
Roles of Process Parameters on the Ricinoleic Acid Production from Castor Oil by Aspergillus flavus BU22S



Abstract
1. Introduction
2. Materials and Methods
2.1. Molecular Biological Identification of the Isolated Species and Its Growth Conditions
2.2. Biotransformation
2.3. Optimization of Process Variables by Response Surface Methodology (RSM)
2.4. Bench Scale Fermentation in the Optimized Conditions
2.5. Sample Analyses
2.5.1. Recovery of the Oil Phase
2.5.2. HPLC Analyses
2.5.3. Statistical Analyses
2.5.4. GenBank Accession Numbers
3. Results and Discussion
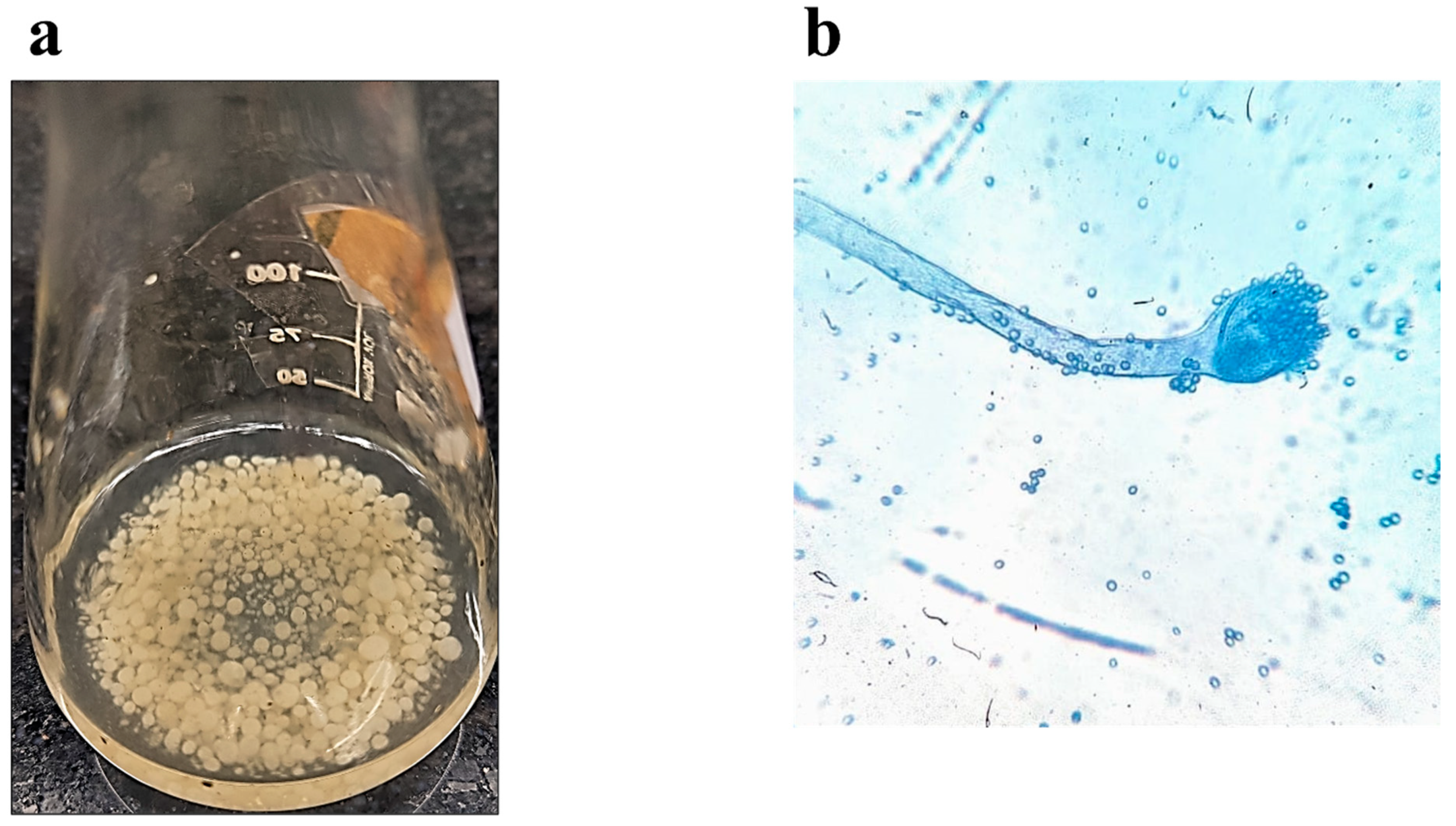
3.1. Molecular Biological Identification of the Isolated Species
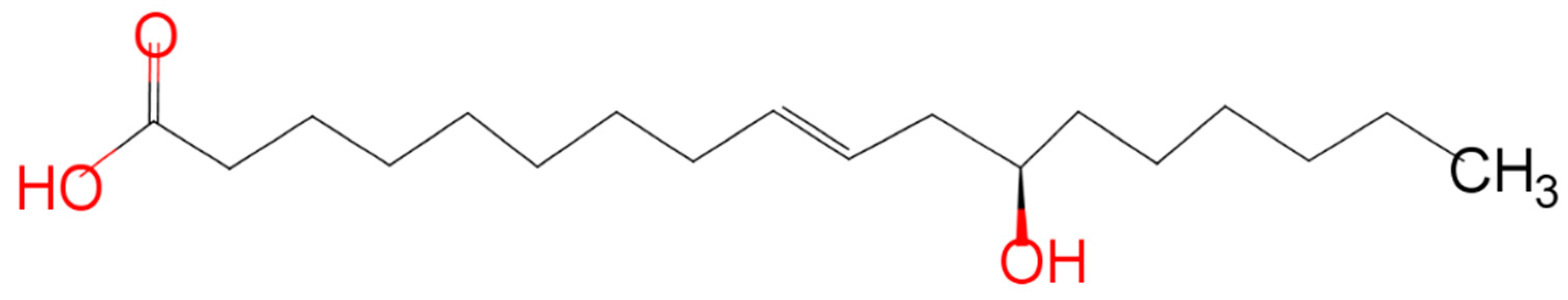
3.2. Biotransformation
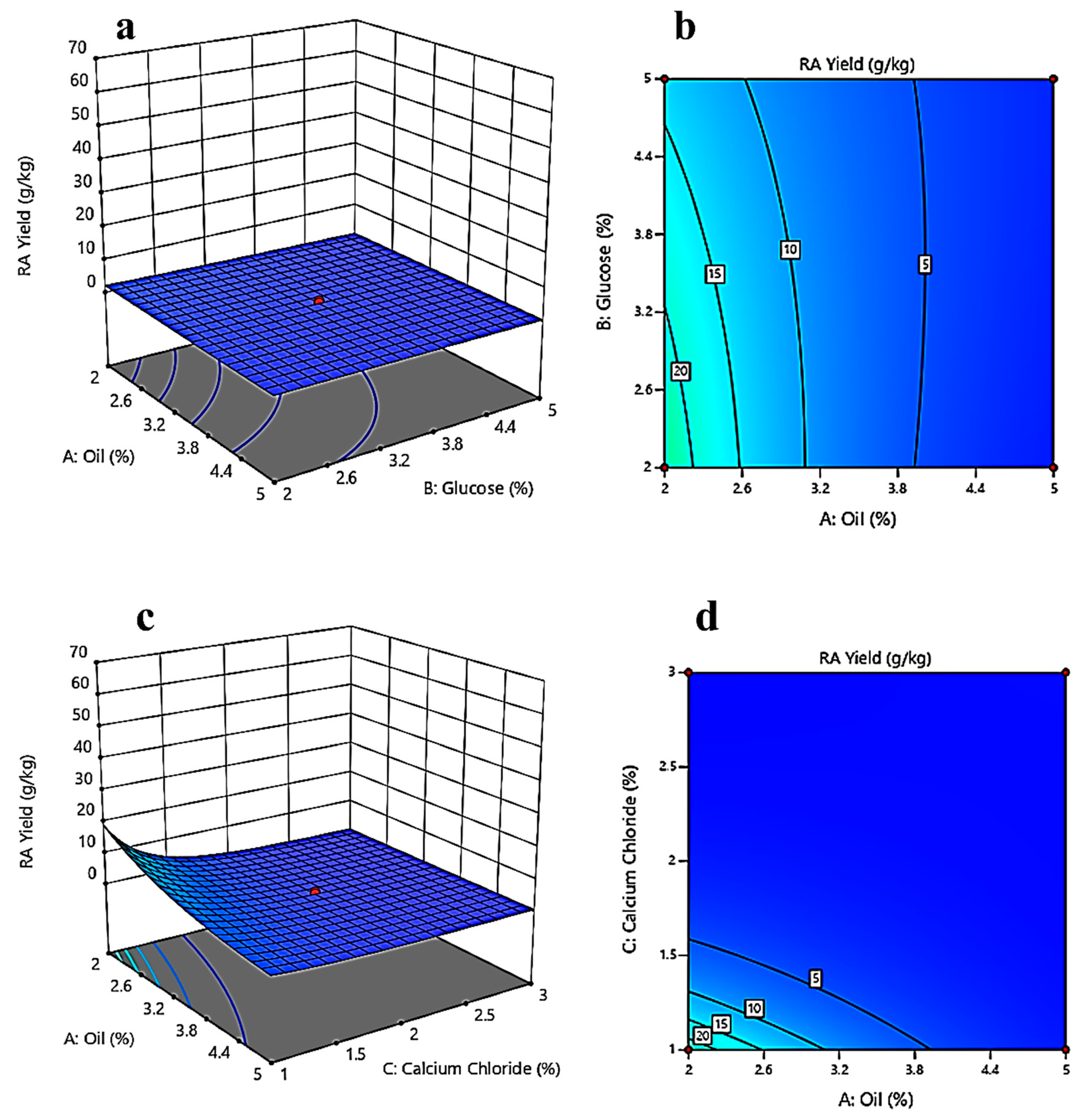
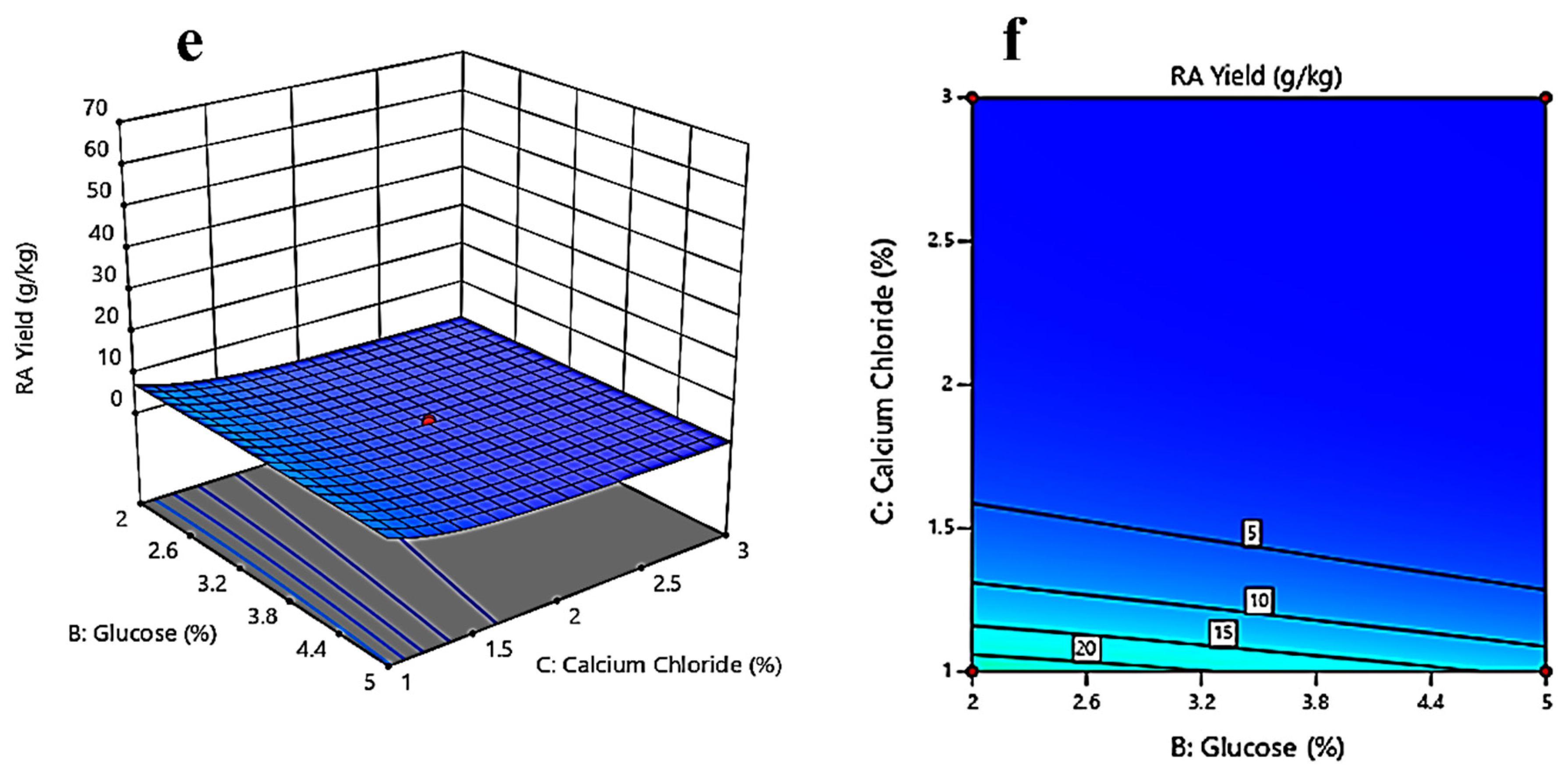
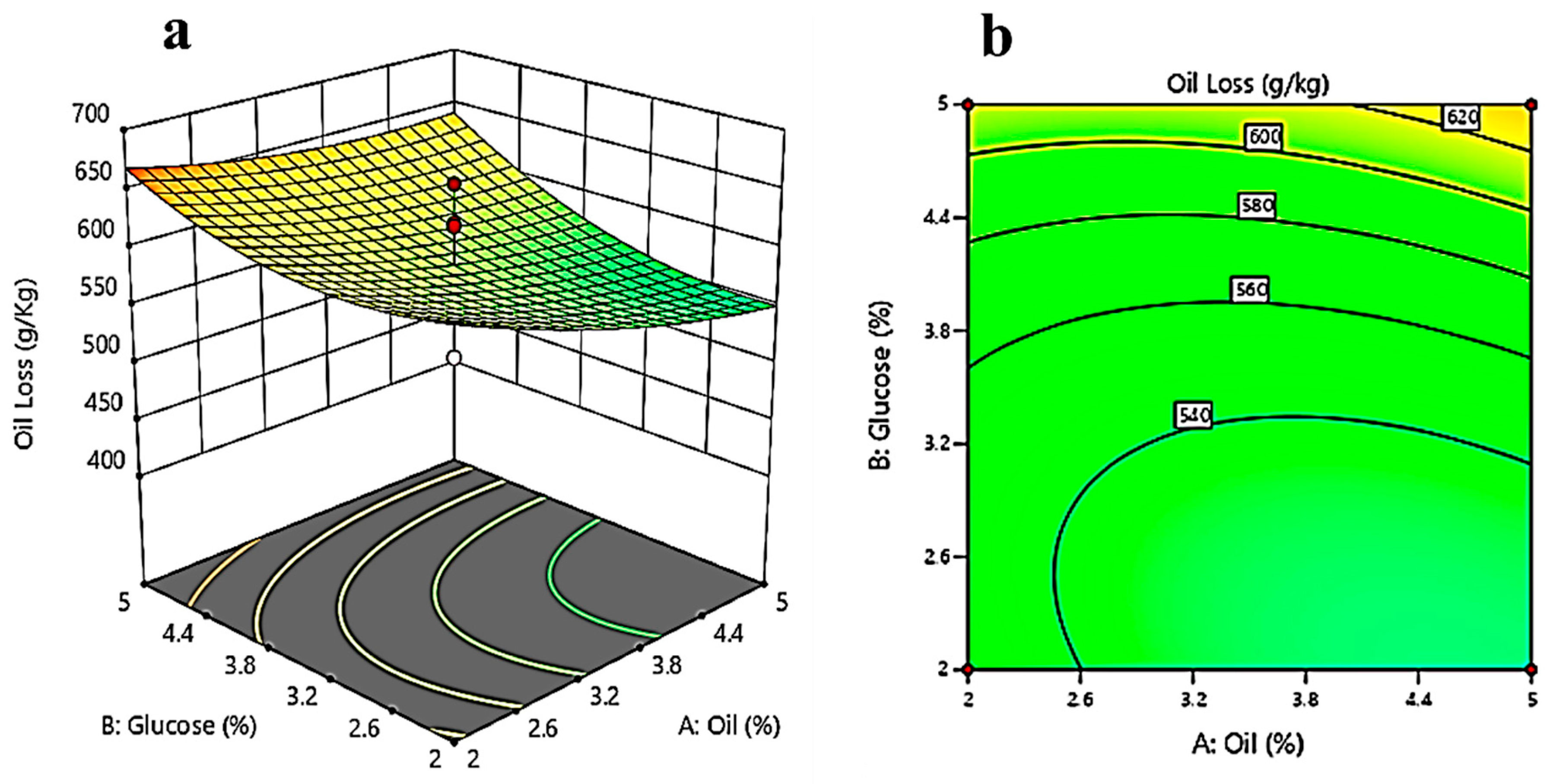
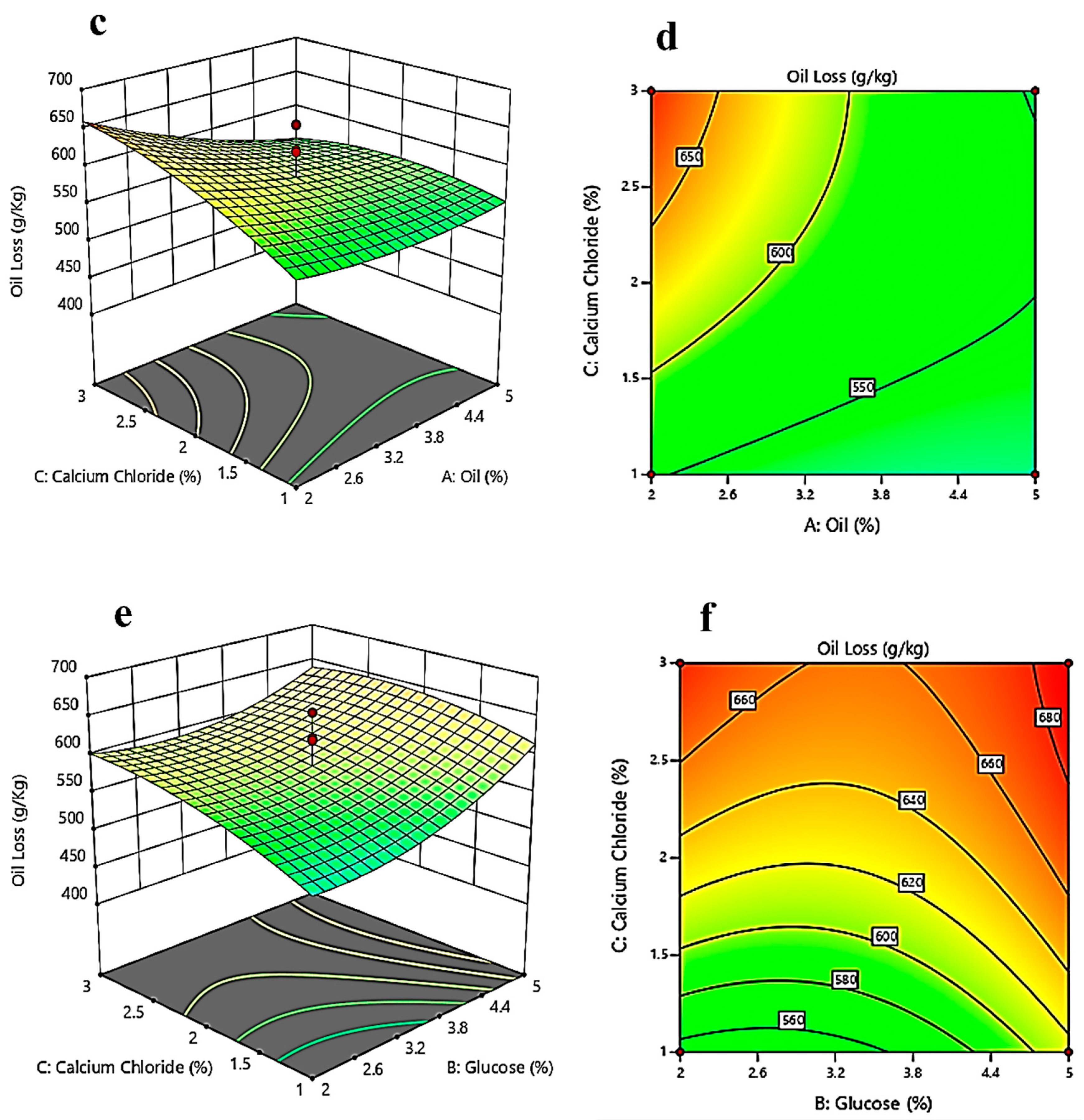
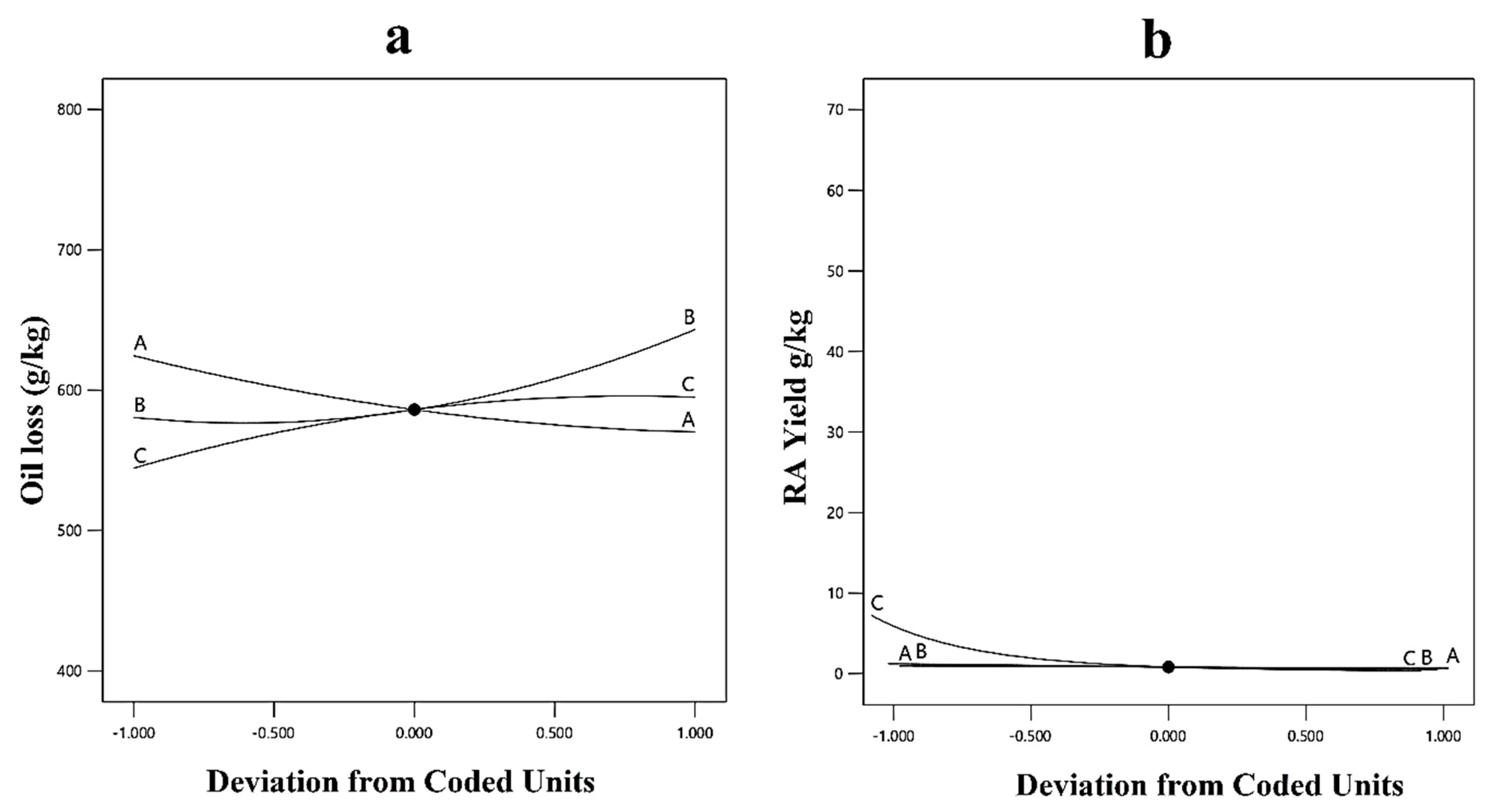
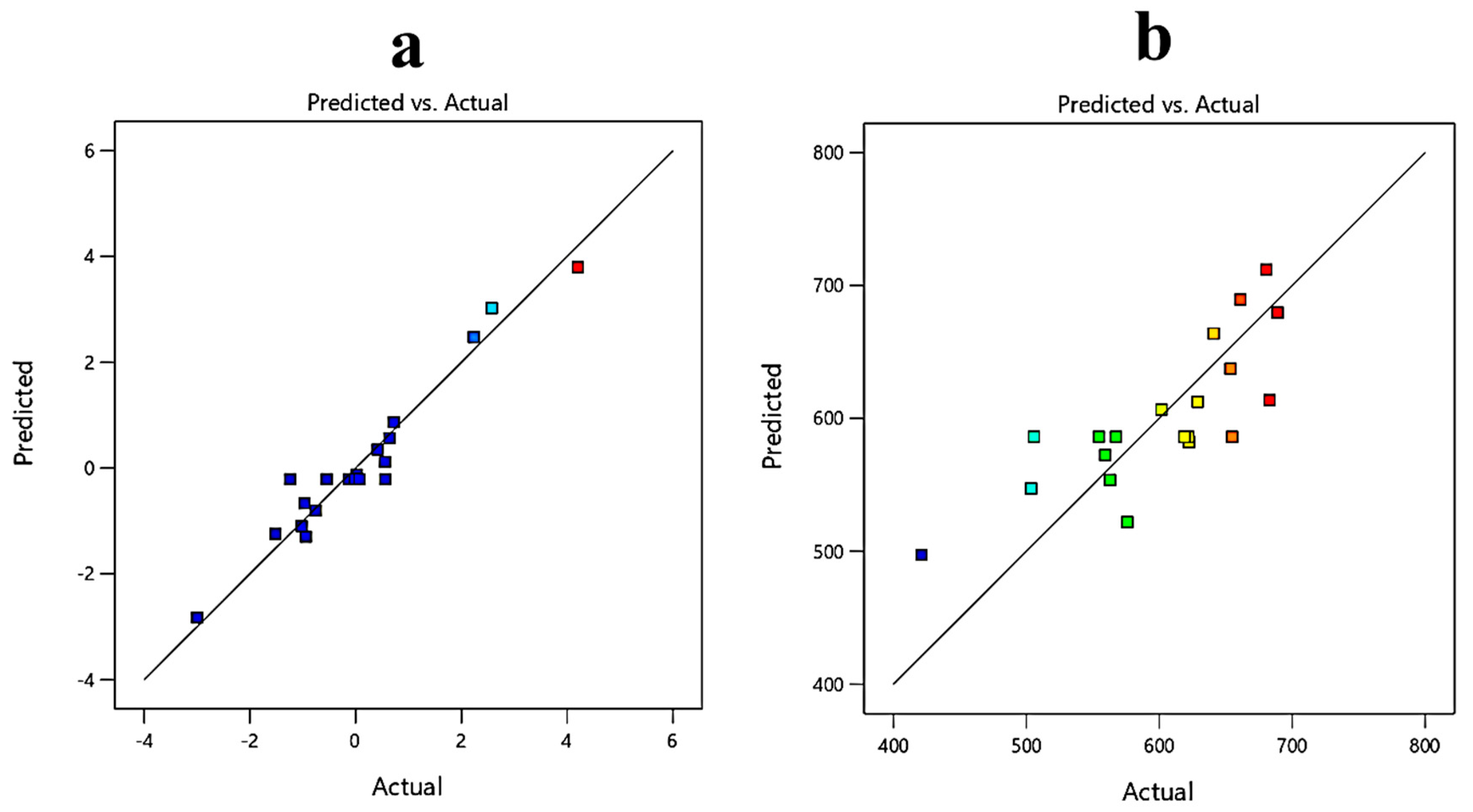
3.3. Response Surface Methodology
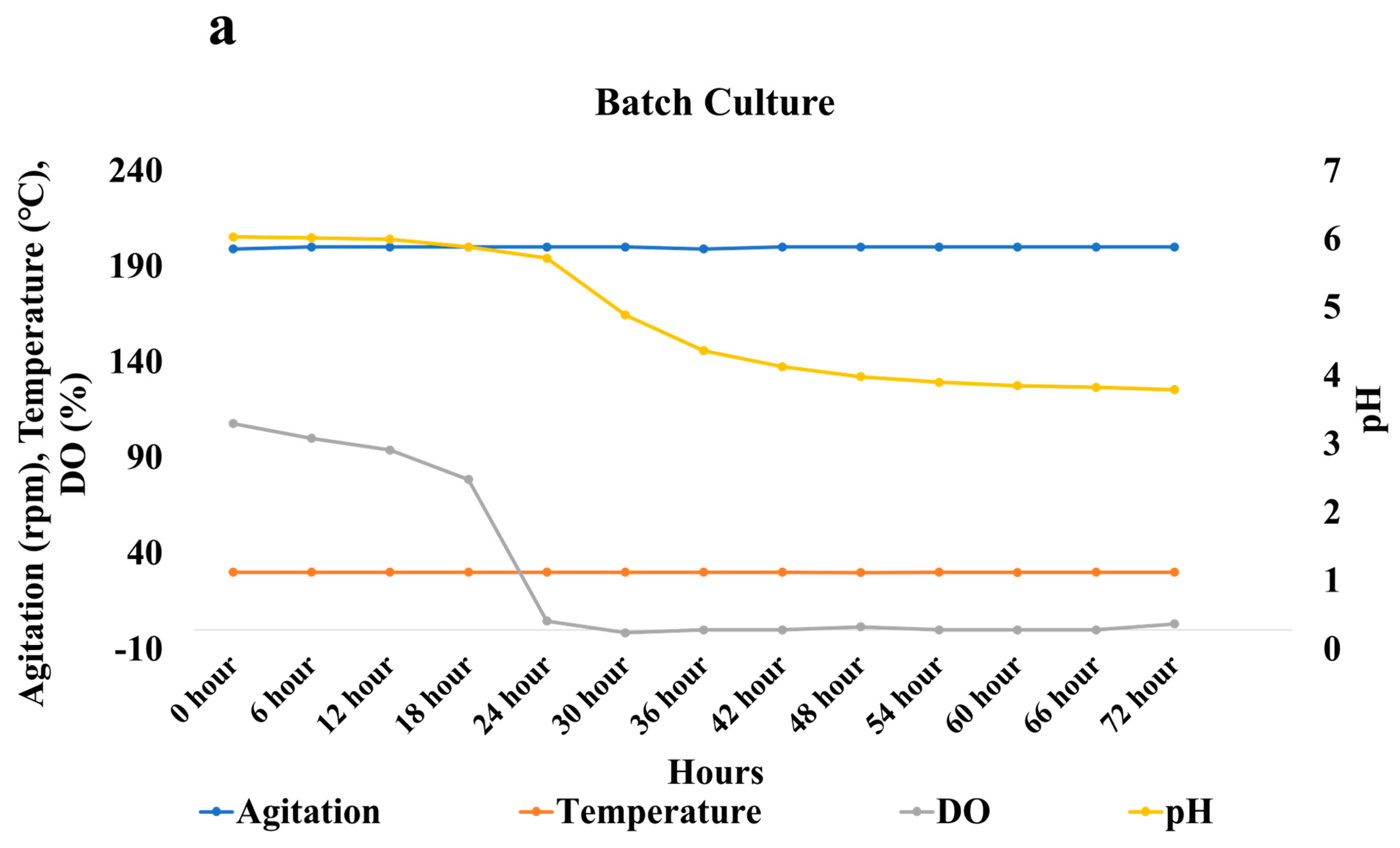
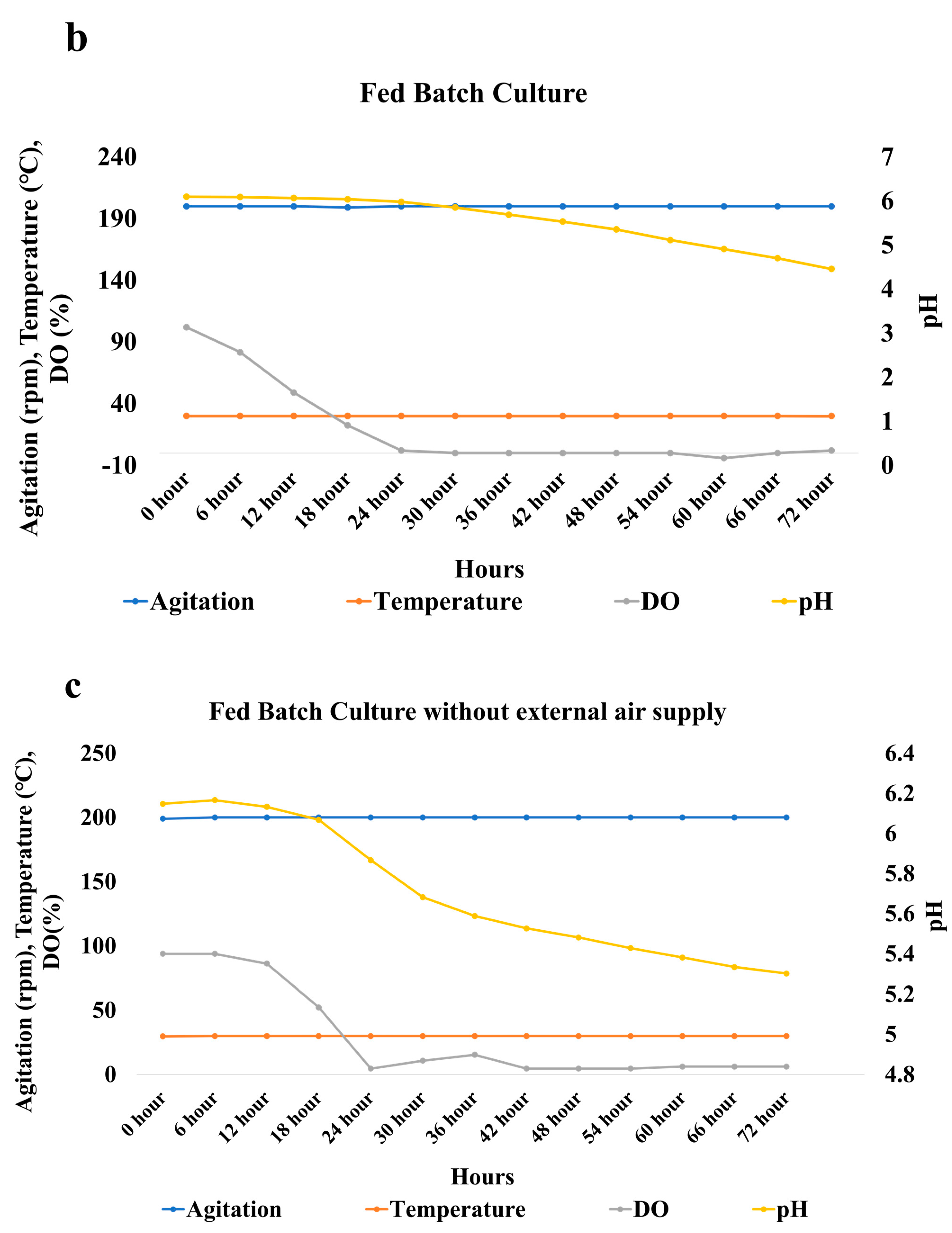
3.4. Bench Scale Fermentation in the Optimized Conditions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, S.; Sharma, S.; Sarma, S.J.; Brar, S.K. A Comprehensive Review of Castor Oil-Derived Renewable and Sustainable Industrial Products. Environ. Prog. Sustain. Energy 2022, 42, e14008. [Google Scholar] [CrossRef]
- Hoon, J.; Hee, S.; Yeub, I.; Byung, J.; Han, S.; Lee, H.; Park, C.; Yeol, E. Preparation of 11-Hexyloxy-9-Undecenoic Acid from Crude Castor Oil Hydrolysates by Recombinant Escherichia Coli Expressing Alcohol Dehydrogenase and Baeyer–Villiger Monooxygenase. Process Biochem. 2016, 51, 362–368. [Google Scholar] [CrossRef]
