

Combined Use of Schizosaccharomyces pombe and a Lachancea thermotolerans Strain with a High Malic Acid Consumption Ability for Wine Production
 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Vinification
2.3. Chemical Parameter Measurements
2.4. Volatile Compounds
2.5. Color Intensity
2.6. Statistical Analyses
3. Results and Discussion
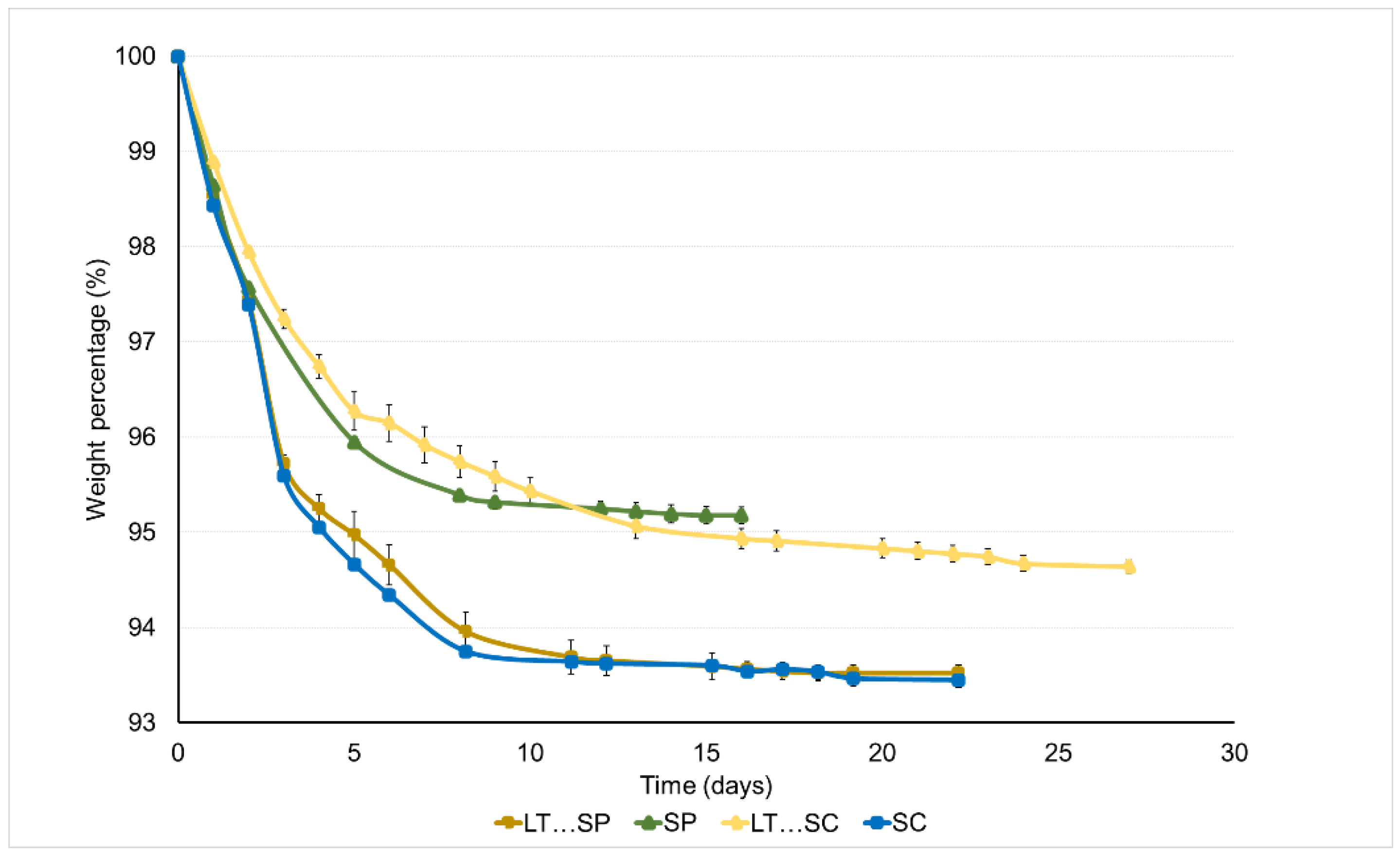
3.1. Fermentation Kinetics
3.2. Glucose and Fructose
3.3. Ethanol
3.4. l-Lactic Acid
3.5. Malic Acid
3.6. Acetic Acid
3.7. Succinic Acid
3.8. Glycerol
3.9. Volatile Compounds
3.10. Color Intensity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benito, S. Combined Use of Lachancea thermotolerans and Schizosaccharomyces pombe in Winemaking: A Review. Microorganisms 2020, 8, 655. [Google Scholar] [CrossRef]
- Hranilovic, A.; Gambetta, J.M.; Schmidtke, L.; Boss, P.K.; Grbin, P.R.; Masneuf-Pomarede, I.; Bely, M.; Albertin, W.; Jiranek, V. Oenological Traits of Lachancea thermotolerans Show Signs of Domestication and Allopatric Differentiation. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Hranilovic, A.; Albertin, W.; Capone, D.L.; Gallo, A.; Grbin, P.R.; Danner, L.; Bastian, S.E.P.; Masneuf-Pomarede, I.; Coulon, J.; Bely, M.; et al. Impact of Lachancea thermotolerans on Chemical Composition and Sensory Profiles of Merlot Wines. Food Chem. 2021, 349, 129015. [Google Scholar] [CrossRef]
- Blanco, P.; Rabuñal, E.; Neira, N.; Castrillo, D. Dynamic of Lachancea thermotolerans Population in Monoculture and Mixed Fermentations: Impact on Wine Characteristics. Beverages 2020, 6, 36. [Google Scholar] [CrossRef]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The Role and Use of Non-Saccharomyces Yeasts in Wine Production. S. Afr. J. Enol. Vitic. 2006, 27, 15–38. [Google Scholar] [CrossRef]
- Benito, S. The Impacts of Lachancea thermotolerans Yeast Strains on Winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 6775–6790. [Google Scholar] [CrossRef]
