
Effects of Lactic Acid Bacteria-Inoculated Corn Silage on Bacterial Communities and Metabolites of Digestive Tract of Sheep

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Silage Making
2.2. Silage Quality Analysis
2.3. Animal Experiments and Sampling
2.4. DNA Extraction PCR Amplification and 16S rDNA Sequencing
2.5. Rumen Metabolites Analysis
2.6. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cholewinska, P.; Gorniak, W.; Wojnarowski, K. Impact of selected environmental factors on microbiome of the digestive tract of ruminants. BMC Vet. Res. 2021, 17, 25. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Fitzsimmons, C.; Plastow, G.; Guan, L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome 2019, 7, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgavi, D.P.; Kelly, W.J.; Janssen, P.H.; Attwood, G.T. Rumen microbial (meta)genomics and its application to ruminant production. Animal 2013, 7, 184–201. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Choi, S.H.; Nogoy, K.M.; Liang, S. Review: The development of the gastrointestinal tract microbiota and intervention in neonatal ruminants. Animal 2021, 15, 100316. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xu, T.; Xu, S.; Ma, L.; Han, X.; Wang, X.; Zhang, X.; Hu, L.; Zhao, N.; Chen, Y.; et al. Effect of dietary concentrate to forage ratio on growth performance, rumen fermentation and bacterial diversity of Tibetan sheep under barn feeding on the Qinghai-Tibetan plateau. Peer. J. 2019, 7, e7462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.E.; Zotti, C.A.; McKinnon, J.J.; McAllister, T.A. Lactobacilli Are Prominent Members of the Microbiota Involved in the Ruminal Digestion of Barley and Corn. Front. Microbiol. 2018, 9, 718. [Google Scholar] [CrossRef] [Green Version]
