Abstract
There is a recent trend of using lactic acid bacteria for the production of gamma-aminobutyric acid (GABA). This study described a method that combines fermentation and self-buffered whole-cell catalysis for the efficient production of GABA using Levilactobacillus brevis CD0817. Upon the completion of GABA fermentation, cells were recovered to conduct whole-cell catalysis by which the substrate L-glutamic acid was catalytically decarboxylated to GABA. L-glutamic acid itself maintained the acidity essential for decarboxylation. To maximize the whole-cell catalysis ability, the effects of the cell culture method, catalysis temperature, catalysis time, cell concentration, and L-glutamic acid dosage were investigated. The results illustrate that the cells that were cultivated for 16 h in a fermentation medium supplemented with 20.0 g/L of glucose were the most suitable for the whole-cell catalytic production of GABA. At 16 h, the fermentative GABA content reached 204.2 g/L. Under optimized whole-cell catalytic conditions (temperature 45.0 °C, time 12.0 h, wet cells 25.0 g/L, and L-glutamic acid 120.0 g/L), 85.1 g/L of GABA was obtained, with 3.7 ± 0.9 g/L of substrate residue. GABA was recovered from the system by sequentially performing rotary vacuum evaporation, precipitation with ethanol, filtration with filter paper, and drying. The purity of the GABA product reached 97.1%, with a recovery rate of 87.0%. These data suggest that the proposed method has potential applications in the production of GABA.
1. Introduction
Gamma-aminobutyric acid (GABA) functions as the key inhibitory neurotransmitter in the central nervous system of mammalian species [1,2]. GABA also has many other well-documented biological activities, such as anti-hypertension [3], anti-diabetes [4], and anti-pain activities [5]. Lactic acid bacteria (LAB) belong to a significant cluster of Gram-positive bacteria that are widely distributed in natural habitats, such as fermented foods, vegetables, and human bodies. LAB strains have numerous vital physiological functions and are generally regarded as safe. In the last three decades, many efforts have been made to produce GABA by employing LAB strains as microbial cell factories [4,6,7].
LAB strains exploit the intracellular glutamic acid decarboxylase (GAD) system to synthesize GABA using L-glutamate as the substrate. The GAD system is composed of GAD enzymes (GadA/GadB) and L-glutamate/GABA antiporters (GadC). The working mechanism for the generation of GABA by the GAD system is as follows: (i) the substrate is imported into the cell by GadC; (ii) the imported substrate is catalyzed to GABA by intracellular GAD; and (iii) the final product, GABA, is exported out of the cell by GadC. Each decarboxylation needs to consume one proton (H+). As a result, the GAD system helps to maintain the intracellular physiological pH when the environmental pH decreases [4,8,9].
The screening and identification of GABA-producing LAB strains is the first essential step for the production of LAB-sourced GABA [4]. Previous investigations have demonstrated that LAB strains with GABA-producing ability are common in nature, especially in fermented foods. A lot of GABA-producing LAB strains have been isolated, such as Levilactobacillus brevis [10], L. buchneri [11], L. paracasei [12], and Lactococcus lactis [13]. In general, lactobacilli strains exhibit a robust GABA-producing ability compared to that of lactococci strains [4]. It is worth emphasizing that the top GABA-producing LAB strains belong to L. brevis. The GABA production of L. brevis strains CD0817 [14], ATCC367ΔglnR [10], and D17 [15] was found to reach 321.9, 284.7, and 177.7 g/L, respectively. These strains have exhibited potential for the industrial production of GABA.
It is equally essential to tailor appropriate GABA bioproduction processes fit to specific LAB strains [4,14]. In the early GABA fermentation studies performed using LAB strains, monosodium L-glutamate (MSG), were used as the substrate [4]. As aforementioned, however, each decarboxylation has to consume one H+ [9,16]. Therefore, MSG-based GABA fermentation suffers from the continuous feed of an exogenous acid agent. Recently, we set up an L-glutamic-acid-based GABA fermentation process [17,18]. It is well known that L-glutamic acid has a low isoelectric point (3.22) and low solubility (~ 6.0 g/L). This low solubility guarantees that little inhibitory effects occur even if the needed L-glutamic acid is added to the fermentation system all at once, because most of the added substrate exists in the form of powder. L-glutamic acid powder is slowly dissolved into the fermentation broth as decarboxylation progresses. The newly dissolved L-glutamic acid maintains the acid environment essential for decarboxylation. This L-glutamic-acid-based process can significantly improve fermentation efficiency and reduce costs [14,18].
The fermentative GABA broth of a LAB strain features rather complicated impurities [19]. One challenging issue in the production of GABA via fermentation performed using LAB is how to purify the product from broth. Several research teams have extracted GABA from the fermentative broths of LAB, but to date the outcomes have not been ideal because of the complex purification steps and the high loss rates of the product [19,20]. Thereafter, related scientific communities attempted to produce GABA using whole-cell catalysis. In these whole-cell bioprocesses, MSG or a mixture of MSG and L-glutamic acid was biotransformed into GABA with the help of an exogenous pH-buffering system [21,22]. Clearly, the available whole-cell bioprocesses introduce impurities, which are the added buffer and Na+ from MSG. The practicability of the available whole-cell catalysis strategies needs to be improved.
This work aimed to develop a combined bioprocess, coupling fermentation with whole-cell catalysis, for L. brevis CD0817 to efficiently produce GABA. The needed L-glutamic acid was added all at once into the whole-cell system, and with the progression of decarboxylation, L-glutamic acid was periodically released into the system to serve as the available substrate and maintain the acidity essential for decarboxylation. This self-buffered bioprocess is free of the contamination of extra buffering agents and the Na+ of MSG. A GABA product of high purity could be readily prepared from the whole-cell reaction broth. The results indicate that the proposed bioprocess is a promising method in the production of LAB-sourced GABA.
2. Materials and Methods
2.1. Reagents
Yeast extract FM408 was obtained from Angel Yeast Co., Ltd. (Wuhan, China). L-glutamic acid was provided by Dragon Biotech Co., Ltd. (Emeishan, China). MSG was obtained from Lanji Technology Development Co., Ltd. (Shanghai, China). GABA standard was obtained from Aladdin Biochemical Technology Co., Ltd. (Shanghai, China). A microcrystalline cellulose thin-layer chromatographic plate was obtained from Shengya Chemical Engineering Co., Ltd. (Shanghai, China). Xinhua grade I chromatographic paper was purchased from Hangzhou Xinhua Paper Industry (Hangzhou, China). The reagents used in the HPLC method were of HPLC grade. The other chemicals employed in this study were of analytical grade.
The amino acid derivatization agent was made by adding 10.0 mg o-phthalaldehyde and 10.0 μL 2-mercaptoethanol to 2.5 mL acetonitrile. The borate buffer was manufactured by adding 4.9 g boric acid into 100.0 mL water, adjusting the pH to 10.4 with NaOH, and then diluting the solution to 200.0 mL with water. The mobile phase was made as follows: 2.7 g sodium acetate trihydrate and 200.0 μL triethylamine were dissolved in 0.9 L water, the pH was adjusted to 7.3 using acetic acid, and then the solution was diluted to 1.0 L with water, followed by mixing with 250.0 mL acetonitrile [14].
2.2. Strain and Media
The used GABA producer was L. brevis CD0817 previously isolated from a fecal sample of a healthy adult [23]. The seed medium (pH 5.0) consisted of 10.0 g/L glucose, 35.0 g/L yeast extract, 28.0 g/L MSG, 1.0 g/L Tween-80, and 50.0 mg/L MnSO4·H2O. The starting fermentation medium contained 25.0 g/L glucose, 35.0 g/L yeast extract, 1.0 g/L Tween-80, and 50.0 mg/L MnSO4·H2O. It was supplemented with L-glutamic acid in the amount of 300.0 g per liter of fresh medium just before inoculation. The substrate, glucose, and the other components were individually autoclaved at 121 °C for 20 min and mixed together prior to use [14,23].
2.3. GABA Fermentation Trial
2.3.1. Preparation of Inoculum
The frozen stock culture of L. brevis CD0817 was resuscitated by incubation at 30 °C for 2 d subsequent to streaking onto the seed medium agar. One colony was transferred into a tube containing 5.0 mL of the seed medium and incubated at 30 °C for 24 h. Then, the 5.0 mL culture broth was transferred into a 250 mL Erlenmeyer flask containing 100.0 mL of the seed medium and cultured in a ZHWY-2102C rotary shaker (Zhicheng Inc., Shanghai, China) at 30 °C and 100 rpm. The culture broth could be used as inoculum when its absorbance at 600 nm (A600) was close to 4.0. The A600 value was monitored using a UV1200B ultra-violet–visible spectrophotometer (Mapada Instruments, Shanghai, China).
