
Xylitol Production by Debaryomyces hansenii in Extracted Olive Pomace Dilute-Acid Hydrolysate
 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Raw Material
2.2. Preparation and Treatment of EOP Hydrolysate
2.3. Detoxification Procedures
2.3.1. Activated Charcoal Treatment
2.3.2. Treatment with Anionic-Exchange Resins
2.3.3. Adsorptive Membranes (Nanofiltration)
2.3.4. Evaporation
2.4. Microorganism and Inoculum Conditions
2.5. Medium and Fermentation Conditions
2.6. Analytical Methods
3. Results and Discussion
3.1. EOP Hydrolysate Composition
3.2. Effect of Detoxification on the Chemical Composition of the Hydrolysate
3.3. Effect of Detoxification on Xylitol Production
3.3.1. Evaporation
3.3.2. Activated Charcoal Adsorption
3.3.3. Anionic Resins
3.3.4. Nanofiltration and Supplementation
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Available online: https://agriculture.ec.europa.eu/farming/crop-productions-and-plant-based-products/olive-oil_en (accessed on 20 September 2023).
- de Queiroz, S.S.; Jofre, F.M.; de Bianchini, I.A.; Bordini, F.W.; da Boaes, T.S.; Chandel, A.K.; Felipe, M.G.A. Fermentative production of xylitol from various lignocellulosic hydrolysates. In Current Advances in Biotechnological Production of Xylitol; de Almeida Felipe, M.d.G., Chandel, A.K., Eds.; Springer: Cham, Switzerland, 2022; pp. 51–56. [Google Scholar] [CrossRef]
- Galbe, M.; Wallberg, O. Pretreatment for biorefineries: A review of common methods for efficient utilisation of lignocellulosic materials. Biotechnol. Biofuels. 2019, 23, 2–26. [Google Scholar] [CrossRef] [PubMed]
- Padilla-Rascón, C.; Ruiz, E.; Romero, I.; Castro, E.; Oliva, J.M.; Ballesteros, M.; Manzanares, P. Valorisation of olive stone by-product for sugar production using a sequential acid/steam explosion pretreatment. Ind. Crops Prod. 2020, 148, 112279. [Google Scholar] [CrossRef]
- Carvalheiro, F.; Duarte, L.C.; Medeiros, R.; Gírio, F.M. Xylitol production by Debaryomyces hansenii CCMI 941 in brewery’s spent grain dilute-acid hydrolysate: Effect of supplementation. Biotechnol. Lett. 2007, 29, 1887–1891. [Google Scholar] [CrossRef] [PubMed]
- Deshavath, N.N.; Mohan, M.; Veeranki, V.D.; Goud, V.V.; Pinnamaneni, S.R.; Benarjee, T. Dilute acid pretreatment of sorghum biomass to maximize the hemicellulose hydrolysis with minimized levels of fermentative inhibitors for bioethanol production. 3Biotech 2017, 7, 139. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Roberto, I.C. Alternatives for detoxification of diluted-acid lignocellulosic hydrolysates for use in fermentative processes: A review. Bioresour. Technol. 2004, 93, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brás, T.; Guerra, V.; Torrado, I.; Lourenço, T.; Carvalheiro, F.; Duarte, L.C.; Neves, L.A. Detoxification of hemicellulosic hydrolysates from extracted olive pomace by diananofiltration. Proc. Biochem. 2014, 49, 173–180. [Google Scholar] [CrossRef]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass: Volume 1-Results of Screening for Potential Candidates from Sugars and Synthesis Gas; Report TP-510-35523; National Renewable Energy Lab. (NREL): Golden, CO, USA, 2004. [Google Scholar]
- Lim, K.L.; Wong, W.Y.; James Rubinsin, N.; Loh, S.K.; Lim, M.T. Techno-economic analysis of an integrated bio-refinery for the production of biofuels and value-added chemicals from oil palm empty fruit bunches. Processes 2022, 10, 1965. [Google Scholar] [CrossRef]
- Available online: https://www.grandviewresearch.com/industry-analysis/xylitol-market (accessed on 18 September 2023).