- Mahanna, B.; Chase, L.E. Practical applications and solutions to silage problems. Silage Science and Technology. Agron. Monogr. 2003, 42, 855–895. [Google Scholar]
- Oliveira, A.S.; Weinberg, Z.G.; Ogunade, I.M.; Cervantes, A.A.P.; Arriola, K.G.; Jiang, Y.; Kim, D.; Li, X.; Gonçalves, M.C.M.; Vyas, D.; et al. Meta-analysis of effects of inoculation with homofermentative and facultative heterofermentative lactic acid bacteria on silage fermentation, aerobic stability, and the performance of dairy cows. J. Dairy Sci. 2017, 100, 4587–4603. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Wang, L.; Wei, Y. Effects of Bacillus amyloliquefaciens and Bacillus pumilus on Rumen and Intestine Morphology and Microbiota in Weanling Jintang Black Goat. Animals 2020, 10, 1604. [Google Scholar] [CrossRef]
- Cherdthong, A.; Suntara, C.; Khota, W.; Wanapat, M. Feed utilization and rumen fermentation characteristics of Thai-indigenous beef cattle fed ensiled rice straw with Lactobacillus casei TH14, molasses, and cellulase enzymes. Livest. Sci. 2021, 245, 104405. [Google Scholar] [CrossRef]
- Basso, F.C.; Adesogan, A.T.; Lara, E.C.; Rabelo, C.H.; Berchielli, T.T.; Teixeira, I.A.; Siqueira, G.R.; Reis, R.A. Effects of feeding corn silage inoculated with microbial additives on the ruminal fermentation, microbial protein yield, and growth performance of lambs. J. Anim. Sci. 2014, 92, 5640–5650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, H.F.; Paula, E.M.; Muck, R.E.; Broderick, G.A.; Faciola, A.P. Effects of lactic acid bacteria in a silage inoculant on ruminal nutrient digestibility, nitrogen metabolism, and lactation performance of high-producing dairy cows. J. Dairy. Sci. 2021, 104, 8826–8834. