
Exploration and Biorefinery Antimicrobial Agent through Solid State Fermentation from Indonesia’s Marine Actinomycetes


 ,
,  ,
,  , ,
, , 
 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Biomaterial and Maintenance
2.2. Chitinolytic Assay and Analysis of Degradation Products
2.3. Screening of Antimicrobial Assay
2.4. Morphological Identification of Actinomycetes Isolate
2.5. Phylogenetic Analysis
2.6. Cultivation and Extraction
3. Results and Discussion
3.1. Isolation of Actinomycetes from Bali and Gorontalo Marine Biota
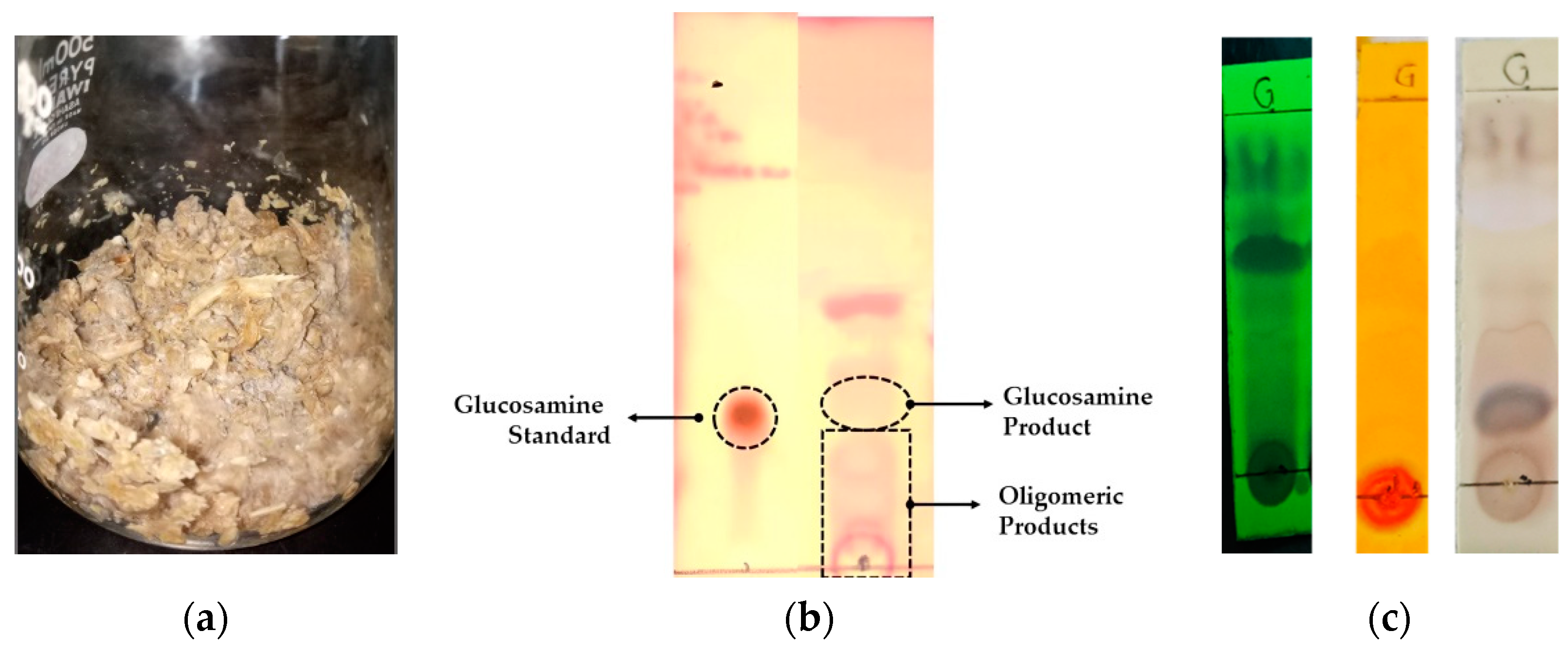
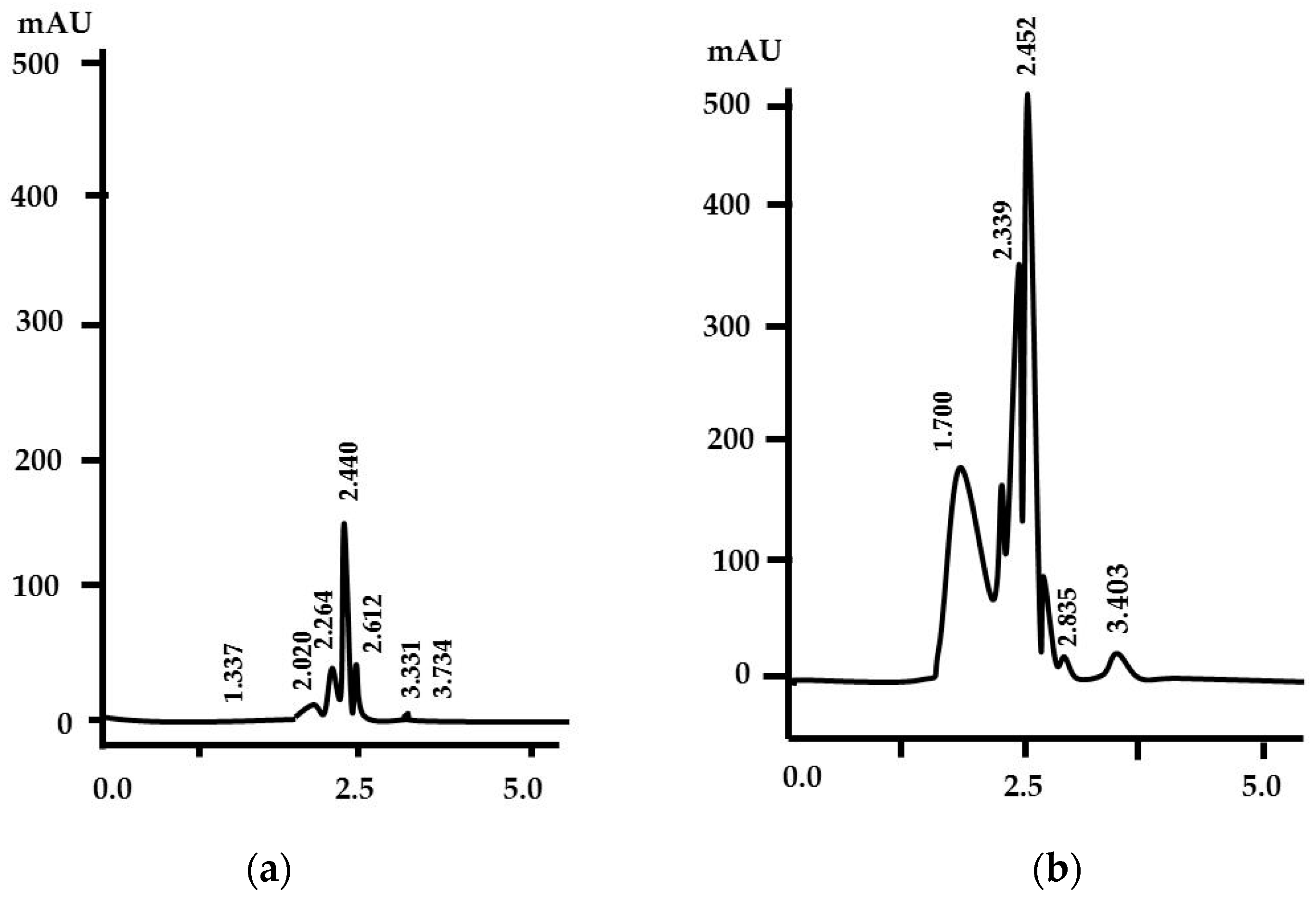
3.2. Chitinolytic Assay and Analysis of Degradation Products
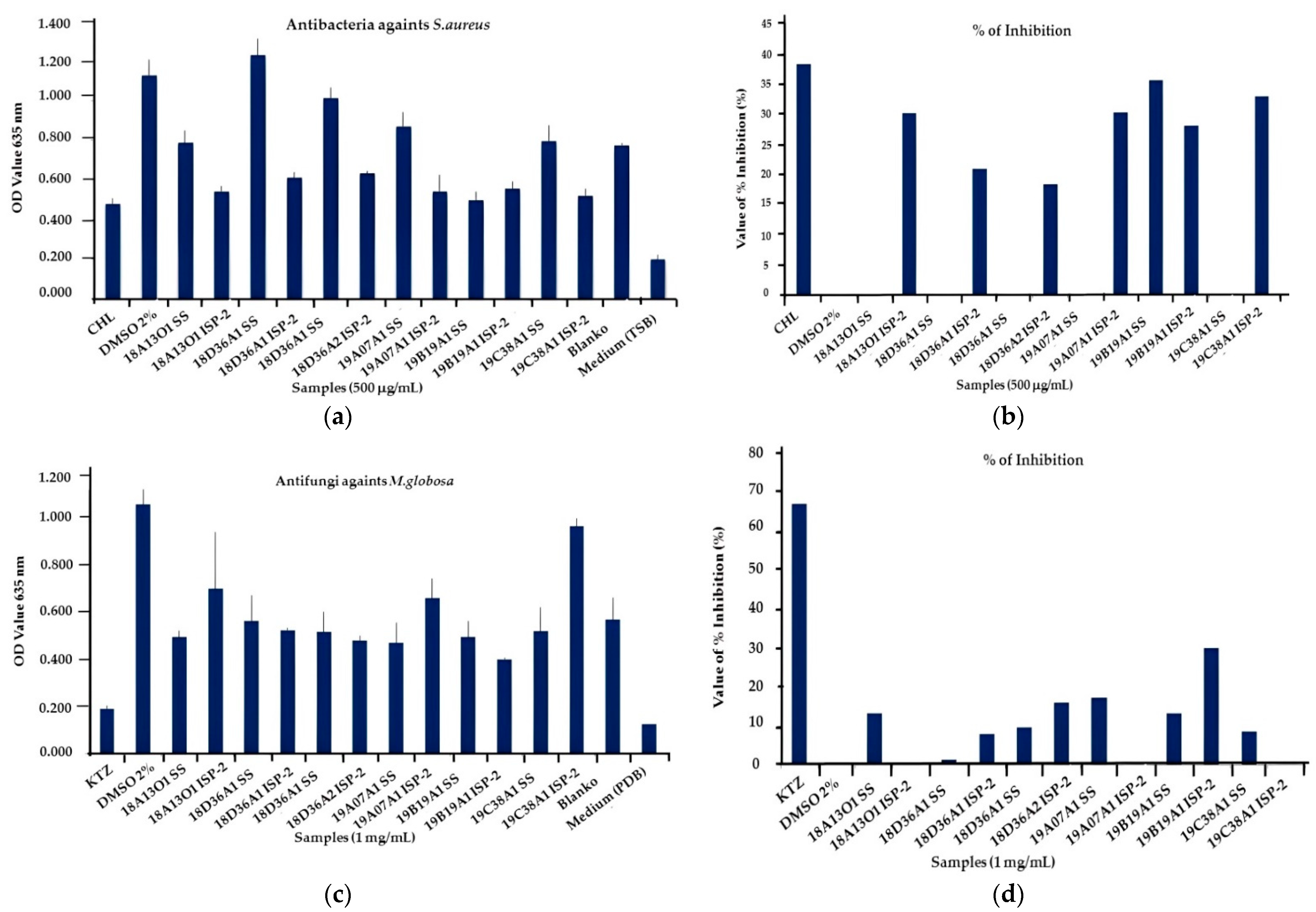
3.3. Screening of Antimicrobial Assay
3.4. Scale up Solid State Fermentation and Extraction
3.5. Morphological Identification of Actinomycetes
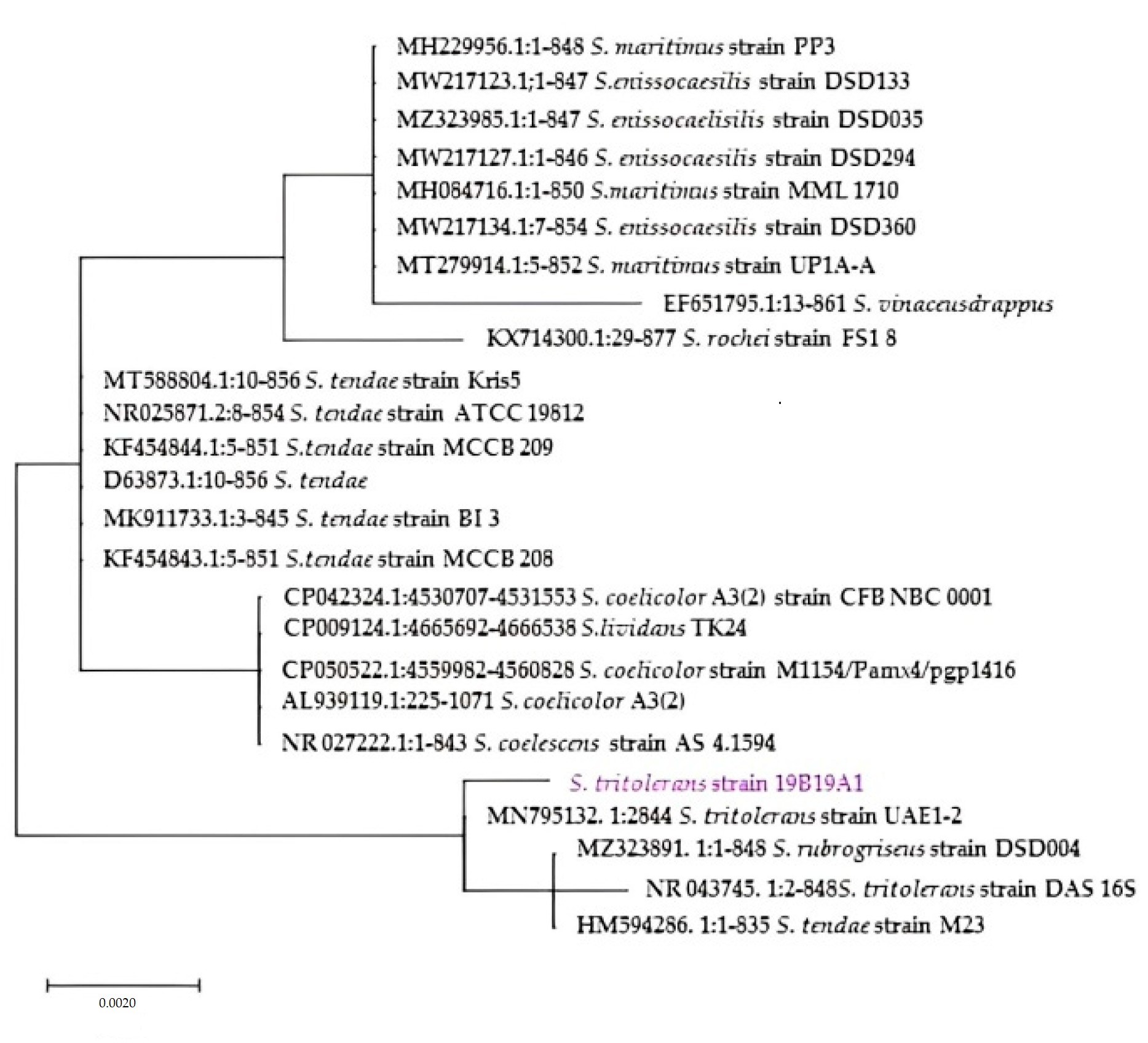
3.6. Phylogenetical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spernovasilis, N.; Tsiodras, S.; Poulakou, G. Emerging and Re-Emerging Infectious Diseases: Humankind’s Companions and Competitors. Microorganisms 2022, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Von Lilienfeld-Toal, M.; Wagener, J.; Einsele, H.; Cornely, O.A.; Kurzai, O. Invasive Fungal Infection: New Treatments To Meet New Challenges. Dtsch. Ärzteblatt Int. 2019, 116, 271. [Google Scholar]
- Rathore, S.S.; Sathiyamoorthy, J.; Lalitha, C.; Ramakrishnan, J. A holistic review on Cryptococcus neoformans. Microb. Pathog. 2022, 166, 105521. [Google Scholar] [CrossRef]
- Huang, C.Y.; Peng, C.C.; Hsu, C.H.; Chang, J.H.; Chiu, N.C.; Chi, H. Systemic Infection Caused by Malassezia pachydermatis In Infants: Case Series and Review of The Literature. Pediatr. Infect. Dis. J. 2020, 39, 444–448. [Google Scholar] [CrossRef]
- Mularoni, A.; Graziano, E.; Todaro, F. Invasive Trichophyton Infection In A Liver Transplant Recipient. Transpl. Infect. Dis. 2022, 24, e13794. [Google Scholar] [CrossRef]