- Heilig, M.L. United States Patent Office. ACM SIGGRAPH Comput. Graph. 1994, 28, 131–134. [Google Scholar] [CrossRef]
- Patel, V.R.; Dumancas, G.G.; Viswanath, L.C.K.; Maples, R.; Subong, B.J.J. Castor Oil: Properties, Uses, and Optimization of Processing Parameters in Commercial Production. Lipid Insights 2016, 9, LPI-S40233. [Google Scholar] [CrossRef]
- Mubofu, E.B. Castor Oil as a Potential Renewable Resource for the Production of Functional Materials. Sustain. Chem. Process. 2016, 4, 11. [Google Scholar] [CrossRef]
- Mutlu, H.; Meier, M.A.R. Castor Oil as a Renewable Resource for the Chemical Industry. Eur. J. Lipid Sci. Technol. 2010, 112, 10–30. [Google Scholar] [CrossRef]
- Hablot, E.; Zheng, D.; Bouquey, M.; Ave, L. Polyurethanes Based on Castor Oil: Kinetics, Chemical, Mechanical and Thermal Properties. Macromol. Mater. Eng. 2002, 293, 922–929. [Google Scholar] [CrossRef]
- Naughton, F. Castor Oil. In Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2000; pp. 1–20. [Google Scholar]
- Kunduru, K.R.; Basu, A.; Zada, M.H.; Domb, A.J. Castor Oil Based Biodegradable Polyesters. Biomacromolecules 2015, 16, 2572. [Google Scholar] [CrossRef]
- Verma, K.K.; Kendurkar, P.S.; Tewari, N.; Prasad, R.N. Physico-Chemical Characteristics of Some Castorbean (Ricinus Communis L.) Varieties, Hybrids and Genotypes. Indian J. Agric. Biochem. 2007, 20, 43–45. [Google Scholar]
- Guimarães, A.; Venâncio, A. The Potential of Fatty Acids and Their Derivatives as Antifungal Agents: A Review. Toxins 2022, 14, 188. [Google Scholar] [CrossRef]
- Naik, S.N.; Saxena, D.K.; Dole, B.R.; Khare, S.K. Chapter 21—Potential and Perspective of Castor Biorefinery. In Waste Biorefinery; Elsevier: Amsterdam, The Netherlands, 2018; pp. 623–656. [Google Scholar] [CrossRef]
