
Bioconversion of Ginsenosides in American Ginseng Extraction Residue by Fermentation with Ganoderma lucidum Improves Insulin-like Glucose Uptake in 3T3-L1 Adipocytes

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Materials
2.3. Total Phenol Content Determination
2.4. Glucosidase Activity
2.5. Cell Culture and Glucose Uptake
2.6. Statistical Analysis
3. Results
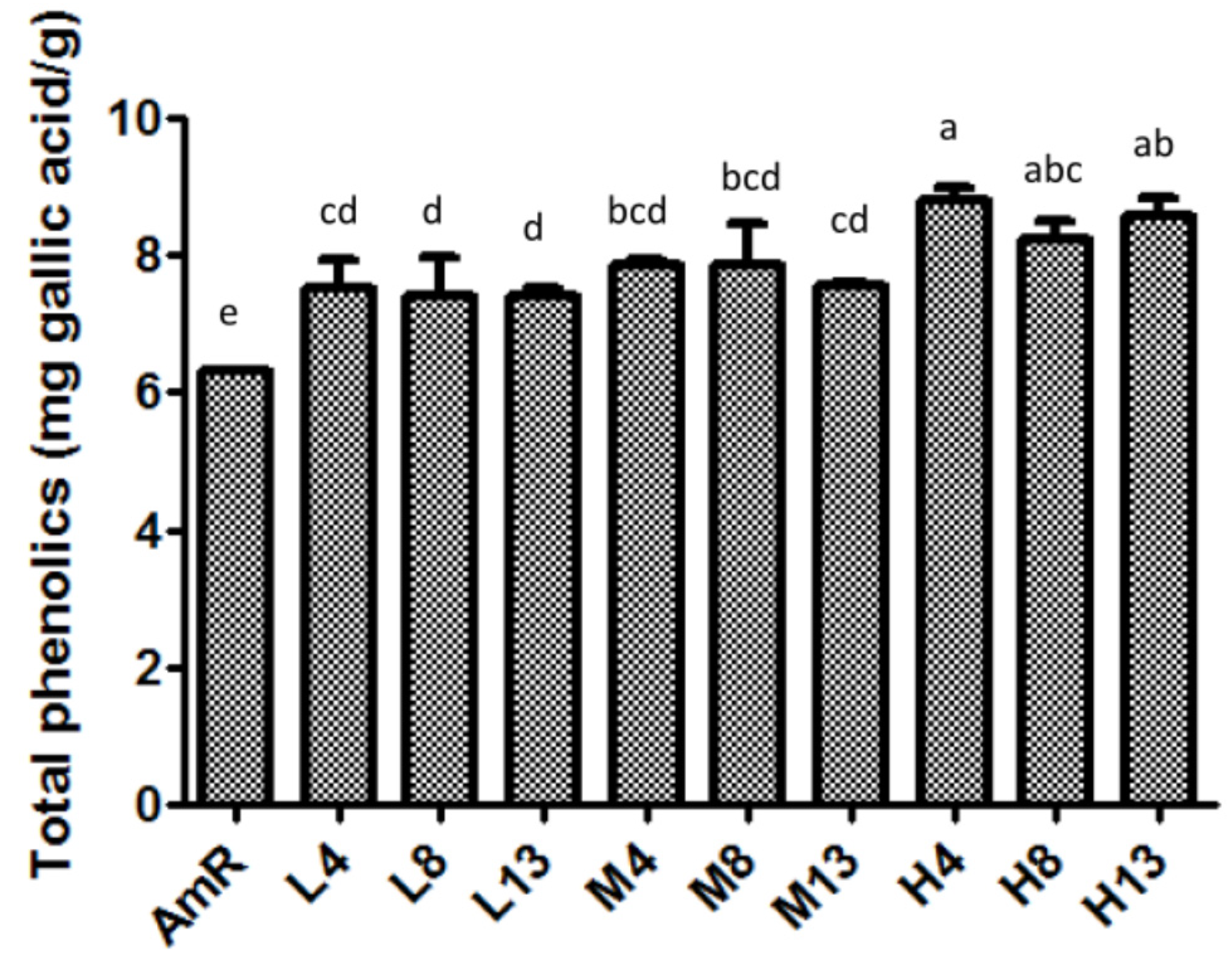
3.1. Total Phenolic Contents of AmR and FAmR
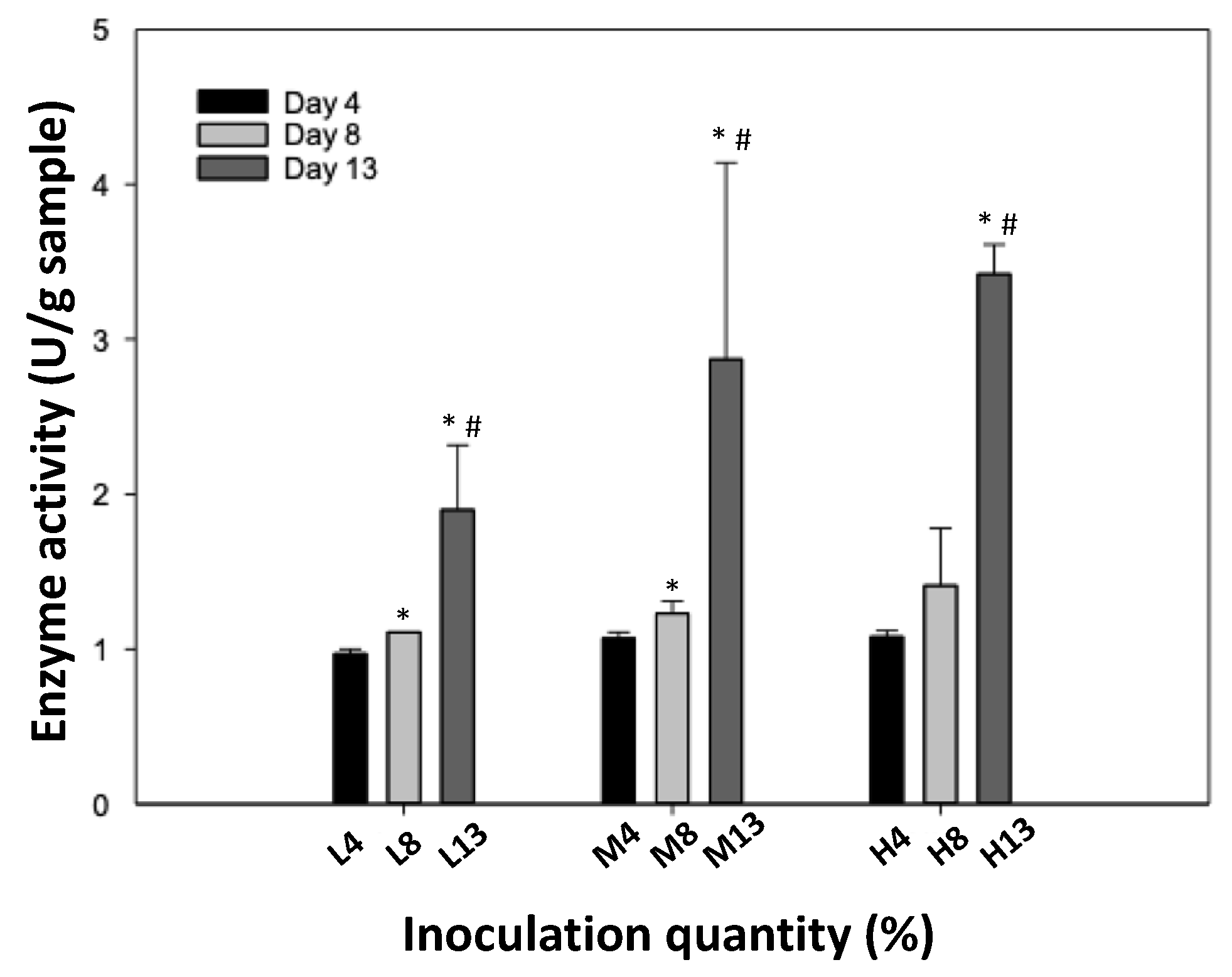
3.2. Glucosidase Activity
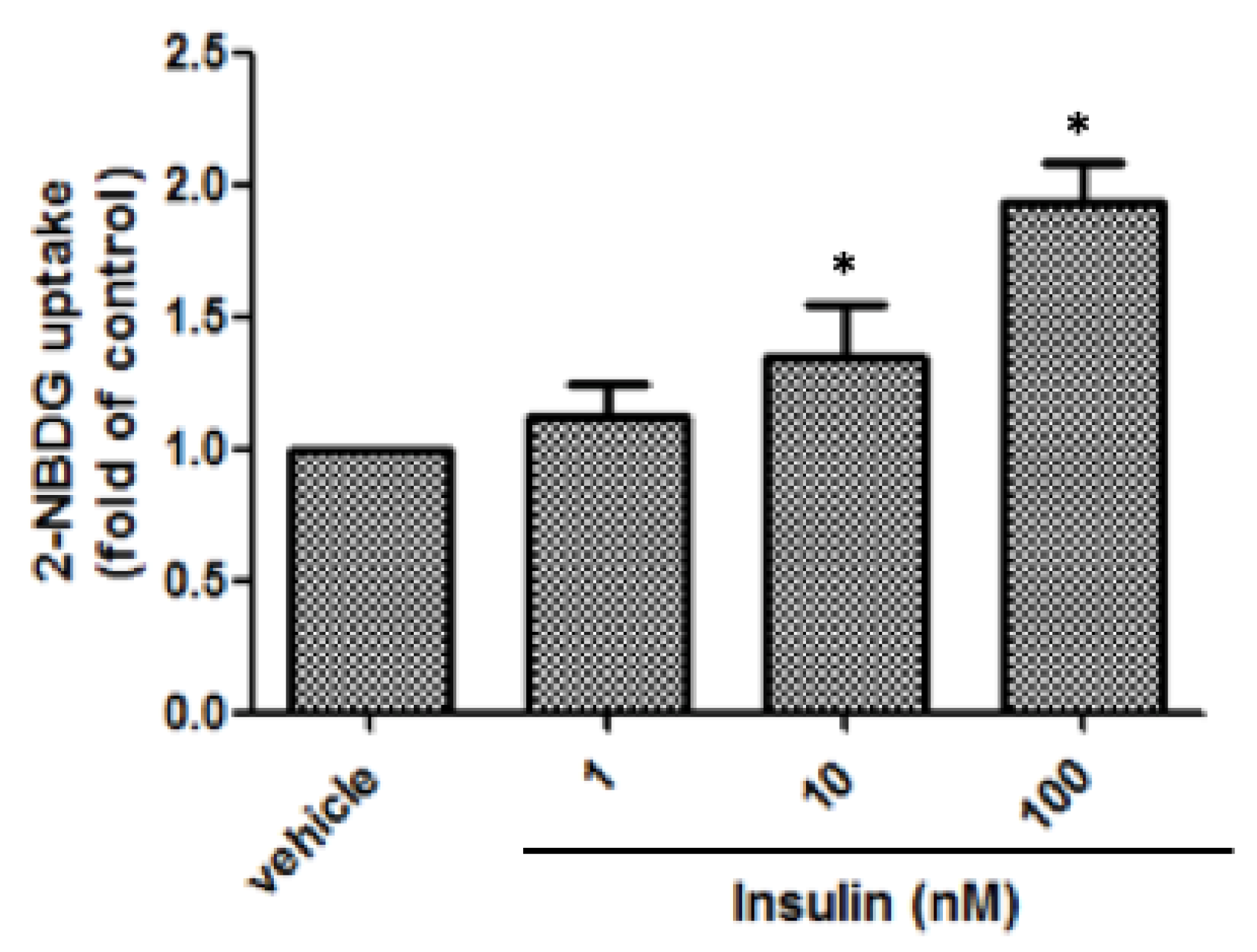
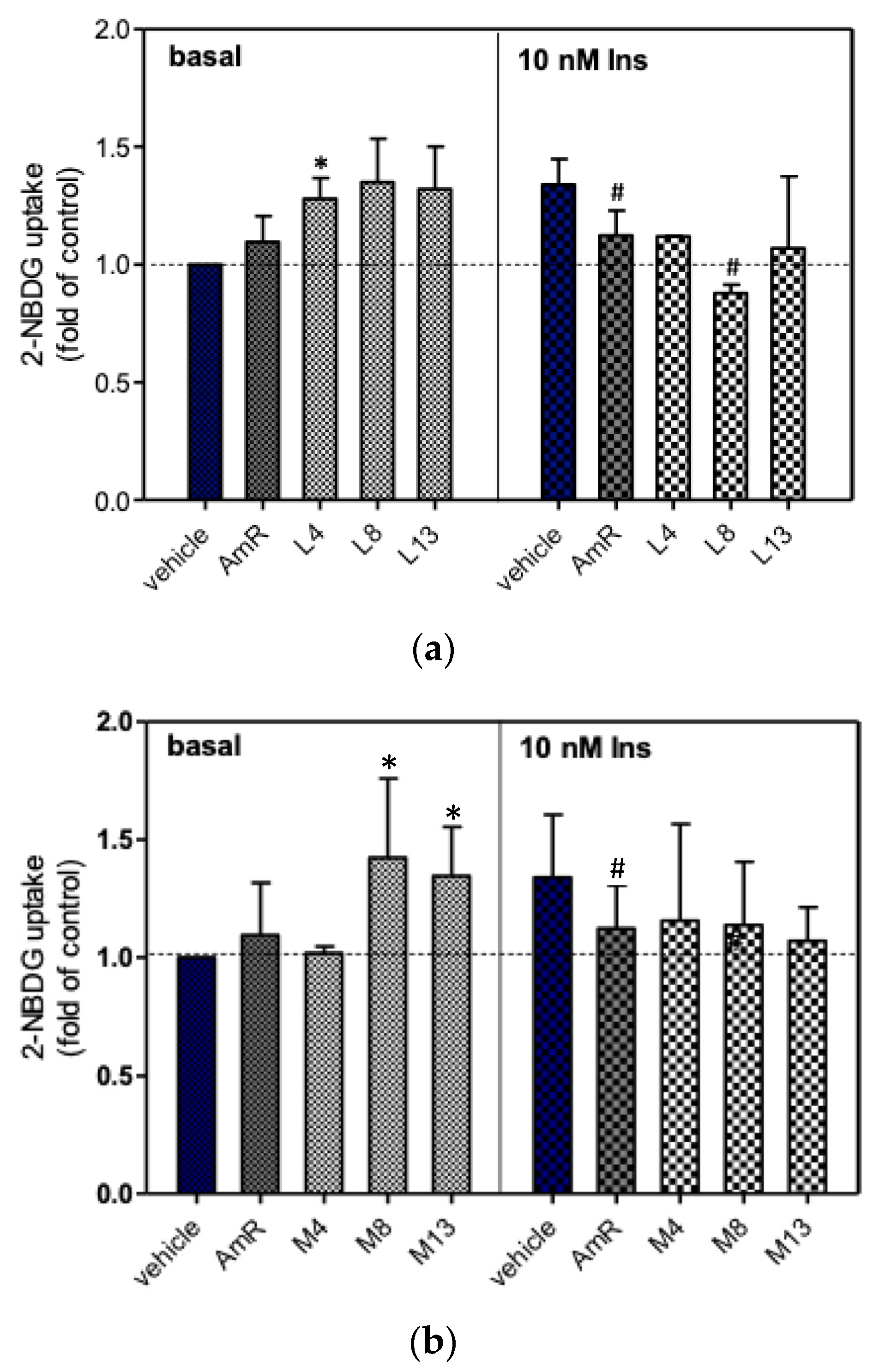
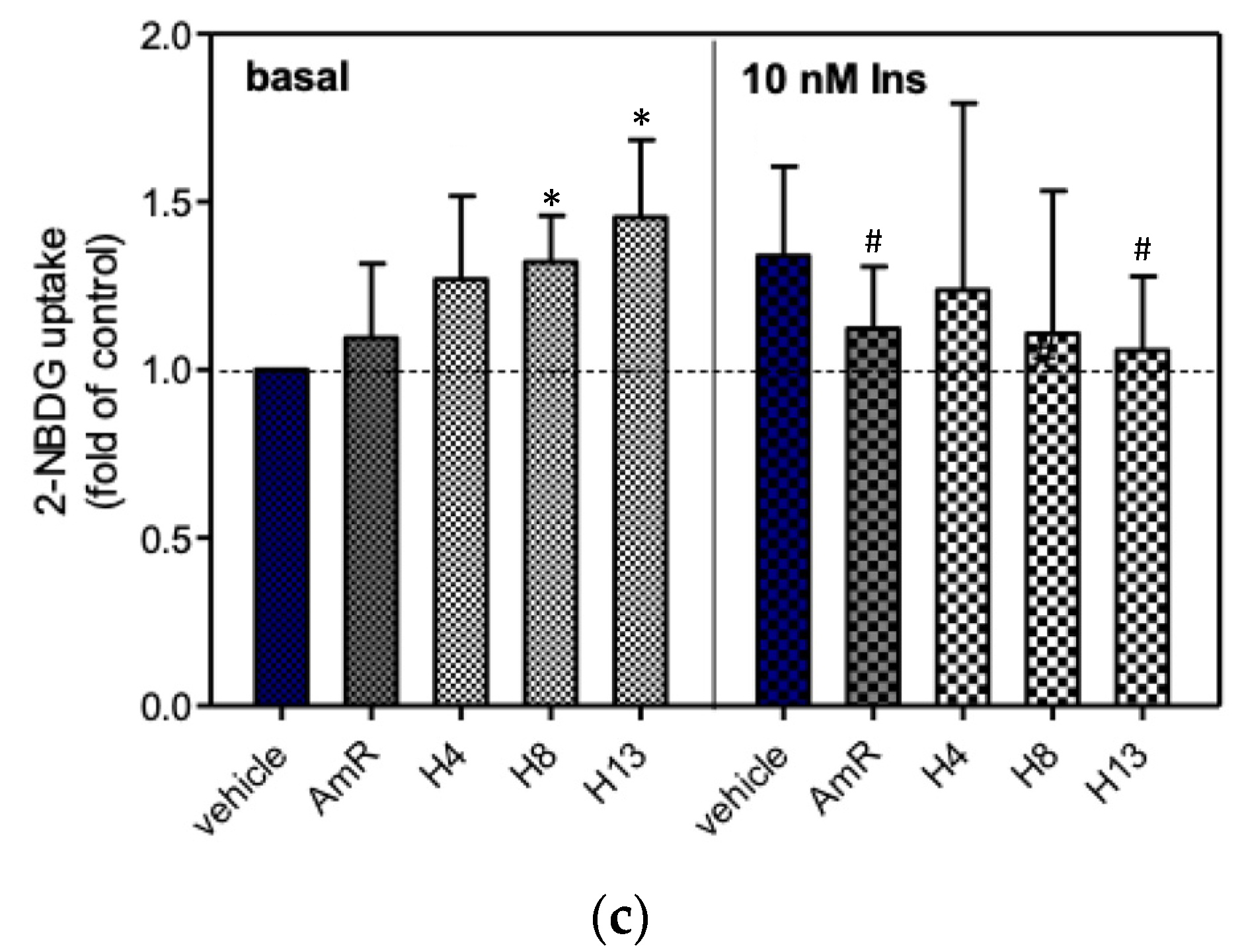
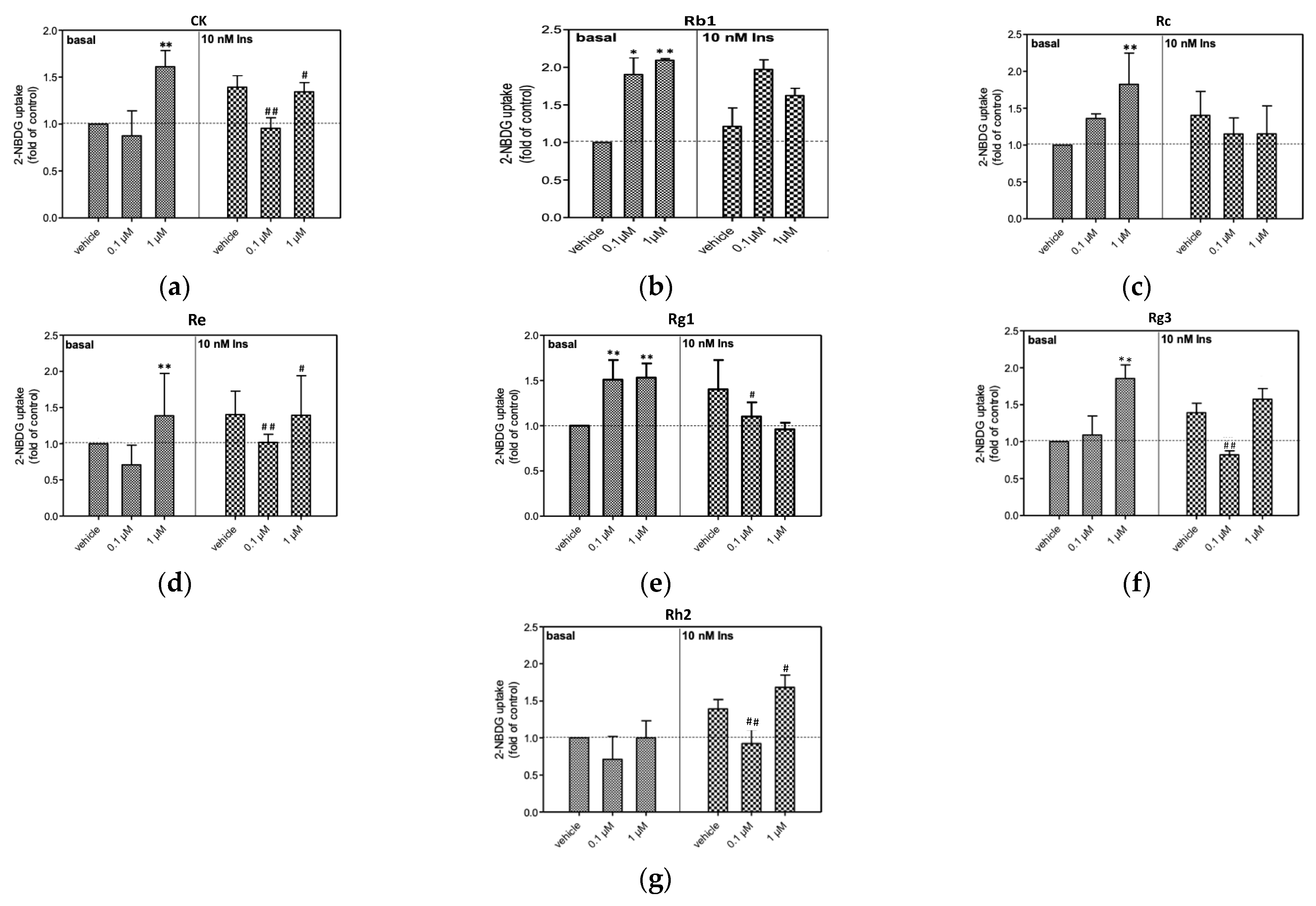
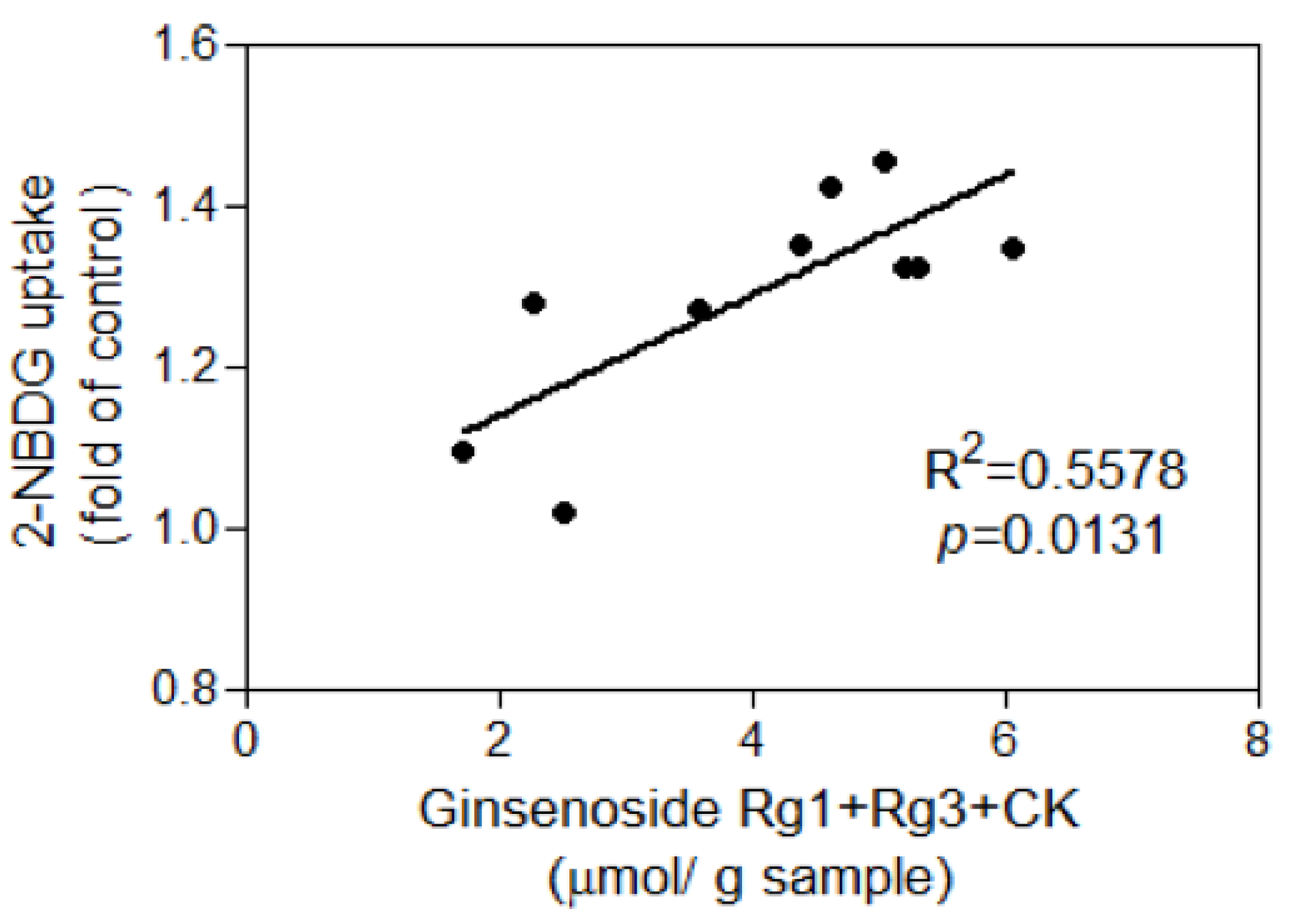
3.3. Glucose Uptake of 3T3-L1 Adipocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shaw, P.C.; But, P.P.H. Authentication of panax species and their adulterants by random-primed polymerase chain-reaction. Planta Med. 1995, 61, 466–469. [Google Scholar] [CrossRef]
- Liu, Z.Q. Chemical insights into ginseng as a resource for natural antioxidants. Chem. Rev. 2012, 112, 3329–3355. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.M.; Xin, Y.Z.; Li, Y.J.; Xu, F.X.; Xi, X.Z.; Guo, H.; Cui, X.W.; Cao, H.; Zhang, X.; Han, C.C. Ginsenosides: A potential neuroprotective agent. Biomed. Res. Int. 2018, 2018, 8174345. [Google Scholar] [CrossRef]
- Yi, Y.S. Roles of ginsenosides in inflammasome activation. J. Ginseng Res. 2019, 43, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.X.; Huang, Y.L.; Zheng, H.; Li, S.Q.; Li, Z.H.; Yuan, L.; Cheng, X.; He, C.S.; Sun, J.F. Ginsenosides for the treatment of metabolic syndrome and cardiovascular diseases: Pharmacology and mechanisms. Biomed. Pharmacother. 2020, 132, 110915. [Google Scholar] [CrossRef]
