Abstract
Foodborne prevention and treatment of hyperuricemia (HUA) has received widespread attention. Lactic acid bacteria (LAB) can improve intestinal function, while traditional medicine dandelion has the functions of detoxification and detumescence. Whether LAB fermented dandelion has any effects on HUA and the underlying mechanism is not clear. To address these questions, Lactobacillus acidophilus was selected or maximal xanthine oxidase activity. The effect of Lactobacillus acidophilus fermented dandelion (LAFD) on uric acid metabolism was evaluated by the HUA mouse model. Expression levels of UA, BUN, CRE, XOD, and inflammatory factors in serum were detected. Paraffin sections and staining were used to observe the kidney and small intestine, and mRNA expression of GLUT9, URAT1, OAT1, and ABCG2 related to uric acid metabolism were investigated. Furthermore, the intestinal flora was studied by contents of the cecum and high throughput 16S rRNA sequencing. The results showed that LAFD had a significant inhibitory effect on XOD in vitro (p < 0.01). LAFD could reduce the levels of UA, BUN, CRE, XOD, IL-1 β, IL-6, and TNF- α in serum (p < 0.05), thus inhibiting inflammatory reaction, and reducing UA by decreasing the mRNA expression of GLUT9, URAT1 in kidney and increasing the mRNA expression of OAT1 and ABCG2 in kidney and small intestine (p < 0.05). In addition, the 16S rRNA gene sequencing analysis demonstrated that LAFD treatment can help restore the imbalance of the intestinal microbial ecosystem and reverse the changes in Bacterodietes/Firmicutes, Muribaculaceae, Lachnospiraceae in mice with HUA. It is suggested that the mechanism of LAFD in treating HUA may be related to the regulation of the mRNA expressions of GLUT9, URAT1, OAT1, and ABCG2 in the kidney and small intestine, as well as the regulation of intestinal flora, which provides the experimental basis for the development of new plant fermented products.
1. Introduction
Hyperuricemia (HUA) is a chronic clinical syndrome, where the blood uric acid content is significantly higher than the normal value due to the disorder of purine metabolism and abnormal excretion of uric acid in the human body [1]. HUA is not only the basic stage of gout but also easily induces cardiovascular disease, diabetes, chronic kidney disease, non-alcoholic fatty liver, etc [2]. At present, the treatment and prevention measures for HUA mainly use the effect of exogenous drugs and restrict the intake of purine substances in the diet. Most clinical uric-acid-lowering drugs rely on xanthine oxidase inhibitors (allopurinol) and uricosuric drugs (benzbromarone). Long-term use will cause serious adverse reactions in the human body [3,4]. Therefore, it is urgent to find new ways to reduce uric acid in the blood of the HUA population, and it is of great significance to explore a new type of HUA treatment and prevention method with low toxicity and side effects and certain food origin.
Dandelion (Taraxacum mongolicum Hand.-Mazz.) is a perennial herb of Compositae. Dandelion contains a variety of functional ingredients, such as phenolic acids, flavonoids, polysaccharides, terpenoids, and sterol compounds, etc., which have anti-inflammatory [5], anti-oxidation [6], anti-cancer [7], hypoglycemic [8], anti-obesity [9], immune regulation [10], and traditional heat-clearing and detoxifying bioactivities. Previous studies have shown that the water extract of dandelion can reduce the serum uric acid content of rats with HUA [11], and has a good protective effect on rat kidney damage caused by persistently elevated uric acid levels [12].
The use of micro-organisms to ferment traditional Chinese medicine has the advantages of reducing toxicity, expanding drug efficacy, and improving absorption rate [13]. During fermentation, substrates such as medicinal plants are decomposed or converted into compatible components, thereby improving the properties of the substrate through the production and enhancement of bioactive compounds. Fermentation mainly improves the pharmacological properties of traditional Chinese medicine through the modification of natural molecules (such as flavonoids, phenolic acids, saponins, phytosterols, etc.) to better promote health and prevent diseases [14]. The fermentation process of lactic acid bacteria (LAB) can produce new active substances, such as exopolysaccharides, and mild microbial fermentation conditions can protect the active ingredients to a certain extent [13]. For example, the use of Lactobacillus plantarum to ferment cinnamon can enhance phenolic substances and flavonoids and improve antioxidant and anticancer activities without producing toxic substances [15]. Mulberry leaf extract fermented with Lactobacillus acidophilus A4 can better improve 5-fluorouracil-induced intestinal mucositis in rats [16]. Artemisia selengensis Turcz fermented by Lactobacillus acidophilus can significantly increase the content of free polyphenols, enhance the inhibition of xanthine oxidase (XOD) in vitro, and have a better effect on alleviating HUA than the unfermented counterpart [17]. Recently, our research group carried out some work in the field of two-way fermentation of traditional Chinese medicine and found that the fermentation liquid obtained by fermenting plants with edible micro-organisms has a variety of active ingredients and functions [18,19,20]. The antioxidant phenols and flavonoids in the fermentation supernatant, such as ellagic acid, vanillin, luteolin, kaempferol, myricetin, isorhamnetin, and (+)-gallocatechin, by bidirectional metabolites from Tremella fuciformis and Acanthopanax trifoliatus was found increased significantly [18]. The exopolysaccharide yield and antioxidant activity of the fermentation products of Schizophyllum commune were increased by the addition of Radix Puerariae [19], and the fermentation supernatant could prolong the lifespan of Caenorhabditis elegans and improve stress resistance [20].
Whether dandelion fermented by LAB has any effects on lowering uric acid and how it works has not been reported. Therefore, this study screened the optimal fermentation strains through in vitro XOD inhibition testing to explore the effect of fermented dandelion on HUA animal models. Additionally, the underlying mechanism is investigated through inflammatory factors, mRNA expression levels, and gut microbiota. This work is aimed to provide the experimental basis for the research and development of new dandelion-fermented products.
2. Materials and Methods
2.1. Materials
Dandelion was purchased from Guangdong Shizhen Pharmaceutical Co., Ltd. (Guangzhou, China), batch number 211101; Lactobacillus plantarum GDMCC1.648, GDMCC1.191, GDMCC1.140, Lactobacillus acidophilus GDMCC1.412, and Lactobacillus casei GDMCC1.159 were purchased from Guangdong microbial strain Preservation Center; Lactobacillus rhamnosus ATCC7469 and MRS culture medium were purchased from Guangdong Huankai Microbiological Technology Co., Ltd. (Guangzhou, China) UA, BUN, CRE, XOD, ALT, AST kits were purchased from Nanjing Jiancheng Reagent Co., Ltd. (Nanjing, China); and IL-6, IL-1 β, and TNF- α Elisa kits were purchased from Jiangsu Enzymatic Immunization Co., Ltd. (Nanjing, China).
2.2. Preparation of Fermented Dandelion
The dried dandelion was crushed and sifted through 50 meshes, and MRS liquid medium was added according to the ratio of material to liquid at 1:10, sterilized at 121 °C for 15 min, then cooled the sterilized liquid medium naturally to room temperature. According to the inoculation amount of 5%, Lactobacillus plantarum, Lactobacillus acidophilus, Lactobacillus casei, and Lactobacillus rhamnose were inoculated twice and fermented at 37 °C for 4 days to obtain dandelion fermentation broth. Unfermented dandelion was set as the control group.
2.3. Determination of the Inhibitory Activity of XOD
The inhibitory activity of XOD was performed with a minor modification described previously [21,22]. Add 50 μL XOD solution of 0.25 U/mL to 50 μL dandelion or fermented dandelion solution to react. After incubating for 10 min at 37 °C, 50 μL xanthine solution was added to start the reaction. After the next 10 min, the absorbance of the reaction system was recorded under 295 nm. The inhibition rates of fermented traditional Chinese medicine extracts with concentrations of 1.25, 2.5, 5, 10, 15, and 20 mg/mL of XOD solution were determined, and three compound holes were set for each concentration. In this experiment, the XOD solution concentrations of 0, 0.125, 0.25, 0.5,1, 1.5 U/mL and enzyme reaction times of 5, 10, 15, 20, and 25 min were investigated, and it was determined that the enzyme concentration of 0.25 U/mL and enzyme reaction time of 10 min were used as the reaction system. The drug concentration where the inhibition rate was 50%, namely the half inhibitory concentration (IC50), was calculated to determine the optimal fermentation strain.