- Vicente, J.; Navascués, E.; Calderón, F.; Santos, A.; Marquina, D.; Benito, S.; Fracassetti, D.; Rustioni, L. An Integrative View of the Role of Lachancea thermotolerans in Wine Technology. Foods 2021, 10, 2878. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. Combined Use of S. pombe and L. thermotolerans in Winemaking. Beneficial Effects Determined through the Study of Wines’ Analytical Characteristics. Molecules 2016, 21, 1744. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. Mixed Alcoholic Fermentation of Schizosaccharomyces pombe and Lachancea thermotolerans and Its Influence on Mannose-Containing Polysaccharides Wine Composition. AMB Express 2019, 9, 17. [Google Scholar] [CrossRef]
- Vicente, J.; Baran, Y.; Navascués, E.; Santos, A.; Calderón, F.; Marquina, D.; Rauhut, D.; Benito, S. Biological Management of Acidity in Wine Industry: A Review. Int. J. Food Microbiol. 2022, 375, 109726. [Google Scholar] [CrossRef]
- del Mónaco, S.M.; Barda, N.B.; Rubio, N.C.; Caballero, A.C. Selection and Characterization of a Patagonian Pichia kudriavzevii for Wine Deacidification. J. Appl. Microbiol. 2014, 117, 451–464. [Google Scholar] [CrossRef]
- van Wyk, N.; Scansani, S.; Beisert, B.; Brezina, S.; Fritsch, S.; Semmler, H.; Pretorius, I.S.; Rauhut, D.; von Wallbrunn, C. The Use of Hanseniaspora occidentalis in a Sequential Must Inoculation to Reduce the Malic Acid Content of Wine. Appl. Sci. 2022, 12, 6919. [Google Scholar] [CrossRef]
- Liu, S.; Laaksonen, O.; Yang, B. Volatile Composition of Bilberry Wines Fermented with Non-Saccharomyces and Saccharomyces Yeasts in Pure, Sequential and Simultaneous Inoculations. Food Microbiol. 2019, 80, 25–39. [Google Scholar] [CrossRef]
- Benito, S. The Impacts of Schizosaccharomyces on Winemaking. Appl. Microbiol. Biotechnol. 2019, 103, 4291–4312. [Google Scholar] [CrossRef] [PubMed]
- Urbina, Á.; Calderón, F.; Benito, S. The Combined Use of Lachancea thermotolerans and Lactiplantibacillus plantarum (Former Lactobacillus plantarum) in Wine Technology. Foods 2021, 10, 1356. [Google Scholar] [CrossRef]
- Benito, Á.; Jeffares, D.; Palomero, F.; Calderón, F.; Bai, F.Y.; Bähler, J.; Benito, S. Selected Schizosaccharomyces pombe Strains Have Characteristics That Are Beneficial for Winemaking. PLoS ONE 2016, 11, e0151102. [Google Scholar] [CrossRef]