- Park, S.K.; Hyun, S.H.; In, G.; Park, C.K.; Kwak, Y.S.; Jang, Y.J.; Kim, B.; Kim, J.H.; Han, C.K. The antioxidant activities of Korean Red Ginseng (Panax ginseng) and ginsenosides: A systemic review through in vivo and clinical trials. J. Ginseng Res. 2021, 45, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.S.T.; Che, C.M.; Leung, K.W. Recent advances in ginseng as cancer therapeutics: A functional and mechanistic overview. Nat. Prod. Rep. 2015, 32, 256–272. [Google Scholar] [CrossRef]
- Chen, W.; Balan, P.; Popovich, D.G. Review of ginseng anti-diabetic studies. Molecules 2019, 24, 4501. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yang, M.F.; Su, Y.P.; Jiang, H.M.; You, X.J.; Yang, Y.J.; Zhang, H.L. Ginsenoside Re reduces insulin resistance through activation of PPAR-gamma pathway and inhibition of TNF-alpha production. J. Ethnopharmacol. 2013, 147, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Ding, L.; Zhang, H.Q.; Chu, Y.F.; Chang, Z.H.; Yu, Y.L.; Guo, D.D.; Zhang, S.P.; Liu, X.Z. Ginsenoside Rb1 increases insulin sensitivity through suppressing 11 beta-hydroxysteroid dehydrogenase type I. Am. J. Transl. Res. 2017, 9, 1049–1057. [Google Scholar]
- Kim, K.S.; Yang, H.J.; Lee, I.S.; Kim, K.H.; Park, J.; Jeong, H.S.; Kim, Y.; Ahn, K.S.; Na, Y.C.; Jang, H.J. The aglycone of ginsenoside Rg3 enables glucagon-like peptide-1 secretion in enteroendocrine cells and alleviates hyperglycemia in type 2 diabetic mice. Sci. Rep. 2015, 5, 18325. [Google Scholar] [CrossRef]