2.4. Determination of Total Phenolic Content and Total Flavonoid Content
The content of total phenol was determined by the Folin-Ciocalteu colorimetric method with slight modifications [23]. The solution to be tested was mixed with 1 mL Folin reagent (0.6 mol/L), and the 2 mL Na2CO3 solution (0.15 g/mL) was added quickly. After mixing, the volume was fixed to 25 mL, and the mixture solution was incubated for 50 min avoiding light at room temperature. The absorbance value was determined under 756 nm. The total phenol content was expressed as the Gallic acid equivalent (mg GAE/g).
The total flavonoid content was determined by the AlCl3 colorimetric method with minor modifications [24]. The total flavonoid contents were determined by the aluminum nitrate colorimetric method. The solution to be measured was placed in a 10 mL measuring flask. Distilled water of 5 mL and 5% sodium nitrite of 0.3 mL was added to the flask separately, then shaken the mixture and standed it for 6 min. Ten percent aluminum nitrate of 0.3 mL was added, After mix and stand it for 6 min, 4 mL sodium hydroxide of 4% concentration was added, Finally, water was added to the scale. The absorbance value of the mixed solution was determined at 510 nm after intensive mixing and standing for 15 min at room temperature. Rutin is used as a standard, and data are calculated in milligrams of rutin equivalent (mg RE/g) per gram of sample.
2.5. Antioxidant Activity Assay
2.5.1. Determination of DPPH Free Radical Scavenging Activity
According to the reference with slight modifications [25], 1 mg of DPPH was dissovled in 10 mL anhydrous ethanol, and add 2 mL DPPH-anhydrous ethanol solution at a concentration of 0.1 mg/mL. Then add the solution to be tested and mix them well, let the mixed solution stand for 5 min in the dark at room temperature, and measure the absorbance at 517 nm. Using ultrapure water zero, ascorbic acid solution as a positive control group, and DPPH solution as a blank control group, effective concentrations of DPPH free radical scavenging activity and 50% free radical scavenging activity(EC50) were calculated according to the following formula.
DPPH free radical scavenging activity (%) = (A0 − A1)/A0 × 100%
In the formula, A0 stands for the absorbance value of the blank solution, while A1 stands for the absorbance value of the sample.
2.5.2. Determination of ABTS Free Radical Scavenging Activity
According to the previous work with slight modifications [26], the test solution of 1 mL was taken into the plug test tube and mixed with ABTS working liquid, and the absorbance A1 was determined at 734 nm for 10 min at 37 °C. The blank group used distilled water instead of the ABTS solution, and the control group used distilled water instead of samples.
ABTS free radical scavenging activity (%) = (1 − (A1 − A2)A0) × 100%
In the formula, A0: the absorbance value of the control solution; A1: The absorbance value of the sample; A2: the absorbance value of the blank solution.
2.5.3. Determination of Reducing Power
Reducing power was determined according to the reference [27] with slight modifications, take 1.5 mL of the solution to be tested into a plug test tube, then add 1.5 mLphosphoric acid solution (0.2 mol/L, pH = 6.0), and 1.5 mL 1% potassium ferricyanide, The mixture was kept in a water bath for 30 min at 50 °C, then taken it out and cooled to room temperature, add 1.5 mL trichloroacetic acid of 10% concentration was added to the mixture, place for 3 min, remove 2 mL 0.1% FeCl3, fully react for 10 min, and determine the absorbance at 700 nm.
2.6. Animal Grouping, Modeling, and Administration
In total, 70 male Kunming mice (20 ± 2 g) were purchased from Zhejiang Weitong Lihua Experimental Animal Technology Co., Ltd. (Jiaxing, China) (quality certificate: SCXK (Zhe) 2019-0001). After 7 days of adaptive feeding, they were randomly divided into the control group, model group, allopurinol group (0.02 g/kg/day), fermented dandelion low and high dose groups (0.1, 0.4 g/kg/day), dandelion low and high dose groups (0.1, 0.4 g/kg/day), with 10 rats in each group. The mouse model of HUA was established by continuous intragastric administration of potassium oxonate(300 mg/kg) and hypoxanthine (300 mg/kg) once per day. All groups except for the control group were established as HUA model mice, and the corresponding treatment drug was administered 1 h after the administration of the modeling drug, starting on day 8 for 4 weeks. During the 5 weeks experiment, the body weight of mice was recorded once a week. At the end of the experiment, serum was collected by centrifugation (4 °C, 4000× g for 10 min) and stored at −80°C until further analysis. Complete kidneys were collected, washed, weighed, and part of the kidney tissue was fixed in 4% paraformaldehyde, and the other part was stored at −80°C. The ileum was cut out and rinsed with saline; part of the ileum was fixed in 4% paraformaldehyde and the rest was stored at −80°C. Cut off the cecum and squeeze out the contents of the cecum with sterile forceps into sterile Ep tubes and store at −80°C.
2.7. Serum Index Detection
Uric acid (UA), creatinine (CRE), and urea nitrogen (BUN) levels in serum and XOD were measured according to the kit instructions; the levels of related inflammatory factors TNF-α, IL-1β, and IL-6 were measured by enzyme-linked immunoassay (ELISA).
2.8. Morphological Observation
The kidney and ileum of mice were transferred from 4% paraformaldehyde solution, cut into small squares, placed in the embedding box, rinsed for 2 h using running water, dehydrated with gradient ethanol, paraffin wax immersion, embedding, sectioning, HE staining, and other steps, observed under the microscope, and images were collected to analyze the condition of the kidney and ileum.
2.9. qRT-PCR Analysis
Total RNA from mouse kidney and ileal tissues was extracted with the Trizol reagent. After homogenization, 1/5 volume of chloroform was added and centrifuged for separation. The supernatant was further precipitated with an equal volume of isopropanol to obtain the RNA precipitate. After two rounds of washing with 75% ethanol, it was then solubilized with DEPC water. Reverse transcription was performed by referring to the Servicebio®RT First Strand cDNA Synthesis Kit. qRT-PCR was run in the following conditions: 95 °C, 30 s; 95 °C, 15 s; 60 °C, 30 s. The primer sequences for genes URAT1, GLUT9, OAT1, ABCG2 are shown in Table 1. GAPDH was used as an internal standard for quantification, and the relative expression of target gene mRNA was calculated by the 2−∆∆Ct method.

Table 1.
Primer sequences of qRT-PCR.
2.10. Gut Microbiological Analysis
DNA extraction from the contents of the cecum of each group of mice was performed according to the instructions of the DNA kit, and the DNA concentration and purity were checked by 1% agarose gel electrophoresis for conformity. V3 and V4 hypervariable regions of 16S rRNA were selected and amplified using the primer pairs: F: ACTCCTACGGGAGGCAGCA; R: GGACTACHVGGGTWTCTAAT. The Illumina Novaseq platform is used for sequencing, and the original sequencing sequence uses Trimmatic v0.33 and cutadapt 1.9.1 software to filter the quality of each sample. The dada2 method in QIIME2 2020.6 is used for noise removal; the two-end sequence is spliced, and the chimeric sequence is removed to obtain the final effective data. Operational taxonomy units (OTUs) with similarity at 97% were selected to calculate the diversity index (USEARCH v10.0). Using QIIME2 2020.6 to generate species abundance tables at different taxonomic levels; additionally, analyze and evaluate Alpha diversity index of the sample, including the Chao1, Shannon, Simpson indexes. Beta diversity analysis was performed using QIIME software to compare the similarity of different samples in terms of species diversity. Principal coordinates analysis (PCoA) examined the abundance and diversity of OTUs. LefSe analysis (http://huttenhower.sph.harvard.edu/lefse/ accessed on 26 October 2022.) was used to estimate the magnitude of the effect of each species’ abundance on the differential effect using linear discriminant analysis (LDA = 3.5).
2.11. Statistical Analysis
The statistical analysis and plots were performed using IBM SPSS Statistics 26 and GraphPadPrism8 software. The results were expressed as mean ± standard deviation (SD). Data were analyzed using one-way ANOVA, and significant differences between the two groups were analyzed using a t-test. p < 0.05 indicates statistical significance.
3. Results
3.1. Determination of XOD Inhibitory Activity
The IC50 of dandelion and each fermentation solution on XOD was determined and the results are shown in Table 2. The IC50 value of Lactobacillus acidophilus fermented dandelion(LAFD) was the smallest at 15.55 ± 2.31 mg/mL, which was significantly different from the unfermented group (p < 0.01) and was therefore determined as the optimal fermenting strain for subsequent experiments.
Table 2.
IC50 of dandelion and each fermentation solution on XOD.
