

Single and Co-Cultures of Proteolytic Lactic Acid Bacteria in the Manufacture of Fermented Milk with High ACE Inhibitory and Antioxidant Activities

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Bacterial Strains
2.2. Preparation of Fermented Milk Samples and Crude Peptide Extracts
2.3. Determination of Proteolysis and Peptide Content
2.4. Determination of ACE Inhibitory Activity
2.5. Determination of Antioxidant Activities
2.5.1. DPPH Scavenging Activity
2.5.2. Ferric-Reducing Antioxidant Power (FRAP)
2.5.3. OH-Radical Scavenging Activity
2.5.4. Total Antioxidant Capacity (Ammonium phosphomolybdate Assay)
2.6. Statistical Analysis
3. Results and Discussion
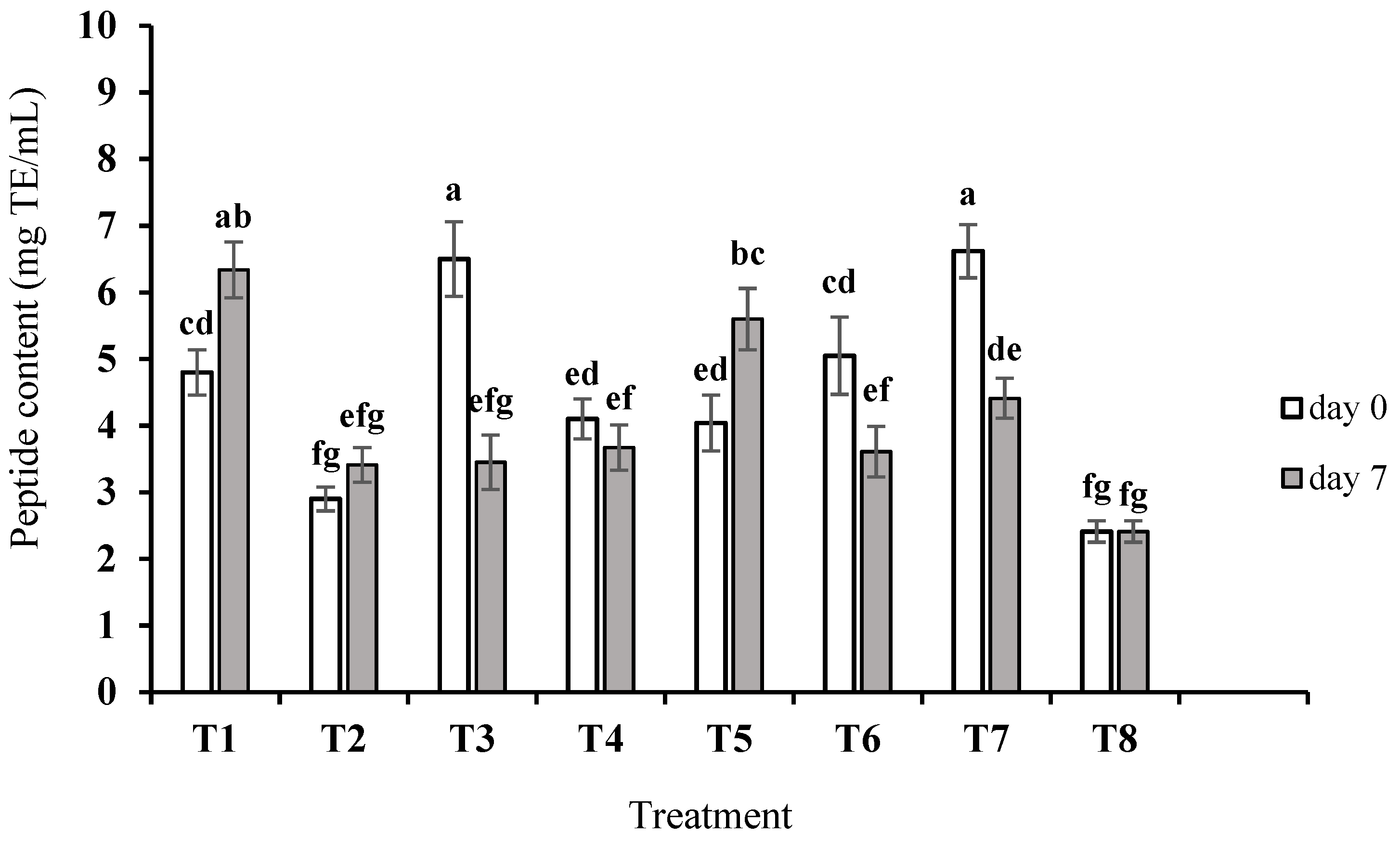
3.1. Proteolysis and Peptide Content in Crude Peptide Extract
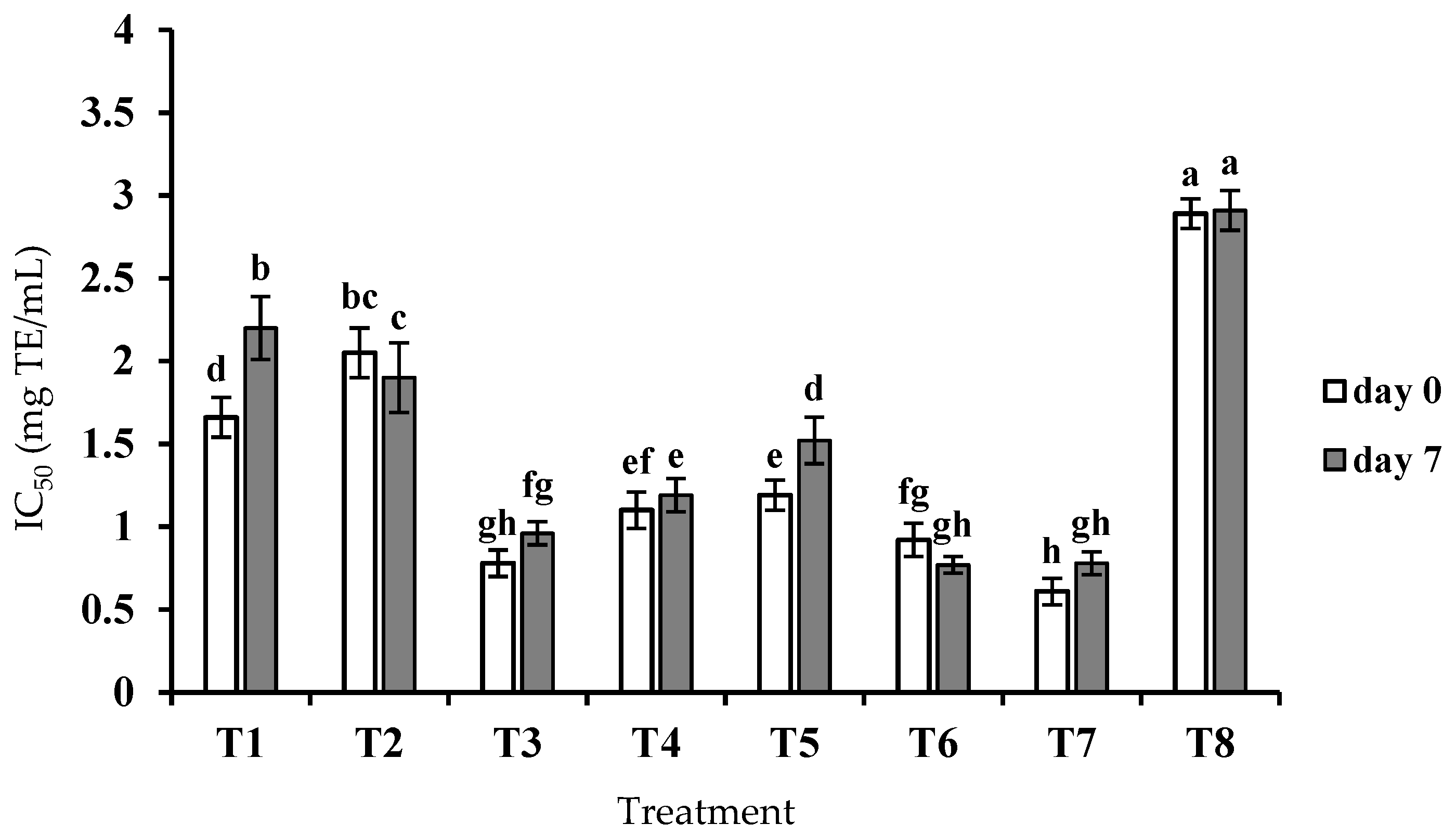
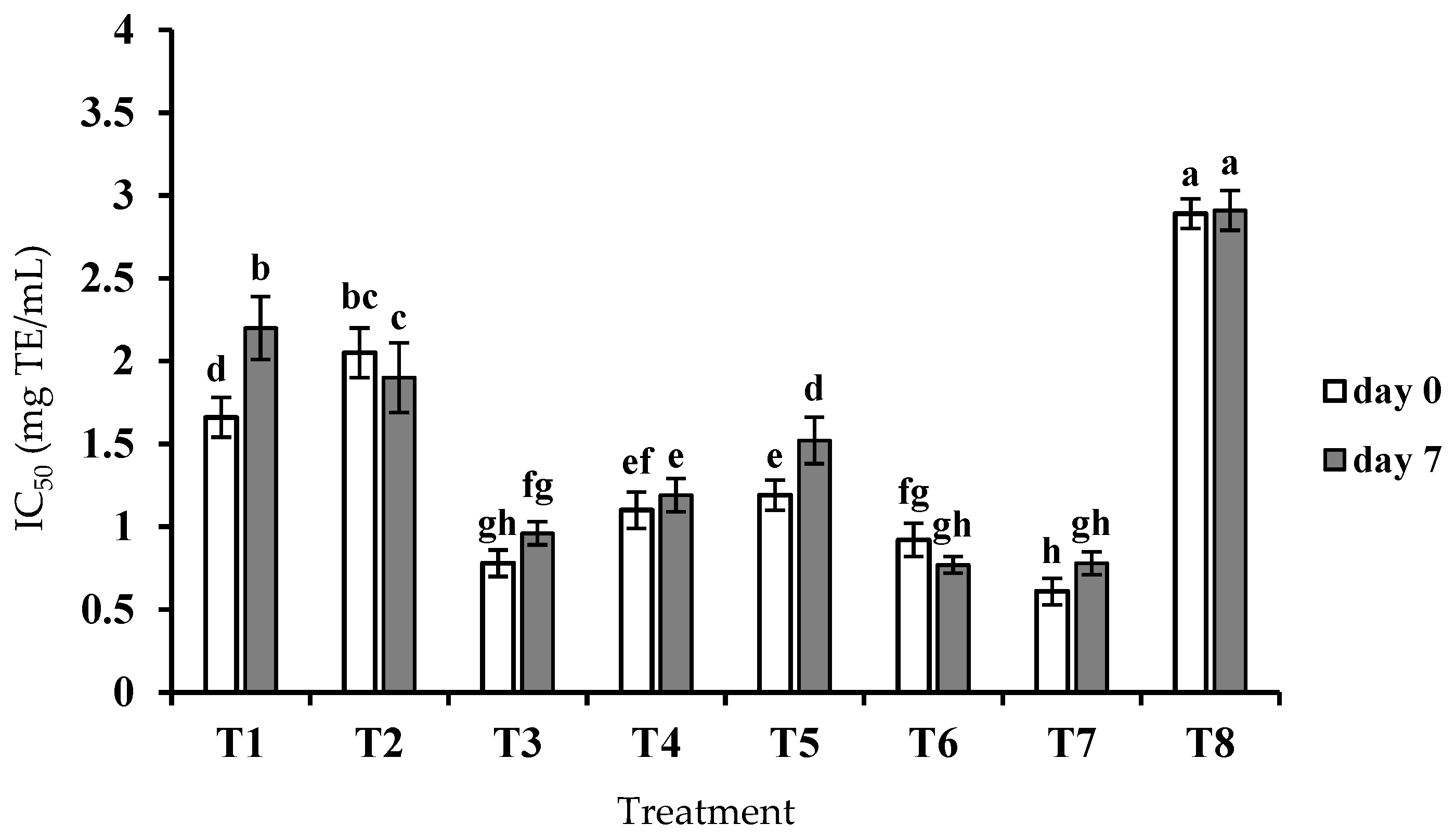
3.2. ACE Inhibitory Activity of Crude Peptide Extract
3.3. Antioxidant Activity
3.3.1. DPPH Radical Inhibitory Activity
3.3.2. Ferric-Reducing Antioxidant Power (FRAP)
3.3.3. Hydroxyl Radical Scavenging Activity
3.3.4. Total Antioxidant Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. A Global Brief on Hypertension: Silent Killer, Global Public Health Crisis; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- Khan, M.D.H.; Imig, J.D. Antihypertensive Drugs. Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–5. [Google Scholar]
- O’Keeffe, M.B.; Conesa, C.; FitzGerald, R.J. Identification of angiotensin converting enzyme inhibitory and antioxidant peptides in a whey protein concentrate hydrolysate produced at semi-pilot scale. Int. J. Food Sci. 2017, 52, 1751–1759. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Sorriento, D.; Luca, N.D.; Trimarco, B.; Iaccarino, G. The Antioxidant Therapy: New Insights in the Treatment of Hypertension. Front. Physiol. 2018, 9, 258. [Google Scholar] [CrossRef]
- Sah, B.N.P.; Vasiljevic, T.; McKechnie, S.; Donkor, O.N. Identification of anticancer peptides from bovine milk proteins and their potential roles in management of cancer: A critical review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 123–138. [Google Scholar] [CrossRef]
- Villaverde, P.; Lajous, M.; MacDonald, C.J.; Fagherazzi, G.; Bonnet, F.; Boutron-Ruault, M.C. High dietary total antioxidant capacity is associated with a reduced risk of hypertension in French women. Nutr. J. 2019, 18, 31. [Google Scholar] [CrossRef]