- Jenkins, A.L.; Morgan, L.M.; Bishop, J.; Jovanovski, E.; Jenkins, D.J.A.; Vuksan, V. Co-administration of a konjac-based fibre blend and American ginseng (Panax quinquefolius L.) on glycaemic control and serum lipids in type 2 diabetes: A randomized controlled, cross-over clinical trial. Eur. J. Nutr. 2018, 57, 2217–2225. [Google Scholar] [CrossRef]
- Vuksan, V.; Xu, Z.Z.; Jovanovski, E.; Jenkins, A.L.; Beljan-Zdravkovic, U.; Sievenpiper, J.L.; Mark Stavro, P.; Zurbau, A.; Duvnjak, L.; Li, M.Z.C. Efficacy and safety of American ginseng (Panax quinquefolius L.) extract on glycemic control and cardiovascular risk factors in individuals with type 2 diabetes: A double-blind, randomized, cross-over clinical trial. Eur. J. Nutr. 2019, 58, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Lee, J.W.; Lee, K.Y.; Yang, D.C. Microbial conversion of major ginsenoside Rb-1 to pharmaceutically active minor ginsenoside Rd. J. Microbiol. 2005, 43, 456–462. [Google Scholar]
- Choi, H.J.; Kim, E.A.; Kim, D.H.; Shin, K.S. The bioconversion of red ginseng ethanol extract into compound K by saccharomyces cerevisiae HJ-014. Mycobiology 2014, 42, 256–261. [Google Scholar] [CrossRef]
- Zheng, M.M.; Xu, F.X.; Li, Y.J.; Xi, X.Z.; Cui, X.W.; Han, C.C.; Zhang, X.L. Study on transformation of ginsenosides in different methods. Biomed. Res. Int. 2017, 2017, 8601027. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.P.; Jin, Y.; Yin, C.R.; Bai, L.L. Co-transformation of Panax major ginsenosides Rb-1 and Rg(1) to minor ginsenosides C-K and F-1 by Cladosporium cladosporioides. J. Ind. Microbiol. Biotechnol. 2012, 39, 521–527. [Google Scholar] [CrossRef]