3.2. Content and Antioxidant Activity Determination
The total phenolic and flavonoid contents of unfermented dandelion were 7.88 ± 0.18 mg GAE/g and 23.39 ± 0.34 mg RE/g, respectively. Meanwhile, the total phenolic content and flavonoid of LAFD were 16.05 ± 0.22 mg GAE/g and 20.81 ± 0.19 mg RE/g, respectively. LAFD at least doubled the total phenolic content compared to unfermented dandelion. The slight decrease in total flavonoid content may be explained by the fermentation process continues which the substances in fermented dandelion are utilized, degraded, and transformed by micro-organisms [28]. As shown in Figure 1, the EC50 values of DPPH radical and ABTS radical of LAFD were 1.25 ± 0.02, 0.36 ± 0.01 mg/mL, while those of unfermented dandelion were 2.95 ± 0.08, 0.81 ± 0.02 mg/mL. It is proved that LAFD has a stronger free radical scavenging ability than unfermented dandelion. The greater the reducing power, the stronger the oxidation resistance. The reducing power of LAFD was significantly higher than that of unfermented dandelion. The above results showed that LAFD had better antioxidant activity than unfermented dandelion.
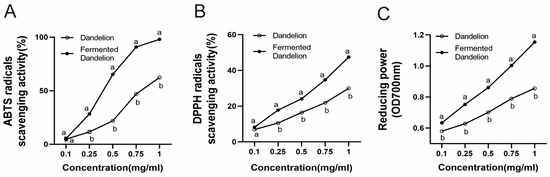
Figure 1.
Determination of the antioxidant activity of dandelion and fermented dandelion. (A) ABTS radical scavenging rate. (B) DPPH radical scavenging rate. (C) Determination of reducing power. The data are expressed as mean ± SD (n = 3). Different lowercase letters indicate statistically significant differences (p < 0.05) between the groups.
3.3. Effects on Serum UA and XOD of Mice with HUA
As shown in Figure 2, the levels of UA and XOD in the model group were significantly higher than those in the blank group (p < 0.001), indicating that potassium oxonate combined with hypoxanthine induced the HUA model successfully. Barakat M. ALRashdi reported that M. quadrangular reduced HUA model serum uric acid, creatinine, urea, and XO activity [29]. Consistent with our study, compared with the model group, each administration group significantly decreased the level of UA in HUA mice (p < 0.001) and showed a certain dose dependence. Among them, LAFD had a better uric-acid-lowering effect than unfermented dandelion (p < 0.05). Allopurinol, D-H, and FD-H, significantly decreased the level of serum XOD in mice (p < 0.001, p < 0.01). Compared with the control group and the ALL group, there was no significant difference in UA and XOD in the FD-H group, which indicated that LAFD had the same curative effect as positive drugs and could effectively restore the levels of UA and XOD.
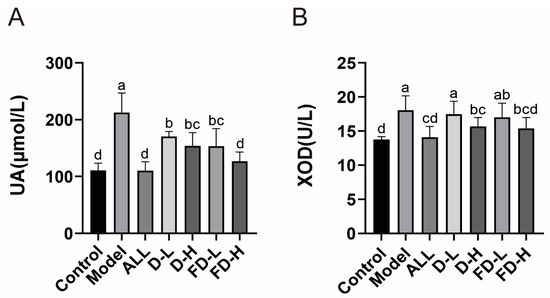
Figure 2.
Effects of fermented dandelion on serum UA and XOD in mice with HUA. (A) Serum UA level. (B) Serum XOD activity. The data are expressed as mean ± SD (n = 8). Different lowercase letters indicate statistically significant differences (p < 0.05) between the groups. ALL, allopurinol; D-L, dandelion low dose group; D-H, dandelion high dose group; FD-L, low dose group of LAFD; FD-H, high dose group of LAFD.
3.4. Effect on Liver and Kidney Function of Mice with HUA
Hyperuricemia can cause certain damages to the liver such as hepatocyte inflammation, apoptosis, and hepatic steatosis [30]. Additionally, the kidneys are one of the most important sites for uric acid excretion, with BUN and CRE also commonly used as indicators of kidney damage from hyperuricaemia [31]. As shown in Figure 3, the levels of ALT, AST, CRE, and BUN in the serum of mice in the model group increased significantly (p < 0.001), indicating that the model of potassium oxonate combined with hypoxanthine caused certain liver and kidney damage, and the levels of ALT and AST in each administration group decreased to varying degrees (p < 0.05, p < 0.001), reducing liver damage. Allopurinol, D-H, and FD-H decreased the CRE level (p < 0.001, p < 0.01, p < 0.01). D-H and FD-H decreased the BUN level (p < 0.01, p < 0.001), reducing renal injury. Interestingly, compared with the control group, there was no significant difference in BUN in the FD-H group, which indicated that LAFD can reduce BUN levels to normal levels. Thus, FD-H was more effective at protecting the kidneys than allopurinol.
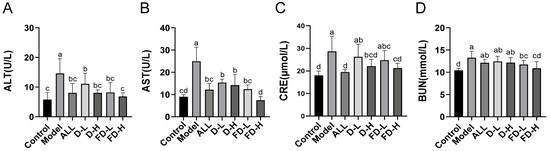
Figure 3.
Effect of fermented dandelion on liver and kidney function in mice with HUA. (A) Serum ALT level. (B) Serum AST level. (C) Serum CRE level. (D) Serum BUN level. The data are expressed as mean ± SD (n = 8). Different lowercase letters indicate statistically significant differences (p < 0.05) between the groups. ALL, allopurinol; D-L, dandelion low dose group; D-H, dandelion high dose group; FD-L, low dose group of LAFD; FD-H, high dose group of LAFD.
3.5. Effects on the Levels of Inflammatory Factors IL-1 β, IL-6, and TNF- α in Mice with HUA
Hyperuricemia leads to inflammation and promotes the production of inflammatory factors, which can lead to gout, urinary stones, uric acid nephropathy, etc [32]. Figure 4. shows that a persistently high level of uric acid would lead to an inflammatory reaction and increase the levels of inflammatory cytokines IL-1 β, IL-6, and TNF- α (p < 0.001). Compared with the model group, the levels of IL-1 β, IL-6, and TNF- α decreased in all treatment groups. Among them, the levels of IL-1 β and IL-6 in the high-dose group before and after LAFD showed significant differences (p < 0.05). There was a significant difference in IL-1β (p < 0.05) and IL-6 (p < 0.001) in the FD-H group and the ALL group, while no significant difference for TNF-α between the FD-H group and the ALL group was detected. Therefore, FD-H had comparable anti-inflammatory activity as the positive drugs.
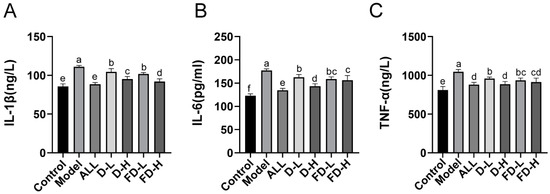
Figure 4.
Effects on the levels of inflammatory factors IL-1 β, IL-6, and TNF- α in mice with HUA. (A) Serum IL-1 β level; (B) Serum IL-6 level; (C) Serum TNF- α level. The data are expressed as mean ± SD (n = 8). Different lowercase letters indicate statistically significant differences (p < 0.05) between the groups. ALL, allopurinol; D-L, dandelion low dose group; D-H, dandelion high dose group; FD-L, low dose group of LAFD; FD-H, high dose group of LAFD.
3.6. Effects on Renal Pathological Changes in Mice with HUA
Elevated serum levels of uric acid can cause structural changes in the kidney, including the glomerulus, capsule, and tubules [33]. As shown in Figure 5A, the histological characteristics of the kidney in the control group were normal, the size of renal tubular epithelial cells was arranged neatly, and no inflammatory cell infiltration was found. Compared with control group, the model group showed glomerular atrophy and more renal tubular injury, such as dilatation of the renal tubule, narrowing of the tubular wall, infiltration of the inflammatory cells, swelling and exfoliation of some renal tubular epithelial cells, and cytoplasmic vacuolization. Compared with the model group, renal tubule dilatation, tube wall narrowing, and inflammatory cell infiltration were alleviated in each treatment group, and the normal histological characteristics were maintained. Additionally, the shape of the glomerulus tended to be more circumference styled in the ALL group, D-H group, and FD-H group compared to the model group.