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Wang, N.; Rinne, M.; Ke, W.; Weinberg, Z.G.; Da, M.; Bai, J.; Zhang, Y.; Li, F.; Guo, X. The bacterial community and metabolome dynamics and their interactions modulate fermentation process of whole crop corn silage prepared with or without inoculants. Microb. Biotechnol. 2021, 14, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Wang, C.; Li, Y.; Yu, Z.; Xu, Q.; Li, G.; Minh, T.T.; Nishino, N. Identification of lactic acid bacteria in the feces of dairy cows fed whole crop maize silage to assess the survival of silage bacteria in the gut. Anim. Sci. J. 2018, 89, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Mu, C.; Xu, Y.; Shen, J.; Zhu, W. Changes in the Solid-, Liquid-, and Epithelium-Associated Bacterial Communities in the Rumen of Hu Lambs in Response to Dietary Urea Supplementation. Front. Microbiol. 2020, 11, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Q.; Xi, Y.M.; Wang, Z.D.; Zeng, H.F.; Han, Z. Combined signature of rumen microbiome and metabolome in dairy cows with different feed intake levels. J. Anim. Sci. 2020, 98, skaa070. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Marlow, UK, 1991. [Google Scholar]
- So, S.; Cherdthong, A.; Wanapat, M.; Uriyapongson, S. Fermented sugarcane bagasse with Lactobacillus combined with cellulase and molasses promotes in vitro gas kinetics, degradability, and ruminal fermentation patterns compared to rice straw. Anim. Biotechnol. 2020, 18, 116–127. [Google Scholar] [CrossRef]
- Kaewpila, C.; Gunun, P.; Kesorn, P.; Subepang, S.; Thip-Uten, S.; Cai, Y.M.; Pholsen, S.; Cherdthong, A.; Khota, W. Improving ensiling characteristics by adding lactic acid bacteria modifies in vitro digestibility and methane production of forage-sorghum mixture silage. Sci. Rep. 2021, 11, 1968. [Google Scholar] [CrossRef]