- Zhou, D.D.; Luo, M.; Shang, A.; Mao, Q.Q.; Li, B.Y.; Gan, R.Y.; Li, H.B. Antioxidant Food Components for the Prevention and Treatment of Cardiovascular Diseases: Effects, Mechanisms, and Clinical Studies. Oxid. Med. Cell. Longev. 2021, 6627355. [Google Scholar] [CrossRef] [PubMed]
- Garavand, F.; Daly, D.F.; Gómez-Mascaraque, L.G. Biofunctional, structural, and tribological attributes of GABA-enriched probiotic yoghurts containing Lacticaseibacillus paracasei alone or in combination with prebiotics. Int. Dairy J. 2022, 129, 105348. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai1, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Iwamoto, H.; Matsubara, T.; Okamoto, T.; Matsumoto, T.; Yoshikawa, M.; Takeda, Y. Ingestion of casein hydrolysate induces oral tolerance and suppresses subsequent epicutaneous sensitization and development of anaphylaxis reaction to casein in mice. Int. Arch. Allergy Imm. 2019, 179, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Elfahri, K.R.; Vasiljevic, T.; Yeager, T.; Donkor, O.N. Anti-colon cancer and antioxidant activities of bovine skim milk fermented by selected Lactobacillus helveticus strains. J. Dairy Sci. 2016, 99, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Ayyash, M.; Al-Dhaheri, A.S.; Al Mahadin, S.; Kizhakkayil, J.; Abushelaibi, A. In vitro investigation of anticancer, antihypertensive, antidiabetic, and antioxidant activities of camel milk fermented with camel milk probiotic: A comparative study with fermented bovine milk. J. Dairy Sci. 2018, 101, 900–911. [Google Scholar] [CrossRef]
- Garavand, F.; Daly, D.F.; Gómez-Mascaraque, L.G. The consequence of supplementing with synbiotic systems on free amino acids, free fatty acids, organic acids, and some stability indexes of fermented milk. Int. Dairy J. 2022, 105477. [Google Scholar] [CrossRef]
- Begunova, A.V.; Savinova, O.S.; Glazunova, O.A.; Moiseenko, K.V.; Rozhkova, I.V.; Fedorova, T.V. Development of Antioxidant and Antihypertensive Properties during Growth of Lactobacillus helveticus, Lactobacillus rhamnosus and Lactobacillus reuteri on Cow’s Milk: Fermentation and Peptidomics Study. Foods 2021, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Li, S.N.; Tang, S.H.; He, Q.; Hu, J.X.; Zheng, J. In vitro antioxidant and angiotensin-converting enzyme inhibitory activity of fermented milk with different culture combinations. J. Dairy Sci. 2020, 103, 1120–1130. [Google Scholar] [CrossRef]