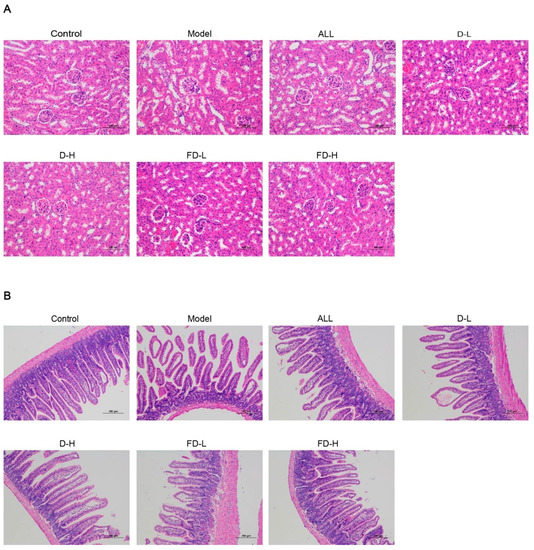
Figure 5.
Effects of fermented dandelion on kidney and ileum of mice with HUA. (A) Renal pathological section (200×). (B) Pathological section of the small intestine (100×). ALL, allopurinol; D-L, dandelion low dose group; D-H, dandelion high dose group; FD-L, low dose group of LAFD; FD-H, high dose group of LAFD.
3.7. Effect on Pathological Changes of Ileum in Mice with HUA
As shown in Figure 5B, the ileal mucosal epithelium of the blank group was intact, the villi were abundant, the glands were rich and orderly arranged, and the epithelial cells were arranged neatly. Compared with the blank group, the ileal mucosa of the model group was exfoliated, the small intestinal villi were sparse and broken, the villi epithelial cells were shedding, and the intestinal glands were atrophied. Compared with the model group, the pathological changes of ileum in ALL group were significantly alleviated. Except for a few villi epithelial cell necrosis, other pathological phenomena were significantly improved. In mice in the dandelion and fermented dandelion groups, the pathological damage of the ileum was alleviated, the morphology was more complete, the epithelial cells of the ileum were arranged closely, and some of the villous epithelial cells were necrotic and exfoliated.
3.8. Effects on the Levels of GLUT9, URAT1, OAT1 mRNA in Kidney and ABCG2 mRNA in Small Intestine in Mice with HUA
As shown in Figure 6, the contents of GLUT9 mRNA and URAT1mRNA in the kidney in the Model group were significantly higher than those in the control group (p < 0.001), while the contents of OAT1 mRNA and ABCG2 mRNA in the small intestine decreased significantly (p < 0.001). Compared with the model group, GLUT9 mRNA expression in the kidney was significantly increased in each group (p < 0.001), URAT1 and OAT1 mRNA expression in the kidney of D-H group was significantly increased (p < 0.01, p < 0.001); the mRNA expression of OAT1 in the kidney in FD-L group was significantly increased (p < 0.001); and the mRNA expression of URAT1 and OAT1 in kidney in the ALL and FD-H groups was significantly increased (p < 0.001). The ALL and FD-H groups could significantly down-regulate the mRNA expression of ABCG2 in the small intestine.
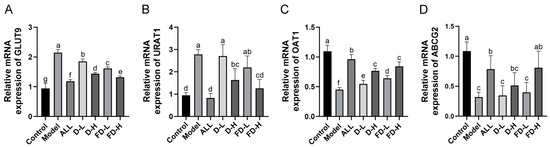
Figure 6.
Effects of fermented dandelion on uric acid transporter mRNA in the kidney and small intestine of mice with HUA. (A) Renal GLUT9 mRNA level; (B) Renal URAT1 mRNA level; (C) Renal OAT1 mRNA level; (D) Small intestine ABCG2 mRNA level. The data are expressed as mean ± SD (n = 4 or 5). Different lowercase letters indicate statistically significant differences (p < 0.05) between the groups. ALL, allopurinol; D-L, dandelion low dose group; D-H, dandelion high dose group; FD-L, low dose group of LAFD; FD-H, high dose group of LAFD.
3.9. Effect on the Intestinal Microflora of Mice with HUA
To detect whether the protective effect of dandelion and fermented dandelion is related to intestinal microflora, the 16S rRNA V3-V4 region of bacteria in the intestinal contents was sequenced. The Venn map showed a decrease in intestinal microflora diversity (Figure 7B) in mice with high uric acid. The Alpha diversity analysis mainly includes the Chao1, Shannon, and Simpson indexes. The results in Figure 7A show that the intestinal microbial diversity in the model group was significantly lower than that in the control group (p < 0.05), while FD-L and FD-H significantly increased the Shannon index (p < 0.05, p < 0.01). In addition, β-diversity analysis was carried out to study the similarity of intestinal microbial community structure among different samples. The PCoA diagram showed that (Figure 7C) the structure of intestinal microbiota in the model group was significantly different from that in the control group. As expected, there was a significant separation between the FD-H group and the model group. The above results show that the diversity of intestinal flora can be restored by fermented dandelion.
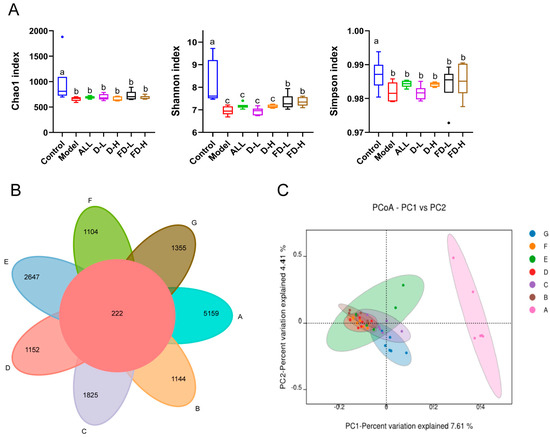
Figure 7.
Effects of fermented dandelion on intestinal microflora diversity in mice with HUA. (A) Alpha diversity analysis; (B) Venn; (C) PCoA. The data are expressed as mean ± SD (n = 6). Different lowercase letters indicate statistically significant differences (p < 0.05) between the groups. A: Control; B: Model; C: ALL, allopurinol; D: D−L, dandelion low dose group; E: D−H, dandelion high dose group; F: FD−L, low dose group of LAFD; G: FD−H, high dose group of LAFD.
The OTU cluster analysis was used to further visualize the composition and structure of each group of intestinal flora at the gate level. As shown in Figure 8A, Bacteroidota and Firmicutes are the main components of intestinal bacterial communities. The Bacteroidota in the model group was significantly higher than that in the control group (p < 0.001) the Firmicutes in the model group was significantly lower than that in the control group (p < 0.001) and the Bacteroidota/Firmicutes in the model group was significantly higher than that in the control group (p < 0.001), while the FD-H group could finally reverse this trend (p < 0.05, p < 0.001, p < 0.001). Similarly, Cao et al. reported that Lactobacillus paracasei X11 could restore the ratio of Bacteroidetes to Firmicutes (Bac/Firm ratio) [34]. Figure 8B shows the results of species flora at the family level. Muribaculaceae, Lachnospiraceae, Prevotellaceae, and Fusobacteriaceae are the dominant bacteria. Yongliang Chu et al. reported that all differential species in Bacteroides, Prevotella, and Fusobacterium were enriched in gout patients [35]. In our study, compared with the control group, Bacteroidetes, Prevotellaceae, and Fusobacteriaceae were increased in the model group. Compared with the control group, Muribaculaceae in the model group increased significantly (p < 0.001), while Lachnospiraceae decreased significantly (p < 0.001). This trend was significantly reversed after the administration of fermented dandelion (p < 0.001). Lee, Y et al. reported that Lacticaseibacillus paracasei MJM60396 modulates hyperuricemia by improving bacterial abundance in the intestinal flora, with increased relative abundance of Lachnospiraceae, which are associated with intestinal barrier integrity [36]. Our findings are consistent with the study, implying that LAFD supplementation may improve the hyperuricemia phenotype in mice via influencing bacterial abundance in the intestinal flora.
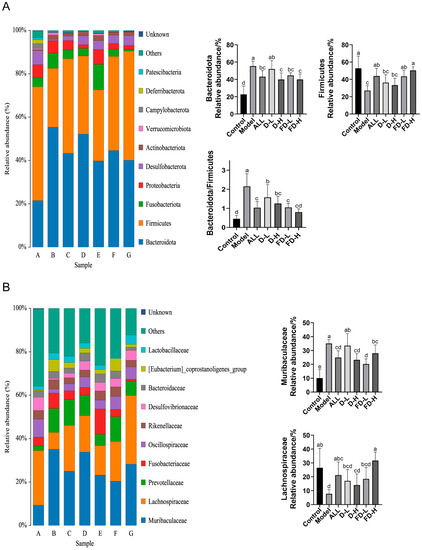
Figure 8.