2.3.2. Fermentation
The fermentations started by inoculating the seed culture at a size of 10% (v/v) into fermentation media supplemented with different levels of glucose, and then they were statically maintained at 30 °C until the end. Samples were aseptically withdrawn every 4 h. All the experiments were conducted in triplicate.
2.4. Optimization of Whole-Cell Catalysis System
Unless elsewhere stated, whole-cell catalysis was initiated by resuspending the cells in an aqueous solution saturated with excess L-glutamic acid, and the duration was 16 h. The factors that may affect GABA formation, namely, catalysis temperature, catalysis time, cell concentration, and L-glutamic acid dosage, were optimized one by one. The recyclability of the whole-cell system was also assessed. The samples were withdrawn at an interval of 4 h. The relative contents of GABA were determined using the sensitivity intensified ninhydrin-amino acid chromogenic system (SINACS) described below.
2.5. Recovery of GABA from Whole-Cell Conversion Solution
A total of 500.0 mL of whole-cell conversion broth was centrifuged at 8000× g and 4 °C for 10 min. The supernatant was moved to a 1 L round-bottomed flask, followed by rotary vacuum evaporation at 60 °C. The evaporation was terminated when explosive boiling occurred. Then, 200.0 mL of anhydrous ethanol was added to the hot solution, and it was kept at room temperature for several minutes to fully precipitate GABA. The precipitate was harvested by suction filtration using the Xinhua grade I chromatographic paper, dried at 60 °C, and weighed.
2.6. Analytic Methods
In the optimization of the whole-cell catalysis system, a large number of samples were involved. To improve work efficiency, the high-throughput SINACS method (unpublished data, in preparation) was employed to relatively quantitate GABA. During whole-cell catalytic decarboxylation, the formation of GABA is synchronized with the dissolution of L-glutamic acid solid powder. Therefore, the more GABA is produced, the higher the A570 value of the chromophore of ninhydrin-GABA/L-glutamic acid. The A570 value represents the relative content of GABA. A SINACS kit (2.9 mL) composed of 1% ninhydrin, 35.0 μL 0.2 M sodium acetate buffer (pH 5.0), 40.0% ethanol, and 25.0% ethyl acetate was used. The color development reaction was performed by adding 0.1 mL of the sample to the SINACS kit and then by heating it at 70 °C for 30 min. The A570 values of the reacted solutions were obtained using the spectrophotometer. Note that, in the optimization of the effects of temperature, the samples were saturated using L-glutamic acid powder at 50 °C for 2.0 h before the SINACS analysis to eliminate the difference in A570 values resulting from the variation in the solubility of L-glutamic acid. The SINACS chromogenic reaction mechanism is identical to that of the classical ninhydrin reagent. However, the SINACS method provides a more stable chromophore and a 30-fold increase in sensitivity.
Pre-staining thin-layer chromatography was conducted as previously described. In brief, samples loaded in 2.0 μL aliquots onto a microcrystalline cellulose thin-layer plate were separated using the ascending technique in n-butanol–acetic acid-water (5:3:2) supplemented with 1.2% (w/v) ninhydrin. The plate was heated at 80 °C for 10 min for the chromogenic reaction afterwards [24,25].
When absolute quantitation was necessary, the HPLC method was adopted to measure GABA as follows: A cell-free sample was subjected to a 5 min boiling treatment, followed by centrifugation at 12,000× g for 5 min. Then, 20.0 μL of supernatant was derivatized at 25.0 °C for 5.0 min by 20.0 μL of derivatization reagent and 100.0 μL of borate buffer. The derivatized sample was analyzed using an Agilent 1200 system (Agilent Technologies Inc., Santa Clara, CA, USA) loaded with Agilent Eclipse XDB-C18 column (4.6 × 150 mm, 5 μm). The HPLC operational parameters were as follows: a column temperature of 30 °C; mobile phase flow rate of 0.8 mL/min; and detection wavelength of 338 nm [14]. All the data are presented as means ± standard deviations.
The cell biomass is expressed as an A600 value. Cells were recovered by centrifugation and resuspended in an appropriate volume of 0.8% NaCl. A600 values were obtained using the spectrophotometer.
2.7. Calculations and Statistical Analyses
The substrate molar conversion rate (k) was calculated by adopting the following formula:
where CGABA is the concentration of GABA (g/L); mGlu is the mass of consumed L-glutamic acid (g); 147.1 is the molar mass of L-glutamic acid (g/mol); 103.1 is the molar mass of GABA (g/mol); and V is the volume (L).
The statistical analyses were completed using Microsoft Office Excel (Microsoft Corp., Redmond, USA). Data are given as means ± standard deviations (n = 3). All charts and graphs were plotted using GraphPad Prism 8 (GraphPad Software Inc., San Diego, CA, USA).
3. Results and Discussion
3.1. Effects of Cell Culture Method on Whole-Cell Catalysis
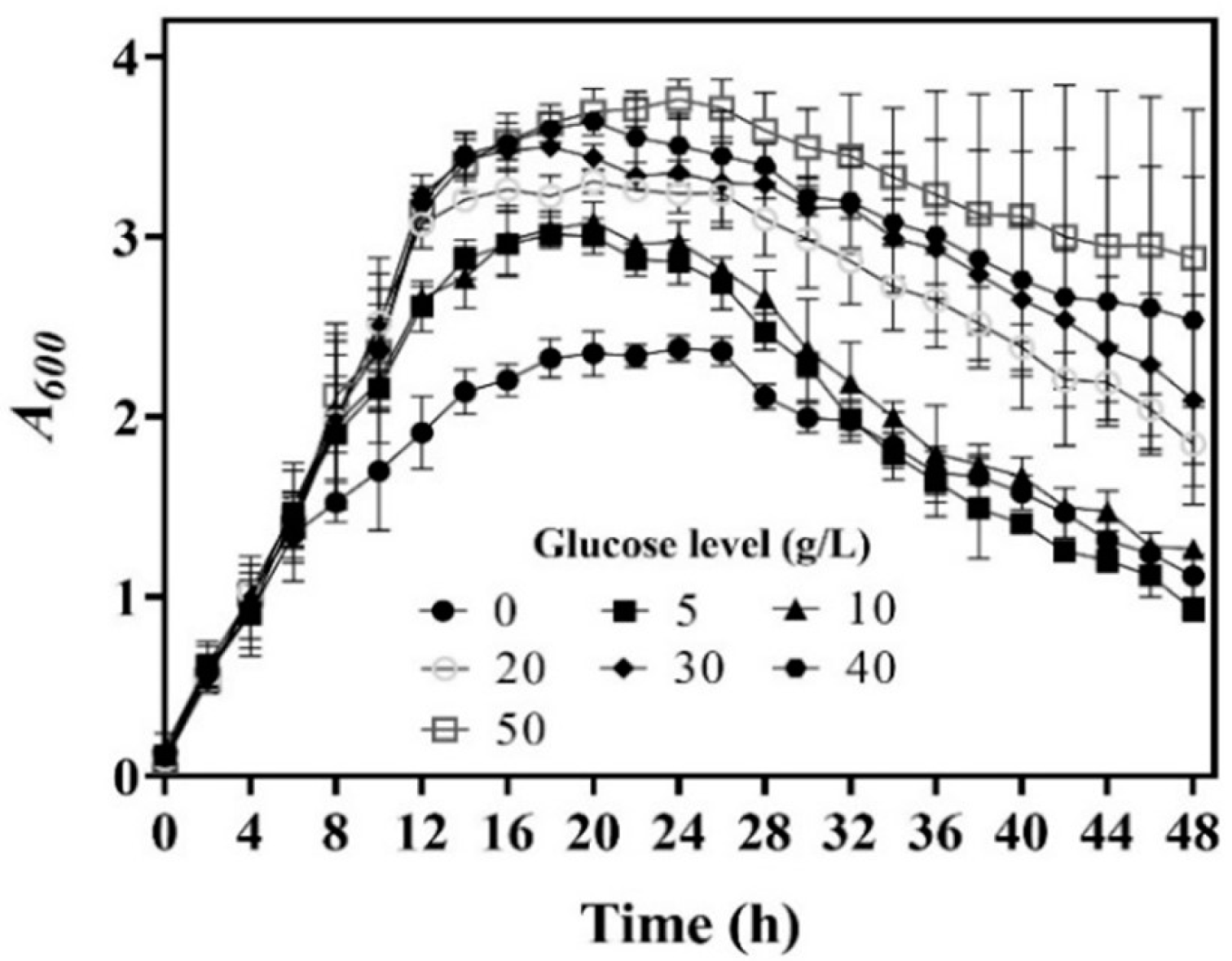
In a previous fermentation experiment, we observed that 5.0 g/L of glucose was most suitable to generate GABA with L. brevis CD0817. A higher level (10.0–40.0 g/L) of glucose did promote cell growth, but it also slightly reduced fermentative GABA production, which was attributed to the negative effect of its metabolites, such as lactic acid, ethanol, and acetic acid [14]. We suspect that whole-cell catalysis differs from fermentation in glucose demand because these metabolites are absent from whole-cell catalysis. To determine the glucose level that maximizes whole-cell catalysis, we performed GABA fermentations using L. brevis CD0817 with various levels of glucose. The results indicate that cell growth increased with the increase in glucose level (Figure 1), which is in line with the previous data [14].