- Bloom, P.; Lee, I.; Reimers, P. Method for the Production of Fatty Acids Having a Low Trans-Fatty Acid Content. U.S. Patent No. 7,126,019, 24 October 2006. [Google Scholar]
- Sonntag, N.O.V. Fat Splitting. J. Am. Oil Chem. Soc. 1979, 56, 729A–732A. [Google Scholar] [CrossRef]
- Goswami, D.; Basu, J.K.; De, S. Ipase Applications in Oil Hydrolysis with a Case Study on Castor Oil: A Review. Crit. Rev. Biotechnol. 2013, 33, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Majid, S.A.; Hossain, A. Study on the Hydrolysis of Fats and Oils by Twitchell Reagent. Bangladesh J. Sci. Ind. Res. 1976, 44–50. [Google Scholar]
- Avelar, M.H.M.; Cassimiro, D.M.J.; Santos, K.C.; Domingues, R.C.C.; de Castro, H.F.; Mendes, A.A. Hydrolysis of Vegetable Oils Catalyzed by Lipase Extract Powder from Dormant Castor Bean Seeds. Ind. Crops Prod. 2013, 44, 452–458. [Google Scholar] [CrossRef]
- Goswami, D.; Basu, J.K.; De, S. Optimization of Process Variables in Castor Oil Hydrolysis by Candida Rugosa Lipase with Buffer as Dispersion Medium. Biotechnol. Bioprocess Eng. 2009, 14, 220–224. [Google Scholar] [CrossRef]
- Edwinoliver, N.G.; Thirunavukarasu, K.; Purushothaman, S.; Rose, C.; Gowthaman, M.K.; Kamini, N.R. Corn Steep Liquor as a Nutrition Adjunct for the Production of Aspergillus Niger Lipase and Hydrolysis of Oils Thereof. J. Agric. food Chem. 2009, 57, 10658–10663. [Google Scholar] [CrossRef]
- Dheeman, D.S.; Antony-Babu, S.; Frías, J.M.; Henehan, G.T.M. Purification and Characterization of an Extracellular Lipase from a Novel StrainPurification and Characterization of an Extracellular Lipase from a Novel Strain Penicillium sp. DS-39 (DSM 23773). J. Mol. Catal. B Enzym. 2011, 72, 256–262. [Google Scholar] [CrossRef]