- Lu, J.H.; He, R.J.; Sun, P.L.; Zhang, F.M.; Linhardt, R.J.; Zhang, A.Q. Molecular mechanisms of bioactive polysaccharides from Ganoderma lucidum (Lingzhi), a review. Int. J. Biol. Macromol. 2020, 150, 765–774. [Google Scholar] [CrossRef]
- Songulashvili, G.; Elisashvili, V.; Wasser, S.P.; Nevo, E.; Hadar, Y. Basidiomycetes laccase and manganese peroxidase activity in submerged fermentation of food industry wastes. Enzym. Microb. Technol. 2007, 41, 57–61. [Google Scholar] [CrossRef]
- Lee, S.H.; Oh, M.; Park, J.; Jang, S.Y.; Cheong, S.H.; Lee, H.; Moon, S.H. Antioxidant and anti-inflammatory activities of the ethanolic extract of fermented red ginseng marc. Food Sci. Biotechnol. 2015, 24, 651–657. [Google Scholar] [CrossRef]
- Hsu, B.Y.; Chen, C.H.; Lu, T.J.; Hwang, L.S. Bioconversion of ginsenosides in the american ginseng (xi yang shen) extraction residue by fermentation with lingzhi (ling zhi, ganoderma lucidum). J. Tradit. Complement. Med. 2013, 3, 95–101. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.Z.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, M.; Obata, A. Beta-glucosidases from soybeans hydrolyze daidzin and genistin. J. Food Sci. 1993, 58, 144–147. [Google Scholar] [CrossRef]
- Xu, Q.F.; Fang, X.L.; Chen, D.F. Pharmacokinetics and bioavailability of ginsenoside Rb-1 and Rg(1) from Panax notoginseng in rats. J. Ethnopharmacol. 2003, 84, 187–192. [Google Scholar] [CrossRef]
- Yu, K.; Chen, F.; Li, C. Absorption, disposition, and pharmacokinetics of saponins from Chinese medicinal herbs: What do we know and what do we need to know more? Curr. Drug Metab. 2012, 13, 577–598. [Google Scholar] [CrossRef]
