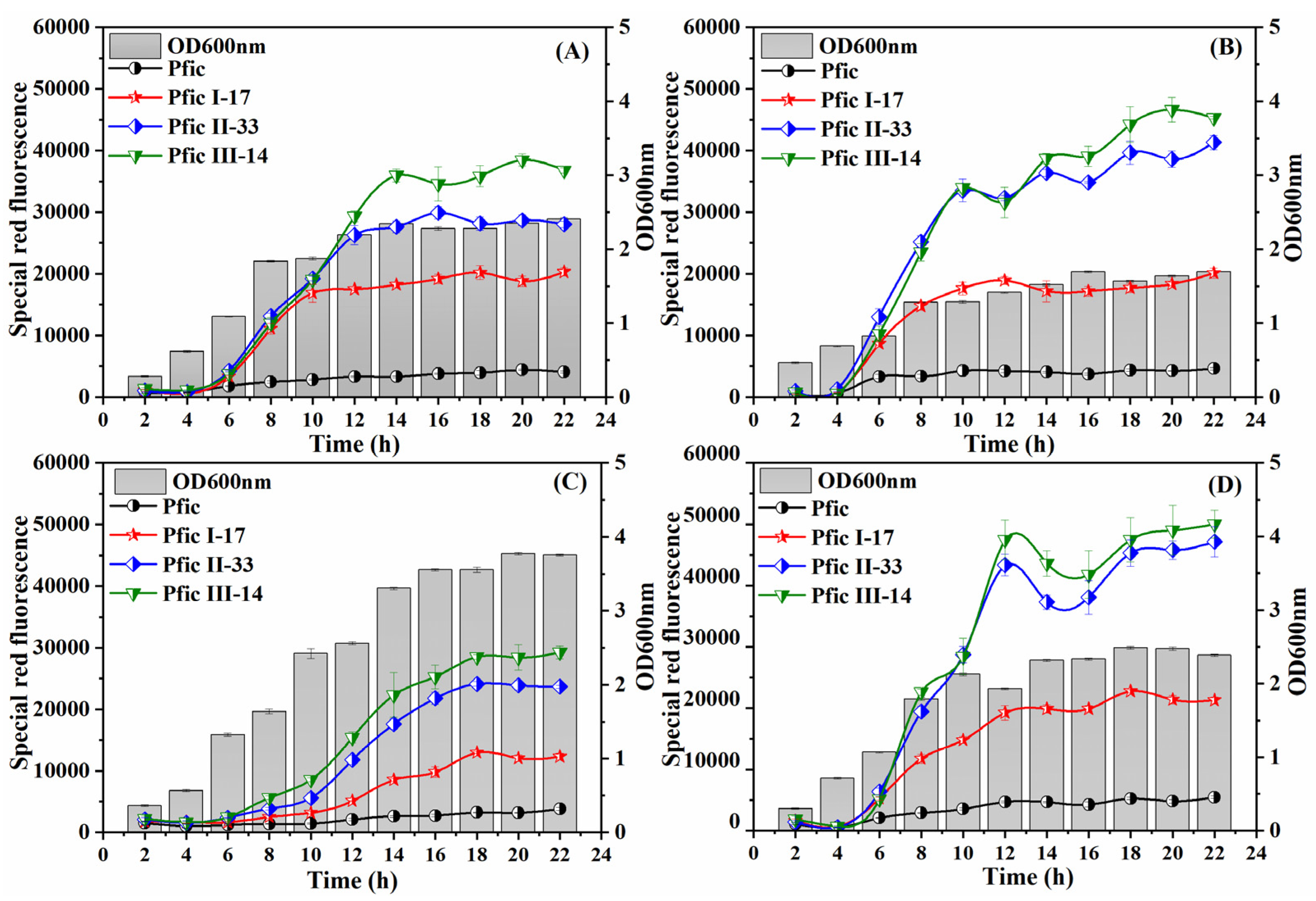
3.5. Dynamics under Different Fermentation Patterns
3.6. Validation of Promoters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Connor, A.; Wigham, C.; Bai, Y.; Rai, M.; Nassifa, S.; Koffas, M.; Zha, R.H. Novel insights into construct toxicity, strain optimization, and primary sequence design for producing recombinant silk fibroin and elastin-like peptide in E. coli. Metab. Eng. Commun. 2023, 16, e00219. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wei, W.; Zong, Y.; Bai, C.; Guo, X.; Zhu, H.; Lou, C. Novel switchable ECF sigma factor transcription system for improving thaxtomin A production in Streptomyces. Synth. Syst. Biotechnol. 2022, 7, 972–981. [Google Scholar] [CrossRef]
- Kaur, J.; Kumar, A.; Kaur, J. Strategies for optimization of heterologous protein expression in E. coli: Roadblocks and reinforcements. Int. J. Biol. Macromol. 2018, 106, 803–822. [Google Scholar] [CrossRef]
- Pasini, M.; Fernandez-Castane, A.; Jaramillo, A.; Mas, C.d.; Caminal, G.; Ferrer, P. Using promoter libraries to reduce metabolic burden due to plasmid-encoded proteins in recombinant Escherichia coli. New Biotechnol. 2016, 33, 78–90. [Google Scholar] [CrossRef]
- Averina, O.; Alekseeva, M.; Shkoporov, A.; Danilenko, V. Functional analysis of the type II toxin-antitoxin systems of the MazEF and RelBE families in Bifidobacterium longum subsp. infantis ATCC 15697. Anaerobe 2015, 35, 59–67. [Google Scholar] [CrossRef]
- Ahmad, I.; Nawaz, N.; Darwesh, N.M.; Rahman, S.u.; Mustafa, M.Z.; Khan, S.B.; Patching, S.G. Overcoming challenges for amplified expression of recombinant proteins using Escherichia coli. Protein Expr. Purif. 2018, 144, 12–18. [Google Scholar] [CrossRef]
- Kwon, S.-K.; Kim, S.K.; Lee, D.-H.; Kim, J.F. Comparative genomics and experimental evolution of Escherichia coli BL21(DE3) strains reveal the landscape of toxicity escape from membrane protein overproduction. Sci. Rep. 2015, 5, 16076. [Google Scholar] [CrossRef]
- Erbakan, M.; Curtis, B.S.; Nixon, B.T.; Kumar, M.; Curtis, W.R. Advancing Rhodobacter sphaeroides as a platform for expression of functional membrane proteins. Protein Expr. Purif. 2015, 115, 109–117. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, Y.; Liu, Y.; Li, J.; Du, G.; Chen, J.; Lv, X.; Liu, L. Sustainable bioproduction of natural sugar substitutes: Strategies and challenges. Trends Food Sci. Technol. 2022, 129, 512–527. [Google Scholar] [CrossRef]
- Cardoso, V.; Brás, J.L.A.; Costa, I.F.; Ferreira, L.M.A.; Gama, L.T.; Vincentelli, R.; Henrissat, B.; Fontes, C.M.G.A. Generation of a Library of Carbohydrate-Active Enzymes for Plant Biomass Deconstruction. Int. J. Mol. Sci. 2022, 23, 4024. [Google Scholar] [CrossRef]