- Kung, L.; Muck, R.E. Effects of Silage Additives on Ensiling. In Proceedings of the Silage: Field to Feedbunk North American Conference, Hershey, PA, USA, 11–13 February 1997; 99, pp. 187–199. [Google Scholar]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Kung, L., Jr.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef]
- Muck, R.E. A Lactic Acid Bacterial Strain to Improve Aerobic Stability of Silages; Research Summaries; US Dairy Forage Research Center: Madison, WI, USA, 1996; pp. 42–43.
- Guo, X.S.; Ke, W.C.; Ding, W.R.; Ding, L.M.; Xu, D.M.; Wang, W.W.; Zhang, P.; Yang, F.Y. Profiling of metabolome and bacterial community dynamics in ensiled Medicago sativa inoculated without or with Lactobacillus plantarum or Lactobacillus buchneri. Sci. Rep. 2018, 8, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; He, L.; Xing, Y.; Zheng, Y.; Zhou, W.; Pian, R.; Yang, F.; Chen, X.; Zhang, Q. Dynamics of Bacterial Community and Fermentation Quality during Ensiling of Wilted and Unwilted Moringa oleifera Leaf Silage with or without Lactic Acid Bacterial Inoculants. mSphere 2019, 4, e00341-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, Y.; Lin, X.; Wang, Z.; Hou, Q.; Wang, Y.; Hu, Z. High-production dairy cattle exhibit different rumen and fecal bacterial community and rumen metabolite profile than low-production cattle. MicrobiologyOpen 2019, 8, e00673. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, H.F.; Lelis, A.L.J.; Fan, P.; Agustinho, B.C.; Lobo, R.R.; Arce-Cordero, J.A.; Dai, X.; Jeong, K.C.; Faciola, A.P. Effects of lactic acid-producing bacteria as direct-fed microbials on the ruminal microbiome. J. Dairy Sci. 2022, 105, 2242–2255. [Google Scholar] [CrossRef]