- Senthil, K.; Veena, V.; Mahalakshmi, M.; Pulla, R.; Yang, D.C.; Parvatham, R. Microbial conversion of major ginsenoside Rb1 to minor ginsenoside Rd by Indian fermented food bacteria. Afr. J. Biotechnol. 2009, 8, 6961–6966. [Google Scholar]
- Son, J.W.; Kim, H.J.; Oh, D.K. Ginsenoside Rd production from the major ginsenoside Rb-1 by beta-glucosidase from Thermus caldophilus. Biotechnol. Lett. 2008, 30, 713–716. [Google Scholar] [CrossRef]
- Ko, S.R.; Choi, K.J.; Suzuki, K.; Suzuki, Y. Enzymatic preparation of ginsenosides Rg(2), Rh-1, and F-1(1)). Chem. Pharm. Bull. 2003, 51, 404–408. [Google Scholar] [CrossRef]
- Park, C.S.; Yoo, M.H.; Noh, K.H.; Oh, D.K. Biotransformation of ginsenosides by hydrolyzing the sugar moieties of ginsenosides using microbial glycosidases. Appl. Microbiol. Biot. 2010, 87, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Lin, C.Y.; Huang, S.F.; Lin, H.C.; Chang, W.L.; Chang, T.C. Effect and mechanism of ginsenosides CK and Rg1 on stimulation of glucose uptake in 3T3-L1 adipocytes. J. Agric. Food Chem. 2010, 58, 6039–6047. [Google Scholar] [CrossRef]
- Lee, O.H.; Lee, H.H.; Kim, J.H.; Lee, B.Y. Effect of ginsenosides Rg3 and Re on glucose transport in mature 3T3-L1 adipocytes. Phytother. Res. 2011, 25, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Shang, W.B.; Yang, Y.; Zhou, L.B.; Jiang, B.R.; Jin, H.; Chen, M.D. Ginsenoside Rb-1 stimulates glucose uptake through insulin-like signaling pathway in 3T3-L1 adipocytes. J. Endocrinol. 2008, 198, 561–569. [Google Scholar] [CrossRef]