- Zhu, L.; Huang, J.; Wang, Y.; Yang, Z.; Chen, X. Chemerin causes lipid metabolic imbalance and induces passive lipid accumulation in human hepatoma cell line via the receptor GPR1. Life Sci. 2021, 278, 119530. [Google Scholar] [CrossRef] [PubMed]
- Korpys-Wozniak, P.; Celinska, E. Global transcriptome profiling reveals genes responding to overproduction of a small secretory, a high cysteine- and a high glycosylation-bearing protein in Yarrowia lipolytica. Biotechnol. Rep. 2021, 31, e00646. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Kojima, K.; Xu, N.; Mobley, J.; Zhou, L.; Yang, S.-T.; Liu, X.M. Comparative proteomics analysis of high n-butanol producing metabolically engineered Clostridium tyrobutyricum. J. Biotechnol. 2015, 193, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.M.; Garcia, D.E.; Redding-Johanson, A.M.; Friedland, G.D.; Chan, R.; Batth, T.S.; Haliburton, J.R.; Chivian, D.; Keasling, J.D.; Petzold, C.J.; et al. Optimization of a heterologous mevalonate pathway through the use of variant HMG-CoA reductases. Metab. Eng. 2011, 13, 588–597. [Google Scholar] [CrossRef] [PubMed]
- D’Huys, P.-J.; Lule, I.; Hove, S.V.; Vercammen, D.; Wouters, C.; Bernaerts, K.; Anné, J.; Impe, J.F.M.V. Amino acid uptake profiling of wild type and recombinant Streptomyces lividans TK24 batch fermentations. J. Biotechnol. 2011, 152, 132–143. [Google Scholar] [CrossRef]
- Zhao, M.; Huang, D.; Zhang, X.; Koffas, M.A.G.; Zhou, J.; Deng, Y. Metabolic engineering of Escherichia coli for producing adipic acid through the reverse adipate-degradation pathway. Metab. Eng. 2018, 47, 254–262. [Google Scholar] [CrossRef]
- Marbach, A.; Bettenbrock, K. lac operon induction in Escherichia coli: Systematic comparison of IPTG and TMG induction and influence of the transacetylase LacA. J. Biotechnol. 2012, 157, 82–88. [Google Scholar] [CrossRef]
- Baptista, I.S.C.; Kandavalli, V.; Chauhan, V.; Bahrudeen, M.N.M.; Almeida, B.L.B.; Palma, C.S.D.; Dash, S.; Ribeiro, A.S. Sequence-dependent model of genes with dual σ factor preference. Biochim. Biophys. Acta—Gene Regul. Mech. 2022, 1865, 194812. [Google Scholar] [CrossRef]
- Morichaud, Z.; Chaloin, L.; Brodolin, K. Regions 1.2 and 3.2 of the RNA Polymerase σ Subunit Promote DNA Melting and Attenuate Action of the Antibiotic Lipiarmycin. J. Mol. Biol. 2016, 428, 463–476. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, S.; Li, Y.; Zhang, K.; Mo, F.; Zhang, J.; Hou, Y.; He, L.; Liu, Z.; Wang, Y.; et al. RssB-mediated σS Activation is Regulated by a Two-Tier Mechanism via Phosphorylation and Adaptor Protein—IraD. J. Mol. Biol. 2021, 433, 166757. [Google Scholar] [CrossRef]