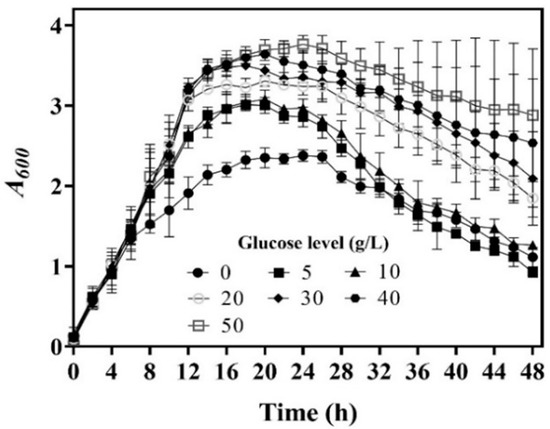
Figure 1.
Effects of glucose level on cell growth of L. brevis CD0817. Cell growth is presented as A600 (n = 3). A600, absorbance at 600 nm.
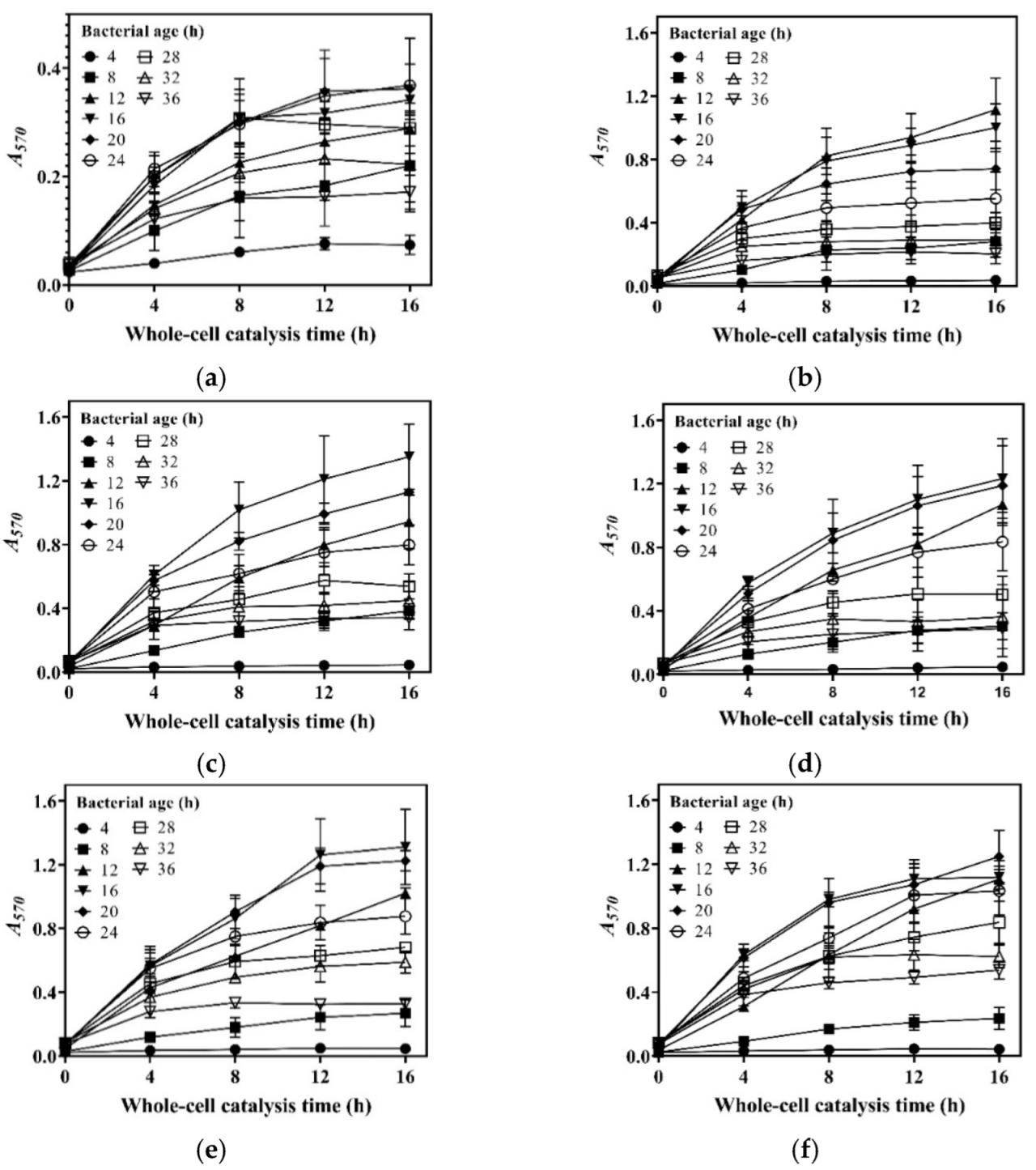
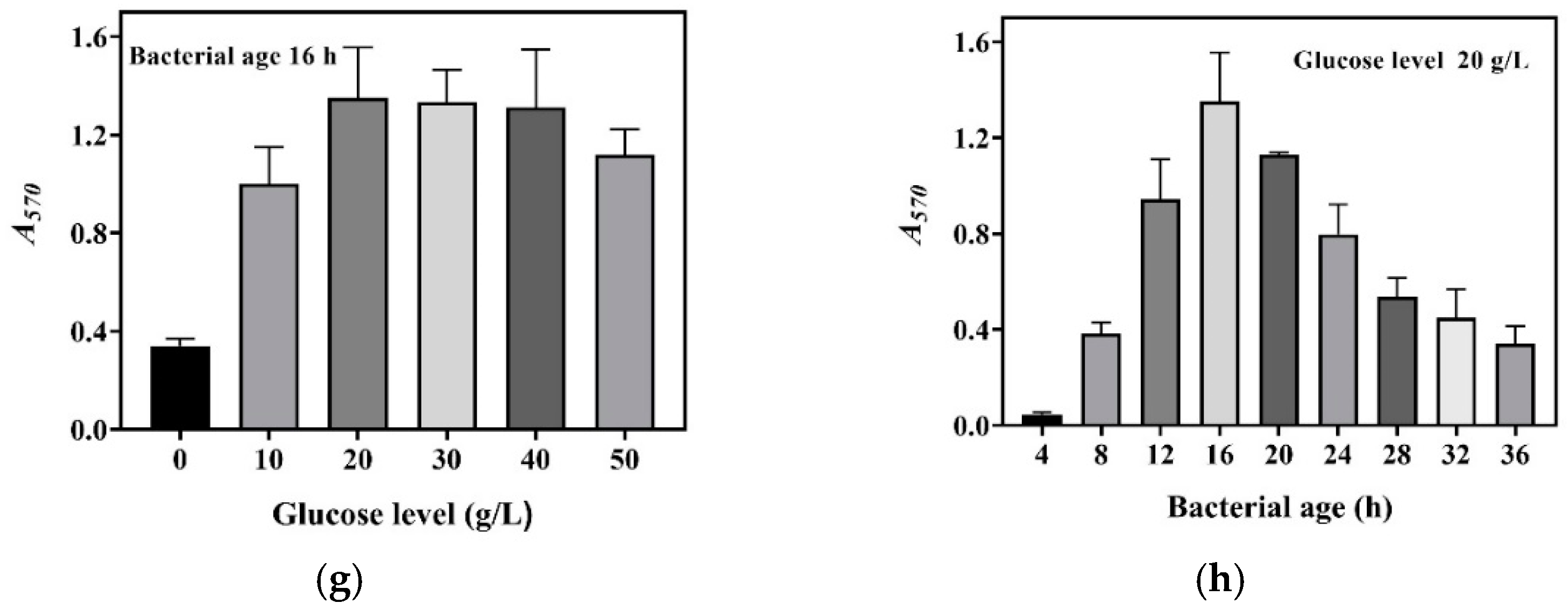
We harvested cells at 4, 8, 12, 16, 20, 24, 28, 32, and 36 h during the aforementioned fermentations; set up a whole-cell catalysis system for each harvest of cells; and then determined the GABA production-time profile up to 16.0 h for each whole-cell catalysis system (Figure 2a–f). For the sake of intuition, the catalysis abilities of the whole-cell systems, constructed by 16 h cells from different concentrations of glucose, were compared alone (Figure 2g). The results indicate that whole-cell catalytic ability increased as the glucose level increased from 0 to 20.0 g/L, then plateaued, and afterwards slightly decreased when the glucose level exceeded 40.0 g/L (Figure 2a–g). The 20.0–40.0 g/L level of glucose maximized whole-cell catalysis, and this was at least partly attributed to the increased number of cells and the elimination of inhibitory metabolites. The whole-cell catalysis system that originated from an excessively high level (50.0 g/L) of glucose had a decreased ability, possibly because the negative effect was too severe to be compensated by the increased number of cells and the removal of metabolites. Currently, the exact mechanism underlying this is poorly understood. In sum, cells cultivated by 20.0–40.0 g/L glucose were the most suitable for the construction of the whole-cell system. Therefore, 20.0 g/L of glucose was used in the following experiments.