- Chen, L.; Li, J.; Dong, Z.; Shao, T. Effects of lactic acid bacteria inoculants and fibrolytic enzymes on the fermentation quality, in vitro degradability, ruminal variables and microbial communities of high-moisture alfalfa silage. Grassl. Sci. 2019, 65, 216–225. [Google Scholar] [CrossRef]
- Chen, L.; Bao, X.; Guo, G.; Huo, W.; Li, Q.; Xu, Q.; Wang, C.; Liu, Q. Evaluation of gallnut tannin and Lactobacillus plantarum as natural modifiers for alfalfa silage: Ensiling characteristics, in vitro ruminal methane production, fermentation profile and microbiota. J. Appl. Microbiol. 2022, 132, 907–918. [Google Scholar] [CrossRef]
- Garro, M.S.; Aguirre, L.; de Giori, G.S. Biological activity of Bifidobacterium longum in response to environmental pH. Appl. Microbiol. Biotechnol. 2006, 70, 612–617. [Google Scholar] [CrossRef]
- Cone, J.W.; Van Gelder, A.H.; Soliman, I.A.; De Visser, H.; Van Vuuren, A.M. Different techniques to study rumen fermentation characteristics of maturing grass and grass silage. J. Dairy Sci. 1999, 82, 957–966. [Google Scholar] [CrossRef]
- Martinez-Fernandez, G.; Jiao, J.; Padmanabha, J.; Denman, S.E.; McSweeney, C.S. Seasonal and Nutrient Supplement Responses in Rumen Microbiota Structure and Metabolites of Tropical Rangeland Cattle. Microorganisms 2020, 8, 1550. [Google Scholar] [CrossRef]
- Hu, Y.; He, Y.; Gao, S.; Liao, Z.; Lai, T.; Zhou, H.; Chen, Q.; Li, L.; Gao, H.; Lu, W. The effect of a diet based on rice straw co-fermented with probiotics and enzymes versus a fresh corn Stover-based diet on the rumen bacterial community and metabolites of beef cattle. Sci. Rep. 2020, 10, 10721. [Google Scholar] [CrossRef]