- Sarada, R.; Joseph, R. Purification and Properties of Lipase from the Anaerobepropionibacterium Acidi-Propionici. J. Am. Oil Chem. Soc. 1992, 69, 974–977. [Google Scholar] [CrossRef]
- Syed, M.N.; Iqbal, S.; Bano, S.; Khan, A.B.; Ali-ul-, S. Purification and Characterization of 60 KD Lipase Linked with Chaperonin from Pseudomonas Aeruginosa BN-1. Afr. J. Biotechnol. 2010, 9, 7724–7732. [Google Scholar] [CrossRef]
- Ozcan, H.M.; Sagiroglu, A. Production of Ricinoleic Acid from Castor Oil by Immobilised Lipases. Prep. Biochem. Biotechnol. 2009, 39, 170–182. [Google Scholar] [CrossRef]
- Er-zheng, S.; Zhou, Y.; You, P.; Dong-zhi, W. Lipases in the Castor Bean Seed of Chinese Varieties: Activity Comparison, Purification and Characterization. J. Shanghai Univ. 2010, 14, 137–144. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, S.; Sarma, S.J.; Brar, S.K. Molecular Characterization of a New Strain of Aspergillus and Ricinoleic Acid Production from Castor Oil by the Fungus. Environ. Prog. Sustain. Energy 2023, Unpublished. [Google Scholar]
- Toscano, L.; Montero, G.; Stoytcheva, M.; Gochev, V.; Cervantes, L.; Campbell, H.; Zlatev, R.; Valdez, B.; Pérez, C.; Gil-Samaniego, M. Lipase Production through Solid-State Fermentation Using Agro-Industrial Residues as Substrates and Newly Isolated Fungal Strains. Biotechnol. Biotechnol. Equip. 2013, 27, 4074–4077. [Google Scholar] [CrossRef]
- Negedu, A.; Ameh, J.; Umoh, V.; Atawodi, S. Lipolytic Activity Of Some Fungal Species On Castor Oil. Afr. J. Food Agric. Nutr. Dev. 2012, 12, 6686–6699. [Google Scholar]
- Rathod, V.K.; Pandit, A.B. Effect of Various Additives on Enzymatic Hydrolysis of Castor Oil. Biochem. Eng. J. 2009, 47, 93–99. [Google Scholar] [CrossRef]