- Micevski, D.; Zeth, K.; Mulhern, T.D.; Schuenemann, V.J.; Zammit, J.E.; Truscott, K.N.; Dougan, D.A. Insight into the RssB-Mediated Recognition and Delivery of σS to the AAA+ Protease, ClpXP. Biomolecules 2020, 10, 615. [Google Scholar] [CrossRef]
- Dorich, V.; Brugger, C.; Tripathi, A.; Hoskins, J.R.; Tong, S.; Suhanovsky, M.M.; Sastry, A.; Wickner, S.; Gottesman, S.; Deaconescu, A.M. Structural basis for inhibition of a response regulator of σS stability by a ClpXP antiadaptor. Genes Dev. 2019, 33, 718–732. [Google Scholar] [CrossRef] [PubMed]
- Cartagena, A.J.; Banta, A.B.; Sathyan, N.; Ross, W.; Gourse, R.L.; Campbell, E.A.; Darst, S.A. Structural basis for transcription activation by Crl through tethering of σS and RNA polymerase. Proc. Natl. Acad. Sci. USA 2019, 116, 18923–18927. [Google Scholar] [CrossRef] [PubMed]
- Lago, M.; Monteil, V.; Douche, T.; Guglielmini, J.; Criscuolo, A.; Maufrais, C.; Matondo, M.; Norel, F. Proteome remodelling by the stress sigma factor RpoS/σS in Salmonella: Identification of small proteins and evidence for post-transcriptional regulation. Sci. Rep. 2017, 7, 2127. [Google Scholar] [CrossRef] [PubMed]
- Levi-Meyrueis, C.; Monteil, V.e.; Sismeiro, O.; Dillies, M.-A.; Kolb, A.; Monot, M.; Dupuy, B.; Duarte, S.S.; Jagla, B.; Coppee, J.-Y.; et al. Repressor activity of the RpoS/σS-dependent RNA polymerase requires DNA binding. Nucleic Acids Res. 2015, 43, 1456–1468. [Google Scholar] [CrossRef]
- Burda, W.N.; Miller, H.K.; Krute, C.N.; Leighton, S.L.; Shaw, R.K.C.a.L.N. Investigating the genetic regulation of the ECF sigma factor σS in Staphylococcus aureus. BMC Microbiol. 2014, 14, 1471–2180. [Google Scholar] [CrossRef]
- Gottesman, S. Trouble is coming: Signaling pathways that regulate general stress responses in bacteria. J. Biol. Chem. 2019, 294, 11685–11700. [Google Scholar] [CrossRef]
- Matin, A. Stress, Bacterial: General and Specific. In Encyclopedia of Microbiology; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Adams, J.; Hoang, J.; Petroni, E.; Ashby, E.; Hardin, J.; Stoebel, D.M. The timing of transcription of RpoS-dependent genes varies across multiple stresses. bioRxiv 2023. [Google Scholar] [CrossRef]
- Wong, G.T.; Bonocora, R.P.; Schep, A.N.; Beeler, S.M.; Fong, A.J.L.; Shull, L.M.; Batachari, L.E.; Dillon, M.; Evans, C.; Becker, C.J.; et al. Genome-Wide Transcriptional Response to Varying RpoS Levels in Escherichia coli K-12. J. Bacteriol. 2017, 199, e00755-16. [Google Scholar] [CrossRef]