- Nobre, A.; Duarte, L.C.; Roseiro, J.C.; Girio, F.M. A physiological and enzymatic study of Debaryomyces hansenii growth on xylose- and oxygen-limited chemostats. Appl. Microb. Biotechnol. 2002, 59, 509–516. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Moncada, J.; Roberto, I.C.; Cardona, C.A. Techno-economic analysis for brewer’s spent grains use on a biorefinery concept: The Brazilian case. Bioresour. Technol. 2013, 148, 302–310. [Google Scholar] [CrossRef]
- Carvalheiro, F.; Duarte, L.C.; Lopes, S.; Parajó, J.C.; Pereira, H.; Gírio, F.M. Evaluation of the detoxification of brewery’s spent grain hydrolysate for xylitol production by Debaryomyces hansenii CCMI 941. Proc. Biochem. 2005, 40, 1215–1223. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Oxid. Antioxid. 1999, 299 Pt A, 152–178. [Google Scholar]
- Niaounakis, M.; Halvadakis, C.P. Olive Processing Waste Management. Literature Review and Patent Survey, 2nd ed.; Elsevier Publisher: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Martínez-Patiño, J.C.; Gómez-Cruz, I.; Romero, I.; Gullón, B.; Ruiz, E.; Brnčićc, M.; Castro, E. Ultrasound-assisted extraction as a first step in a biorefinery strategy for valorisation of extracted olive pomace. Energies 2019, 12, 2679. [Google Scholar] [CrossRef]
- Guo, X.; Cavka, A.; Jönsson, L.J.; Hong, F. Comparison of methods for detoxification of spruce hydrolysate for bacterial cellulose production. Microb. Cell Fact. 2013, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Chandel, A.K.; Kapoor, R.K.; Singh, A.; Kuhad, R.C. Detoxification of sugarcane bagasse hydrolysate improves ethanol production by Candida shehatae NCIM 3501. Bioresour Technol. 2007, 98, 1947–1950. [Google Scholar] [CrossRef]