- Hu, M.; Li, Y.; Decker, E.A.; McClements, D.J. Role of Calcium and Calcium-Binding Agents on the Lipase Digestibility of Emulsified Lipids Using an in Vitro Digestion Model. Food Hydrocoll. 2010, 24, 719–725. [Google Scholar] [CrossRef]
- Goswami, D.; Sen, R.; Basu, J.K.; De, S. Maximization of Bioconversion of Castor Oil into Ricinoleic Acid by Response Surface Methodology. Bioresour. Technol. 2009, 100, 4067–4073. [Google Scholar] [CrossRef]
- Barrak, N.; Mannai, R.; Zaidi, M.; Kechida, M.; Helal, A.N.; Barrak, N.; Mannai, R.; Zaidi, M.; Kechida, M.; Helal, A.N. Experimental Design Approach with Response Surface Methodology for Removal of Indigo Dye by Electrocoagulation. J. Geosci. Environ. Prot. 2016, 4, 50–61. [Google Scholar] [CrossRef]
- Fatoba, O.S.; Akanji, O.L.; Aasa, A.S.; Fatoba, O.S.; Akanji, O.L.; Aasa, A.S. Optimization of Carburized UNS G10170 Steel Process Parameters Using Taguchi Approach and Response Surface Model (RSM). J. Miner. Mater. Charact. Eng. 2014, 2, 566–578. [Google Scholar] [CrossRef]
- Sen, R.; Babu, K.S. Modeling and Optimization of the Process Conditions for Biomass Production and Sporulation of a Probiotic Culture. Process Biochem. 2005, 40, 2531–2538. [Google Scholar] [CrossRef]
- Namal Senanayake, S.P.J.; Shahidi, F. Lipase-Catalyzed Incorporation of Docosahexaenoic Acid (DHA) into Borage Oil: Optimization Using Response Surface Methodology. Food Chem. 2002, 77, 115–123. [Google Scholar] [CrossRef]
- Shieh, C.-J.; Liao, H.-F.; Lee, C.-C. Optimization of Lipase-Catalyzed Biodiesel by Response Surface Methodology. Bioresour. Technol. 2003, 88, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Carter-House, D.; E Stajich, J.; Unruh, S.; Kurbessoian, T. Fungal CTAB DNA Extraction. Protocols.io 2020. [Google Scholar] [CrossRef]