Effects of fermented dandelion on intestinal microbiota of mice with HUA. (A) Phylum level; (B) Family level. The data are expressed as mean ± SD (n = 6). Different lowercase letters indicate statistically significant differences (p < 0.05) between the groups. A: Control; B: Model; C: ALL, allopurinol; D: D-L, dandelion low dose group; E: D-H, dandelion high dose group; F: FD-L, low dose group of LAFD; G: FD-H, high dose group of LAFD.
To further explore the changes in key microbiota among the groups, LEfSe analysis was used to obtain the cladistic diagram of phyla and genera, and the dominant micro-organisms were shown in each group. As shown in Figure 9A, Clostridia and Firmicutes are the main microorganisms in the control group; Bacteroidales and Muribaculaceae are the main microorganisms in the model group; Lachnospiraceae, Desulfovibrio, and Ligilactobacillusare are the dominant bacteria in the FD-H group. According to the correlation analysis between intestinal microflora and the HUA index (Figure 9B), the correlation between Bacteroidota and the HUA index was the highest, including UA, BUN, CRE, and XOD (p < 0.001). In addition, Firmicutes and Campylobacter were correlated with these four indices (p < 0.05). Acidobacteriota and Verrucomicrobiota were correlated with UA (p < 0.05); Desulfobacterota and Fusobacterioata were correlated with BUN (p < 0.001), Fusobacterioata and Acidobacteriota were correlated with CRE (p < 0.01, p < 0.05), Acidobacteriota and Gemmatimonadota were correlated with XOD(p < 0.01). Based on the above results, we speculate that the intestinal microflora of HUA mice treated with fermented dandelion may play a key role in reducing uric acid.
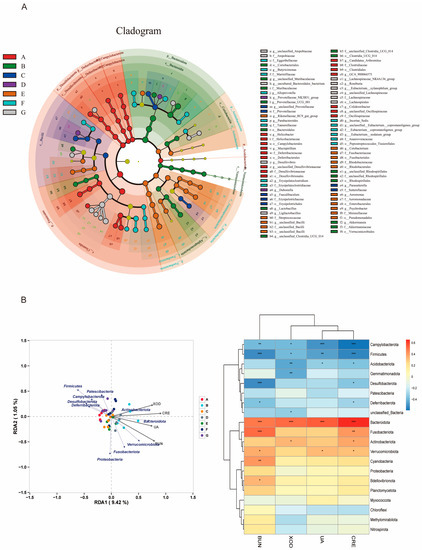
Figure 9.
Effects of fermented dandelion on intestinal microbiota of mice with HUA. (A) LEfSe analysis; (B) Relativity analysis. The data are expressed as mean ± SD (n = 6). Compared with HUA index, * p < 0.05, ** p < 0.01, *** p < 0.001. A: Control; B: Model; C: ALL, allopurinol; D: D−L, dandelion low dose group; E: D−H, dandelion high dose group; F: FD−L, low dose group of LAFD; G: FD−H, high dose group of LAFD.
4. Discussion
The remarkable characteristic of HUA is that the content of uric acid in the blood is higher than the normal level. How to reduce the concentration of serum uric acid has become the key to the treatment of HUA. In recent years, the search for effective or new bioactive compounds from a variety of traditional herbs, natural products, or LAB and other micro-organisms to treat HUA has attracted the people’s attention.
As an important member of human intestinal flora, probiotics have the characteristics of being non-toxic, harmless, having no side effects, and having more advantages than drugs in the treatment of HUA [37]. Additionally, probiotics have shown a variety of benefits, such as immune regulation [38], improving intestinal function and preventing intestinal flora ecological disorders [39], and alleviating inflammation caused by HUA [40]. Additionally probiotics also reduce purine absorption, improve purine metabolism, and inhibit the increase in serum uric acid levels by maintaining intestinal microbial homeostasis [41]. Hsieh, MW et al. screened three kinds of LAB with the best purine degradation. LAB could not only protect renal function but also effectively regulate the concentration of uric acid in the body [42]. Yuanxun Xiao et al. isolated LAB from 18 pickles and proved that LAB can relieve HUA and renal injury through animal experiments [43].
Currently, the comprehensive cooperative research on the two natural active substances of medicinal plants and probiotics has gradually attracted widespread attention. At the same time, people pay more and more attention to the study of traditional Chinese medicine fermented by probiotics. Pei Y et al. fermented bovine bile with Massa Medicata Fermentata. Fermented bovine bile may have anti-inflammatory effects by weakening the activation of NLRP3 inflammatory bodies and enhancing the anti-inflammatory effect of bovine bile [44]. Le Su et al. found that compared with Salvia miltiorrhiza, the fermentation products of Salvia miltiorrhiza could more effectively alleviate the experimental ulcerative colitis induced by sodium dextran sulfate in mice [45]. Therefore, in this experiment, LAB was selected to ferment dandelion to explore the effect and mechanism of HUA.
Dandelion polyphenols have strong antioxidant, anticancer, antibacterial, antiviral, and anti-inflammatory activities [46]. The content of polyphenols in LAFD was doubled and the antioxidant activity was significantly enhanced. XOD is a key factor in purine catabolism, which catalyzes the metabolism of hypoxanthine and xanthine to uric acid. Inhibition of XOD activity is beneficial to reducing uric acid [47]. To effectively screen the fermentation strains to reduce HUA, six kinds of LAB were evaluated by the XOD inhibition test in vitro. We found that LAFD has the smallest IC50 and the strongest inhibitory effect on XOD. As an important intestinal probiotic in the family of LAB, Lactobacillus acidophilus can participate in the host’s intestinal regulation through the production of metabolites and the regulation of intestinal microflora [48]. Studies have shown that Lactobacillus acidophilus reduces renal fibrosis in mice with chronic kidney disease through immunomodulatory effects [49]. Lactobacillus acidophilus fermented Artemisia annua (AST) can increase the expression of OAT1, decrease the expression of URAT1 and GLUT9 in HUA rats, and increase the abundance of intestinal beneficial bacteria [17].
In our study, LAFD has better uric-acid-lowering effect and liver and kidney protective function than unfermented dandelion. Uric acid stimulates inflammatory mediators and causes inflammatory responses [50]. HUA promotes the expression of inflammatory cytokines, such as IL-1 β, TNF- α, and IL-6 [51]. Studies have confirmed that heat-clearing and detoxifying herbs can improve the injury induced by high uric acid by inhibiting the inflammatory reaction of the body. In addition to dandelion, Poria cocos could alleviate paw edema in mice with HUA and gout, decrease serum UA, TNF- α, IL-1β, IL-6, and IL-12, and improve the infiltration of inflammatory cells in renal tubule and joint synovium [52]. In this experiment, LAFD significantly improved the levels of IL-6, TNF- α, and IL-1β in the serum of HUA mice, and alleviated renal and intestinal inflammation.
Uric acid is mainly excreted by the kidney (65–75%) and the intestinal tract (25–35%). Uric acid transporter is necessary for uric acid renal treatment, which can be divided into reabsorption-related proteins and secretion-related proteins. Among them, URAT1 reabsorbs uric acid in the apical membrane of renal proximal tubular epithelial cells (TECs) [53]. As a transporter, GLUT9 can reabsorb uric acid and glucose into renal tubular epithelial cells [54]. The intestinal tract is an important organ for uric acid excretion. The endogenous uric acid in the blood is secreted directly into the intestinal lumen of all intestinal segments through the urate transporter [55]. The excretion of uric acid through the intestine depends on the intestinal homeostasis maintained by the intestinal mucosal barrier [56]. ABCG2 is expressed in the parietal membrane and plays an important role in uric acid excretion [57]. Apigenin reduces uric acid by reducing renal GLUT9 and URAT1 levels and increasing OAT1 levels [58]. Chicory promotes intestinal uric acid excretion by regulating ABCG2 in the intestinal tract of the HUA model [59]. In this study, LAFD decreased the mRNA expression of GLUT9 and URAT1 in the kidney of HUA mice, increased the mRNA expression of OAT1 in the kidney and ABCG2 in the small intestine, and effectively excreted uric acid in the kidney and intestine of HUA mice.