- Weber, H.; Polen, T.; Heuveling, J.; Wendisch, V.F.; Hengge, R. Genome-wide analysis of the general stress response network in Escherichia coli: σS-dependent genes, promoters, and sigma factor selectivity. J. Bacteriol. 2005, 187, 1591–1603. [Google Scholar] [CrossRef]
- Schellhorn, H.E. Function, Evolution, and Composition of the RpoS Regulon in Escherichia coli. Front. Microbiol. 2020, 11, 560099. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Gómez, P.; López, M.; Prieto, M.; González-Raurich, M.; Alvarez-Ordóñez, A. The role of the general stress response regulator RpoS in Cronobacter sakazakii biofilm formation. Food Res. Int. 2020, 136, 109508. [Google Scholar] [CrossRef] [PubMed]
- Alattas, H.; Wong, S.; Slavcev, R.A. Identification of Escherichia coli Host Genes That Influence the Bacteriophage Lambda (λ) T4rII Exclusion (Rex) Phenotype. Genetics 2020, 216, 1087–1102. [Google Scholar] [CrossRef]
- Guo, Y.; Li, Y.; Zhan, W.; Wood, T.K.; Wang, X. Resistance to oxidative stress by inner membrane protein ElaB is regulated by OxyR and RpoS. Microbical Biotechnol. 2019, 12, 392–404. [Google Scholar] [CrossRef]
- Bouillet, S.; Genest, O.; Méjean, V.; Iobbi-Nivol, C. Protection of the general stress response σS factor by the CrsR regulator allows a rapid and efficient adaptation of Shewanella oneidensis. J. Biol. Chem. 2017, 292, 14921–14928. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Y.; Chen, Y.; Xu, F.; Halliday, N.; Gao, K.; Chan, K.G.; Cámara, M. RpoS differentially affects the general stress response and biofilm formation in the endophytic Serratia plymuthica G3. Res. Microbiol. 2016, 167, 168–177. [Google Scholar] [CrossRef]
- Campos-Gomez, J.; Benitez, J.A. A simple cell-based high throughput screening (HTS) assay for inhibitors of Salmonella enterica RNA polymerase containing the general stress response regulator RpoS (σS). J. Microbiol. Methods 2018, 150, 1–4. [Google Scholar] [CrossRef]
- Chen, J.; Wassarman, K.M.; Feng, S.; Walz, T.; Campbell, E.A.; Darst, S.A. 6S RNA Mimics B-Form DNA to Regulate Escherichia coli RNA Polymerase. Mol. Cell 2017, 68, 388–397.e6. [Google Scholar] [CrossRef]
- Samuels, D.S.; Lybecker, M.C.; Yang, X.F.; Ouyang, Z.; Bourret, T.J.; Boyle, W.K.; Stevenson, B.; Drecktrah, D.; Caimano, M.J. Gene Regulation and Transcriptomics. Curr. Issues Mol. Biol. 2021, 42, 223–266. [Google Scholar] [CrossRef]
- Oguienko, A.; Petushkov, I.; Pupov, D.; Esyunina, D.; Kulbachinskiy, A. Universal functions of the σ finger in alternative sigma factors during transcription initiation by bacterial RNA polymerase. RNA Biol. 2021, 18, 2028–2037. [Google Scholar] [CrossRef]
- Gaal, T.; Ross, W.; Estrem, S.T.; Nguyen, L.H.; Burgess, R.R.; Gourse, R.L. Promoter recognition and discrimination by EσS RNA polymerase. Mol. Microbiol. 2001, 42, 939–954. [Google Scholar] [CrossRef] [PubMed]