- Șanli, T.; Akal, H.C.; Yetisemiyen, A.; HayaLoglu, A.A. Influence of adjunct cultures on angiotensin-converting enzyme (ACE)-inhibitory activity, organic acid content and peptide profile of kefir. Int. J. Dairy Technol. 2018, 71, 131–139. [Google Scholar] [CrossRef]
- Wang, S.Y.; Huang, R.F.; Ng, K.S.; Chen, Y.P.; Shiu, J.S.; Chen, M.J. Co-Culture Strategy of Lactobacillus kefiranofaciens HL1 for Developing Functional Fermented Milk. Foods 2021, 10, 2098. [Google Scholar] [CrossRef]
- Sharma, S. Antivirulence activities of Bioactive Peptides Produced by Lactobacillus helveticus and Lactobacillus acidophilus against Salmonella enterica Serovar Typhimurium. Master’s Thesis, University of Guelph, Guelph, ON, Canada, 2014. [Google Scholar]
- Tulini, F.L.; Hymery, N.; Choiset, Y.; Chobert, J.M.; Haertlé, T.; De Martinis, E.C.P.; Le Blay, G. Milk fermented with the probiotic candidate Lactobacillus paracasei FT700 induces differentiation of monocytes toward macrophages in vitro. J. Funct Foods 2015, 15, 533–540. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Losito, I.; Gobbetti, M.; Carbonara, T.; De Bari, M.D.; Zambonin, P.G. Antibacterial Activities of Peptides from the Water-Soluble Extracts of Italian Cheese Varieties. J. Dairy Sci. 2005, 88, 2348–2360. [Google Scholar] [CrossRef]
- Donkor, O.N.; Henriksson, A.; Vasiljevic, T.; Shah, N.P. Probiotic strains as starter cultures improve angiotensin-converting enzyme inhibitory activity in soy yogurt. J. Food Sci. 2005, 70, 375–381. [Google Scholar] [CrossRef]
- Jemil, I.; Jridi, M.; Nasri, R.; Ktari, N.; Slama-Ben Salem, R.B.; Mehiri, M.; Hajji, M.; Nasri, M. Functional, antioxidant and antibacterial properties of protein hydrolysates prepared from fish meat fermented by Bacillus subtilis A26. Process Biochem. 2014, 49, 963–972. [Google Scholar] [CrossRef]
- Yildirim, A.; Mavi, A.; Kara, A.A. Determination of antioxidant and antimicrobial activities of Rumex crispus L. extracts. J. Agric. Food Chem. 2001, 49, 4083–4089. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ding, L.; Wang, Y.; Zhang, Y.; Liu, J. Isolation and Characterisation of in Vitro and Cellular Free Radical Scavenging Peptides from Corn Peptide Fractions. Molecules 2015, 20, 3221–3237. [Google Scholar] [CrossRef] [PubMed]
