
Biomethanation of Carbon Monoxide by Hyperthermophilic Artificial Archaeal Co-Cultures

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Media
2.3. Strains and Cultivation Conditions
2.4. Pure-Culture Closed Batch Experiments
2.5. Co-Culture Closed Batch Experiments
2.6. Sampling
2.7. Analytical Procedures
3. Results
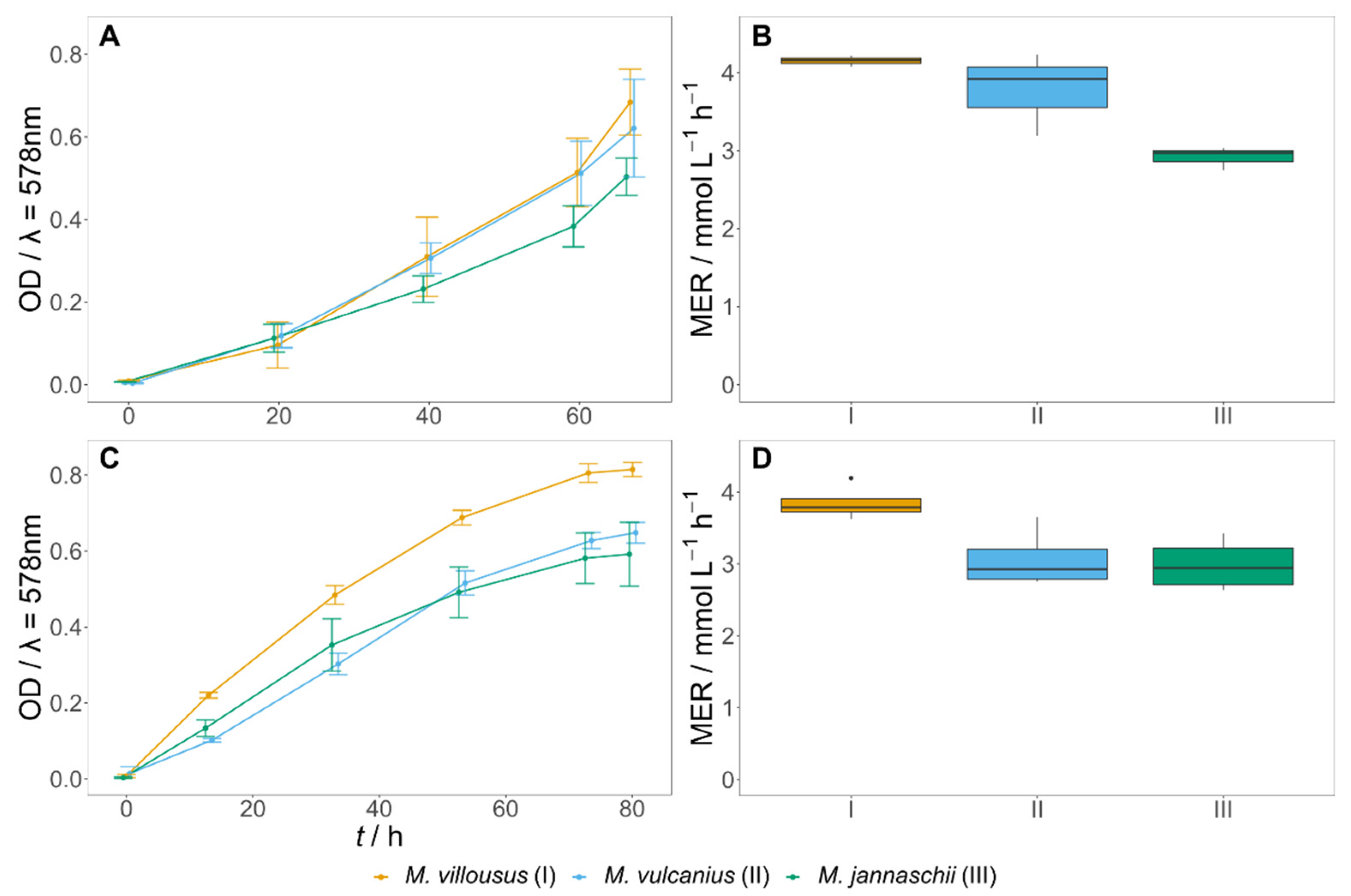
3.1. Growth Kinetics of Methanogens in Defined Medium