- Jaishankar, J.; Srivastava, P. Molecular Basis of Stationary Phase Survival and Applications. Front. Microbiol. 2017, 8, 2000. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Xu, J.; Liu, X.; Chu, X.; Wang, P.; Tian, J.; Wu, N.; Fan, Y. Identification of a highly efficient stationary phase promoter in Bacillus subtili. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Utsumi, R.; Kusafuka, S.; Nakayama, T.; Tanaka, K.; Takayanagi, Y.; Takahashi, H.; Noda, M.; Kawamukai, M. Stationary phase-specific expression of the fic gene in Escherichia coli K-12 is controlled by the rpoS gene product (σ38). FEMS Microbiol. Lett. 1993, 113, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Logel, D.Y.; Trofimova, E.; Jaschke, P.R. Codon-Restrained Method for Both Eliminating and Creating Intragenic Bacterial Promoters. ACS Synth. Biol. 2022, 11, 689–699. [Google Scholar] [CrossRef]
- Yin, K.; Guan, Y.; Ma, R.; Wei, L.; Liu, B.; Liu, X.; Zhou, X.; Ma, Y.; Zhang, Y.; Waldor, M.K.; et al. Critical role for a promorter discriminator in RpoS control of virulence in Edwardsiella piscicida. PLoS Pathog. 2018, 14, e1007272. [Google Scholar] [CrossRef]
- Dudin, O.; Lacour, S.; Geiselmann, J. Expression dynamics of RpoS/Crl-dependent genes in Escherichia coli. Res. Microbiol. 2013, 164, 838–847. [Google Scholar] [CrossRef]
- Typas, A.; Hengge, R. Role of the spacer between the −35 and −10 regions in sigmas promoter selectivity in Escherichia coli. Mol. Microbiol. 2006, 59, 1037–1051. [Google Scholar] [CrossRef]
- Rosenthal, A.Z.; Hu, M.; Gralla, J.D. Osmolyte-induced transcription: −35 region elements and recognition by sigma38 (rpoS). Mol. Microbiol. 2006, 59, 1052–1061. [Google Scholar] [CrossRef]
- Hengge-Aronis, R. Stationary phase gene regulation: What makes an Escherichia coli promoter σS-selective? Curr. Opin. Microbiol. 2002, 5, 591–595. [Google Scholar] [CrossRef]
- Miksch, G.; Bettenworth, F.; Friehs, K.; Flaschel, E. The sequence upstream of the −10 consensus sequence modulates the strength and induction time of stationary-phase promoters in Escherichia coli. Appl. Microbiol. Biotechnol. 2005, 69, 312–320. [Google Scholar] [CrossRef]
- Labun, K.; Montague, T.G.; Krause, M.; Cleuren, Y.N.T.; Tjeldne, H.; Valen, E. CHOPCHOP v3: Expanding the CRISPR web toolbox beyond genome editing. Nucleic Acids Res. 2019, 47, W171–W174. [Google Scholar] [CrossRef]
- Todor, H.; Osadnik, H.; Campbell, E.A.; Myers, K.S.; Li, H.; Donohue, T.J.; Gross, C.A. Rewiring the specificity of extracytoplasmic function sigma factors. Proc. Natl. Acad. Sci. USA 2020, 117, 33496–33506. [Google Scholar] [CrossRef]