- Chen, K.; Escott, C.; Loira, I.; del Fresno, J.M.; Morata, A.; Tesfaye, W.; Calderon, F.; Suárez-Lepe, J.A.; Han, S.; Benito, S. Use of Non-Saccharomyces Yeasts and Oenological Tannin in Red Winemaking: Influence on Colour, Aroma and Sensorial Properties of Young Wines. Food Microbiol. 2018, 69, 51–63. [Google Scholar] [CrossRef]
- YuHua, W.; WenJun, S.; Min, L.; Lan, M.; YuMei, J.; Jing, W. Effect of Sequential Fermentation with Lachancea thermotolerans and Schizosaccharomyces pombe on the Quality of Merlot Dry Red Wine. Shipin Kexue/Food Sci. 2019, 40, 102–111. [Google Scholar]
- Ruiz, J.; Kiene, F.; Belda, I.; Fracassetti, D.; Marquina, D.; Navascués, E.; Calderón, F.; Benito, A.; Rauhut, D.; Santos, A.; et al. Effects on Varietal Aromas during Wine Making: A Review of the Impact of Varietal Aromas on the Flavor of Wine. Appl. Microbiol. Biotechnol. 2019, 103, 7425–7450. [Google Scholar] [CrossRef] [PubMed]
- Gardoni, E.; Benito, S.; Scansani, S.; Brezina, S.; Fritsch, S.; Rauhut, D. Biological Deacidification Strategies for White Wines. S. Afr. J. Enol. Vitic. 2021, 42, 114–122. [Google Scholar] [CrossRef]
- Scansani, S.; Rauhut, D.; Brezina, S.; Semmler, H.; Benito, S. The Impact of Chitosan on the Chemical Composition of Wines Fermented with Schizosaccharomyces pombe and Saccharomyces Cerevisiae. Foods 2020, 9, 1423. [Google Scholar] [CrossRef] [PubMed]
- Benito, Á.; Calderón, F.; Benito, S. The Combined Use of Schizosaccharomyces pombe and Lachancea thermotolerans—Effect on the Anthocyanin Wine Composition. Molecules 2017, 22, 739. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| SC | S. cerevisiae (106 CFU/mL) alone. |
| LT…SC | L. thermotolerans (106 CFU/mL) followed by S. cerevisiae (106 CFU/mL) 5 days later. |
| LT…SP | L. thermotolerans (106 CFU/mL) followed by S. pombe (106 CFU/mL) 5 days later. |
| SP | S. pombe (106 CFU/mL) alone. |
| SC | LT…SC | LT…SP | SP | |
|---|---|---|---|---|
| l-lactic acid (g/L) | 0.11 ± 0.03 a | 2.26 ± 0.64 c | 1.52 ± 0.09 b | 0.16 ± 0.02 a |
| l-malic acid (g/L) | 1.41 ± 0.03 d | 1.11 ± 0.10 c | 0.11 ± 0.03 a | 0.48 ± 0.07 b |