- Nargesi, S.; Abastabar, M.; Valadan, R.; Mayahi, S.; Youn, J.H.; Hedayati, M.T.; Seyedmousavi, S. Differentiation of Aspergillus Flavus from Aspergillus Oryzae Targeting the Cyp51A Gene. Pathogens 2021, 10, 1279. [Google Scholar] [CrossRef]
- Mehmood, T.; Ahmed, A.; Ahmad, A.; Ahmad, M.S.; Sandhu, M.A. Optimization of Mixed Surfactants-Based β-Carotene Nanoemulsions Using Response Surface Methodology: An Ultrasonic Homogenization Approach. Food Chem. 2018, 253, 179–184. [Google Scholar] [CrossRef]
- Bódalo-Santoyo, A.; Bastida-Rodríguez, J.; Máximo-Martín, M.F.; Montiel-Morte, M.C.; Murcia-Almagro, M.D. Enzymatic Biosynthesis of Ricinoleic Acid Estolides. Biochem. Eng. J. 2005, 26, 155–158. [Google Scholar] [CrossRef]
- Joyce, R.M. Experiment Optimization in Chemistry and Chemical Engineering, S. Akhnazarova and V. Kafarov, Mir Publishers, Moscow and Chicago, 1982, 312 Pp. Price: $9.95. J. Polym. Sci. Polym. Lett. Ed. 1984, 22, 372. [Google Scholar] [CrossRef]
- Khuri, A.I.; Cornell, J.A. Response Surfaces Designs and Analyses; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar] [CrossRef]
- Box, G.E.P.; Hunter, J.S.; Hunter, W.G. Statistics Experimenters; John Wiley & Sons: Hoboken, NJ, USA, 2009; pp. 1–655. [Google Scholar]
- Box, G.E.P.; Wilson, K.B. On the Experimental Attainment of Optimum Conditions. J. R. Stat. Soc. Ser. B 1951, 13, 1–38. [Google Scholar] [CrossRef]
- Osho, M.; Popoola, T.; Adeleye, T.; Adetunji, C. Response Surface Methodology for Optimal Immobilization of Aspergillus Niger ATCC 1015 Lipase by Adsorption Method. Int. J. Biol. Res. 2016, 4, 56. [Google Scholar] [CrossRef]
- Sun, S.; Guo, J. Enhanced Ricinoleic Acid Preparation Using Lipozyme TLIM as a Novel Biocatalyst: Optimized by Response Surface Methodology. Catalysts 2018, 8, 486. [Google Scholar] [CrossRef]