- Ionescu, M.; Belkin, S. Overproduction of exopolysaccharides by an Escherichia coli K-12 rpoS mutant in response to osmotic stress. Appl. Environ. Microbiol. 2009, 75, 483–492. [Google Scholar] [CrossRef]
- Patange, O.; Schwall, C.; Jones, M.; Villava, C.; Griffith, D.A.; Phillips, A.; Locke, J.C.W. Escherichia coli can survive stress by noisy growth modulation. Nat. Commun. 2018, 9, 5333. [Google Scholar] [CrossRef]
- Utsumia, R.; Nakayamaa, T.; Iwamotoa, N.; Kohdaa, K.; Kawamukaib, M.; Tanabea, H.; Tanakac, K.; Takahashic, H.; Nodaa, M. Mutational analysis of the fic promoter recognized by RpoS (σ38) in Escherichia coli. Biosci. Biotechnol. Biochem. 1995, 59, 1573–1575. [Google Scholar] [CrossRef]
- Gao, L.; Wu, X.; Li, C.; Xia, X. Exploitation of strong constitutive and stress-driven promoters from Acetobacter pasteurianus for improving acetic acid tolerance. J. Biotechnol. 2022, 350, 24–30. [Google Scholar] [CrossRef]
- Song, Y.; Nikoloff, J.M.; Fu, G.; Chen, J.; Li, Q.; Xie, N.; Zheng, P.; Sun, J.; Zhang, D. Promoter Screening from Bacillus subtilis in Various Conditions Hunting for Synthetic Biology and Industrial Applications. PLoS ONE 2016, 11, e0158447. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Q.; Weng, H.; Shi, Y.; Chen, J.; Du, G.; Kang, Z. Construction of Synthetic Promoters by Assembling the Sigma Factor Binding −35 and −10 Boxes. Biotechnol. J. 2019, 14, e1800298. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Tong, S.; Amin, F.R.; Khalid, H.; Chen, K.; Zhao, X.; Cai, J.; Li, D. Enhancing the Activity of a Self-Inducible Promoter in Escherichia coli through Saturation Mutation and High-Throughput Screening. Fermentation 2023, 9, 468. https://doi.org/10.3390/fermentation9050468
Li J, Tong S, Amin FR, Khalid H, Chen K, Zhao X, Cai J, Li D. Enhancing the Activity of a Self-Inducible Promoter in Escherichia coli through Saturation Mutation and High-Throughput Screening. Fermentation. 2023; 9(5):468. https://doi.org/10.3390/fermentation9050468
Chicago/Turabian StyleLi, Jinyang, Sheng Tong, Farrukh Raza Amin, Habiba Khalid, Kai Chen, Xiaoguang Zhao, Jinling Cai, and Demao Li. 2023. "Enhancing the Activity of a Self-Inducible Promoter in Escherichia coli through Saturation Mutation and High-Throughput Screening" Fermentation 9, no. 5: 468. https://doi.org/10.3390/fermentation9050468
APA StyleLi, J., Tong, S., Amin, F. R., Khalid, H., Chen, K., Zhao, X., Cai, J., & Li, D. (2023). Enhancing the Activity of a Self-Inducible Promoter in Escherichia coli through Saturation Mutation and High-Throughput Screening. Fermentation, 9(5), 468. https://doi.org/10.3390/fermentation9050468