- Meshginfar, N.; Sadeghi Mahoonak, A.; Hosseinian, F.; Ghorbani, M.; Tsopmo, A. Production of antioxidant peptide fractions from a by-product of tomato processing: Mass spectrometry identification of peptides and stability to gastrointestinal digestion. JFST 2018, 55, 3498–3507. [Google Scholar] [CrossRef]
- Rasika, D.M.D.; Ueda, T.; Jayakody, L.N.; Suriyagoda, L.D.B.; Silva, K.F.S.T.; Ando, S.; Vidanarachchi, J.K. ACE-inhibitory activity of milk fermented with Saccharomyces cerevisiae K7 and Lactococcus lactis subsp. lactis NBRC 12007. J. Natl. Sci. Found. 2015, 43, 141–151. [Google Scholar] [CrossRef]
- Nielsen, M.S.; Martinussen, T.; Flambard, B.; Sørensen, K.I.; Otte, J. Peptide profiles and angiotensin-I-converting enzyme inhibitory activity of fermented milk products: Effect of bacterial strain, fermentation pH, and storage time. Int. Dairy J. 2009, 19, 155–165. [Google Scholar] [CrossRef]
- Shori, A.B. Antioxidant activity and viability of lactic acid bacteria in soybean-yogurt made from cow and camel milk. J. Taibah Univ. Sci. 2013, 7, 202–208. [Google Scholar] [CrossRef]
- Pihlanto, A.; Virtanen, T.; Korhonen, H. Angiotensin I converting enzyme (ACE) inhibitory activity and antihypertensive effect of fermented milk. Int. Dairy J. 2010, 20, 3–10. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Okubo, A.; Yamazaki, S.; Takano, T. Purification and characterization of angiotensin I-converting enzyme inhibitors from sour milk. J. Dairy Sci. 1995, 78, 777–783. [Google Scholar] [CrossRef]
- Rodríguez-Gómez, F.; Romero-Gil, V.; Bautista-Gallego, J.; Garrido-Fernández, A.; Arroyo-López, F.N. Multivariate analysis to discriminate yeast strains with technological applications in table olive processing. World J. Microbiol. Biotechnol. 2012, 28, 1761–1770. [Google Scholar] [CrossRef]
- Meira, S.M.M.; Daroit, D.J.; Helfer, V.E.; Corrêa, A.P.F.; Segalin, J.; Carro, S.; Brandelli, A. Bioactive peptides in water-soluble extracts of ovine cheeses from Southern Brazil and Uruguay. Food Res. Int. 2012, 48, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Habibi Najafi, M.B.; Fatemi, S.; Tavakkoli Fadiheh, M. Release of proteolysis products with ACE-inhibitory and antioxidant activities in probiotic yogurt containing different levels of fat and prebiotics. Int. J. Pept. Res. Ther. 2019, 25, 367–377. [Google Scholar] [CrossRef]