- Gong, J.; Hou, X.; Geng, Y.; Dai, R.C.; Zhao, F.; He, L. Rapid Identification of Four Fusarium spp. Complex by High-Resolution Melting Curve Analysis and Their Antifungal Susceptibility Profiles. Mycopathologia 2022, 187, 345–354. [Google Scholar]
- Kawabe, Y.; Sakurai, A.; Sasaki, T.; Hasegawa, M.; Suzuki, M.; Hoshino, N.; Doi, Y. Native Valve Infective Endocarditis Due To Sequence Type 97 Community-Associated Methicillin-Resistant Staphylococcus aureus Complicated by Meningitis and Multiple Septic Emboli In A Young Healthy Adult. J. Infect. Chemother. 2022, 28, 828–832. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- Roncarati, D.; Scarlato, V.; Vannini, A. Targeting of Regulators as a Promising Approach in the Search for Novel Antimicrobial Agents. Microorganisms 2022, 10, 185. [Google Scholar] [CrossRef]
- De Simeis, D.; Serra, S. Actinomycetes: A Never-Ending Source of Bioactive Compounds—An Overview on Antibiotics Production. Antibiotics 2021, 10, 483. [Google Scholar] [CrossRef] [PubMed]
- Bérdy, J. Thoughts and facts about antibiotics: Where we are now and where we are heading. J. Antibiot. 2012, 65, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Do Khac, A.L. The Political Sea: Conservation Policies, State Power, and Symbolic Violence: The Case of the Bajau in the Wakatobi Marine National Park. Explorations 2018, 14, 37–50. [Google Scholar]
- Hu, X.; Tang, Y.; Liu, Y.; Pei, X.; Huang, Z.; Song, F.; Zhang, H. Comprehensive Genomic Analysis of Marine Strain Streptomyces sp. 891, an Excellent Producer of Chrysomycin A with Therapeutic Potential. Mar. Drugs 2022, 20, 287. [Google Scholar] [CrossRef] [PubMed]
- Yi, W.; Newaz, A.W.; Yong, K.; Ma, M.; Lian, X.-Y.; Zhang, Z. New Hygrocins K–U and Streptophenylpropanamide A and Bioactive Compounds from the Marine-Associated Streptomyces sp. ZZ1956. Antibiotics 2022, 11, 1455. [Google Scholar] [CrossRef]
- Donald, L.; Pipite, A.; Subramani, R.; Owen, J.; Keyzers, R.A.; Taufa, T. Streptomyces: Still the biggest producer of new natural secondary metabolites, a current perspective. Microbiol. Res. 2022, 13, 418–465. [Google Scholar] [CrossRef]