The intestinal microbiota is another key factor in the pathogenesis of HUA [60]. In general, the changes in intestinal microflora observed after treatment may be attributed to the remission of the disease or the effect of therapeutic drugs [61]. Therefore, the study of the relationship between intestinal microflora and drug therapy is helpful to determine the biomarkers of disease remission or to clarify the mechanism of therapeutic drugs. It is reported that HUA leads to the imbalance of intestinal microflora, including the increase in pathogenic bacteria and the decrease in intestinal microbial diversity [62]. In this study, LAFD restored the composition of intestinal microflora of HUA mice and enhanced the α and β diversity of intestinal microflora.
Based on the gut kidney axis, we investigated the changes in the composition of intestinal flora, which may lead to renal damage. Under normal circumstances, the human intestinal microbiome is in equilibrium, and the main bacteria are Firmicutes and Bacteroidota, accounting for >95% of intestinal bacteria [63,64]. In this study, LAFD decreased the relative abundance of Bacteroidota, increased the relative abundance of Firmicutes, and decreased the ratio of Bacteroides to Firmicutes. At the family level, it decreased the relative abundance of Muribaculaceae and restored the relative abundance of Lachnospiraceae. Supplementation of short-chain fatty acids (SCFAs) could significantly improve the intestinal barrier function and renal function [65,66]. Studies have reported that Bacteroides mainly produce acetate and propionate, while thick-walled bacteria produce more butyrate [67]. Butyrate is considered to be a health-promoting molecule because it can increase insulin sensitivity [68], exert anti-inflammatory activity [69], and regulate energy metabolism [70]. Lachnospiraceae is one of the major producers of SCFAs and can regulate inflammation and the immune system by producing butyrate [71,72]. It can be inferred from these results that LAFD may regulate intestinal immune homeostasis by regulating intestinal microflora. To the best of our knowledge, this is the first report related to the Lactobacillus acidophilus fermented dandelion. However, the exact mechanism between the chemical composition of LAFD and its regulation of HUA needs further study.
5. Conclusions
In this study, we screened the optimal fermentative strain of Lactobacillus acidophilus from lactic acid bacteria with in vitro XOD inhibition assay. LAFD treats hyperuricemia in multiple ways: (1) reducing XOD concentration and decreasing uric acid synthesis; (2) improving liver and kidney function and inhibiting inflammation; (3) promoting uric acid excretion by regulating the mRNA expression of GLUE9, URAT4, OAT1, ABCG2; (4) enhancing flora diversity and maintaining intestinal homeostasis. In conclusion, our findings may provide the key evidence that LAFD may be a promising drug candidate for the treatment of HUA and related diseases.
Author Contributions
Conceptualization, Q.M., M.C. and Y.L.; Funding acquisition, L.Z. and X.H.; Investigation, Q.M.; Methodology, Q.M., M.C., Y.L., Y.T. and L.W.; Project administration, L.Z. and X.H.; Supervision, L.Z. and X.H.; Validation, M.C., T.L. and J.W.; Writing—original draft, Q.M.; Writing—review and editing, B.H., L.Z. and X.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China ( grants 82274357 to Lin Zhou), China.
Institutional Review Board Statement
The animal study protocol was approved by the Tab of GuangDong Pharmaceutical University Experimental Animal Ethics Committee Inspection (protocol code: gdpulacspf2017689, date of approval: 29 April 2022).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yanai, H.; Adachi, H.; Hakoshima, M.; Katsuyama, H. Molecular Biological and Clinical Understanding of the Pathophysiology and Treatments of Hyperuricemia and Its Association with Metabolic Syndrome, Cardiovascular Diseases and Chronic Kidney Disease. Int. J. Mol. Sci. 2021, 22, 9221. [Google Scholar] [CrossRef] [PubMed]
- Agnoletti, D.; Cicero, A.F.; Borghi, C. The Impact of Uric Acid and Hyperuricemia on Cardiovascular and Renal Systems. Cardiol. Clin. 2021, 39, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Stamp, L.K.; Chapman, P.T. Allopurinol hypersensitivity: Pathogenesis and prevention. Best Pr. Res. Clin. Rheumatol. 2020, 34, 101501. [Google Scholar] [CrossRef] [PubMed]
- Imai, S.; Nasuhara, Y.; Momo, K.; Oki, H.; Kashiwagi, H.; Sato, Y.; Miyai, T.; Sugawara, M.; Takekuma, Y. Implementation Status of Liver Function Tests for Monitoring Benzbromarone-Induced Hepatotoxicity: An Epidemiological Survey Using the Japanese Claims Database. Biol. Pharm. Bull. 2021, 44, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhao, H.; Gao, Y.; Meng, Y.; Zhao, X.X.; Pan, S.N. Effects of Dandelion Extract on the Proliferation of Rat Skeletal Muscle Cells and the inhibition of a Lipopolysaccharide-lnduced Inflammatory Reaction. Chin. Med. J. 2018, 131, 1724–1731. [Google Scholar] [CrossRef]
- Sun, Y.; Wu, Y.; Wang, Z.; Chen, J.; Yang, Y.; Dong, G. Dandelion Extract Alleviated Lipopolysaccharide-Induced Oxidative Stress through the Nrf2 Pathway in Bovine Mammary Epithelial Cells. Toxins 2020, 12, 496. [Google Scholar] [CrossRef]
- Kang, L.; Miao, M.S.; Song, Y.G.; Fang, X.Y.; Zhang, J.; Zhang, Y.N.; Miao, J.X. Total flavonoids of Taraxacum mongolicum inhibit non-small cell lung cancer by regulating immune function. J. Ethnopharmacol. 2021, 281, 114514. [Google Scholar] [CrossRef]
- Wirngo, F.E.; Lambert, M.N.; Jeppesen, P.B. The Physiological Effects of Dandelion (Taraxacum officinale) in Type 2 Diabetes. Rev. Diabet. Stud. RDS 2016, 13, 113–131. [Google Scholar] [CrossRef]
- Aabideen, Z.U.; Mumtaz, M.W.; Akhtar, M.T.; Mukhtar, H.; Raza, S.A.; Touqeer, T.; Saari, N. Anti-Obesity Attributes; UHPLC-QTOF-MS/MS-Based Metabolite Profiling and Molecular Docking Insights of Taraxacum officinale. Molecules 2020, 25, 4935. [Google Scholar] [CrossRef]
- Hosseini Shekarabi, S.P.; Mostafavi, Z.S.; Mehrgan, M.S.; Islami, H.R. Dietary supplementation with dandelion (Taraxacum officinale) flower extract provides immunostimulation and resistance against Streptococcus iniae infection in rainbow trout (Oncorhynchusmykiss). Fish Shellfish Immunol. 2021, 118, 180–187. [Google Scholar] [CrossRef]
- Zou, C.Z.; Wang, H.J.; Li, J.; Liu, Z.; Li, Y.T.; Ma, H.X. Effect of Pugongying(Taraxacum mongolicum Hand.-Mazz.) on Acute Hyperuricemia Rats(in Chinese). Chin. Arch. Tradit. Chin. Med. 2020, 38, 170–172. [Google Scholar] [CrossRef]
- Zou, C.Z.; Wang, H.J.; Li, J.; Liu, Z.; Li, Y.T.; Ma, H.X. The Protection of T. Hand.-Mazz. on Renal Injury Rats with Persistent Hyperuricemia(in Chinese). Chin. J. Ethnomedicine Ethnopharmacy 2020, 29, 10–12. [Google Scholar]
- Li, L.; Wang, L.; Fan, W.; Jiang, Y.; Zhang, C.; Li, J.; Peng, W.; Wu, C. The Application of Fermentation Technology in Traditional Chinese Medicine: A Review. Am. J. Chin. Med. 2020, 48, 899–921. [Google Scholar] [CrossRef] [PubMed]