3.2. T. onnurineus Grown on CO
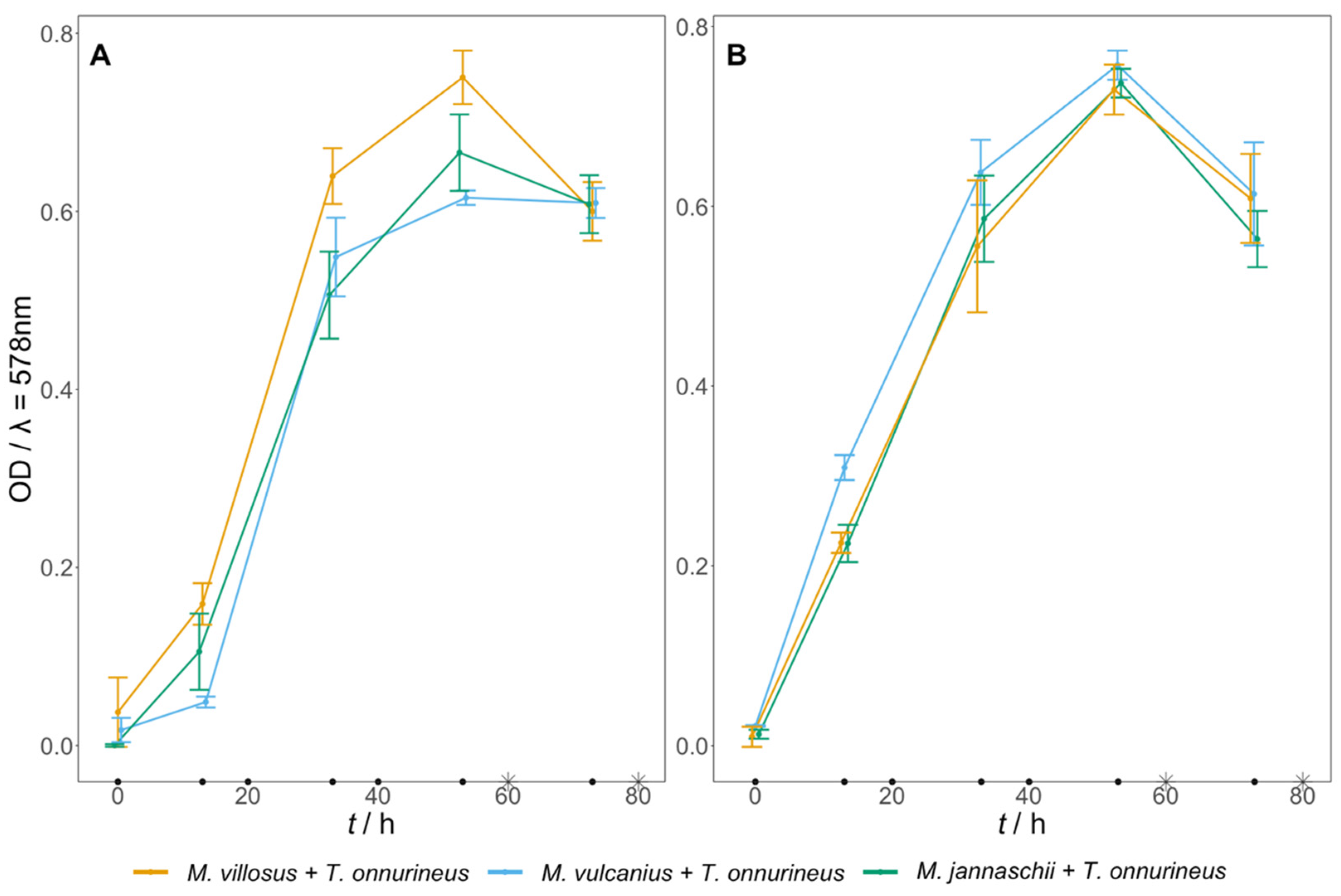
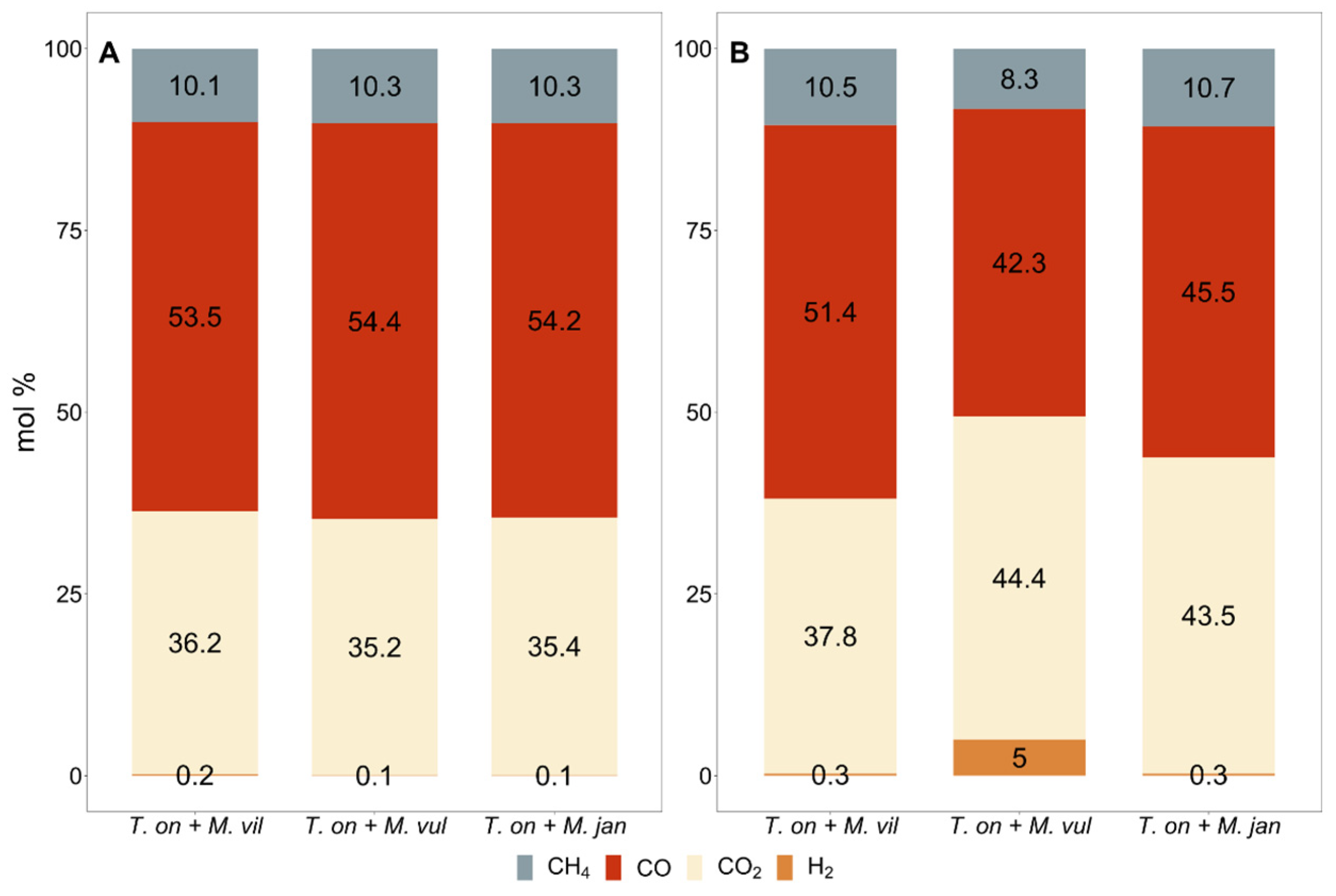
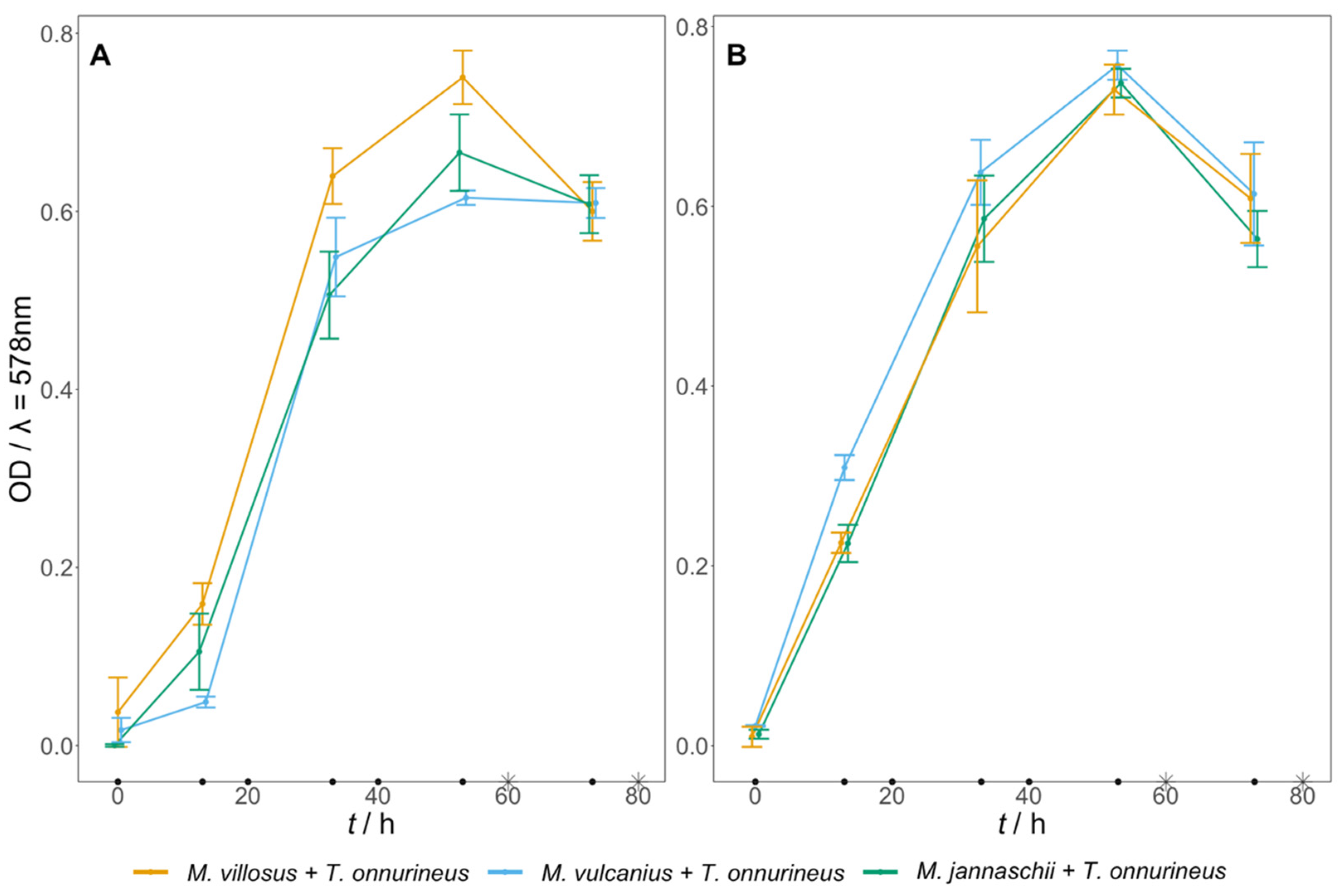
3.3. Artificial Archaeal Co-Culture Engineering
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CO2 | Carbon dioxide |
| CH4 | Methane |
| CO | Carbon monoxide |
| H2 | Molecular hydrogen |
| WGSR | Water gas shift reaction |
| DSMZ | Deutsche Sammlung von Mikroorganismen und Zellkulturen |
| OD | Optical density |
| GC | Gas chromatography |
| MER | Methane evolution rate |
| HER | Molecular hydrogen evolution rate |
| COUR | Carbon monoxide uptake rate |
| CER | Carbon dioxide evolution rate |
| HUR | Molecular hydrogen uptake rate |
| CUR | Carbon dioxide uptake rate |
References
- EEA (European Environment Agency). Primary Energy Consumption by Fuel in the EU-27; EEA: København, Denmark, 2020; Volume 28. [Google Scholar]
- EEA (European Environment Agency). National Emissions Reported to the UNFCCC and to the EU Greenhouse Gas Monitoring Mechanism; EEA: København, Denmark, 2019; Available online: https://www.eea.europa.eu/data-and-maps/data/national-emissions-reported-to-the-unfccc-and-to-the-eu-greenhouse-gas-monitoring-mechanism-14 (accessed on 10 November 2021).