- Setiawan, A.; Widyastuti, W.; Irawan, A.; Wijaya, O.S.; Laila, A.; Setiawan, W.A.; Juliasih, N.L.G.R.; Nonaka, K.; Arai, M.; Hendri, J. Solid State Fermentation of Shrimp Shell Waste Using Pseudonocardia carboxydivorans 18A13O1 to Produce Bioactive Metabolites. Fermentation 2021, 7, 247. [Google Scholar] [CrossRef]
- Raju, K.J.; Rittapalli, T. Production and Optimization Studies of Protease by Soil Isolate Actinomycetes Using Mixed Substrate of Pomegranate Peel Powder and Banana Peel Powder under Solid State Fermentation. Asian J. Biotechnol. Bioresour. Technol. 2022, 8, 39–47. [Google Scholar]
- Wu, D.; Wei, Z.; Mohamed, T.A.; Zheng, G.; Qu, F.; Wang, F.; Zhao, Y.; Song, C. Lignocellulose biomass bioconversion during composting: Mechanism of action of lignocellulase, pretreatment methods and future perspectives. Chemosphere 2022, 286, 131635. [Google Scholar] [CrossRef]
- Swiontek Brzezinska, M.; Jankiewicz, U.; Kalwasińska, A.; Świątczak, J.; Żero, K. Characterization of chitinase from Streptomyces luridiscabiei U05 and its antagonist potential against fungal plant pathogens. J. Phytopathol. 2019, 167, 404–412. [Google Scholar] [CrossRef]
- Alzahrani, O.M.; Sohail, M.; Mahmoud, S.F.; Alswat, A.S.; El-Halmouch, Y. Preparation of a Pectinase-Enriched Multienzyme under Solid State Fermentation of Sugarcane Bagasse. Fermentation 2023, 9, 141. [Google Scholar] [CrossRef]
- Vicente, F.A.; Ventura, S.P.; Passos, H.; Dias, A.C.; Torres-Acosta, M.A.; Novak, U.; Likozar, B. Crustacean waste biorefinery as a sustainable cost-effective business model. Chem. Eng. J. 2022, 442, 135937. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquacultures: Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Lu, D.L.; Zhang, M.S.; Deng, J.J.; Lu, W.J.; Yang, Z.D.; Li, Z.W.; Chen, L.-C.; Luo, X.C. Highly Efficient Shrimp Shell Recovery by Solid-State Fermentation with Streptomyces sp. SCUT-3. Chem. Eng. J. 2023, 458, 141256. [Google Scholar] [CrossRef]
- Ofori, S.A.; Kodikara, S.K.; Jayatissa, L.P.; Madarasinghe, S.K.; Nijamdeen, T.M.; Dahdouh-Guebas, F. What is the ecological footprint of aquaculture after 5 decades of competition between mangrove conservation and shrimp farm development? Aquat. Conserv. Mar. Freshw. Ecosyst. 2023, 33, 15–28. [Google Scholar] [CrossRef]