- Bai, L.T.; Gao, J.L.; Wei, F.; Zhao, J.; Wang, D.W.; Wei, J.P. Therapeutic potential of ginsenosides as an adjuvant treatment for diabetes. Front. Pharmacol. 2018, 9, 423. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.W.; Jiang, J.L.; Zou, J.J.; Yang, M.Y.; Chen, F.M.; Zhang, Y.J.; Jia, L. Therapeutic potential of ginsenosides on diabetes: From hypoglycemic mechanism to clinical trials. J. Funct. Foods 2020, 64, 103630. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, S.G.; Chae, I.G.; Kim, M.J.; Im, N.K.; Yu, M.H.; Lee, E.J.; Lee, I.S. Antioxidant Effects of Fermented Red Ginseng Extracts in Streptozotocin-Induced Diabetic Rats. J. Ginseng Res. 2011, 35, 129–137. [Google Scholar] [CrossRef]
- Malathy, R.; Prabakaran, M.; Kalaiselvi, K.; Chung, I.M.; Kim, S.H. Comparative polyphenol composition, antioxidant and anticorrosion properties in various parts of panax ginseng extracted in different solvents. Appl. Sci. 2021, 11, 93. [Google Scholar] [CrossRef]
- Vinayagam, R.; Jayachandran, M.; Xu, B.J. Antidiabetic effects of simple phenolic acids: A comprehensive review. Phytother. Res. 2016, 30, 184–199. [Google Scholar] [CrossRef]
- Ghorbani, A. Mechanisms of antidiabetic effects of flavonoid rutin. Biomed. Pharmacother. 2017, 96, 305–312. [Google Scholar] [CrossRef]
- Chen, R.; Qi, Q.L.; Wang, M.T.; Li, Q.Y. Therapeutic potential of naringin: An overview. Pharm. Biol. 2016, 54, 3203–3210. [Google Scholar] [CrossRef] [PubMed]
- Roffey, B.; Atwal, A.; Kubow, S. Cinnamon water extracts increase glucose uptake but inhibit adiponectin secretion in 3T3-L1 adipose cells. Mol. Nutr. Food Res. 2006, 50, 739–745. [Google Scholar] [CrossRef]
- Liu, F.; Kim, J.K.; Li, Y.H.; Liu, X.Q.; Li, J.; Chen, X.H. An extract of Lagerstroemia speciosa L. has insulin-like glucose uptake-stimulatory and adipocyte differentiation-inhibitory activities in 3T3-L1 cells. J. Nutr. 2001, 131, 2242–2247. [Google Scholar] [CrossRef]