- Carvalheiro, F.; Duarte, L.C.; Lopes, S.; Parajó, J.C.; Pereira, H.; Gírio, F.M. Supplementation requirements of brewery’s spent grain hydrolysate for biomass and xylitol production by Debaryomyces hansenii CCMI 941. J. Ind. Microbiol. Biotechnol. 2006, 33, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Preziosi-Belloy, L.; Nolleau, V.; Navarro, J.M. Xylitol production from aspenwood hemicellulose hydrolysate by Candida guilliermondii. Biotechnol. Lett. 2000, 22, 239–243. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Santos, J.C.; Roberto, I.C. Effect of pH and activated charcoal adsorption on hemicellulosic hydrolysate detoxification for xylitol production. J. Chem. Technol. Biotechnol. 2004, 79, 590–596. [Google Scholar] [CrossRef]
- Torrado, I. Optimization of Dilute Acid Hydrolysis for the Selective Fractionation of Sorghum Straw Hemicelluloses: Study of the Bioconversion of Hydrolysates to Produce Xylitol. Master’s Thesis, University of Algarve, Algarve, Portugal, 2010. [Google Scholar]
- Villarreal, M.L.M.; Prata, A.M.R.; Felipe, M.G.A.; Almeida e Silva, J.B. Detoxification procedures of eucalyptus hemicellulose hydrolysate for xylitol production by Candida guilliermondii. Enz. Microb. Technol. 2006, 40, 17–24. [Google Scholar] [CrossRef]
- Eryasar-Orer, K.; Karasu-Yalcin, S. Optimization of activated charcoal detoxification and concentration of chestnut shell hydrolysate for xylitol production. Biotechnol. Lett. 2021, 43, 1195–1209. [Google Scholar] [CrossRef]
- Martin, J.F.G.; Cuevas, M.; Bravo, V.; Sanchez, S. Ethanol production from olive prunings by autohydrolysis and fermentation with Candida tropicalis. Ren. Energ. 2010, 35, 1602–1608. [Google Scholar] [CrossRef]
- Cuevas, M.; Sanchez, S.; Bravo, V.; Cruz, N.; Garcia, J.F. Fermentation of enzymatic hydrolysates from olive stones by Pachysolen tannophilus. J. Chem. Technol. Biotechnol. 2009, 84, 461–467. [Google Scholar] [CrossRef]