- Ruslan, N.F.; Ahmad, N.; Abas, A.; Sanfilippo, A.; Mahmoud, K.; Munaim, M.S.A.; Nour, A.H. Sustainable Bioethanol Production by Solid State Fermentation: A Systematic Review. Res. Sq. 2023; preprint. [Google Scholar]
- Yan, N.; Xi, C. Sustainability: Don’t waste seafood waste. Nature 2015, 524, 155–157. [Google Scholar] [CrossRef]
- Tan, Y.N.; Lee, P.P.; Chen, W.N. Microbial extraction of chitin from seafood waste using sugars derived from fruit waste-stream. AMB Expr. 2020, 10, 17. [Google Scholar] [CrossRef]
- Suryawanshi, N.; Eswari, J.S. Chitin from seafood waste: Particle swarm optimization and neural network study for the improved chitinase production. J. Chem. Technol. Biotechnol. 2022, 97, 509–519. [Google Scholar] [CrossRef]
- Agrawal, T.; Kotasthane, A.S. Chitinolytic Assay of Indigenous Trichoderma Isolates Collected from Different Geographical Locations of Chhattisgarh in Central India. SpringerPlus 2012, 1, 73. [Google Scholar] [CrossRef]
- Zhu, X.F.; Zhou, Y.; Feng, J.L. Analysis of both chitinase and chitosanase produced by Sphingomonas sp. CJ-5. J. Zhejiang Univ. Sci. B 2007, 8, 831–838. [Google Scholar] [CrossRef]
- Zbîrcea, R.I.; Menghiu, G.; Matica, A.; Ostafe, V. Use of 3, 5-dinitrosalicylic Acid Reaction To Study The Chitosan Hydrolysis. New Front. Chem. 2016, 25, 145–153. [Google Scholar]
- Setiawan, A.; Lutfiah, R.; Juliasih, N.L.G.R.; Setiawan, W.A.; Hendri, J.; Arai, M. Antibacterial activity of EtOAc extract from marine-derived fungus Aspergillus nomiae A12-RF against clinical pathogen bacteria, Staphylococcus aureus. AACL Bioflux 2022, 15, 1413–1421. [Google Scholar]
- Leong, C.; Buttafuoco, A.; Glatz, M.; Bosshard, P.P. Antifungal Susceptibility Testing of Malassezia spp. With an Optimized Colorimetric Broth Microdilution Method. J. Clin. Microbiol. 2017, 55, 1883–1893. [Google Scholar] [CrossRef] [PubMed]