- Donkor, O.N.; Henriksson, A.; Vasiljevic, T.; Shah, N.P. Proteolytic activity of dairy lactic acid bacteria and probiotics as determinant of growth and in vitro angiotensin-converting enzyme inhibitory activity in fermented milk. Lait 2007, 87, 21–38. [Google Scholar] [CrossRef]
- Shori, A.B.; Rashid, F.; Baba, A.S. Effect of the addition of phytomix-3+ mangosteen on antioxidant activity, viability of lactic acid bacteria, type 2 diabetes key-enzymes, and sensory evaluation of yogurt. LWT 2018, 94, 33–39. [Google Scholar] [CrossRef]
- Qian, B.; Xing, M.; Cui, L.; Deng, Y.; Xu, Y.; Huang, M.; Zhang, S. Antioxidant, antihypertensive, and immunomodulatory activities of peptide fractions from fermented skim milk with Lactobacillus delbrueckii ssp. bulgaricus LB340. J. Dairy Res. 2011, 78, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Kariyawasam, K.M.G.M.M.; Lee, N.K.; Paik, H.D. Synbiotic yoghurt supplemented with novel probiotic Lactobacillus brevis KU200019 and fructooligosaccharides. Food Biosci. 2021, 39, 100835. [Google Scholar] [CrossRef]
- Corrêa, A.P.F.; Daroit, D.J.; Coelho, J.; Meira, S.M.M.; Lopes, F.C.; Segalin, J.; Risso, P.H.; Brandelli, A. Antioxidant, antihypertensive and antimicrobial properties of ovine milk caseinate hydrolyzed with a microbial protease. J. Sci. Food Agric. 2011, 91, 2247–2254. [Google Scholar] [CrossRef]
- Caleja, C.; Barros, L.; Antonio, A.L.; Carocho, M.; Oliveira, M.B.P.P.; Ferreira, I.C.F.R. Fortification of yogurts with different antioxidant preservatives: A comparative study between natural and synthetic additives. Food Chem. 2016, 210, 262–268. [Google Scholar] [CrossRef]
- Biadała, A.; Adzahan, N.M. Storage Stability of Antioxidant in Milk Products Fermented with Selected Kefir Grain Microflora. Molecules 2021, 26, 3307. [Google Scholar] [CrossRef]
- Tyagi, A.; Shabbir, U.; Chelliah, R.; Daliri, E.B.M.; Chen, X.; Oh, D.H. Limosilactobacillus reuteri Fermented Brown Rice: A Product with Enhanced Bioactive Compounds and Antioxidant Potential. Antioxidants 2021, 10, 1077. [Google Scholar] [CrossRef]
- Panchal, G.; Hati, S.; Sakure, A. Characterization and production of novel antioxidative peptides derived from fermented goat milk by L. fermentum. LWT 2020, 119, 108887. [Google Scholar] [CrossRef]