- López-Linares, J.C.; Ruiz, E.; Romero, I.; Castro, E.; Manzanares, P. Xylitol production from exhausted olive pomace by Candida boidinii. Appl. Sci. 2020, 10, 6966. [Google Scholar] [CrossRef]
- Wannawilai, S.; Chisti, Y.; Sirisansaneeyakul, S. A model of furfural-inhibited growth and xylitol production by Candida magnoliae TISTR 5663. Food Bioprod. Proc. 2017, 105, 129–140. [Google Scholar] [CrossRef]
- Salgado, J.M.; Rodríguez, N.; Cortés, S.; Domínguez, J.M. Effect of nutrient supplementation of crude or detoxified concentrated distilled grape marc hemicellulosic hydrolysates on the xylitol production by Debaryomyces hansenii. Prep Biochem Biotechnol. 2012, 42, 1–14. [Google Scholar] [CrossRef]
- Diz, J.; Cruz, J.M.; Dominguez, H.; Parajó, J.C. Xylitol production from Eucalyptus wood hydrolysates in low-cost fermentation media. Food Technol. Biotechnol. 2002, 40, 191–197. [Google Scholar]
- López-Linares, J.C.; Romero, I.; Cara, C.; Castro, E.; Mussatto, S.I. Xylitol production by Debaryomyces hansenii and Candida guilliermondii from rapeseed straw hemicellulosic hydrolysate. Bioresour. Technol. 2017, 247, 736–743. [Google Scholar] [CrossRef]
 ); xylitol (
); xylitol ( ); arabinose (
); arabinose ( ); glucose (
); glucose ( ); acetic acid (×); cell density (
); acetic acid (×); cell density ( ).
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().
).
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().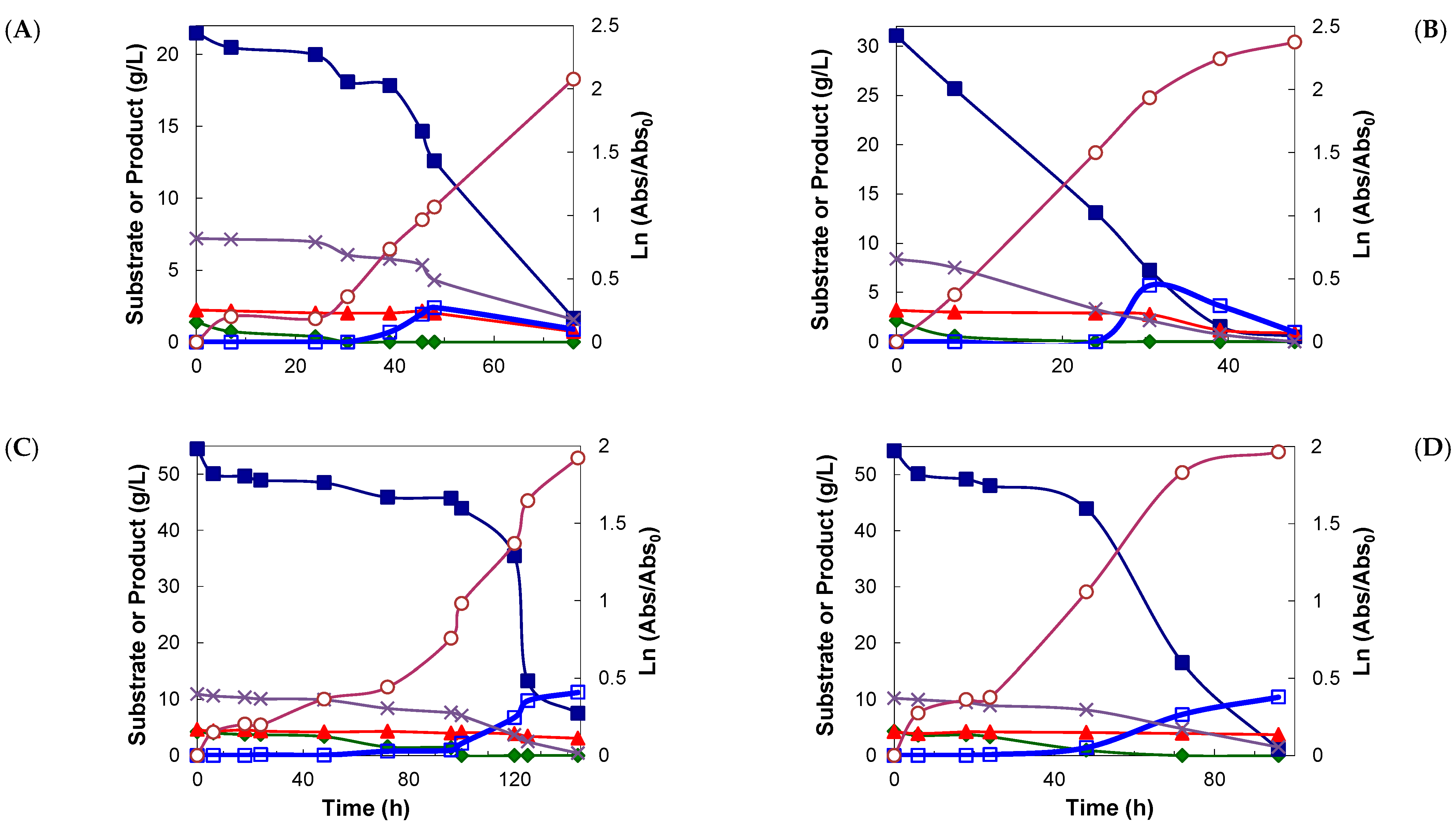 ); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density (). ); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().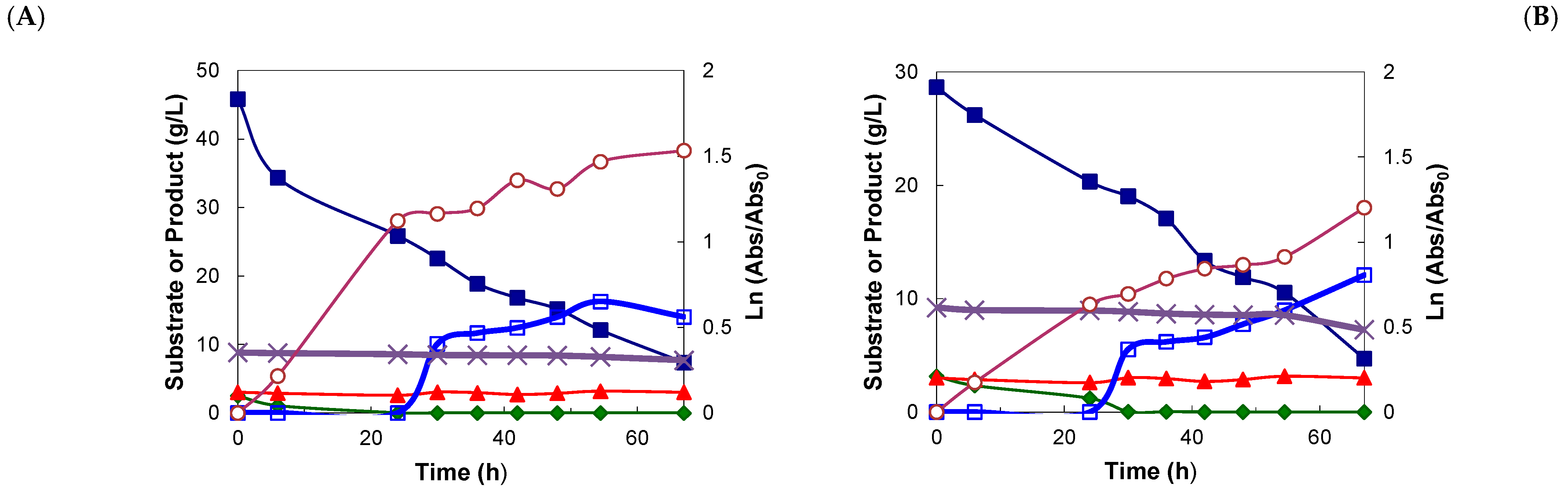 ); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().