- Widyastuti, W.; Setiawan, F.; Al Afandy, C.; Irawan, A.; Laila, A.; Juliasih, N.L.G.R.; Setiawan, W.A.; Arai, M.; Hendri, J.; Setiawan, A. Antifungal Agent Chitooligosaccharides Derived from Solid-State Fermentation of Shrimp Shell Waste by Pseudonocardia antitumoralis 18D36-A1. Fermentation 2022, 8, 353. [Google Scholar] [CrossRef]
- Heuer, H.; Krsek, M.; Baker, P.; Smalla, K.; Wellington, E.M.H. Analysis of Actinomycete Communities by Specific Amplification of Genes Encoding 16S rRNA and Gel-Electrophoretic Separation In Denaturing Gradients. Appl. Environ. Microbiol. 1997, 63, 3233–3241. [Google Scholar] [CrossRef]
- Baskaran, R.; Mohan, P.M.; Sivakumar, K.; Kumar, A. Antimicrobial Activity and Phylogenetic Analysis of Streptomyces Parvulus Dosmb-D105 Isolated from the Mangrove Sediments of Andaman Islands. Acta Microbiol. Immunol. Hung. 2016, 63, 27–46. [Google Scholar] [CrossRef] [PubMed]
- Setyohadi, D.; Zakiyah, U.; Sambah, A.B.; Wijaya, A. Upwelling impact on sardinella lemuru during the indian ocean dipole in the bali strait, Indonesia. Fishes 2021, 6, 8. [Google Scholar] [CrossRef]
- Djunarsjah, E.; Julian, M.M.; Muhammad, F.; Putra, A.P.; Lubis, N.S.; Welly, T.K.; Wahab, W.A.; Pamungkas, B. Utilization of Marine Geospatial Data for Determining Optimal FAD Locations in Improving the Living Standards of the North Gorontalo Coastal Community. Sustainability 2023, 15, 2242. [Google Scholar] [CrossRef]
- Edlund, A.; Loesgen, S.; Fenical, W.; Jensen, P.R. Geographic distribution of secondary metabolite genes in the marine actinomycete Salinispora arenicola. Appl. Environ. Microbiol. 2011, 77, 5916–5925. [Google Scholar] [CrossRef]
- Ekundayo, F.O.; Folorunsho, A.E.; Ibisanmi, T.A.; Olabanji, O.B. Antifungal activity of chitinase produced by Streptomyces species isolated from grassland soils in Futa Area, Akure. Bull. Natl. Res. Cent. 2022, 46, 95. [Google Scholar] [CrossRef]
- Buchanan, R.E.; Gibbons, N.E. (Eds.) Bergey’s Manual of Determinative Bacteriology, 8th ed.; Williams & Wilkins Co.: Baltimore, MD, USA, 1974; pp. xxvi + 1246. [Google Scholar]
- Jahn, M.T. Physiology, Syntrophy and Viral Interplay in the Marine Sponge Holobiont. Ph.D. Dissertation, Christian-Albrechts-Universität Kiel, Kiel, Germany, 2019. [Google Scholar]