- Gadhoumi, H.; Hayouni, E.L.A.; Martinez-Rojas, E.; Yeddes, W.; Tounsi, M.S. Biochemical composition, antimicrobial and antifungal activities assessment of the fermented medicinal plants extract using lactic acid bacteria. Arch. Microbiol. 2022, 204, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Eweys, A.S.; Zhao, Y.-S.; Darwesh, O.M. Improving the antioxidant and anticancer potential of Cinnamomum cassia via fermentation with Lactobacillus plantarum. Biotechnol. Rep. (Amst. Neth.) 2022, 36, e00768. [Google Scholar] [CrossRef]
- Oh, N.S.; Lee, J.Y.; Lee, J.M.; Lee, K.W.; Kim, Y. Mulberry leaf extract fermented with Lactobacillus acidophilus A4 ameliorates 5-fluorouracil-induced intestinal mucositis in rats. Lett. Appl. Microbiol. 2017, 64, 459–468. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, D.; Shen, Y.; Chen, Y.; Qian, J.; Fu, G. Effect of Lactobacillus acidophilus fermentation on the composition of chlorogenic acids and anti-hyperuricemia activity of Artemisia selengensis Turcz. Food Funct. 2022, 13, 11780–11793. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, Y.; Wu, J.; Ou, K.; Huang, Q.; Cao, J.; Duan, T.; Zhou, L.; Pan, Y. Chemical profile and antioxidant activity of bidirectional metabolites from Tremella fuciformis and Acanthopanax trifoliatus as assessed using response surface methodology. Front. Nutr. 2022, 9, 1035788. [Google Scholar] [CrossRef]
- Deng, Y.; Huang, Q.; Hu, L.; Liu, T.; Zheng, B.; Lu, D.; Guo, C.; Zhou, L. Enhanced exopolysaccharide yield and antioxidant activities of Schizophyllum commune fermented products by the addition of Radix Puerariae. RSC Adv. 2021, 11, 38219–38234. [Google Scholar] [CrossRef]
- Deng, Y.; Liu, H.; Huang, Q.; Tu, L.; Hu, L.; Zheng, B.; Sun, H.; Lu, D.; Guo, C.; Zhou, L. Mechanism of Longevity Extension of Caenorhabditis elegans Induced by Schizophyllum commune Fermented Supernatant With Added Radix Puerariae. Front. Nutr. 2022, 9, 847064. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, G.; Liao, Y.; Pan, J. Inhibition of chrysin on xanthine oxidase activity and its inhibition mechanism. Int. J. Biol. Macromol. 2015, 81, 274–282. [Google Scholar] [CrossRef]
- Liu, Y.; Han, C.; Lu, T.; Liu, Y.; Chen, H.; Yang, C.; Tu, Y.; Li, Y. Investigation of the interaction between Chrysoeriol and xanthine oxidase using computational and in vitro approaches. Int. J. Biol. Macromol. 2021, 190, 463–473. [Google Scholar] [CrossRef]
- Wang, Z.N.; Feng, Y.Z.; Yang, N.N.; Jiang, T.; Xu, H.D.; Lei, H.J. Fermentation of kiwifruit juice from two cultivars by probiotic bacteria: Bioactive phenolics, antioxidant activities and flavor volatiles. Food Chem. 2022, 373, 131455. [Google Scholar] [CrossRef] [PubMed]
- Yin, N.; Wang, Y.; Ren, X.R.; Zhao, Y.; Liu, N.; An, X.P.; Qi, J.W. Isolation and Characterization of Flavonoids from Fermented Dandelion (Taraxacum mongolicum Hand.-Mazz.), and Assessment of Its Antioxidant Actions In Vitro and In Vivo. Fermentation 2022, 8, 306. [Google Scholar] [CrossRef]
- Xie, J.H.; Zhang, F.; Wang, Z.J.; Shen, M.Y.; Nie, S.P.; Xie, M.Y. Preparation, characterization and antioxidant activities of acetylated polysaccharides from Cyclocarya paliurus leaves. Carbohydr. Polym. 2015, 133, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Asghar, M.T.; Yusof, Y.A.; Mokhtar, M.N.; Ya’acob, M.E.; Mohd Ghazali, H.; Chang, L.S.; Manaf, Y.N. Coconut (Cocos nucifera L.) sap as a potential source of sugar: Antioxidant and nutritional properties. Food Sci. Nutr. 2020, 8, 1777–1787. [Google Scholar] [CrossRef]
- Chen, S.; Huang, H.; Huang, G. Extraction, derivatization and antioxidant activity of cucumber polysaccharide. Int. J. Biol. Macromol. 2019, 140, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jin, X.; Zhang, X.; Xie, X.; Tu, Z.; He, X. From Function to Metabolome: Metabolomic Analysis Reveals the Effect of Probiotic Fermentation on the Chemical Compositions and Biological Activities of Perilla frutescens Leaves. Front. Nutr. 2022, 9, 933193. [Google Scholar] [CrossRef] [PubMed]
- BM, A.L.; Elgebaly, H.A.; Germoush, M.O.; Qarmush, M.M.; Azab, M.S.; Alruhaimi, R.S.; Ahmeda, A.F.; Abukhalil, M.H.; Kamel, E.M.; Arab, H.H.; et al. A flavonoid-rich fraction of Monolluma quadrangula inhibits xanthine oxidase and ameliorates potassium oxonate-induced hyperuricemia in rats. Env. Sci. Pollut. Res. Int. 2022, 29, 63520–63532. [Google Scholar] [CrossRef]
- Huang, L.; He, X.; Peng, W.; He, X.; Xu, B.; Xu, H.; Wang, Y.; Xu, W.; Chen, W.; Wang, S.; et al. Hyperuricemia induces liver injury by upregulating HIF-1α and inhibiting arginine biosynthesis pathway in mouse liver and human L02 hepatocytes. Biochem. Biophys. Res. Commun. 2022, 617, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.Y.; Kim, D.S. Synergistic Impacts of Alpinia oxyphylla Seed Extract and Allopurinol against Experimental Hyperuricemia. Biomed Res. Int. 2022, 2022, 2824535. [Google Scholar] [CrossRef]
- Wang, M.; Lin, X.; Yang, X.; Yang, Y. Research progress on related mechanisms of uric acid activating NLRP3 inflammasome in chronic kidney disease. Ren. Fail. 2022, 44, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Ohno, I. Relationship between hyperuricemia and chronic kidney disease. Nucl. Nucl. Nucleic Acids 2011, 30, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Liu, Q.; Hao, H.; Bu, Y.; Tian, X.; Wang, T.; Yi, H. Lactobacillus paracasei X11 Ameliorates Hyperuricemia and Modulates Gut Microbiota in Mice. Front. Immunol. 2022, 13, 940228. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Sun, S.; Huang, Y.; Gao, Q.; Xie, X.; Wang, P.; Li, J.; Liang, L.; He, X.; Jiang, Y.; et al. Metagenomic analysis revealed the potential role of gut microbiome in gout. NPJ Biofilms Microbiomes 2021, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Werlinger, P.; Suh, J.W.; Cheng, J. Potential Probiotic Lacticaseibacillus paracasei MJM60396 Prevents Hyperuricemia in a Multiple Way by Absorbing Purine, Suppressing Xanthine Oxidase and Regulating Urate Excretion in Mice. Microorganisms 2022, 10, 851. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.Y.; Lu, Z.X.; Lu, Y.J. The potential of probiotics in the amelioration of hyperuricemia. Food Funct. 2022, 13, 2394–2414. [Google Scholar] [CrossRef]
- Xu, J.; Xu, J.; Shi, T.; Zhang, Y.; Chen, F.; Yang, C.; Guo, X.; Liu, G.; Shao, D.; Leong, K.W.; et al. Probiotic-inspired nanomedicine restores intestinal homeostasis in colitis by regulating redox balance, immune responses, and the gut microbiome. Adv. Mater. 2022, 35, e2207890. [Google Scholar] [CrossRef]
- Wu, Y.; Jha, R.; Li, A.; Liu, H.; Zhang, Z.; Zhang, C.; Zhai, Q.; Zhang, J. Probiotics (Lactobacillus plantarum HNU082) Supplementation Relieves Ulcerative Colitis by Affecting Intestinal Barrier Functions, Immunity-Related Gene Expression, Gut Microbiota, and Metabolic Pathways in Mice. Microbiol. Spectr. 2022, 10, e0165122. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, S.; Xu, L.; Fang, X.; Wan, Y.; Yu, D.; Guo, Y. A tetrapeptide from maize combined with probiotics exerted strong anti-inflammatory effects and modulated gut microbiota in DSS-induced colitis mice. Food Funct. 2022, 13, 12602–12618. [Google Scholar] [CrossRef]
- James, A.; Ke, H.M.; Yao, T.; Wang, Y.S. The Role of Probiotics in Purine Metabolism, Hyperuricemia and Gout: Mechanisms and Interventions. Food Rev. Int. 2023, 39, 261–277. [Google Scholar] [CrossRef]