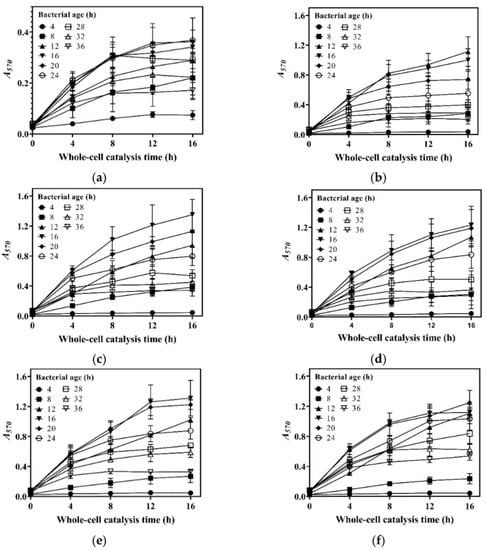
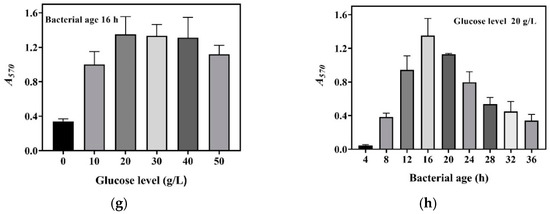
Figure 2.
Effects of glucose level and bacterial age on whole-cell catalysis. The GABA fermentations were individually conducted in fermentation media containing 0 (a), 10.0 (b), 20.0 (c), 30.0 (d), 40.0 (e), and 50.0 g/L (f) of glucose. Cells were recovered from each fermentation broth at 4, 8, 12, 16, 20, 24, 28, 32, and 36 h and used to conduct 16 h whole-cell catalysis. A GABA production-time profile was created for each whole-cell system. (g) Effects of glucose level on whole-cell catalytic ability (the bacterial age was 16 h). (h) Effects of bacterial age on whole-cell catalytic ability (cells were cultivated in the fermentation medium with 20.0 g/L glucose). Bacterial age means GABA fermentation time of L. brevis CD0817, shown in the blank area in the upper left corner of subfigures (a–f). The relative content of GABA was determined using the SINACS method, and it is expressed as A570 (n = 3). A570, absorbance at 570 nm; SINACS: sensitivity intensified ninhydrin-amino acid chromogenic system.
Next, the effects of bacterial age on whole-cell catalysis were evaluated. To achieve this, GABA fermentation was conducted in the fermentation medium supplemented with 20.0 g/L of glucose. Cells were recovered from the fermentation broth at 4, 8, 12, 16, 20, 24, 28, 32, and 36 h and used to conduct whole-cell catalysis. The results demonstrate that bacterial age had an important effect on the catalysis ability of the whole-cell system. The catalysis ability was enhanced with bacterial age from 0 to 16 h, and, thereafter, it gradually decreased with bacterial age (Figure 2h). This resulted from the increase in cell density with fermentation time from 0 to 16 h, followed by a decline with fermentation time beyond 16 h (Figure 1). At 16 h, the fermentative GABA content reached 204.2 ± 2.7 g/L, which can cause a rather high osmotic pressure [13,26]. It is difficult for cells to survive in such a hypertonic environment. With the progression of fermentation, a cell is more vulnerable to dying, possibly resulting from a continuous increase in osmotic pressure. Therefore, the cells fermented for 16 h were chosen for whole-cell catalysis.
3.2. Effects of Temperature on Whole-Cell Catalysis
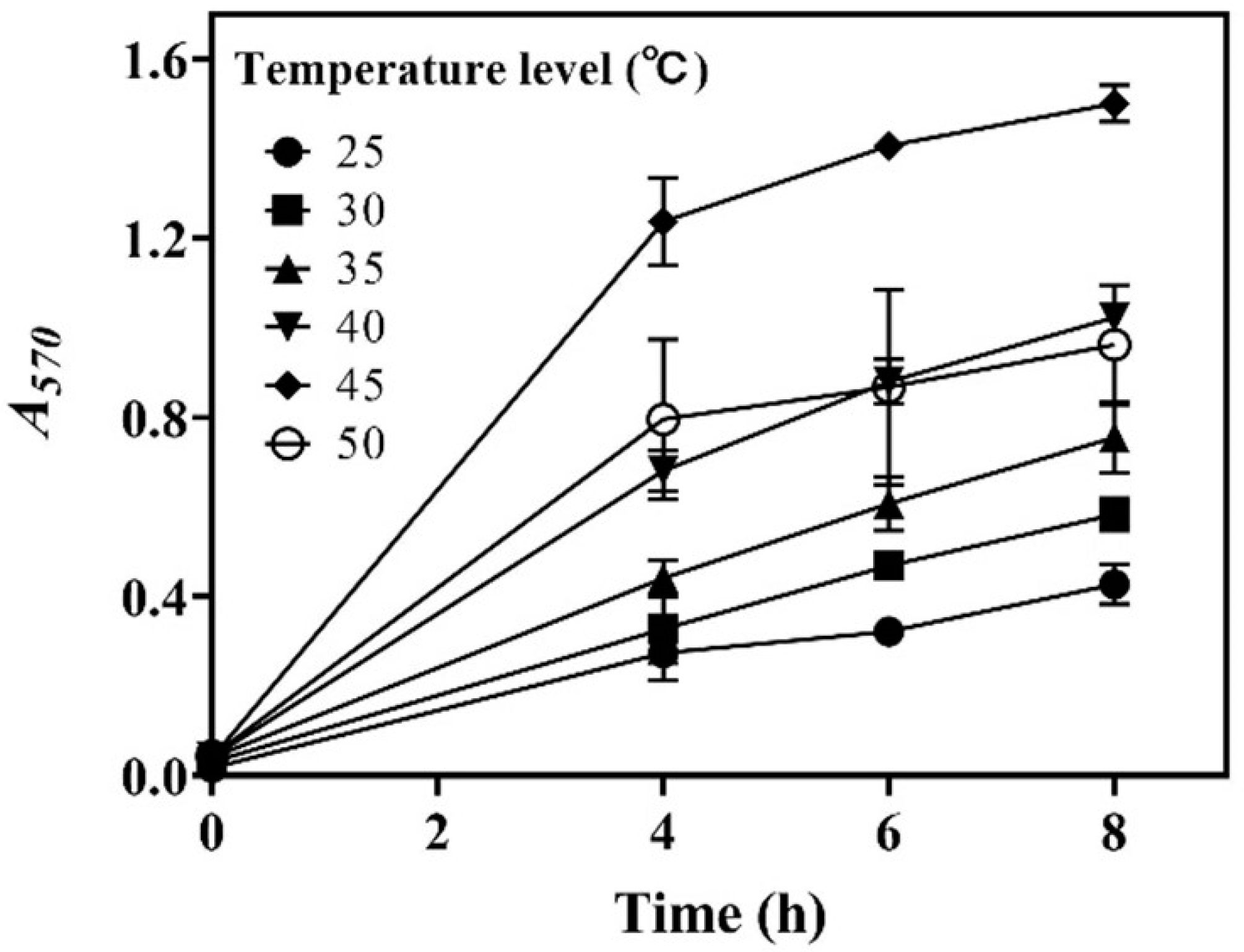
In this section, the effects of incubation temperature (25.0–50.0 °C) on whole-cell catalysis were assessed. The results demonstrate that temperature substantially influenced decarboxylation efficiency. Decarboxylation was enhanced as the temperature was increased from 25.0 to 45.0 °C, and then it decreased with further increases in temperature. Clearly, 45.0 °C was the optimal temperature for the strain to produce GABA in the whole-cell system (Figure 3). Interestingly, the literature has shown that a temperature of around 45.0 °C is favorable for the whole-cell catalytic GABA synthesis performed by L. brevis strains [21,27,28].