- Paulsen, S.S.; Andersen, B.; Gram, L.; Machado, H. Biological Potential of Chitinolytic Marine Bacteria. Mar. Drugs 2016, 14, 230. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Kimoto, H. Bacterial chitinase system as a model of chitin biodegradation. Target. Chitin-Contain. Org. 2019, 26, 131–151. [Google Scholar]
- Wang, S.; Fang, X.; Liang, K.; Li, S.; Han, S.; Zhu, T. Cloning, expression and antifungal effect of the recombinant chitinase from Streptomyces sampsonii KJ40. Ciência Rural 2023, 53, e20210663. [Google Scholar] [CrossRef]
- Craig, M.; Lambert, S.; Jourdan, S.; Tenconi, E.; Colson, S.; Maciejewska, M.; Ongena, M.; Martin, J.F.; van Wezel, G.; Rigali, S. Unsuspected control of siderophore production by N-acetylglucosamine in streptomycetes. Environ. Microbiol. Rep. 2012, 4, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Van der Meij, A.; Worsley, S.F.; Hutchings, M.I.; van Wezel, G.P. Chemical ecology of antibiotic production by actinomycetes. FEMS Microbiol. Rev. 2017, 41, 392–416. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Gözaydın, G.; Yang, H.; Ning, W.; Han, X.; Poon, N.Y.; Liang, H.; Ning, Y.; Zhou, K. Upcycling Chitin-Containing Waste Into Organonitrogen Chemicals Via An Integrated Process. Proc. Natl. Acad. Sci. USA 2020, 117, 7719–7728. [Google Scholar] [CrossRef]
- De Rop, A.S.; Rombaut, J.; Willems, T.; De Graeve, M.; Vanhaecke, L.; Hulpiau, P.; De Maeseneire, S.L.; De Mol, M.L.; Soetaert, W.K. Novel Alkaloids From Marine Actinobacteria: Discovery and Characterization. Mar. Drugs 2022, 20, 6. [Google Scholar] [CrossRef]
- Manucharova, N.A.; Trosheva, E.V.; Kol’tsova, E.M.; Demkina, E.V.; Karaevskaya, E.V.; Rivkina, E.M.; Mardanov, A.V.; El’-Registan, G.I. Characterization of the structure of the prokaryotic complex of antarctic permafrost by molecular genetic techniques. Mikrobiologiya 2016, 85, 83–91. [Google Scholar] [CrossRef]