); xylitol (); arabinose (); glucose (); acetic acid (×); cell density ().

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Concentration a (g/L) |
|---|---|
| Xylose | 32.6 |
| Arabinose | 1.98 |
| Glucose | 2.30 |
| Acetic acid | 9.10 |
| HMF | 0.06 |
| Furfural | 2.66 |
| Phenolic compounds | 5.02 |
| Detoxification Treatment | Monosac. a | Acetic Acid | HMF | Furfural | Phenolics b |
|---|---|---|---|---|---|
| None | 100 | 100 | 100 | 100 | 100 |
| pH 5.5 | 93 | 95 | 74 | 59 | 94 |
| Concentration (1.5×) | 87 | 87 | 93 | 5 | 84 |
| Concentration (2×) | 88 | 83 | 84 | 0 | 90 |
| Activated charcoal | 99 | 89 | 38 | 13 | 27 |
| Activated charcoal + conc. (2×) | 99 | 54 | 27 | 44 | 52 |
| Anionic resins | 99 | 99 | 74 | 37 | 41 |
| Anionic resins + conc. (2×) | 100 | 72 | 47 | 0 | 78 |
| Nanofiltration | 90 | 46 | 59 | 0 | 33 |
| Nanofiltration + conc. (2×) | 84 | 41 | 69 | 0 | 83 |
| Hydrolysate Treatment | pH | Xyl | Ara | Glc | Acetic Acid | Furfural | HMF | Phenolics |
|---|---|---|---|---|---|---|---|---|
| Control | 5.5 | 23.5 | 2.54 | 1.60 | 7.75 | 1.85 | 0.05 | 4.11 |
| Concentration (1.5×) | 5.5 | 32.6 | 3.52 | 2.31 | 9.33 | 0.15 | 0.05 | 3.45 |
| Concentration (2×) | 5.5 | 56.9 | 4.94 | 4.30 | 11.9 | n.d | 0.06 | 3.87 |
| Concentration (2×) | 6.5 | 56.1 | 4.92 | 4.28 | 10.9 | n.d | 0.05 | 3.90 |
| Concentration (2×) | 7.5 | 55.6 | 4.91 | 4.15 | 10.7 | n.d | 0.05 | 4.02 |
| Activated charcoal + conc. (2×) | 5.5 a | 40.9 | 4.71 | 4.69 | 9.80 | n.d | 0.02 | 1.90 |
| Activated charcoal + conc. (2×) | 6.5 a | 39.8 | 4.62 | 4.69 | 9.78 | n.d | 0.02 | 2.10 |
| Activated charcoal + conc. (2×) | 7.5 a | 39.4 | 4.39 | 4.48 | 9.76 | n.d | 0.02 | 3.00 |
| Anionic resins + conc. (2×) | 5.5 | 42.6 | 3.50 | 2.70 | 9.33 | n.d | 0.03 | 4.20 |
| Anionic resins + conc. (2×) | 6.5 a | 31.8 | 3.24 | 3.24 | 10.3 | n.d | 0.02 | 4.70 |
| Nanofiltration + conc. (2×) | 5.5 | 51.7 | 5.23 | 3.97 | 6.18 | n.d | 0.05 | 3.50 |
| Detoxification Treatment | pH | Oxygen Availability | Supplements | X0 (g/L) | Xyl0 (g/L) | Xyl (%) | YXOH (g/g) | QXOH (g/L/h) | Qx (g/L/h) |
|---|---|---|---|---|---|---|---|---|---|
| Control | 5.5 | High | TEVM | 0.9 | 21.5 | 78.0 | 0.17 | 0.06 | 0.15 |