- Shu, G.; Shi, X.; Chen, L.; Kou, J.; Meng, J.; Chen, H. Antioxidant peptides from goat milk fermented by Lactobacillus casei L61: Preparation, optimization, and stability evaluation in simulated gastrointestinal fluid. Nutrients 2018, 10, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Concentration (mg/mL) | ||||
|---|---|---|---|---|
| Treatment | Time (Day) | 20 | 40 | 80 |
| T1 | 0 | 40.65 ± 3.72 t | 63.90 ± 1.06 no | 77.15 ± 1.77 ghij |
| 7 | 37.19 ± 1.42 u | 66.1 ± 1.24 lmn | 79.93 ± 0.65 efg | |
| T2 | 0 | 13.19 ± 2.56 x | 47.83 ± 0.99 r | 64.37 ± 0.52 no |
| 7 | 9.71 ± 0.89 x | 62.93 ± 0.60 no | 70.41 ± 1.27 k | |
| T3 | 0 | 61.22 ± 0.71 op | 78.41 ± 0.30 fghi | 87.01 ± 2.40 b |
| 7 | 49.02 ± 0.79 r | 58.14 ± 1.12 pq | 82.94 ± 2.27 cde | |
| T4 | 0 | 47.97 ± 1.89 r | 64.64 ± 0.75 no | 77.65 ± 1.37 fghij |
| 7 | 31.50 ± 4.16 v | 75.13 ± 0.61 ij | 84.15 ± 2.84 bcd | |
| T5 | 0 | 43.01 ± 1.76 st | 69.10 ± 0.70 kl | 81.06 ± 0.51 def |
| 7 | 45.67 ± 1.56 rs | 68.31 ± 1.29 klm | 82.61 ± 1.70 cde | |
| T6 | 0 | 55.53 ± 1.59 q | 70.17 ± 1.41 k | 84.80 ± 3.57 bc |
| 7 | 48.29 ± 1.68 r | 76.11 ± 0.63 hij | 76.44 ± 2.02 ghij | |
| T7 | 0 | 47.66 ± 4.61 r | 74.24 ± 0.83 j | 92.68 ± 3.60 a |
| 7 | 56.95 ± 0.86 q | 65.53 ± 1.55 mn | 79.67 ± 0.71 efgh | |
| T8 | 0 | 11.87 ± 2.74 x | 24.05 ± 3.10 w | 48.11 ± 1.18 r |
| 7 | 11.87 ± 2.74 x | 24.05 ± 3.10 w | 48.11 ± 1.18 r | |
| Concentration (mg/mL) | ||||
|---|---|---|---|---|
| Treatment | Time (Day) | 20 | 40 | 80 |
| T1 | 0 | 43.80 ± 2.78 u | 75.57 ± 0.77 n | 91.50 ± 1.03 cde |
| 7 | 45.33 ± 1.85 u | 80.70 ± 1.20 lm | 94.43 ± 0.68 ab | |
| T2 | 0 | 31.70 ± 1.84 v | 75.87 ± 0.07 n | 93.50 ± 1.03 abc |
| 7 | 51.50 ± 0.25 rs | 79.40 ± 1.54 m | 95.47 ± 0.88 a | |
| T3 | 0 | 52.20 ± 0.70 qr | 84.70 ± 0.70 hij | 91.40 ± 0.72 cde |
| 7 | 60.97 ± 1.05 o | 87.60 ± 0.73 fg | 93.53 ± 1.11 ab | |
| T4 | 0 | 47.80 ± 3.48 t | 80.43 ± 0.82 lm | 89.27 ± 1.86 ef |
| 7 | 50.60 ± 1.20 rs | 85.50 ± 1.20 ghi | 93.60 ± 1.35 abc | |
| T5 | 0 | 44.60 ± 2.09 u | 79.57 ± 2.05 m | 90.30 ± 0.00 de |
| 7 | 56.10 ± 2.79 p | 81.23 ± 1.96 klm | 93.40 ± 0.00 abc | |
| T6 | 0 | 47.80 ± 0.70 t | 81.50 ± 0.70 klm | 92.53 ± 1.10bcd |
| 7 | 49.60 ± 0.46 st | 86.73 ± 1.20 ghi | 93.37 ± 0.62 abc | |
| T7 | 0 | 47.80 ± 1.84 t | 82.47 ± 0.91 jkl | 90.30 ± 0.00 de |