- Sterner, M.; Specht, M. Power-to-Gas and Power-to-X—The History and Results of Developing a New Storage Concept. Energies 2021, 14, 6594. [Google Scholar] [CrossRef]
- Götz, M.; Lefebvre, J.; Mörs, F.; McDaniel Koch, A.; Graf, F.; Bajohr, S.; Reimert, R.; Kolb, T. Renewable Power-to-Gas: A Technological and Economic Review. Renew. Energy 2016, 85, 1371–1390. [Google Scholar] [CrossRef] [Green Version]
- Rönsch, S.; Schneider, J.; Matthischke, S.; Schlüter, M.; Götz, M.; Lefebvre, J.; Prabhakaran, P.; Bajohr, S. Review on Methanation – From Fundamentals to Current Projects. Fuel 2016, 166, 276–296. [Google Scholar] [CrossRef]
- Eyl-Mazzega, M.-A.; Mathieu, C.; Boesgaard, K.; Daniel-Gromke, J.; Denysenko, V.; Liebetrau, J.; Cornot-Gandolphe, S. Biogas and Bio-Methane in Europe: Lessons from Denmark, Germany and Italy; Études de l’Ifri; French Institute of International Relations (IFRI): Paris, France, 2019; ISBN 979-10-373-0025-6. [Google Scholar]
- Rittmann, S.K.-M.; Seifert, A.H.; Bernacchi, S. Kinetics, Multivariate Statistical Modelling, and Physiology of CO2-Based Biological Methane Production. Appl. Energy 2018, 216, 751–760. [Google Scholar] [CrossRef]
- Mauerhofer, L.-M.; Zwirtmayr, S.; Pappenreiter, P.; Bernacchi, S.; Seifert, A.H.; Reischl, B.; Schmider, T.; Taubner, R.-S.; Paulik, C.; Rittmann, S.K.-M.R. Hyperthermophilic Methanogenic Archaea Act as High-Pressure CH4 Cell Factories. Commun. Biol. 2021, 4, 289. [Google Scholar] [CrossRef]
- Mauerhofer, L.-M.; Reischl, B.; Schmider, T.; Schupp, B.; Nagy, K.; Pappenreiter, P.; Zwirtmayr, S.; Schuster, B.; Bernacchi, S.; Seifert, A.H.; et al. Physiology and Methane Productivity of Methanobacterium Thermaggregans. Appl. Microbiol. Biotechnol. 2018, 102, 7643–7656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel Azim, A.; Pruckner, C.; Kolar, P.; Taubner, R.-S.; Fino, D.; Saracco, G.; Sousa, F.L.; Rittmann, S.K.-M.R. The Physiology of Trace Elements in Biological Methane Production. Bioresour. Technol. 2017, 241, 775–786. [Google Scholar] [CrossRef]
- Seifert, A.H.; Rittmann, S.; Herwig, C. Analysis of Process Related Factors to Increase Volumetric Productivity and Quality of Biomethane with Methanothermobacter Marburgensis. Appl. Energy 2014, 132, 155–162. [Google Scholar] [CrossRef]
- Seifert, A.H.; Rittmann, S.; Bernacchi, S.; Herwig, C. Method for Assessing the Impact of Emission Gasses on Physiology and Productivity in Biological Methanogenesis. Bioresour. Technol. 2013, 136, 747–751. [Google Scholar] [CrossRef]
- Burkhardt, M.; Jordan, I.; Heinrich, S.; Behrens, J.; Ziesche, A.; Busch, G. Long Term and Demand-Oriented Biocatalytic Synthesis of Highly Concentrated Methane in a Trickle Bed Reactor. Appl. Energy 2019, 240, 818–826. [Google Scholar] [CrossRef]
- Burkhardt, M.; Koschack, T.; Busch, G. Biocatalytic Methanation of Hydrogen and Carbon Dioxide in an Anaerobic Three-Phase System. Bioresour. Technol. 2015, 178, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.B.; Strübing, D.; de Jonge, N.; Nielsen, J.L.; Ottosen, L.D.M.; Koch, K.; Kofoed, M.V.W. Stick or Leave—Pushing Methanogens to Biofilm Formation for Ex Situ Biomethanation. Bioresour. Technol. 2019, 291, 121784. [Google Scholar] [CrossRef]