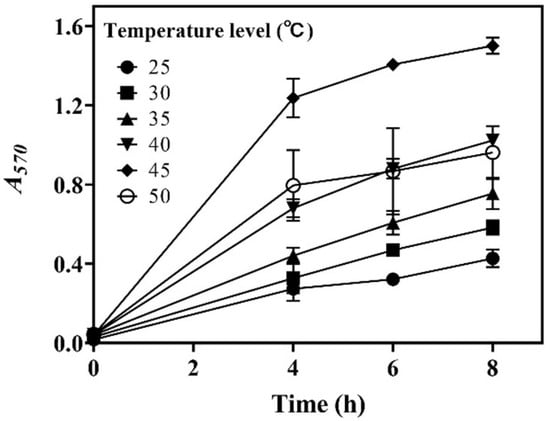
Figure 3.
Effects of temperature on whole-cell catalysis. The relative content of GABA is expressed as A570 (n = 3). A570, absorbance at 570 nm.
3.3. Effects of Wet Cell Level on Whole-Cell Catalysis
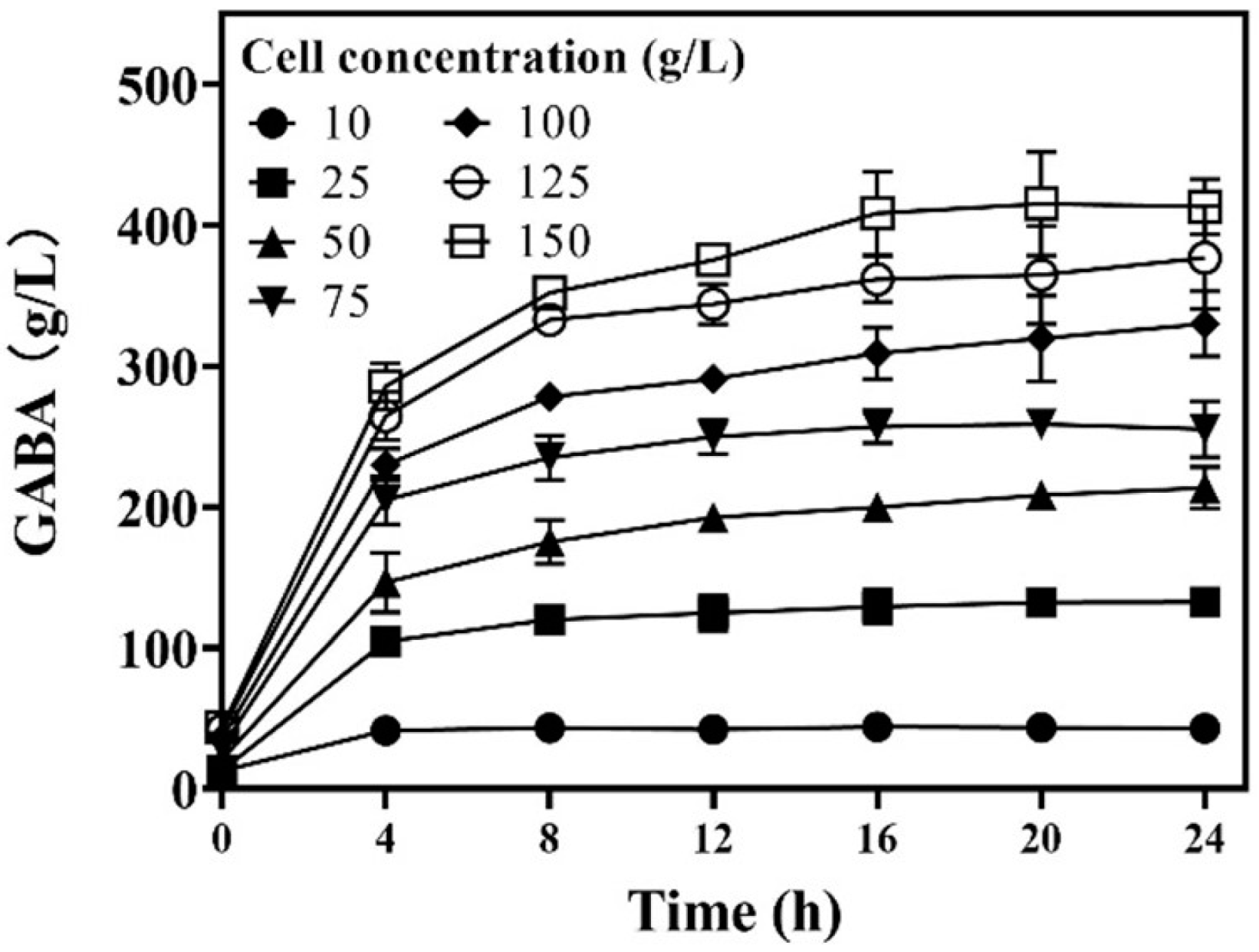
Each cell functioned as a catalyst. Cell concentration should affect GABA synthesis efficiency. Therefore, the effect of cell concentration on whole-cell catalysis was examined. The results show that GABA production gradually elevated as the wet cell concentration increased from 10.0 to 150.0 g/L. However, GABA productivity also increased as the wet cell concentration increased from 10.0 to 25.0 g/L, and then it gradually decreased with further increases in wet cell concentration. For all the whole-cell catalyzes, reactions were completed at around 8.0 h (Figure 4). Therefore, a catalysis period of 12 h was proposed to ensure the completion of the reaction. During the first 12 h, GABA productivity increased from 4.2 ± 0.8 to 5.0 ± 0.4 g/g/h (g GABA/g wet cells/h) as cell concentration increased from 10.0 to 25.0 g/L; afterwards, it gradually declined to 2.5 ± 0.01 g/g/h as cell concentration increased to 150.0 g/L. In detail, the GABA productivities at wet cell levels of 10, 25, 50, 75, 100, 125, and 150 g/L were 4.2 ± 0.8, 5.0 ± 0.4, 3.9 ± 0.1, 3.3 ± 0.1, 2.9 ± 0.1, 2.8 ± 0.1, and 2.5 ± 0.01 g/g/h, respectively. Clearly, 25.0 g/L of wet cells showed the highest GABA productivity. From the aspect of full utilization of cellular catalysis, the wet cell concentration was suggested to be 25.0 g/L.
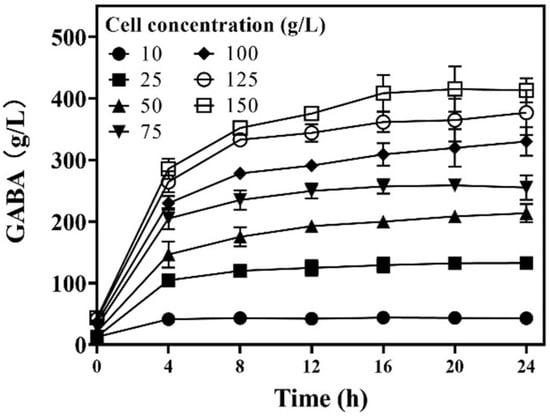
Figure 4.
Effects of cell concentration on GABA production. GABA was determined using the HPLC method (n = 3).