| Succinic acid (g/L) | 1.47 ± 0.06 c | 1.41 ± 0.07 bc | 1.34 ± 0.03 b | 1.26 ± 0.04 a |
| Acetic acid (g/L) | 0.36 ± 0.05 a | 0.51 ± 0.07 b | 0.45 ± 0.03 b | 0.45 ± 0.01 b |
| pH | 3.77 ± 0.02 b | 3.62 ± 0.02 a | 3.78 ± 0.04 b | 3.83 ± 0.01 c |
| Ethanol (g/L) | 11.63 ± 0.06 c | 11.21 ± 0.06 a | 11.46 ± 0.06 b | 11.72 ± 0.04 c |
| Glucose + Fructose (g/L) | 1.24 ± 0.21 a | 4.73 ± 0.06 c | 1.85 ± 0.24 b | 1.56 ± 0.14 ab |
| Glycerol (g/L) | 9.25 ± 0.32 b | 8.96 ± 0.47 ab | 9.13 ± 0.26 b | 8.26 ± 0.14 a |
| Compound (Area Units) | SC | SP | LT…SC | LT…SP |
|---|---|---|---|---|
| Ethyl acetate | 1.42 ± 0.02 a | 1.59 ± 0.2 a | 0.77 ± 0.41 b | 1.43 ± 0.14 a |
| 3-Methyl butanal * | 2.70 ± 0.62 a | 0.00 ± 0.00 a | 0.87 ± 0.70 a | 1.88 ± 2.45 a |
| 2-Methylpropyl acetate | 0.03 ± 0.00 a | 0.02 ± 0.00 a | 0.01 ± 0.01 b | 0.03 ± 0.00 a |
| Ethyl butanoate | 0.05 ± 0.01 a | 0.05 ± 0.01 a | 0.01 ± 0.01 b | 0.053 ± 0.01 a |
| Toluene | 0.15 ± 0.14 a | 0.11 ± 0.10 a | 0.13 ± 0.11 a | 0.10 ± 0.09 a |
| Ethyl 3-methylbutanoate * | 5.56 ± 1.04 a | 2.93 ± 0.28 ab | 1.97 ± 1.89 b | 4.95 ± 1.43 ab |
| 1-(1-Ethoxyethoxy)pentane * | 0.00 ± 0.00 b | 3.24 ± 0.99 a | 0.00 ± 0.00 b | 3.17 ± 1.89 a |
| 2-Methyl-1-propanol | 0.89 ± 0.14 a | 0.69 ± 0.09 ab | 0.48 ± 0.08 b | 0.92 ± 0.25 a |
| 3-Methyl-1-butyl acetate | 0.55 ± 0.07 a | 0.39 ± 0.08 a | 0.25 ± 0.20 a | 0.26 ± 0.07 a |
| Butanol * | 2.58 ± 2.65 b | 3.73 ± 3.35 b | 31.42 ± 13.54 a | 7.88 ± 0.95 b |
| 3-Methyl-1-butanol | 6.65 ± 0.37 a | 5.88 ± 0.43 a | 5.06 ± 0.23 a | 6.25 ± 1.07 a |
| Ethyl hexanoate | 0.48 ± 0.10 a | 0.51 ± 0.09 a | 0.21 ± 0.08 a | 0.42 ± 0.26 a |
| Ethyl lactate | 0.03 ± 0.01 c | 0.02 ± 0.00 c | 0.19 ± 0.04 a | 0.11 ± 0.01 b |
| Hexanol | 0.29 ± 0.01 a | 0.28 ± 0.02 a | 0.26 ± 0.00 a | 0.29 ± 0.02 a |
| cis-3-Hexen-1-ol * | 20.35 ± 1.91 a | 18.19 ± 1.51 ab | 17.41 ± 1.23 ab | 13.38 ± 3.91 b |
| Ethyl octanoate | 0.11 ± 0.03 b | 0.15 ± 0.02 ab | 0.01 ± 0.00 c | 0.19 ± 0.03 a |
| 2-Nonanol * | 4.82 ± 2.04 b | 10.51 ± 1.95 a | 2.71 ± 0.23 b | 4.85 ± 1.33 b |
| Ethyl 3-hydroxybutyrate * | 3.76 ± 1.12 a | 4.44 ± 0.34 a | 1.70 ± 0.08 b | 2.99 ± 0.85 ab |