- Nock, W.J.; Serna-Maza, A.; Heaven, S.; Banks, C.J. Evaluation of Microporous Hollow Fibre Membranes for Mass Transfer of H2 into Anaerobic Digesters for Biomethanization. J. Chem. Technol. Biotechnol. 2019, 94, 2693–2701. [Google Scholar] [CrossRef] [Green Version]
- Tao, B.; Alessi, A.M.; Zhang, Y.; Chong, J.P.J.; Heaven, S.; Banks, C.J. Simultaneous Biomethanisation of Endogenous and Imported CO2 in Organically Loaded Anaerobic Digesters. Appl. Energy 2019, 247, 670–681. [Google Scholar] [CrossRef]
- Rittmann, S.K.-M.; Seifert, A.; Herwig, C. Essential Prerequisites for Successful Bioprocess Development of Biological CH4 Production from CO2 and H2. Crit. Rev. Biotechnol. 2015, 35, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Rittmann, S.K.-M.R.; Seifert, A.H.; Krajete, A. Biomethanisierung—ein Prozess zur Ermöglichung der Energiewende? BIOspektrum 2014, 20, 816–817. [Google Scholar] [CrossRef]
- Rittmann, S.K.-M.R. A Critical Assessment of Microbiological Biogas to Biomethane Upgrading Systems. In Biogas Science and Technology; Guebitz, G.M., Bauer, A., Bochmann, G., Gronauer, A., Weiss, S., Eds.; Advances in Biochemical Engineering/Biotechnology; Springer International Publishing: Cham, Switzerland, 2015; pp. 117–135. ISBN 978-3-319-21993-6. [Google Scholar]
- Van der Stricht, W.; De Maré, C.; Plattner, T.; Fleischanderl, A.; Haselgribler, M.; Nair, P.; Wolf, C. Sustainable Production of Low Carbon, Renewable Fuels by Fermenting Industrial Process Gasses from the Iron and Steel Industry; ArcelorMittal, Primetals Technologies, LanzaTech: Gent, Belgium, 2017. [Google Scholar]
- Safari, F.; Tavasoli, A.; Ataei, A.; Choi, J.-K. Hydrogen and Syngas Production from Gasification of Lignocellulosic Biomass in Supercritical Water Media. Int. J. Recycl. Org. Waste Agric. 2015, 4, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Dürre, P.; Eikmanns, B.J. C1-Carbon Sources for Chemical and Fuel Production by Microbial Gas Fermentation. Curr. Opin. Biotechnol. 2015, 35, 63–72. [Google Scholar] [CrossRef]
- de Medeiros, E.M.; Noorman, H.; Maciel Filho, R.; Posada, J.A. Multi-Objective Sustainability Optimization of Biomass Residues to Ethanol via Gasification and Syngas Fermentation: Trade-Offs between Profitability, Energy Efficiency, and Carbon Emissions. Fermentation 2021, 7, 201. [Google Scholar] [CrossRef]
- Phillips, J.R.; Huhnke, R.L.; Atiyeh, H.K. Syngas Fermentation: A Microbial Conversion Process of Gaseous Substrates to Various Products. Fermentation 2017, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Westman, S.Y.; Chandolias, K.; Taherzadeh, M.J. Syngas Biomethanation in a Semi-Continuous Reverse Membrane Bioreactor (RMBR). Fermentation 2016, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Devarapalli, M.; Lewis, R.S.; Atiyeh, H.K. Continuous Ethanol Production from Synthesis Gas by Clostridium Ragsdalei in a Trickle-Bed Reactor. Fermentation 2017, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Rittmann, S.K.-M.; Lee, H.S.; Lim, J.K.; Kim, T.W.; Lee, J.-H.; Kang, S.G. One-Carbon Substrate-Based Biohydrogen Production: Microbes, Mechanism, and Productivity. Biotechnol. Adv. 2015, 33, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Thauer, R.K. Citric-Acid Cycle, 50 Years on: Modifications and an Alternative Pathway in Anaerobic Bacteria. Eur. J. Biochem. 1988, 176, 497–508. [Google Scholar] [CrossRef]