- Palevich, N.; Kelly, W.J.; Leahy, S.C.; Denman, S.; Altermann, E.; Rakonjac, J.; Attwood, G.T. Comparative Genomics of Rumen Butyrivibrio spp. Uncovers a Continuum of Polysaccharide-Degrading Capabilities. Appl. Environ. Microbiol. 2019, 86, e01993-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, F.S.; Oikonomou, G.; Lima, S.F.; Bicalho, M.L.; Ganda, E.K.; Filho, J.C.; Lorenzo, G.; Trojacanec, P.; Bicalhoa, R.C. Prepartum and postpartum rumen fluid microbiomes: Characterization and correlation with production traits in dairy cows. Appl. Environ. Microbiol. 2015, 81, 1327–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]

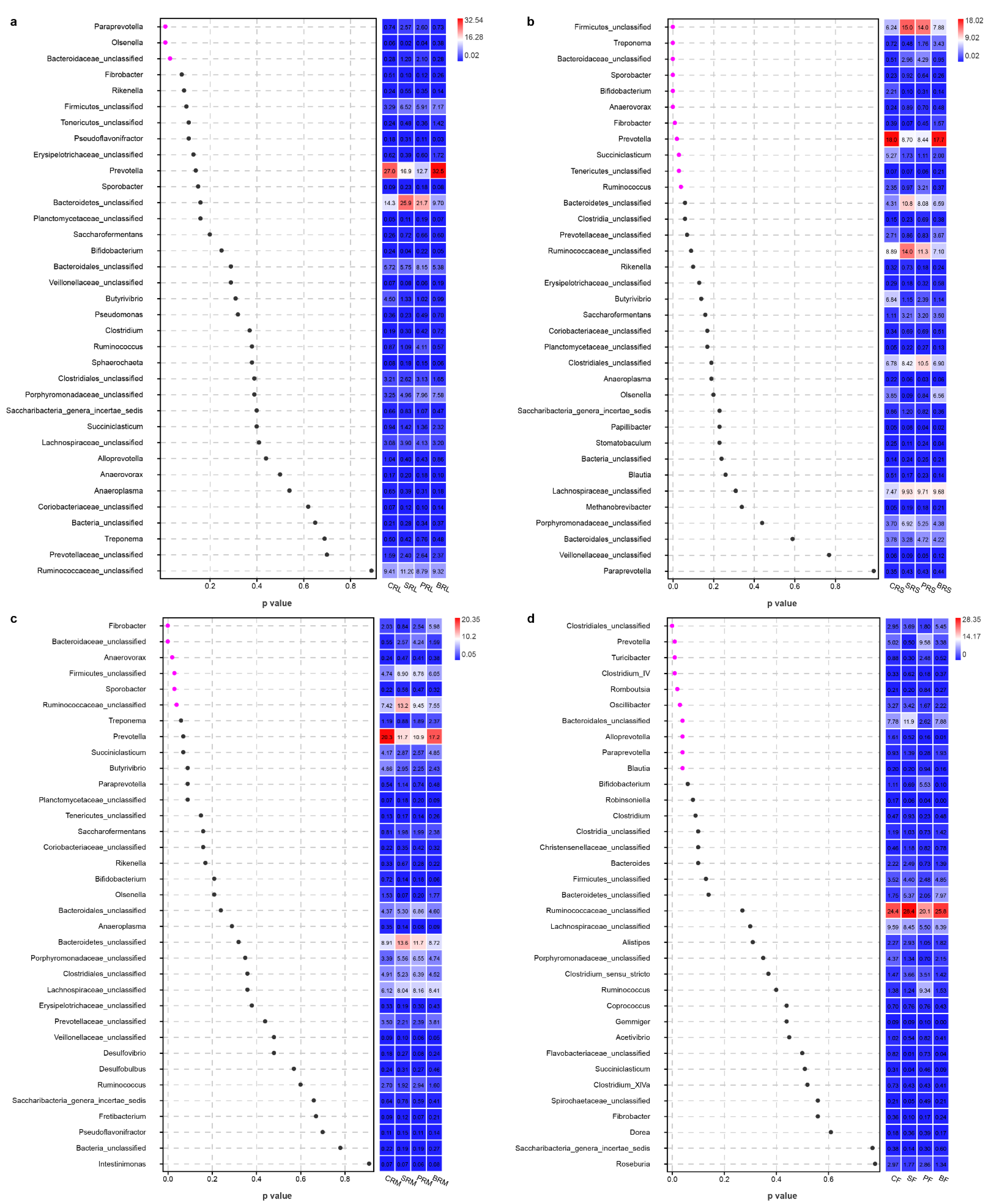


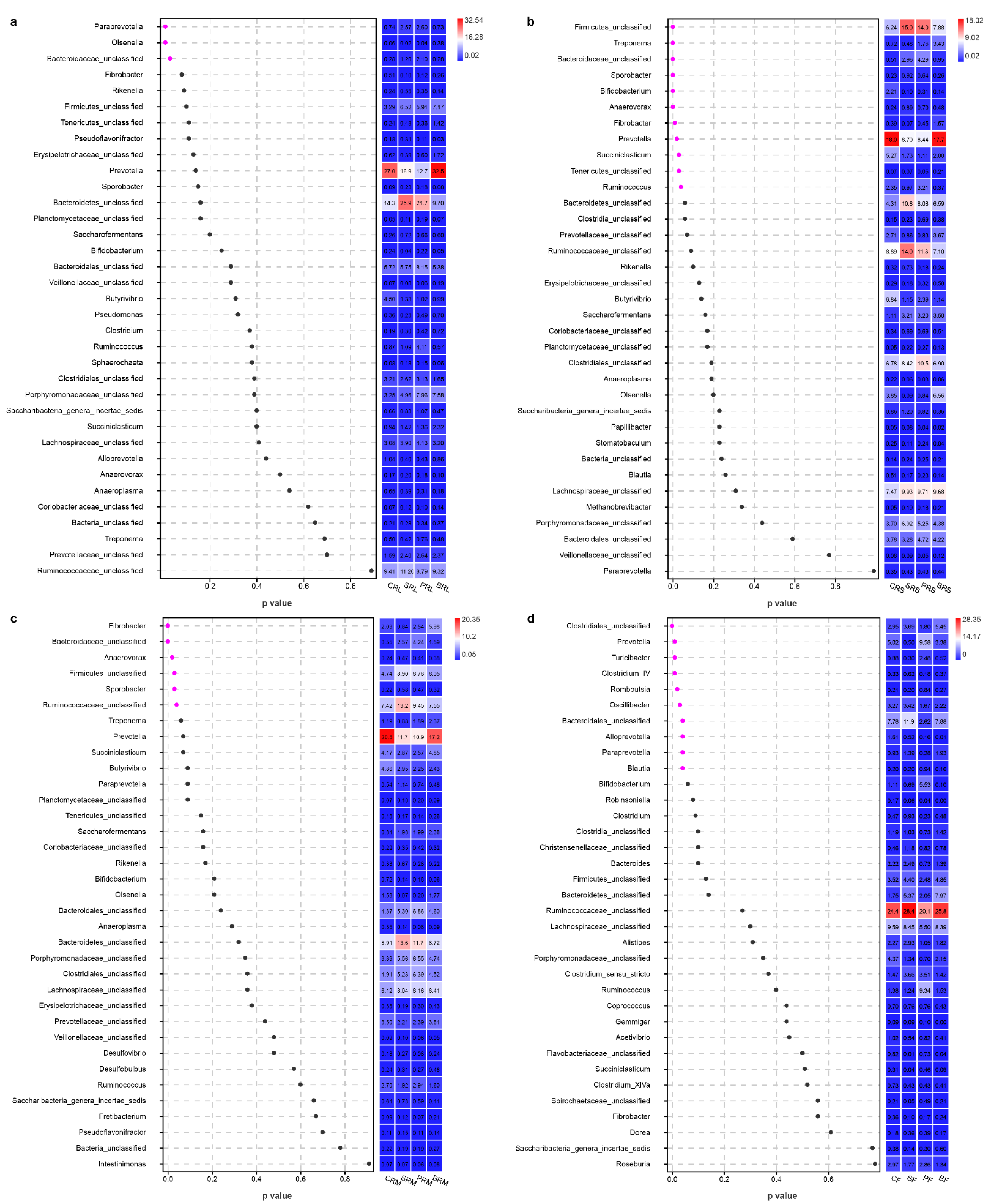
{kind=link}
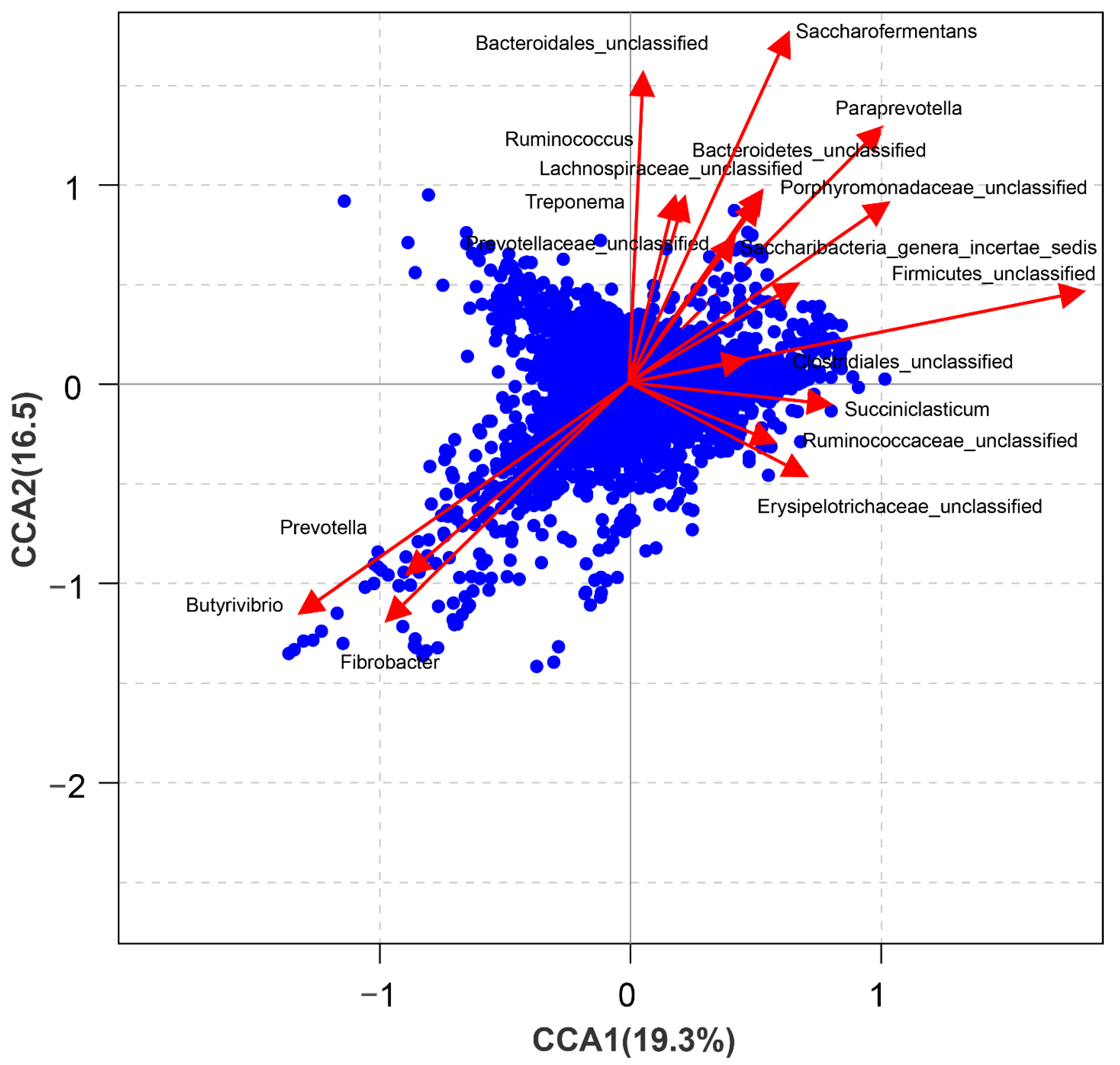
{kind=link}
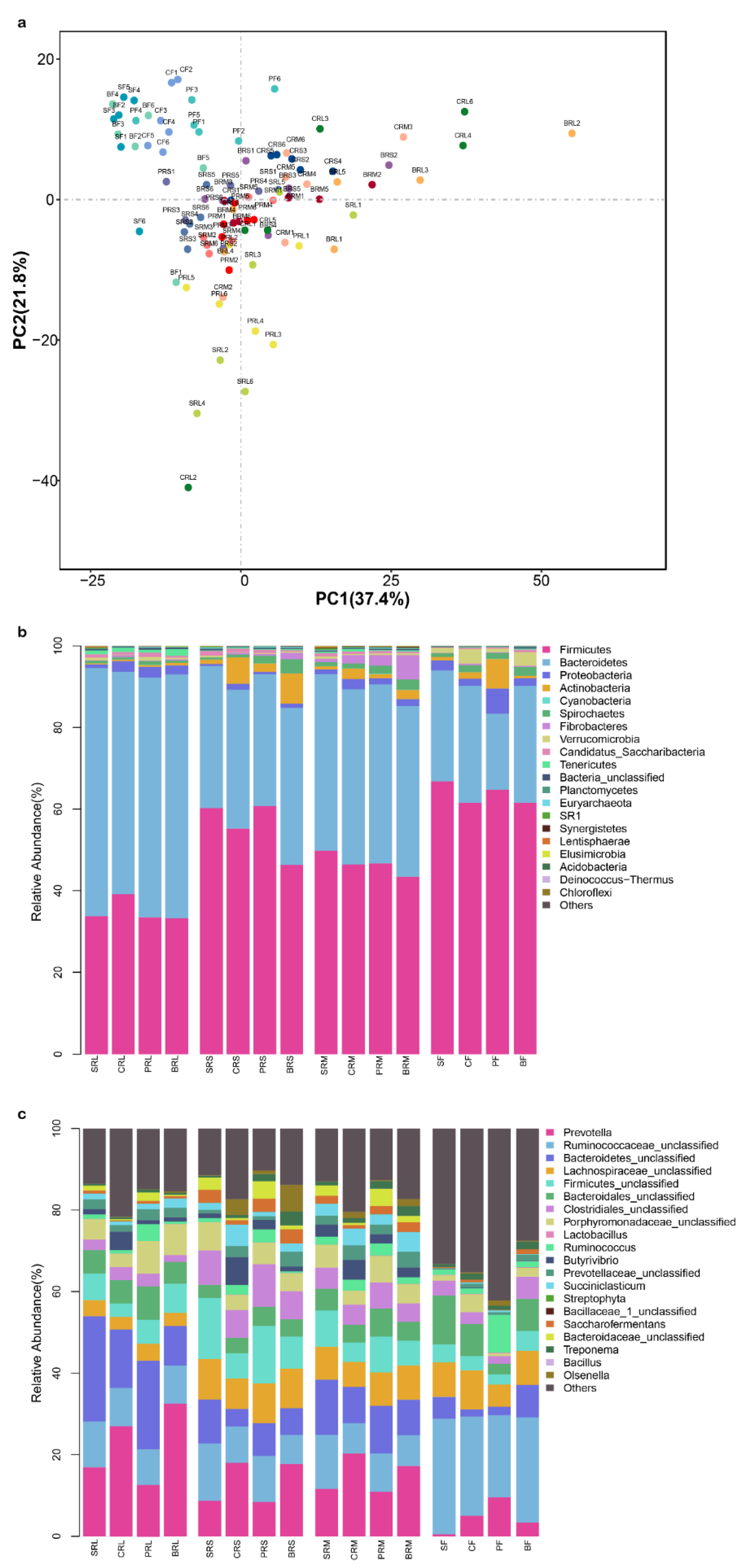
{kind=link}
{kind=link}
| Ingredient Composition (DM%) | S | C | P | B |
|---|---|---|---|---|
| Corn stalk | 40.00 | 0.00 | 0.00 | 0.00 |
| Corn stalk silage | 0.00 | 40.00 | 40.00 | 40.00 |
| Ground corn | 33.00 | 30.73 | 30.73 | 30.73 |
| Rice bran | 2.27 | 11.50 | 11.50 | 11.50 |
| Soybean meal | 6.00 | 5.00 | 5.00 | 5.00 |
| Cottonseed meal | 11.20 | 8.59 | 8.59 | 8.59 |
| Mineral powder | 0.59 | 0.74 | 0.74 | 0.74 |
| Molasses | 6.00 | 2.50 | 2.50 | 2.50 |
| Salt | 0.71 | 0.71 | 0.71 | 0.71 |
| Premix | 0.23 | 0.23 | 0.23 | 0.23 |
| Metabolic energy (MJ/kg DM) | 10.80 | 11.80 | 11.80 | 11.80 |
| CP (%) | 15.00 | 15.10 | 15.10 | 15.10 |
| Ca (%) | 0.45 | 0.49 | 0.49 | 0.49 |
| P (%) | 0.41 | 0.48 | 0.48 | 0.48 |
| NDF (%) | 28.25 | 31.02 | 31.02 | 31.02 |
| ADF (%) | 18.00 | 16.77 | 16.77 | 16.77 |
| Item | C | P | B | SEM | p-Value |
|---|---|---|---|---|---|
| DM (%) | 32.1 b | 37.0 a | 32.9 b | 0.80 | 0.001 |
| pH | 3.89 | 3.93 | 3.86 | 0.01 | 0.089 |