| Benzaldehyde | 0.03 ± 0.00 a | 0.02 ± 0.00 b | 0.02 ± 0.00 ab | 0.02 ± 0.00 ab |
| Ethyl nonanoate * | 7.92 ± 1.35 a | 7.96 ± 2.38 a | 1.53 ± 0.62 b | 8.51 ± 0.86 a |
| Ethyl 2-hydroxy-4-methylpentanoate * | 14.75 ± 0.28 a | 10.69 ± 0.67 b | 11.09 ± 0.59 b | 15.78 ± 1.04 a |
| Octanol * | 11.93 ± 3.63 a | 13.07 ± 1.85 a | 3.66 ± 1.36 b | 13.54 ± 2.62 a |
| 2-Methyl propanoic acid | 0.03 ± 0.00 a | 0.02 ± 0.00 a | 0.03 ± 0.01 a | 0.02 ± 0.00 a |
| Isoamyl lactate | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.03 ± 0.00 a | 0.03 ± 0.00 a |
| Ethyl 2-furoate * | 1.32 ± 0.14 a | 1.01 ± 0.08 b | 1.03 ± 0.11 ab | 1.11 ± 0.12 ab |
| Methyl benzoate * | 1.95 ± 0.27 a | 0.98 ± 0.03 c | 1.73 ± 0.11 ab | 1.33 ± 0.15 bc |
| Butanoic acid * | 2.04 ± 0.30 a | 1.94 ± 0.31 a | 1.62 ± 0.16 a | 1.68 ± 0.52 a |
| Ethyl decanoate | 0.01 ± 0.00 a | 0.02 ± 0.00 a | 0.00 ± 0.00 b | 0.02 ± 0.04 a |
| Butyrolactone * | 7.92 ± 0.11 a | 4.08 ± 0.99 b | 6.52 ± 0.71 a | 8.26 ± 1.12 a |
| 4-methylbenzaldehyde * | 9.11 ± 1.74 a | 6.36 ± 1.15 a | 7.42 ± 1.12 a | 7.30 ± 1.12 a |
| Acetophenone * | 2.23 ± 0.90 ab | 1.50 ± 0.36 b | 2.17 ± 0.77 ab | 3.79 ± 0.80 a |
| 2-methyl butanoic acid | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a |
| Diethyl succinate | 0.08 ± 0.01 b | 0.05 ± 0.01 b | 0.04 ± 0.01 b | 0.27 ± 0.11 a |
| Dodecanal * | 4.52 ± 0.57 ab | 4.68 ± 0.39 a | 3.26 ± 0.15 b | 4.21 ± 0.70 ab |
| 3-(methylthio)-1-propanol * | 7.06 ± 1.32 b | 6.63 ± 1.27 b | 3.70 ± 0.89 c | 10.83 ± 0.72 a |
| Ethyl phenylacetate * | 1.70 ± 0.13 ab | 1.22 ± 0.12 b | 1.76 ± 0.31 a | 1.84 ± 0.14 a |
| 2-phenylethyl acetate | 0.02 ± 0.00 a | 0.01 ± 0.00 a | 0.02 ± 0.00 a | 0.01 ± 0.00 a |
| β-damascenone * | 4.75 ± 0.92 a | 3.73 ± 0.58 a | 4.61 ± 0.30 a | 4.82 ± 0.68 a |
| Ethyl dodecanoate * | 0.80 ± 0.12 a | 1.12 ± 0.13 a | 0.00 ± 0.00 b | 1.26 ± 0.36 a |
| Hexanoic acid | 0.07 ± 0.00 a | 0.07 ± 0.01 a | 0.04 ± 0.00 b | 0.07 ± 0.01 a |
| N-(3-Methylbutyl)acetamide | 0.06 ± 0.01 b | 0.05 ± 0.00 b | 0.05 ± 0.00 b | 0.09 ± 0.01 a |