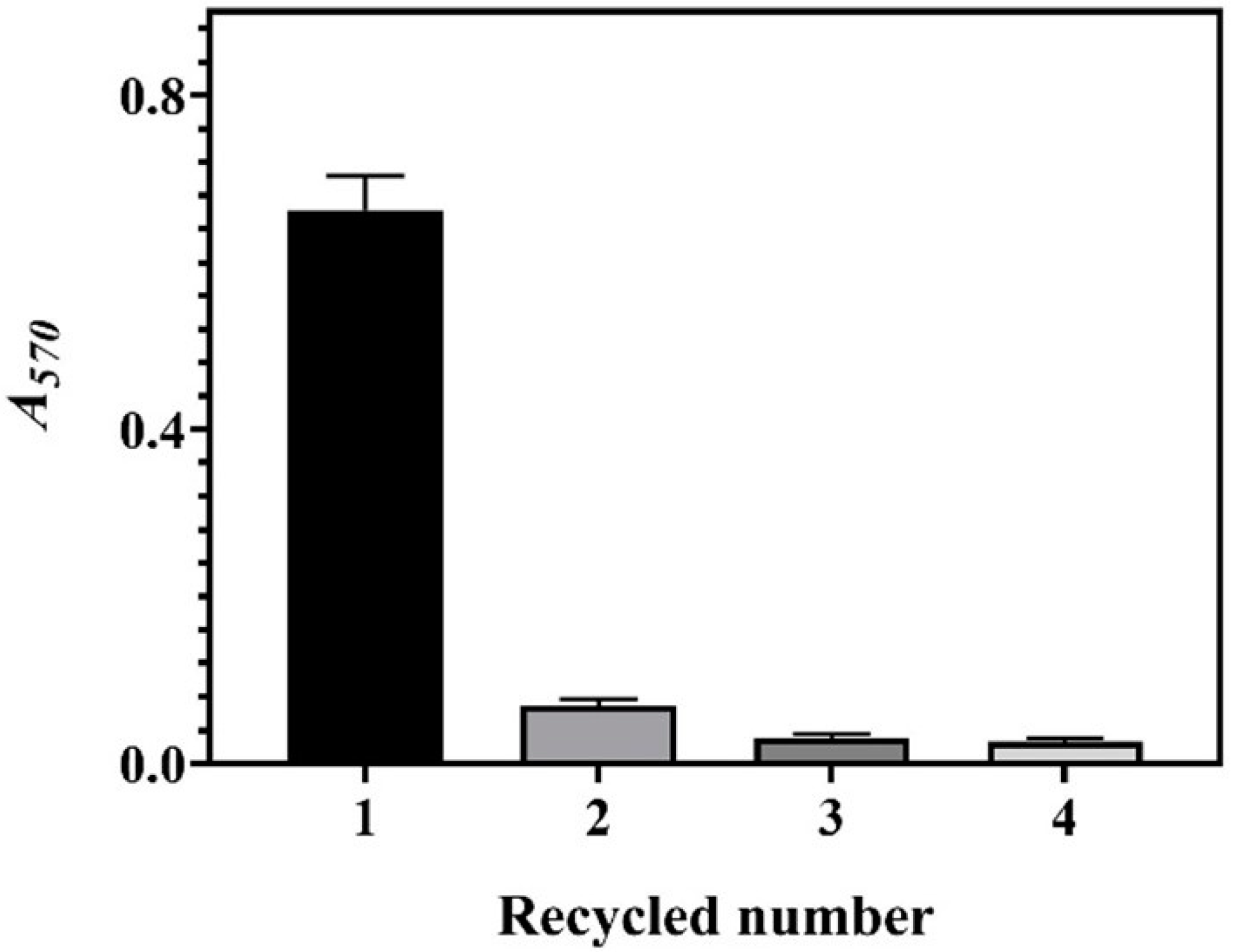
3.4. Recyclability of Whole-Cell System
We successively ran the system four times to determine recyclability. Each run was only performed for a duration of 4 h. The results indicate that the current whole-cell system can only be used once (Figure 5). We attributed this to the cell lysis during the first cycle of the reaction. As aforementioned, the cells were obtained from a fermentative broth with a high osmotic pressure arising from 204.2 ± 2.7 g/L of GABA. As a result, cell lysis occurred when the hypertonic cell entered a hypotonic environment. After the first cycle reaction was completed, few cells were grown when the whole-cell bioconversion broth was plated onto the medium agar, confirming that almost 100% of the cells were lysed. The bioconversion of GABA, however, lasted for approximately 8 h (Figure 2, Figure 3 and Figure 4). This may be interpreted from the role of the released intracellular GAD enzyme. The available LAB whole-cell systems could be used about four times [27], possibly because the cells were from a low osmotic environment. Accordingly, these systems produced low levels of GABA (4.0–12.0 g/L) in a single run [27,28]. Although our system was disposable, it produced 85.1 ± 0.04 g/L of GABA (as discussed below), surpassing the accumulative GABA of any available hypotonic system. Therefore, our system is convenient and efficient relative to the currently available ones.
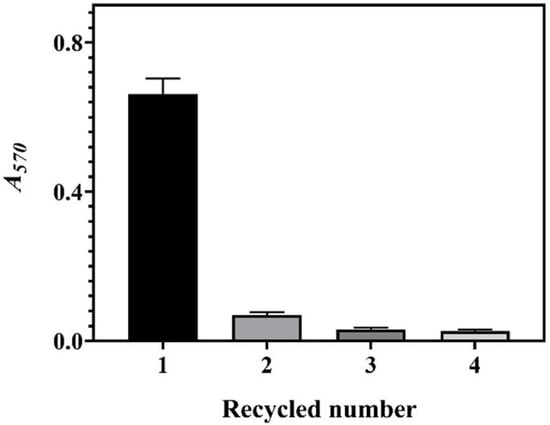
Figure 5.
GABA productions of four sequential runs of the whole-cell catalysis system. The relative content of GABA is expressed as A570 (n = 3). A570, absorbance at 570 nm.
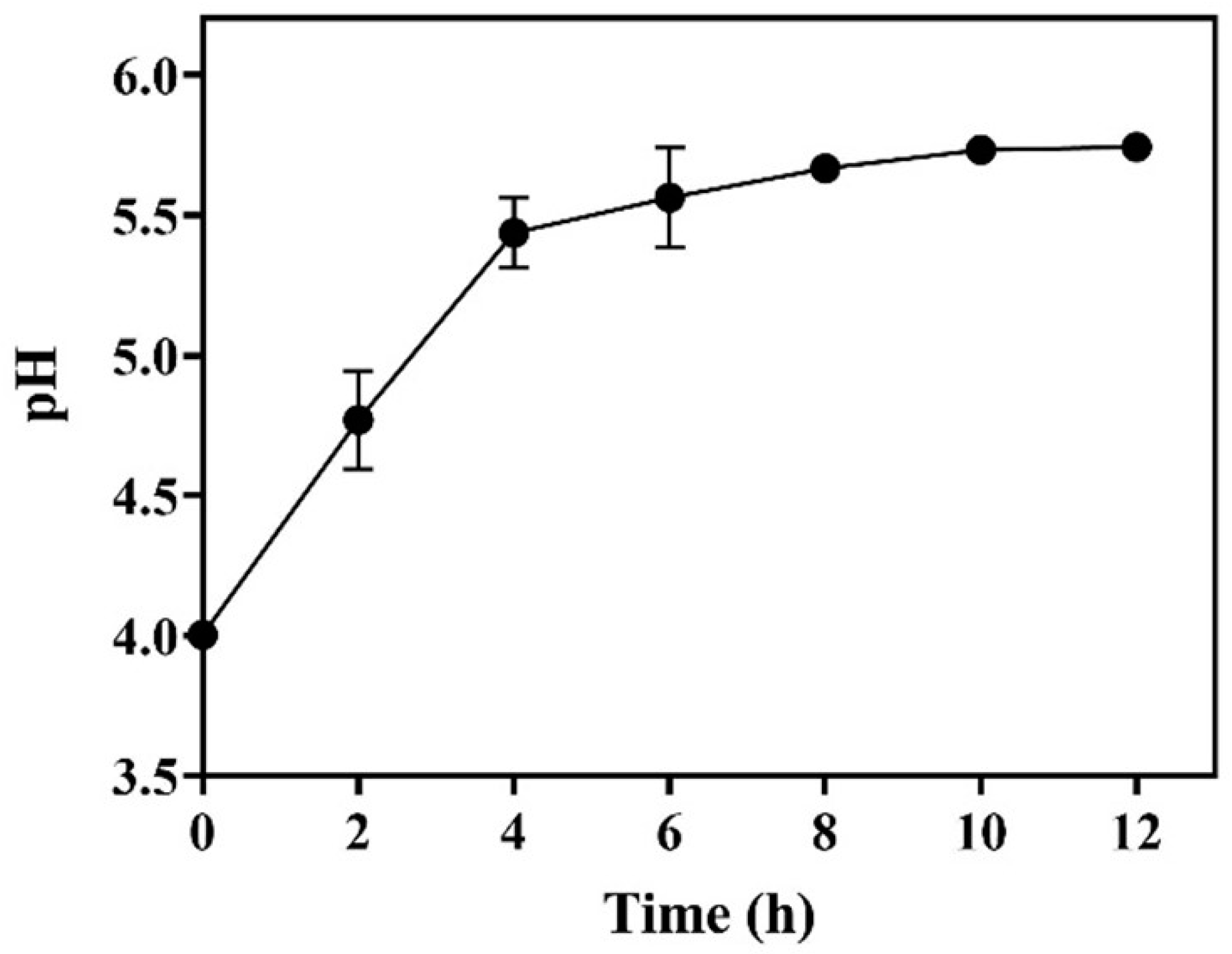
The required L-glutamic acid could be added all at once, but most of it exists in the form of powder, which is ascribed to its low solubility (~6.0 g/L). The dissolution of substrate powder is synchronized with decarboxylation [14,18]. As a result, the dissolved substrate buffers the whole-cell system. We determined the pH profile with time during the whole-cell bioconversion driven by 25.0 g/L of wet cells. As shown in Figure 6, the pH gradually increased with time and followed a trend identical to that of GABA formation (Figure 2, Figure 3 and Figure 4). The increase in pH resulted from the substitution of neutral GABA for acid L-glutamic acid [29]. Reportedly, the optimal pH for the GADs of LAB strains is around 5.0 [4,6]. The pH of the current reaction system was always maintained at around 5.0 (from 4.0 to 5.8). This acidity led to a satisfactory decarboxylation efficiency. A similar outcome was previously obtained in L-glutamic-acid-based GABA fermentations [14,18].