| Evaporation (1.5×) | 5.5 | High | TEVM | 1.6 | 31.0 | 98.4 | 0.19 | 0.19 | 0.39 |
| Evaporation (2×) | 5.5 | High | TEVM | 2.5 | 54.8 | - | - | - | - |
| Evaporation (2×) | 6.5 | High | TEVM | 1.0 | 54.5 | 86.3 | 0.24 | 0.08 | 0.15 |
| Evaporation (2×) | 7.5 | High | TEVM | 1.8 | 54.3 | 98.0 | 0.20 | 0.13 | 0.15 |
| Activated charcoal | 5.5 | High | TEVM | 3.2 | 39.1 | 69.6 | 0.21 | 0.13 | 0.18 |
| Activated charcoal | 6.5 | High | TEVM | 4.6 | 38.6 | 92.2 | 0.17 | 0.10 | 0.28 |
| Activated charcoal | 6.5 | Low | TEVM | 4.2 | 36.8 | 99.9 | 0.22 | 0.13 | 0.25 |
| Activated charcoal | 7.5 | High | TEVM | 2.6 | 36.8 | 83.2 | 0.16 | 0.11 | 0.27 |
| Anionic resins | 5.5 | Low | TEVM | 3.2 | 45.8 | 83.9 | 0.42 | 0.30 | 0.09 |
| Anionic resins | 6.5 | Low | TEVM | 2.3 | 28.7 | 83.6 | 0.45 | 0.20 | 0.12 |
| Nanofiltration | 5.5 | Low | TEVM | 2.2 | 48.8 | 90.8 | 0.39 | 0.14 | 0.07 |
| Nanofiltration | 5.5 | Low | YE | 2.2 | 46.4 | 92.9 | 0.55 | 0.17 | 0.04 |
| Nanofiltration | 5.5 | Low | CSL | 2.1 | 49.9 | 92.5 | 0.57 | 0.19 | 0.06 |
| Nanofiltration | 5.5 | Low | BSGE | 1.5 | 50.7 | 92.7 | 0.47 | 0.16 | 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morais, A.R.C.; Duarte, L.C.; Lourenço, P.; Torrado, I.; Brás, T.; Neves, L.A.; Carvalheiro, F. Xylitol Production by Debaryomyces hansenii in Extracted Olive Pomace Dilute-Acid Hydrolysate. Fermentation 2023, 9, 1020. https://doi.org/10.3390/fermentation9121020
Morais ARC, Duarte LC, Lourenço P, Torrado I, Brás T, Neves LA, Carvalheiro F. Xylitol Production by Debaryomyces hansenii in Extracted Olive Pomace Dilute-Acid Hydrolysate. Fermentation. 2023; 9(12):1020. https://doi.org/10.3390/fermentation9121020
Chicago/Turabian StyleMorais, Ana Rita C., Luís C. Duarte, Pedro Lourenço, Ivone Torrado, Teresa Brás, Luísa A. Neves, and Florbela Carvalheiro. 2023. "Xylitol Production by Debaryomyces hansenii in Extracted Olive Pomace Dilute-Acid Hydrolysate" Fermentation 9, no. 12: 1020. https://doi.org/10.3390/fermentation9121020
APA StyleMorais, A. R. C., Duarte, L. C., Lourenço, P., Torrado, I., Brás, T., Neves, L. A., & Carvalheiro, F. (2023). Xylitol Production by Debaryomyces hansenii in Extracted Olive Pomace Dilute-Acid Hydrolysate. Fermentation, 9(12), 1020. https://doi.org/10.3390/fermentation9121020