- Klasson, K.; Cowger, J.; Ko, C.; Vega, J.; Clausen, E.; Gaddy, J. Methane Production from Synthesis Gas Using a Mixed Culture OfR. Rubrum M. Barkeri, and M. Formicicum. Appl. Biochem. Biotechnol. 1990, 24, 317–328. [Google Scholar] [CrossRef]
- Daniels, L.; Fuchs, G.; Thauer, R.; Zeikus, J. Carbon Monoxide Oxidation by Methanogenic Bacteria. J. Bacteriol. 1977, 132, 118–126. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.M.; Wolkin, R.; Moench, T.; Morgan, J.; Zeikus, J. Association of Hydrogen Metabolism with Unitrophic or Mixotrophic Growth of Methanosarcina Barkeri on Carbon Monoxide. J. Bacteriol. 1984, 158, 373–375. [Google Scholar] [CrossRef] [Green Version]
- Diender, M.; Pereira, R.; Wessels, H.J.; Stams, A.J.; Sousa, D.Z. Proteomic Analysis of the Hydrogen and Carbon Monoxide Metabolism of Methanothermobacter Marburgensis. Front. Microbiol. 2016, 7, 1049. [Google Scholar] [CrossRef] [Green Version]
- Diender, M.; Uhl, P.S.; Bitter, J.H.; Stams, A.J.; Sousa, D.Z. High Rate Biomethanation of Carbon Monoxide-Rich Gases via a Thermophilic Synthetic Coculture. ACS Sustain. Chem. Eng. 2018, 6, 2169–2176. [Google Scholar] [CrossRef] [Green Version]
- Kohlmayer, M.; Robert, H.; Raimund, B.; Wolfgang, M. Simultaneous CO2 and CO methanation using microbes. Microbiology 2018. [Google Scholar] [CrossRef] [Green Version]
- Schmider, T. Ecophysiology and Methane Productivity of Carboxydotrophy-Based Archael Co-Cultures. Master’s Thesis, University of Vienna, Vienna, Austria, 2018. [Google Scholar]
- Kim, M.-S.; Fitriana, H.N.; Kim, T.W.; Kang, S.G.; Jeon, S.G.; Chung, S.H.; Park, G.W.; Na, J.-G. Enhancement of the Hydrogen Productivity in Microbial Water Gas Shift Reaction by Thermococcus Onnurineus NA1 Using a Pressurized Bioreactor. Int. J. Hydrog. Energy 2017, 42, 27593–27599. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, M.-S.; Lee, J.-H.; Kim, T.W.; Bae, S.S.; Lee, S.-M.; Jung, H.C.; Yang, T.-J.; Choi, A.R.; Cho, Y.-J.; et al. Adaptive Engineering of a Hyperthermophilic Archaeon on CO and Discovering the Underlying Mechanism by Multi-Omics Analysis. Sci. Rep. 2016, 6, 22896. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-S.; Bae, S.S.; Kim, Y.J.; Kim, T.W.; Lim, J.K.; Lee, S.H.; Choi, A.R.; Jeon, J.H.; Lee, J.-H.; Lee, H.S.; et al. CO-Dependent H2 Production by Genetically Engineered Thermococcus Onnurineus NA1. Appl. Environ. Microbiol. 2013, 79, 2048–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, S.S.; Kim, T.W.; Lee, H.S.; Kwon, K.K.; Kim, Y.J.; Kim, M.-S.; Lee, J.-H.; Kang, S.G. H2 Production from CO, Formate or Starch Using the Hyperthermophilic Archaeon, Thermococcusonnurineus. Biotechnol. Lett. 2012, 34, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.-S.; Kim, Y.-J.; Yang, S.-H.; Lim, J.-K.; Jeon, J.-H.; Lee, H.-S.; Kang, S.-G.; Kim, S.; Lee, J. Thermococcus onnurineus Sp. Nov., a Hyperthermophilic Archaeon Isolated from a Deep-Sea Hydrothermal Vent Area at the PACMANUS Field. J. Microbiol. Biotechnol. 2006, 16, 1826. [Google Scholar]
- Bellack, A.; Huber, H.; Rachel, R.; Wanner, G.; Wirth, R. Methanocaldococcus villosus Sp. Nov., a Heavily Flagellated Archaeon That Adheres to Surfaces and Forms Cell–Cell Contacts. Int. J. Syst. Evol. Microbiol. 2011, 61, 1239–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeanthon, C.; l’Haridon, S.; Reysenbach, A.-L.; Corre, E.; Vernet, M.; Messner, P.; Sleytr, U.; Prieur, D. Methanococcus vulcanius Sp. Nov., a Novel Hyperthermophilic Methanogen Isolated from East Pacific Rise, and Identification of Methanococcus Sp. DSM 4213Tas Methanococcus fervens Sp. Nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 583–589. [Google Scholar] [CrossRef]