| Butanedioic acid, ethyl 3-methylbutyl ester * | 2.30 ± 0.16 b | 1.77 ± 0.19 b | 1.22 ± 0.21 b | 10.54 ± 4.61 a |
| Phenylethyl alcohol | 1.44 ± 0.08 a | 1.22 ± 0.11 a | 1.43 ± 0.25 a | 1.55 ± 0.13 a |
| Octanoic acid | 0.20 ± 0.02 a | 0.23 ± 0.01 a | 0.08 ± 0.00 b | 0.20 ± 0.01 a |
| Nonanoic acid | 0.04 ± 0.00 a | 0.05 ± 0.02 a | 0.05 ± 0.01 a | 0.04 ± 0.02 a |
| Decanoic acid * | 25.98 ± 2.68 a | 31.34 ± 2.19 a | 28.92 ± 2.54 a | 17.17 ± 1.96 b |
| SC | LT…SC | LT…SP | SP | |
|---|---|---|---|---|
| 420 nm | 0.63 ± 0.02 a | 0.65 ± 0.04 a | 0.66 ± 0.03 a | 0.69 ± 0.04 a |
| 520 nm | 1.22 ± 0.04 a | 1.30 ± 0.06 a | 1.26 ± 0.05 a | 1.29 ± 0.06 a |
| 620 nm | 0.19 ± 0.01 a | 0.21 ± 0.02 a | 0.19 ± 0.02 a | 0.21 ± 0.03 a |
| CI | 2.04 ± 0.07 a | 2.16 ± 0.12 a | 2.11 ± 0.10 a | 2.19 ± 0.13 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vicente, J.; Kelanne, N.; Navascués, E.; Calderón, F.; Santos, A.; Marquina, D.; Yang, B.; Benito, S. Combined Use of Schizosaccharomyces pombe and a Lachancea thermotolerans Strain with a High Malic Acid Consumption Ability for Wine Production. Fermentation 2023, 9, 165. https://doi.org/10.3390/fermentation9020165
Vicente J, Kelanne N, Navascués E, Calderón F, Santos A, Marquina D, Yang B, Benito S. Combined Use of Schizosaccharomyces pombe and a Lachancea thermotolerans Strain with a High Malic Acid Consumption Ability for Wine Production. Fermentation. 2023; 9(2):165. https://doi.org/10.3390/fermentation9020165
Chicago/Turabian StyleVicente, Javier, Niina Kelanne, Eva Navascués, Fernando Calderón, Antonio Santos, Domingo Marquina, Baoru Yang, and Santiago Benito. 2023. "Combined Use of Schizosaccharomyces pombe and a Lachancea thermotolerans Strain with a High Malic Acid Consumption Ability for Wine Production" Fermentation 9, no. 2: 165. https://doi.org/10.3390/fermentation9020165
APA StyleVicente, J., Kelanne, N., Navascués, E., Calderón, F., Santos, A., Marquina, D., Yang, B., & Benito, S. (2023). Combined Use of Schizosaccharomyces pombe and a Lachancea thermotolerans Strain with a High Malic Acid Consumption Ability for Wine Production. Fermentation, 9(2), 165. https://doi.org/10.3390/fermentation9020165