- Hsieh, M.W.; Chen, H.Y.; Tsai, C.C. Screening and Evaluation of Purine-Nucleoside-Degrading Lactic Acid Bacteria Isolated from Winemaking Byproducts In Vitro and Their Uric Acid-Lowering Effects In Vivo. Fermentation 2021, 7, 74. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, C.; Zeng, X.; Yuan, Z. Microecological treatment of hyperuricemia using Lactobacillus from pickles. BMC Microbiol. 2020, 20, 195. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Cheng, F.; Li, W.; Yu, Q.; Ma, C.; Zou, Y.; Xu, T.; Liu, S.; Zhang, S.; Wang, Q. Enhancement of anti-inflammatory effect of cattle bile by fermentation and its inhibition of neuroinflammation on microglia by inhibiting NLRP3 inflammasome. J. Biosci. Bioeng. 2022, 133, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Su, Y.; An, Z.; Zhang, P.; Yue, Q.; Zhao, C.; Sun, X.; Zhang, S.; Liu, X.; Li, K.; et al. Fermentation products of Danshen relieved dextran sulfate sodium-induced experimental ulcerative colitis in mice. Sci. Rep. 2021, 11, 16210. [Google Scholar] [CrossRef]
- González-Castejón, M.; Visioli, F.; Rodriguez-Casado, A. Diverse biological activities of dandelion. Nutr. Rev. 2012, 70, 534–547. [Google Scholar] [CrossRef]
- Jhang, J.J.; Ong, J.W.; Lu, C.C.; Hsu, C.L.; Lin, J.H.; Liao, J.W.; Yen, G.C. Hypouricemic effects of Mesona procumbens Hemsl. through modulating xanthine oxidase activity in vitro and in vivo. Food Funct. 2016, 7, 4239–4246. [Google Scholar] [CrossRef]
- Gao, H.; Li, X.; Chen, X.; Hai, D.; Wei, C.; Zhang, L.; Li, P. The Functional Roles of Lactobacillus acidophilus in Different Physiological and Pathological Processes. J. Microbiol. Biotechnol. 2022, 32, 1226–1233. [Google Scholar] [CrossRef]
- Kim, H.; Nam, B.Y.; Park, J.; Song, S.; Kim, W.K.; Lee, K.; Nam, T.W.; Park, J.T.; Yoo, T.H.; Kang, S.W.; et al. Lactobacillus acidophilus KBL409 Reduces Kidney Fibrosis via Immune Modulatory Effects in Mice with Chronic Kidney Disease. Mol. Nutr. Food Res. 2022, 66, 2101105. [Google Scholar] [CrossRef]
- Luis-Rodriguez, D.; Donate-Correa, J.; Martin-Nunez, E.; Ferri, C.; Tagua, V.G.; Castro, A.P.; Mora-Fernandez, C.; Navarro-Gonzalez, J.F. Serum urate is related to subclinical inflammation in asymptomatic hyperuricaemia. Rheumatology 2021, 60, 371–379. [Google Scholar] [CrossRef]
- Kluck, V.; Liu, R.Q.; Joosten, L.A.B. The role of interleukin-1 family members in hyperuricemia and gout. Jt. Bone Spine 2021, 88, 105092. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Nie, Y.; Chang, Y.; Zeng, S.; Liang, C.; Zheng, X.; Xiao, D.; Zhan, S.; Zheng, Q. Protective effects of Rhizoma smilacis glabrae extracts on potassium oxonate- and monosodium urate-induced hyperuricemia and gout in mice. Phytomedicine 2019, 59, 152772. [Google Scholar] [CrossRef]
- Enomoto, A.; Kimura, H.; Chairoungdua, A.; Shigeta, Y.; Jutabha, P.; Cha, S.H.; Hosoyamada, M.; Takeda, M.; Sekine, T.; Igarashi, T.; et al. Molecular identification of a renal urate-anion exchanger that regulates blood urate levels. Nature 2002, 417, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Kim, G.-H. Urate Transporters in the Kidney: What Clinicians Need to Know. Electrolyte Blood Press. 2021, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.X.; Li, C.H.; Zhou, P.; Jiang, T.L. Uric acid transporters hiding in the intestine. Pharm. Biol. 2016, 54, 3151–3155. [Google Scholar] [CrossRef] [PubMed]
- Marchiando, A.M.; Shen, L.; Graham, W.V.; Edelblum, K.L.; Duckworth, C.A.; Guan, Y.F.; Montrose, M.H.; Turner, J.R.; Watson, A.J.M. The Epithelial Barrier Is Maintained by In Vivo Tight Junction Expansion During Pathologic Intestinal Epithelial Shedding. Gastroenterology 2011, 140, 1208–1218. [Google Scholar] [CrossRef]
- Eckenstaler, R.; Benndorf, R.A. The Role of ABCG2 in the Pathogenesis of Primary Hyperuricemia and Gout-An Update. Int. J. Mol. Sci. 2021, 22, 6678. [Google Scholar] [CrossRef]
- Liu, T.; Gao, H.; Zhang, Y.; Wang, S.; Lu, M.; Dai, X.; Liu, Y.; Shi, H.; Xu, T.; Yin, J.; et al. Apigenin Ameliorates Hyperuricemia and Renal Injury through Regulation of Uric Acid Metabolism and JAK2/STAT3 Signaling Pathway. Pharmaceuticals 2022, 15, 1442. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, Z.; Zhang, B.; Nie, A.; Bian, M. Cichorium intybus L. promotes intestinal uric acid excretion by modulating ABCG2 in experimental hyperuricemia. Nutr. Metab. 2017, 14, 38. [Google Scholar] [CrossRef]
- Gao, Y.; Sun, J.; Zhang, Y.; Shao, T.J.; Li, H.C.; Wang, M.J.; Zhang, L.; Bian, H.; Wen, C.P.; Xie, Z.J.; et al. Effect of a Traditional Chinese Medicine Formula (CoTOL) on Serum Uric Acid and Intestinal Flora in Obese Hyperuricemic Mice Inoculated with Intestinal Bacteria. Evid.-Based Complement. Altern. Med. 2020, 2020, 8831937. [Google Scholar] [CrossRef]
- Sommer, F.; Anderson, J.M.; Bharti, R.; Raes, J.; Rosenstiel, P. The resilience of the intestinal microbiota influences health and disease. Nat. Rev. Microbiol. 2017, 15, 630–638. [Google Scholar] [CrossRef]
- Wang, Z.L.; Li, Y.C.; Liao, W.H.; Huang, J.; Liu, Y.P.; Li, Z.Y.; Tang, J.Y. Gut microbiota remodeling: A promising therapeutic strategy to confront hyperuricemia and gout. Front. Cell. Infect. Microbiol. 2022, 12, 935723. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, J.; Wang, Z.; Ang, K.Y.; Huang, S.; Hou, Q.; Su, X.; Qiao, J.; Zheng, Y.; Wang, L.; et al. Intestinal Microbiota Distinguish Gout Patients from Healthy Humans. Sci. Rep. 2016, 6, 20602. [Google Scholar] [CrossRef]
- Haller, D.; Hoermannsperger, G. Interaction between humans and intestinal bacteria as a determinant for intestinal health. Intestinal microbiome and inflammatory bowel diseases. Bundesgesundheitsblatt-Gesundheitsforschung-Gesundheitsschutz 2015, 58, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Martinez, I.; Walter, J.; Keshavarzian, A.; Rose, D.J. In vitro characterization of the impact of selected dietary fibers on fecal microbiota composition and short chain fatty acid production. Anaerobe 2013, 23, 74–81. [Google Scholar] [CrossRef]
- Marzocco, S.; Fazeli, G.; Di Micco, L.; Autore, G.; Adesso, S.; Dal Piaz, F.; Heidland, A.; Di Iorio, B. Supplementation of Short-Chain Fatty Acid, Sodium Propionate, in Patients on Maintenance Hemodialysis: Beneficial Effects on Inflammatory Parameters and Gut-Derived Uremic Toxins, A Pilot Study (PLAN Study). J. Clin. Med. 2018, 7, 315. [Google Scholar] [CrossRef] [PubMed]
- Fei, N.; Zhao, L. An opportunistic pathogen isolated from the gut of an obese human causes obesity in germfree mice. ISME J. 2013, 7, 880–884. [Google Scholar] [CrossRef]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Jiang, Q.; Yin, Y. Butyrate in Energy Metabolism: There Is Still More to Learn. Trends Endocrinol. Metab. TEM 2021, 32, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Han, P.; Ma, S.; Peng, R.; Wang, C.; Kong, W.; Cong, L.; Fu, J.; Zhang, Z.; Yu, H.; et al. Abnormal metabolism of gut microbiota reveals the possible molecular mechanism of nephropathy induced by hyperuricemia. Acta Pharm. Sinica B 2020, 10, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).