| 7 | 53.90 ± 0.56 q | 83.43 ± 1.95 ijk | 94.43 ± 0.68 ab | |
| T8 | 0 | 13.30 ± 1.3 y | 13.50 ± 0.70 xy | 15.60 ± 0.6 wx |
| 7 | 12.40 ± 0.4 y | 16.25 ± 1.25 w | 16.20 ± 1.2 w | |
| Concentration (mg/mL) | ||||
|---|---|---|---|---|
| Treatment | Time (Day) | 20 | 40 | 80 |
| T1 | 0 | 0.29 ± 0.01p | 0.41 ± 0.01 lmno | 0.57 ± 0.01 ghij |
| 7 | 0.53 ± 0.00 hijk | 0.62 ± 0.19 gh | 0.86 ± 0.04 bcd | |
| T2 | 0 | 0.39 ± 0.01 lmno | 0.54 ± 0.01 hijk | 0.89 ± 0.01 ab |
| 7 | 0.38 ± 0.00 mnop | 0.45 ± 0.12 klm | 0.86 ± 0.01 abcd | |
| T3 | 0 | 0.31 ± 0.01 op | 0.42 ± 0.01 lmn | 0.77 ± 0.01 de |
| 7 | 0.37 ± 0.02 mnop | 0.56 ± 0.04 ghij | 0.89 ± 0.01 ab | |
| T4 | 0 | 0.40 ± 0.00 lmno | 0.71 ± 0.11 ef | 0.96 ± 0.05 a |
| 7 | 0.57 ± 0.02 ghij | 0.86 ± 0.02 abcd | 0.96 ± 0.01 a | |
| T5 | 0 | 0.57 ± 0.02 ghij | 0.80 ± 0.02 bcde | 0.95 ± 0.05 a |
| 7 | 0.59 ± 0.01 ghi | 0.92 ± 0.02 a | 0.94 ± 0.01 a | |
| T6 | 0 | 0.37 ± 0.03 mnop | 0.48 ± 0.01 jkl | 0.65 ± 0.04 fg |
| 7 | 0.34 ± 0.02 nop | 0.52 ± 0.22 ijk | 0.88 ± 0.01 abc | |
| T7 | 0 | 0.34 ± 0.02 nop | 0.45 ± 0.02 klm | 0.79 ± 0.01 cde |
| 7 | 0.29 ± 0.01 p | 0.39 ± 0.02 lmno | 0.89 ± 0.01 ab | |
| T8 | 0 | 0.08 ± 0.01 q | 0.12 ± 0.01 q | 0.16 ± 0.01 q |
| 7 | 0.07 ± 0.02 q | 0.15 ± 0.01 q | 0.13 ± 0.02 q | |
| Concentration (mg/mL) | ||||
|---|---|---|---|---|
| Treatment | Time (Day) | 20 | 40 | 80 |
| T1 | 0 | 1.00 ± 0.87 tu | 46.50 ± 1.37 h | 90.03 ± 2.00 d |
| 7 | 3.90 ± 0.2 rs | 51.37 ± 2.49 g | 95.37 ± 0.94 ab | |
| T2 | 0 | 0.00 ± 0.00 u | 30.80 ± 1.78 k | 78.53 ± 3.37 f |
| 7 | 6.20 ± 1.36 qr | 39.37 ± 1.29 i | 95.43 ± 2.72 ab | |
| T3 | 0 | 2.67 ± 0.47 st | 27.10 ± 2.14 l | 92.30 ± 1.59 c |
| 7 | 3.97 ± 0.58 rs | 39.33 ± 1.08 i | 93.43 ± 0.36 bc | |
| T4 | 0 | 2.30 ± 0.00 stu | 31.20 ± 1.21k | 88.30 ± 1.03 de |
| 7 | 6.57 ± 1.08 q | 37.33 ± 0.10 ij | 93.57 ± 1.95 bc | |
| T5 | 0 | 1.60 ± 0.82 stu | 35.43 ± 1.95 j | 88.77 ± 1.53 de |
| 7 | 0.97 ± 0.88 tu | 21.83 ± 1.34 mn | 86.50 ± 1.03 e | |
| T6 | 0 | 0.00 ± 0.00 u | 21.10 ± 0.88 n | 87.70 ± 0.00 de |
| 7 | 0.67 ± 0.31tu | 23.50 ± 0.18 m | 88.37 ± 0.98 de | |
| T7 | 0 | 1.60 ± 0.94 qr | 44.73 ± 3.12 h | 93.23 ± 1.70 bc |
| 7 | 7.40 ± 1.26 pq | 50.83 ± 1.96 g | 96.37 ± 0.10 a | |
| T8 | 0 | 0.00 ± 0.00 u | 8.40 ± 0.10 pq | 11.40 ± 0.10 o |
| 7 | 0.60 ± 0.10 tu | 9.75 ± 0.10 op | 11.75 ± 0.10 o | |
| Concentration (mg/mL) | ||||