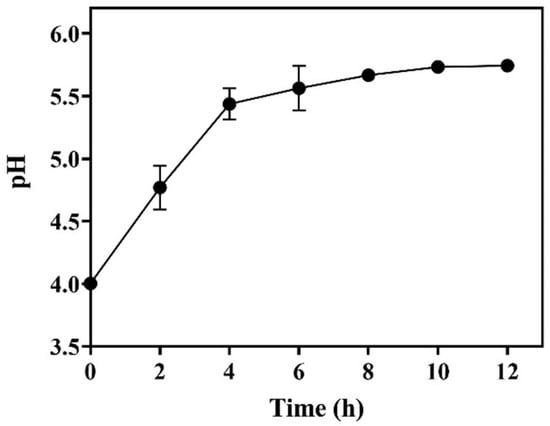
Figure 6.
Time course of pH in whole-cell catalysis (n = 3).
Another feature of our system is that it does not involve any exogenous buffering reagents. The omitting of exogenous buffering agents implies that GABA can be easily purified and recovered. The available whole-cell catalysis systems adopted MSG or a mixture of MSG and L-glutamic acid as the substrate [21,28]. As a result, an extra buffering agent has to be designed to offset the increase in pH arising from decarboxylation. This buffer complicates the operation, increases the cost, and is also not conducive to the purification of GABA. In addition, we suspect that an exogenous buffer is unlikely to always maintain pH 5.0, which is optimal for GADs activity. In sum, the proposed whole-cell system using L-glutamic acid as the substrate shows some advantages over the currently available ones.
3.5. Recovery of GABA from Whole-Cell System
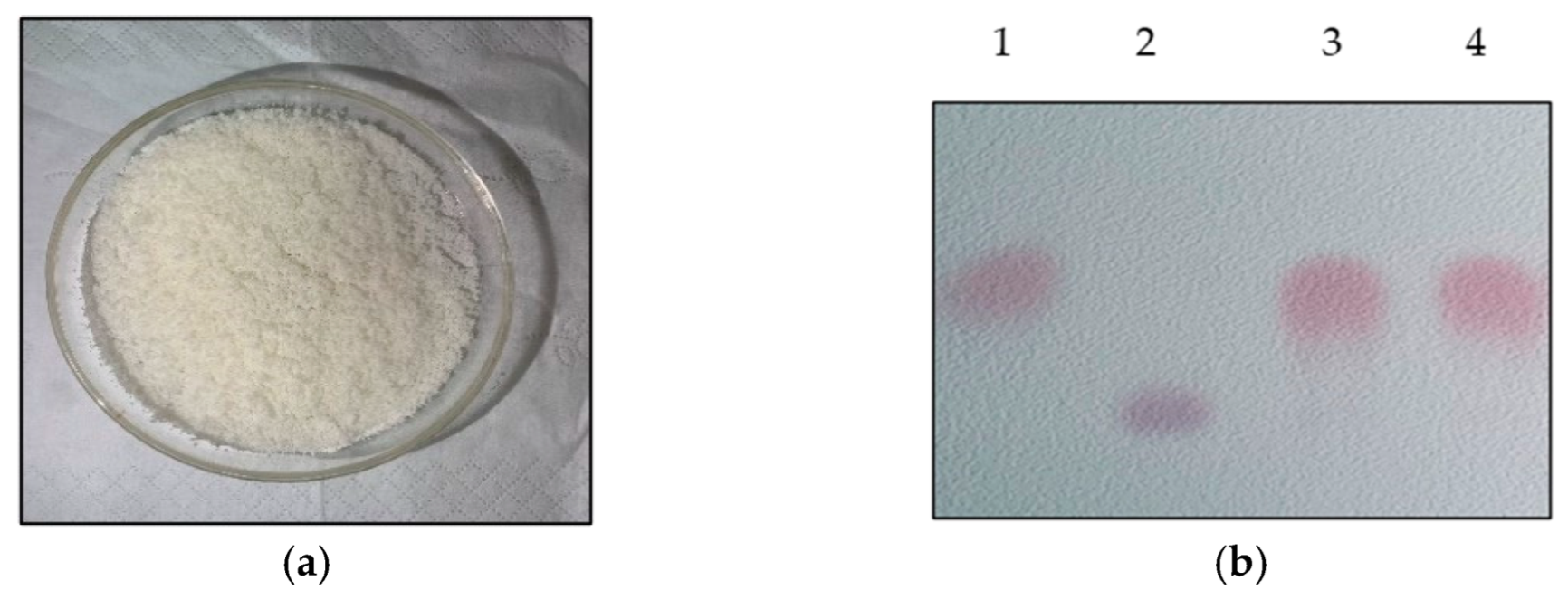
L-glutamic-acid-based GABA fermentation was conducted by using L. brevis CD0817 in the fermentation medium with 20 g/L of glucose. The fermentation was ended after being statically maintained at 30 °C for 16 h. The fermentative GABA concentration was quantitated to be 204.2 ± 2.7 g/L using the HPLC method. The cells were harvested to construct a 500.0 mL whole-cell catalysis system. The system was composed of 25.0 g/L of wet cells and 300.0 g/L of L-glutamic acid (mostly existing as powder). Whole-cell catalysis was carried out statically at 45 °C for 12 h and shaken well every 1.0 h. Upon the completion of the reaction, the solution was boiled for 10 min and then centrifuged at 8000× g and 4.0 °C for 10 min. The GABA titer was determined to be 85.1 g/L using the HPLC method. The substrate residue was 3.7 ± 0.9 g/L. GABA was recovered using the method described in the Materials and Methods Section. The recovery was simple and benefited from the concise bioconversion process. The manufactured GABA product was a white solid powder (Figure 7a). The pre-staining thin-layer chromatogram of the product is shown in Figure 7b.
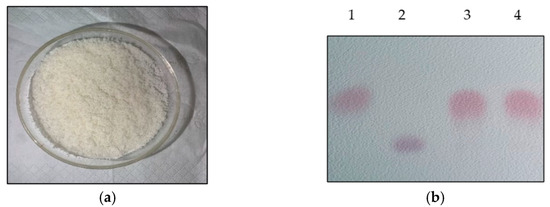
Figure 7.
GABA product (a) and its pre-staining thin-layer chromatogram (b). Lane 1: 1.0 g/L GABA standard; lane 2: 2.0 g/L L-glutamic acid standard; lane 3: purified GABA product; and lane 4: 100-fold dilution of whole-cell reaction broth.
The purity of the GABA product was measured using the HPLC method. The results indicate that the purity of the product was 97.1%, with a recovery of 87.0%. The only possible impurities may be trace L-glutamic acid, intracellular molecules, and the debris of cells. As a proteinaceous amino acid, L-glutamic acid is an ingredient in our diets. Moreover, as a food-associated strain, L. brevis is prevalent in fermented foods and human bodies [30,31]. Thus, these impurities are acceptable.
The purification of fermentative GABA involves a complex combination of multiple steps, including, but not limited to, centrifugation, decolorization, filtration, desalination, ion-exchange chromatography, and precipitation [19,20]. These operations increase the cost of production. From economical and practical points of view, we suggest that fermentative GABA be directly applied in feed industries and that GABA produced by whole-cell catalysis be applied in food industries. The proposed combined bioprocess for GABA production is worthy of promotion.
4. Conclusions
In this work, a bioprocess combining fermentation and self-buffered whole-cell catalysis was set up for L. brevis CD0817 to efficiently synthesize GABA. After 16 h of fermentation, the GABA titer reached 204.2 g/L. Moreover, the whole-cell synthetic GABA titer reached up to 85.1 g/L after a 12 h reaction using the 16 h cells as catalysts. A GABA product with a purity of 97.1% was easily recovered from the whole-cell system, with a recovery rate of 87.0%. The fermentative GABA may be directly applied in feed industries, while the GABA purified from the whole-cell catalysis system may be used in food industries. These findings demonstrate that the current bioprocess is promising for the production of LAB-sourced GABA.
Author Contributions
Conceptualization and funding acquisition, H.L.; investigation, methodology, data collection, and analysis, T.S.; writing—original draft preparation, M.J., data validation and interpretation, L.W. and C.W.; writing—review and editing, J.P.; formal analysis, Z.L.; supervision, S.W. All authors have read and agreed to the published version of the manuscript.