- Bhattacharjee, D.; Goswami, D. Surfactant Assisted Production of Ricinoleic Acid Using Cross-Linked and Entrapped Porcine Pancreas Lipase. J. Dispers. Sci. Technol. 2021, 42, 947–955. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Symbols | Range and Levels | ||||
|---|---|---|---|---|---|---|
| −α | −1 | 0 | +1 | +α | ||
| Concentration of oil (%) | x1 | 0.9773 | 2 | 3.5 | 5 | 6.02 |
| Glucose concentration (%) | x2 | 0.9773 | 2 | 3.5 | 5 | 6.02 |
| Concentration of calcium chloride (%) | x3 | 0.3182 | 1 | 2 | 3 | 3.68 |
| Run | Independent Variables | Response/Output | |||
|---|---|---|---|---|---|
| x1 (%) | x2 (%) | x3 (%) | RA Yield (g/kg) | Oil Loss (g/kg) | |
| 1 | 3.5 | 3.5 | 2 | 0.29 | 654.85 |
| 2 | 3.5 | 3.5 | 0.32 | 66.85 | 421.14 |
| 3 | 3.5 | 3.5 | 2 | 0.88 | 621.71 |
| 4 | 2 | 5 | 3 | 0.05 | 661 |
| 5 | 2 | 5 | 1 | 9.35 | 683 |
| 6 | 3.5 | 3.5 | 3.68 | 0.22 | 622.28 |
| 7 | 5 | 2 | 1 | 1.9 | 576 |
| 8 | 3.5 | 6.02 | 2 | 0.36 | 680.57 |
| 9 | 3.5 | 3.5 | 2 | 1.08 | 567.42 |
| 10 | 5 | 2 | 3 | 1.02 | 503.6 |
| 11 | 3.5 | 0.99 | 2 | 1.51 | 601.71 |
| 12 | 5 | 5 | 1 | 2.06 | 653.6 |
| 13 | 2 | 2 | 3 | 0.47 | 689 |
| 14 | 5 | 5 | 3 | 0.39 | 628.8 |
| 15 | 6.02 | 3.5 | 2 | 0.38 | 559.32 |
| 16 | 0.97 | 3.5 | 2 | 1.74 | 640.81 |
| 17 | 3.5 | 3.5 | 2 | 0.58 | 618.85 |
| 18 | 3.5 | 3.5 | 2 | 0.99 | 554.54 |
| 19 | 2 | 2 | 1 | 13.13 | 563 |
| 20 | 3.5 | 3.5 | 2 | 1.75 | 505.71 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 44.50 | 9 | 4.94 | 17.14 | <0.0001 |
| Pure error | 1.89 | 5 | 0.3780 | ||
| Cor Total | 47.39 | 19 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 56,493.00 | 9 | 6277.00 | 1.79 | 0.1894 |
| Pure error | 14,867.73 | 5 | 2973.55 | ||
| COR. Total | 91,624.89 | 19 |
| Factors | RA Yield (g/kg) | Oil Loss (g/kg) | ||||
|---|---|---|---|---|---|---|