- Liu, H.M.; Liu, M.H.; Jin, Z.B.; Yaqoob, S.; Zheng, M.Z.; Cai, D.; Liu, J.S.; Guo, S.D. Ginsenoside Rg2 inhibits adipogenesis in 3T3-L1 preadipocytes and suppresses obesity in high-fat-diet-induced obese mice through the AMPK pathway. Food Funct. 2019, 10, 3603–3614. [Google Scholar] [CrossRef] [PubMed]
- Salt, I.P.; Connell, J.M.C.; Gould, G.W. 5-aminoimidazole-4-carboxamide ribonucleoside (AICAR) inhibits insulin-stimulated glucose transport in 3T3-L1 adipocytes. Diabetes 2000, 49, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Sakoda, H.; Ogihara, T.; Anai, M.; Fujishiro, M.; Ono, H.; Onishi, Y.; Katagiri, H.; Abe, M.; Fukushima, Y.; Shojima, N.; et al. Activation of AMPK is essential for AICAR-induced glucose uptake by skeletal muscle but not adipocytes. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E1239–E1244. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, M.L.; Hu, J.J.; Zhao, W.W.; Gao, X.J.; Jiang, C.H.; Liu, K.; Liu, B.L.; Huang, F. Quercetin differently regulates insulin-mediated glucose transporter 4 translocation under basal and inflammatory conditions in adipocytes. Mol. Nutr. Food Res. 2014, 58, 931–941. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
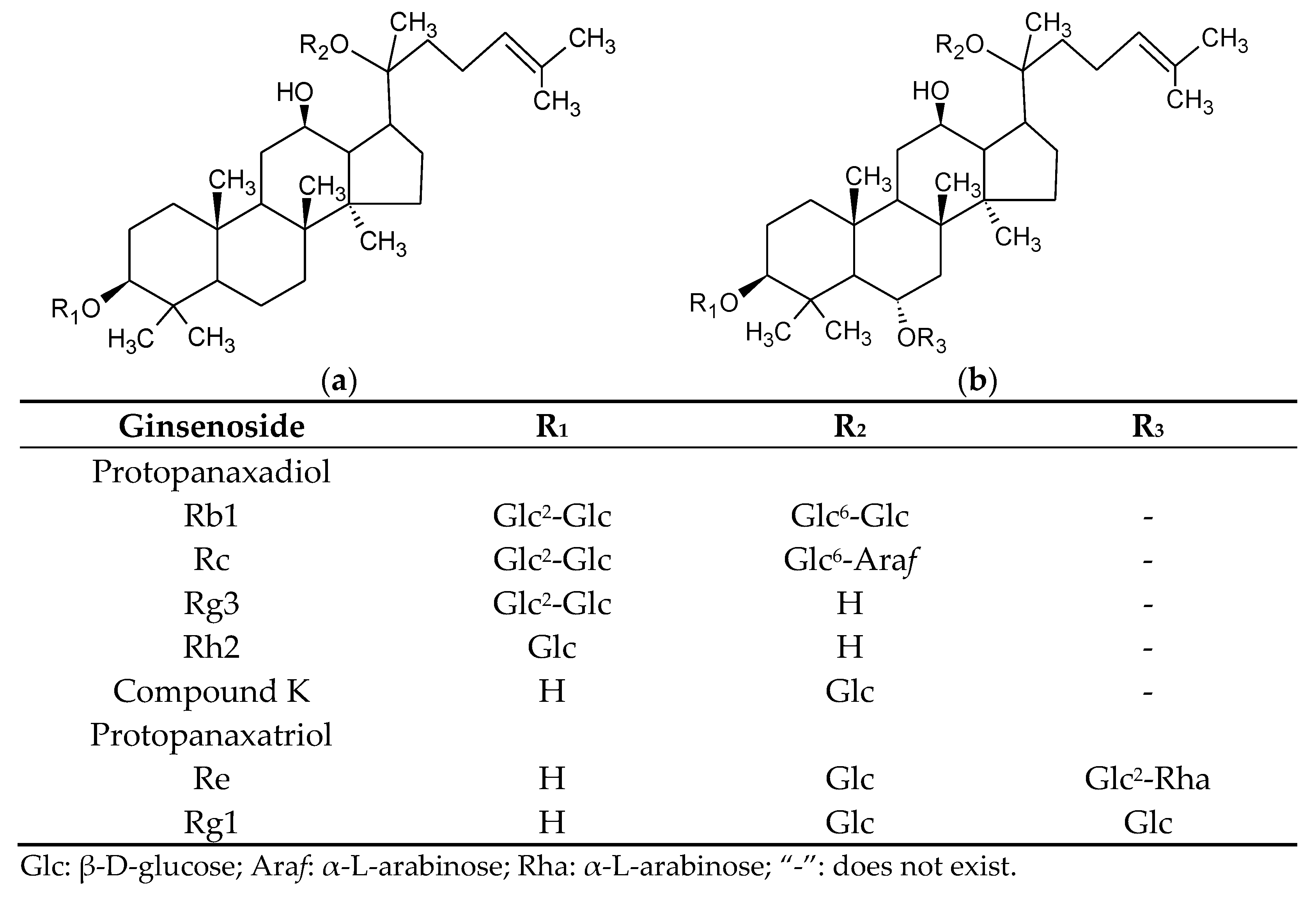
| Percentage of the Sum of Seven Ginsenosides | |||
|---|---|---|---|
| Rg1 | Rg3 | CK | |
| AmR | 0.00 ± 0.00 | 1.69 ± 0.25 | 0.16 ± 0.03 |
| L4 | 0.00 ± 0.00 | 11.41 ± 0.43 ** | 0.00 ± 0.00 ** |
| L8 | 7.71 ± 10.91 | 14.89 ± 1.63 ** | 0.07 ± 0.10 |
| L13 | 9.31 ± 12.69 | 17.15 ± 5.19 ** | 0.10 ± 0.14 |
| M4 | 0.00 ± 0.00 | 13.05 ± 0.91 *** | 0.00 ± 0.00 ** |