- Umar, A.A.; Hussaini, A.B.; Yahayya, J.; Sani, I.; Aminu, H. Chitinolytic and Antagonistic Activity of Streptomyces Isolated from Fadama Soil against Phytopathogenic Fungi. Trop. Life Sci. Res. 2021, 32, 25–38. [Google Scholar] [CrossRef]
- Fan, Z.; Wang, L.; Qin, Y.; Li, P. Activity of chitin/chitosan/chitosan oligosaccharide against plant pathogenic nematodes and potential modes of application in agriculture: A review. Carbohydr. Polym. 2023, 306, 120592. [Google Scholar] [CrossRef]
- Xu, T.; Qi, M.; Liu, H.; Cao, D.; Xu, C.; Wang, L.; Qi, B. Chitin degradation potential and whole-genome sequence of Streptomyces diastaticus strain CS1801. AMB Express 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Setiawan, A.; Setiawan, F.; Juliasih, N.L.G.R.; Widyastuti, W.; Laila, A.; Setiawan, W.A.; Djailani, F.M.; Mulyono, M.; Hendri, J.; Arai, M. Fungicide Activity of Culture Extract from Kocuria palustris 19C38A1 against Fusarium oxysporum. J. Fungi 2022, 8, 280. [Google Scholar] [CrossRef] [PubMed]
- Lacey, H.J.; Rutledge, P.J. Recently Discovered Secondary Metabolites from Streptomyces Species. Molecules 2022, 27, 887. [Google Scholar] [CrossRef]
- Moran, M.A.; Rutherford, L.T.; Hodson, R.E. Evidence for indegenous Streptomyces population in a marine environment determined with a 16S rRNA probe. Appl. Environ. Microbiol. 1995, 61, 3695–3700. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, K.; Gupta, R.K. Diversity and isolation of rare actinomycetes: An overview. Crit. Rev. Microbiol. 2013, 39, 256–294. [Google Scholar] [CrossRef] [PubMed]
- Siro, G.; Pipite, A.; Christi, K.; Srinivasan, S.; Subramani, R. Marine Actinomycetes Associated with Stony Corals: A Potential Hotspot for Specialized Metabolites. Microorganisms 2022, 10, 1349. [Google Scholar] [CrossRef] [PubMed]
- Kathiresan, K.; Nabeel, M.A.; Manivannan, S. Bioprospecting of marine organisms for novel bioactive compounds. Sci. Trans. Environ. Technovation 2008, 1, 107–120. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Code | Antibacteria (0.5 mg/mL) | Antifungi (1 mg/mL) | ||
|---|---|---|---|---|
| ISP-2 | Shrimp Shell | ISP-2 | Shrimp Shell | |
| 18A13O1 | + | + | + | - |