Funding
We are grateful to the National Natural Science Foundation of China (grant Nos. 32160014 and 31570070) and the State Key Laboratory of Food Science and Technology at Nanchang University (grant No. SKLF-ZZB-202118).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wong, C.G.T.; Bottiglieri, T.; Snead, O.C. GABA, gamma-hydroxybutyric acid, and neurological disease. Ann. Neurol. 2003, 54, S3–S12. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Rui, Q.Y.; Han, S.T.; Wu, X.J.; Wang, X.Y.; Wu, P.; Shen, Y.P.; Dai, H.; Xue, Q.; Li, Y.G. Reduced GABA levels in the medial prefrontal cortex are associated with cognitive impairment in patients with NMOSD. Mult. Scler. Relat. Disord. 2022, 58, 103496. [Google Scholar] [CrossRef] [PubMed]
- Han, S.M.; Lee, J.S. Production and its anti-hyperglycemic effects of gamma-aminobutyric acid from the wild yeast strain Pichia silvicola UL6-1 and Sporobolomyces carnicolor 402-JB-1. Mycobiology 2017, 45, 199–203. [Google Scholar] [CrossRef] [Green Version]
- Li, H.X.; Cao, Y.S. Lactic acid bacterial cell factories for gamma-aminobutyric acid. Amino Acids 2010, 39, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Francois, A.; Low, S.A.; Sypek, E.I.; Christensen, A.J.; Sotoudeh, C.; Beier, K.T.; Ramakrishnan, C.; Ritola, K.D.; Sharif-Naeini, R.; Deisseroth, K.; et al. A brainstem-spinal cord inhibitory circuit for mechanical pain modulation by GABA and enkephalins. Neuron 2017, 93, 822–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.L.; Shah, N.P. High gamma-aminobutyric acid production from lactic acid bacteria: Emphasis on Lactobacillus brevis as a functional dairy starter. Crit. Rev. Food Sci. Nutr. 2016, 57, 3661–3672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhakal, R.; Bajpai, V.K.; Baek, K.-H. Production of gaba (γ-aminobutyric acid) by microorganisms: A review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotter, P.D.; Hill, C. Surviving the acid test: responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, P.L.C.; Waterman, S.R. Acid stress, anaerobiosis and gadCB: lessons from Lactococcus lactis and Escherichia coli. Trends Microbiol. 1998, 6, 214–216. [Google Scholar] [CrossRef]
- Gong, L.; Ren, C.; Xu, Y. GlnR negatively regulates glutamate-dependent acid resistance in Lactobacillus brevis. Appl. Environ. Microbiol. 2020, 86, e02615–e02619. [Google Scholar] [CrossRef]
- Zhao, A.; Hu, X.; Pan, L.; Wang, X. Isolation and characterization of a gamma-aminobutyric acid producing strain Lactobacillus buchneri WPZ001 that could efficiently utilize xylose and corncob hydrolysate. Appl. Microbiol. Biotechnol. 2015, 99, 3191–3200. [Google Scholar] [CrossRef]
- Komatsuzaki, N.; Shima, J.; Kawamoto, S.; Momose, H.; Kimura, T. Production of gamma-aminobutyric acid (GABA) by Lactobacillus paracasei isolated from traditional fermented foods. Food Microbiol. 2005, 22, 497–504. [Google Scholar] [CrossRef]
- Laroute, V.; Mazzoli, R.; Loubiere, P.; Pessione, E.; Cocaign-Bousquet, M. Environmental conditions affecting GABA production in Lactococcus lactis NCDO 2118. Microorganisms 2021, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Zhu, Y.; Wang, L.; Sun, T.; Pan, H.; Li, H. pH auto-sustain-based fermentation supports efficient gamma-aminobutyric acid production by Lactobacillus brevis CD0817. Fermentation 2022, 8, 208. [Google Scholar] [CrossRef]
- Gong, L.C.; Ren, C.; Xu, Y. Deciphering the crucial roles of transcriptional regulator GadR on gamma-aminobutyric acid production and acid resistance in Lactobacillus brevis. Microb. Cell Fact. 2019, 18, 108. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, T.; Hayashi, H.; Abe, K. Exchange of glutamate and gamma-aminobutyrate in a Lactobacillus strain. J. Bacteriol. 1997, 179, 3362–3364. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Qiu, T.; Huang, G.; Cao, Y. Production of gamma-aminobutyric acid by Lactobacillus brevis NCL912 using fed-batch fermentation. Microb. Cell Fact. 2010, 9, 85. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Liu, X.; Fu, J.; Wang, S.; Chen, Y.; Chang, K.; Li, H. Substrate sustained release-based high efficacy biosynthesis of GABA by Lactobacillus brevis NCL912. Microb. Cell Fact. 2018, 17, 80. [Google Scholar] [CrossRef] [Green Version]
- Li, H.X.; Qiu, T.; Chen, Y.; Cao, Y.S. Separation of gamma-aminobutyric acid from fermented broth. J. Ind. Microbiol. Biotechnol. 2011, 38, 1955–1959. [Google Scholar] [CrossRef]
- Gao, Q.; Duan, Q.; Wang, D.P.; Zhang, Y.Z.; Zheng, C.Y. Separation and purification of gamma-aminobutyric acid from fermentation broth by flocculation and chromatographic methodologies. J. Agric. Food Chem. 2013, 61, 1914–1919. [Google Scholar] [CrossRef]
- Shi, X.; Chang, C.; Ma, S.; Cheng, Y.; Zhang, J.; Gao, Q. Efficient bioconversion of L-glutamate to gamma-aminobutyric acid by Lactobacillus brevis resting cells. J. Ind. Microbiol. Biotechnol. 2017, 44, 697–704. [Google Scholar] [CrossRef]
- Sun, L.H.; Li, S.N.; Gong, Y.Q. Synthesis of γ-aminobutyric acid by whole cells of Lactobacillus brevis DLF-19076. Food Sci. Technol. 2019, 44, 31–36. (In Chinese) [Google Scholar]
- Gao, D.D.; Chang, K.P.; Ding, G.T.; Wu, H.J.; Chen, Y.H.; Jia, M.Y.; Liu, X.H.; Wang, S.X.; Jin, Y.Y.; Pan, H.; et al. Genomic insights into a robust gamma-aminobutyric acid-producer Lactobacillus brevis CD0817. AMB Express 2019, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Li, H.X.; Cao, Y.S. Pre-staining thin layer chromatography method for amino acid detection. Afr. J. Biotechnol. 2010, 9, 8679–8681. [Google Scholar]
- Li, H.X.; Qiu, T.; Cao, Y.S.; Yang, J.Y.; Huang, Z.B. Pre-staining paper chromatography method for quantification of gamma-aminobutyric acid. J. Chromatogr. A 2009, 1216, 5057–5060. [Google Scholar] [CrossRef]
- Bremer, E.; Kramer, R. Responses of microorganisms to osmotic stress. Annu. Rev. Microbiol. 2019, 73, 313–334. [Google Scholar] [CrossRef]
- Choi, S.I.; Lee, J.W.; Park, S.M.; Lee, M.Y.; Ji, G.E.; Park, M.S.; Heo, T.R. Improvement of gamma-aminobutyric acid (GABA) production using cell entrapment of Lactobacillus brevis GABA 057. J. Microbiol. Biotechnol. 2006, 16, 562–568. [Google Scholar]
- Huang, J.; Mei, L.H.; Wu, H.; Lin, D.Q. Biosynthesis of gamma-aminobutyric acid (GABA) using immobilized whole cells of Lactobacillus brevis. World J. Microbiol. Biotechnol. 2007, 23, 865–871. [Google Scholar] [CrossRef]
- Wu, Q.L.; Shah, N.P. Restoration of GABA production machinery in Lactobacillus brevis by accessible carbohydrates, anaerobiosis and early acidification. Food Microbiol. 2018, 69, 151–158. [Google Scholar] [CrossRef]
- Pourbaferani, M.; Modiri, S.; Norouzy, A.; Maleki, H.; Heidari, M.; Alidoust, L.; Derakhshan, V.; Zahiri, H.S.; Noghabi, K.A. A newly characterized potentially probiotic strain, Lactobacillus brevis MK05, and the toxicity effects of its secretory proteins against MCF-7 breast cancer cells. Probiotics Antimicrob. Proteins 2021, 13, 982–992. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, N.; Werlinger, P.; Suh, D.A.; Lee, H.; Cho, J.H.; Cheng, J. Probiotic characterization of Lactobacillus brevis MJM60390 and in vivo assessment of its antihyperuricemic activity. J. Med. Food 2022, 25, 367–380. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).