| Coefficient Estimate | Standard Error | p-Value | Coefficient Estimate | Standard Error | p-Value | |
| Intercept | −0.2101 | 0.2191 | - | 586.14 | 24.17 | 0.1894 |
| –Concentration of Oil (%) | −0.2325 | 0.1454 | 0.1408 | −27.17 | 16.04 | 0.1211 |
| –Glucose Concentration | −0.4300 | 0.1454 | 0.0143 | 31.30 | 16.04 | 0.0796 |
| −Concentration of Calcium Chloride | −1.50 | 0.1454 | <0.0001 | 25.27 | 16.04 | 0.1462 |
| 0.2125 | 0.1899 | 0.2894 | 13.85 | 20.96 | 0.5236 | |
| 0.7843 | 0.1899 | 0.0020 | −25.15 | 20.96 | 0.2577 | |
| −0.3679 | 0.1899 | 0.0815 | −12.55 | 20.96 | 0.5626 | |
| −0.0227 | 0.1415 | 0.8757 | 11.33 | 15.61 | 0.4848 | |
| −0.0573 | 0.1415 | 0.6939 | 25.85 | 15.61 | 0.1288 | |
| 0.5257 | 0.1415 | 0.0040 | −16.38 | 15.61 | 0.3189 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.; Sharma, S.; Sarma, S.J.; Brar, S.K. Roles of Process Parameters on the Ricinoleic Acid Production from Castor Oil by Aspergillus flavus BU22S. Fermentation 2023, 9, 318. https://doi.org/10.3390/fermentation9040318
Singh S, Sharma S, Sarma SJ, Brar SK. Roles of Process Parameters on the Ricinoleic Acid Production from Castor Oil by Aspergillus flavus BU22S. Fermentation. 2023; 9(4):318. https://doi.org/10.3390/fermentation9040318
Chicago/Turabian StyleSingh, Shikha, Sumit Sharma, Saurabh Jyoti Sarma, and Satinder Kaur Brar. 2023. "Roles of Process Parameters on the Ricinoleic Acid Production from Castor Oil by Aspergillus flavus BU22S" Fermentation 9, no. 4: 318. https://doi.org/10.3390/fermentation9040318
APA StyleSingh, S., Sharma, S., Sarma, S. J., & Brar, S. K. (2023). Roles of Process Parameters on the Ricinoleic Acid Production from Castor Oil by Aspergillus flavus BU22S. Fermentation, 9(4), 318. https://doi.org/10.3390/fermentation9040318