|---|---|---|---|---|
| Treatment | Time (Day) | 20 | 40 | 80 |
| T1 | 0 | 0.35 ± 0.01 q | 0.66 ± 0.01 gh | 0.86 ± 0.01 d |
| 7 | 0.26 ± 0.01 s | 0.58 ± 0.01 k | 0.85 ± 0.02 d | |
| T2 | 0 | 0.37 ± 0.01 pq | 0.61 ± 0.03 j | 0.90 ± 0.01 c |
| 7 | 0.42 ± 0.03 o | 0.63 ± 0.03 ij | 0.89 ± 0.02 c | |
| T3 | 0 | 0.42 ± 0.01 o | 0.65 ± 0.01 hi | 0.76 ± 0.02 f |
| 7 | 0.36 ± 0.00 pq | 0.45 ± 0.01 n | 0.68 ± 0.03 g | |
| T4 | 0 | 0.37 ± 0.03 pq | 0.61 ± 0.02 j | 0.96 ± 0.02 b |
| 7 | 0.32 ± 0.01 r | 0.41 ± 0.02 o | 0.89 ± 0.00 c | |
| T5 | 0 | 0.32 ± 0.02 r | 0.38 ± 0.01 p | 0.77 ± 0.01 ef |
| 7 | 0.31 ± 0.01r | 0.35 ± 0.01 q | 0.79 ± 0.00 e | |
| T6 | 0 | 0.37 ± 0.02 pq | 0.43 ± 0.01 no | 0.67 ± 0.03 gh |
| 7 | 0.37 ± 0.02 pq | 0.48 ± 0.00 m | 0.75 ± 0.01 f | |
| T7 | 0 | 0.36 ± 0.00 pq | 0.56 ± 0.01 l | 0.99 ± 0.01 a |
| 7 | 0.36 ± 0.00 pq | 0.45 ± 0.01 n | 0.68 ± 0.01 g | |
| T8 | 0 | 0.06 ± 0.01 vw | 0.08 ± 0.01 u | 0.12 ± 0.01 t |
| 7 | 0.04 ± 0.01 w | 0.07 ± 0.01 uv | 0.13 ± 0.01 t | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loghman, S.; Moayedi, A.; Mahmoudi, M.; Khomeiri, M.; Gómez-Mascaraque, L.G.; Garavand, F. Single and Co-Cultures of Proteolytic Lactic Acid Bacteria in the Manufacture of Fermented Milk with High ACE Inhibitory and Antioxidant Activities. Fermentation 2022, 8, 448. https://doi.org/10.3390/fermentation8090448
Loghman S, Moayedi A, Mahmoudi M, Khomeiri M, Gómez-Mascaraque LG, Garavand F. Single and Co-Cultures of Proteolytic Lactic Acid Bacteria in the Manufacture of Fermented Milk with High ACE Inhibitory and Antioxidant Activities. Fermentation. 2022; 8(9):448. https://doi.org/10.3390/fermentation8090448
Chicago/Turabian StyleLoghman, Shahram, Ali Moayedi, Mandana Mahmoudi, Morteza Khomeiri, Laura G. Gómez-Mascaraque, and Farhad Garavand. 2022. "Single and Co-Cultures of Proteolytic Lactic Acid Bacteria in the Manufacture of Fermented Milk with High ACE Inhibitory and Antioxidant Activities" Fermentation 8, no. 9: 448. https://doi.org/10.3390/fermentation8090448
APA StyleLoghman, S., Moayedi, A., Mahmoudi, M., Khomeiri, M., Gómez-Mascaraque, L. G., & Garavand, F. (2022). Single and Co-Cultures of Proteolytic Lactic Acid Bacteria in the Manufacture of Fermented Milk with High ACE Inhibitory and Antioxidant Activities. Fermentation, 8(9), 448. https://doi.org/10.3390/fermentation8090448