| 18D36A1 | + | - | + | - |
| 18D36A2 | + | - | + | + |
| 19A07A1 | + | - | + | - |
| 19B19A1 | + | + | + | + |
| 19C38A1 | + | + | + | - |
| Analysis of 16S rRNA | Product | Potential Use |
|---|---|---|
| Pseudonocardia carboxydivorans 18A13O1 | Branymicin B | Antibacteria |
| Pseudonocardia antitumoralis 18D36A1 | Chitinase | Antifungi |
| Micrococcus sp 18D36A2 | Chitinase | Chitinase producing |
| Brevibacterium linens 19A07A1 | Alkaloid | Antibiofilm |
| Streptomyces tritolerans 19B19A1 | Alkaloid | Antibacteria, Antifungi |
| Kocuria palustris 19C38A1 | Benzimidazole | Antifungi |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laila, A.; Setiawan, F.; Widyastuti, W.; Fadhilah, M.R.; Setiawan, A.; Juliasih, N.L.G.R.; Setiawan, W.A.; Apriliana, E.; Ahmadi, P.; Arai, M.; et al. Exploration and Biorefinery Antimicrobial Agent through Solid State Fermentation from Indonesia’s Marine Actinomycetes. Fermentation 2023, 9, 334. https://doi.org/10.3390/fermentation9040334
Laila A, Setiawan F, Widyastuti W, Fadhilah MR, Setiawan A, Juliasih NLGR, Setiawan WA, Apriliana E, Ahmadi P, Arai M, et al. Exploration and Biorefinery Antimicrobial Agent through Solid State Fermentation from Indonesia’s Marine Actinomycetes. Fermentation. 2023; 9(4):334. https://doi.org/10.3390/fermentation9040334
Chicago/Turabian StyleLaila, Aspita, Fendi Setiawan, Widyastuti Widyastuti, Muhammad Rizky Fadhilah, Andi Setiawan, Ni Luh Gede Ratna Juliasih, Wawan Abdullah Setiawan, Ety Apriliana, Peni Ahmadi, Masayoshi Arai, and et al. 2023. "Exploration and Biorefinery Antimicrobial Agent through Solid State Fermentation from Indonesia’s Marine Actinomycetes" Fermentation 9, no. 4: 334. https://doi.org/10.3390/fermentation9040334
APA StyleLaila, A., Setiawan, F., Widyastuti, W., Fadhilah, M. R., Setiawan, A., Juliasih, N. L. G. R., Setiawan, W. A., Apriliana, E., Ahmadi, P., Arai, M., & Hendri, J. (2023). Exploration and Biorefinery Antimicrobial Agent through Solid State Fermentation from Indonesia’s Marine Actinomycetes. Fermentation, 9(4), 334. https://doi.org/10.3390/fermentation9040334