| M8 | 0.20 ± 0.35 | 21.98 ± 2.76 ** | 0.43 ± 0.39 |
| M13 | 14.39 ± 4.19 * | 18.78 ± 1.56 *** | 2.09 ± 0.83 |
| H4 | 1.35 ± 2.33 | 10.97 ± 1.35 ** | 0.00 ± 0.00 ** |
| H8 | 2.09 ± 0.55 * | 26.20 ± 5.50 * | 0.57 ± 0.70 |
| H13 | 10.88 ± 1.23 ** | 17.39 ± 2.90 * | 1.88 ± 1.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, B.-Y.; Chen, C.-H.; Lu, T.-J.; Pan, M.-H.; Ho, C.-T.; Hwang, L.S.; Hung, W.-L. Bioconversion of Ginsenosides in American Ginseng Extraction Residue by Fermentation with Ganoderma lucidum Improves Insulin-like Glucose Uptake in 3T3-L1 Adipocytes. Fermentation 2021, 7, 297. https://doi.org/10.3390/fermentation7040297
Hsu B-Y, Chen C-H, Lu T-J, Pan M-H, Ho C-T, Hwang LS, Hung W-L. Bioconversion of Ginsenosides in American Ginseng Extraction Residue by Fermentation with Ganoderma lucidum Improves Insulin-like Glucose Uptake in 3T3-L1 Adipocytes. Fermentation. 2021; 7(4):297. https://doi.org/10.3390/fermentation7040297
Chicago/Turabian StyleHsu, Bo-Yang, Chia-Hui Chen, Ting-Jang Lu, Min-Hsiung Pan, Chi-Tang Ho, Lucy Sun Hwang, and Wei-Lun Hung. 2021. "Bioconversion of Ginsenosides in American Ginseng Extraction Residue by Fermentation with Ganoderma lucidum Improves Insulin-like Glucose Uptake in 3T3-L1 Adipocytes" Fermentation 7, no. 4: 297. https://doi.org/10.3390/fermentation7040297
APA StyleHsu, B.-Y., Chen, C.-H., Lu, T.-J., Pan, M.-H., Ho, C.-T., Hwang, L. S., & Hung, W.-L. (2021). Bioconversion of Ginsenosides in American Ginseng Extraction Residue by Fermentation with Ganoderma lucidum Improves Insulin-like Glucose Uptake in 3T3-L1 Adipocytes. Fermentation, 7(4), 297. https://doi.org/10.3390/fermentation7040297