| LAB (log cfu/g) | 3.60 | 5.28 | 4.11 | 0.35 | 0.128 |
| Coliform bacteria (log cfu/g) | <2.00 | <2.00 | <2.00 | <0.01 | 1.00 |
| Lactic acid (%DM) | 5.55 | 3.64 | 5.39 | 0.99 | 0.737 |
| Acetic acid (%DM) | 2.00 | 1.45 | 2.28 | 0.44 | 0.788 |
| Aerobic stability (h) | 68 | 36 | 128 | 17.9 | 0.081 |
| Item | S | C | P | B | SEM | p-Value |
|---|---|---|---|---|---|---|
| Acetic acid (mg/mL) | 2.46 | 2.63 | 2.32 | 2.19 | 0.15 | 0.785 |
| Propionic acid (mg/mL) | 0.66 | 0.66 | 0.62 | 0.65 | 0.05 | 0.989 |
| Butyric acid (mg/mL) | 0.34 | 0.44 | 0.39 | 0.36 | 0.04 | 0.803 |
| pH | 7.26 | 6.73 | 6.94 | 6.83 | 0.09 | 0.157 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, H.; Wang, C.; Huang, Z.; Zhang, Y.; Sun, L.; Xue, Y.; Guo, X. Effects of Lactic Acid Bacteria-Inoculated Corn Silage on Bacterial Communities and Metabolites of Digestive Tract of Sheep. Fermentation 2022, 8, 320. https://doi.org/10.3390/fermentation8070320
Han H, Wang C, Huang Z, Zhang Y, Sun L, Xue Y, Guo X. Effects of Lactic Acid Bacteria-Inoculated Corn Silage on Bacterial Communities and Metabolites of Digestive Tract of Sheep. Fermentation. 2022; 8(7):320. https://doi.org/10.3390/fermentation8070320
Chicago/Turabian StyleHan, Hongyan, Chao Wang, Zhipeng Huang, Ying Zhang, Lin Sun, Yanlin Xue, and Xusheng Guo. 2022. "Effects of Lactic Acid Bacteria-Inoculated Corn Silage on Bacterial Communities and Metabolites of Digestive Tract of Sheep" Fermentation 8, no. 7: 320. https://doi.org/10.3390/fermentation8070320
APA StyleHan, H., Wang, C., Huang, Z., Zhang, Y., Sun, L., Xue, Y., & Guo, X. (2022). Effects of Lactic Acid Bacteria-Inoculated Corn Silage on Bacterial Communities and Metabolites of Digestive Tract of Sheep. Fermentation, 8(7), 320. https://doi.org/10.3390/fermentation8070320