- Jones, W.; Leigh, J.; Mayer, F.; Woese, C.; Wolfe, R. Methanococcus jannaschii Sp. Nov., an Extremely Thermophilic Methanogen from a Submarine Hydrothermal Vent. Arch. Microbiol. 1983, 136, 254–261. [Google Scholar] [CrossRef]
- Diender, M.; Stams, A.J.; Sousa, D.Z. Pathways and Bioenergetics of Anaerobic Carbon Monoxide Fermentation. Front. Microbiol. 2015, 6, 1275. [Google Scholar] [CrossRef] [Green Version]
- Balch, W.E.; Wolfe, R. New Approach to the Cultivation of Methanogenic Bacteria: 2-Mercaptoethanesulfonic Acid (HS-CoM)-Dependent Growth of Methanobacterium Ruminantium in a Pressureized Atmosphere. Appl. Environ. Microbiol. 1976, 32, 781–791. [Google Scholar] [CrossRef] [Green Version]
- Taubner, R.-S.; Rittmann, S.K.-M. Method for Indirect Quantification of CH4 Production via H2O Production Using Hydrogenotrophic Methanogens. Front. Microbiol. 2016, 7, 532. [Google Scholar] [CrossRef] [Green Version]
- Rittmann, S.K.-M.; Herwig, C. A Comprehensive and Quantitative Review of Dark Fermentative Biohydrogen Production. Microb. Cell Factories 2012, 11, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappenreiter, P.A.; Zwirtmayr, S.; Mauerhofer, L.-M.; Rittmann, S.K.-M.R.; Paulik, C. Development of a Simultaneous Bioreactor System for Characterization of Gas Production Kinetics of Methanogenic Archaea at High Pressure. Eng. Life Sci. 2019, 19, 537–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittmann, S.K.-M.; Seifert, A.; Herwig, C. Quantitative Analysis of Media Dilution Rate Effects on Methanothermobacter Marburgensis Grown in Continuous Culture on H2 and CO2. Biomass Bioenergy 2012, 36, 293–301. [Google Scholar] [CrossRef]
- Bernacchi, S.; Rittmann, S.K.-M.; Seifert, A.H.; Krajete, A.; Herwig, C. Experimental Methods for Screening Parameters Influencing the Growth to Product Yield (Y (x/CH4)) of a Biological Methane Production (BMP) Process Performed with Methanothermobacter Marburgensis. AIMS Bioeng. 2014, 1, 72–86. [Google Scholar] [CrossRef]
- Reischl, B.; Ergal, İ.; Rittmann, S.K.-M. Biohydrogen Production Characteristics of Desulfurococcus Amylolyticus DSM 16532. Int. J. Hydrog. Energy 2018, 43, 8747–8753. [Google Scholar] [CrossRef]
- Pruckner, C. Evolutionary and Physiological Aspects of Potentially Carboxydotrophic Archaea. Master’s Thesis, University of Vienna, Vienna, Austria, 2019. [Google Scholar]
- Pfeifer, K.; Ergal, İ.; Koller, M.; Basen, M.; Schuster, B.; Rittmann, S.K.-M.R. Archaea Biotechnology. Biotechnol. Adv. 2021, 47, 107668. [Google Scholar] [CrossRef]
- Ergal, İ.; Bochmann, G.; Fuchs, W.; Rittmann, S.K.-M.R. Design and Engineering of Artificial Microbial Consortia for Biohydrogen Production. Curr. Opin. Biotechnol. 2022, 73, 74–80. [Google Scholar] [CrossRef]
- Guiot, S.R.; Cimpoia, R.; Carayon, G. Potential of Wastewater-Treating Anaerobic Granules for Biomethanation of Synthesis Gas. Environ. Sci. Technol. 2011, 45, 2006–2012. [Google Scholar] [CrossRef] [Green Version]
- Straub, C.T.; Counts, J.A.; Nguyen, D.M.N.; Wu, C.-H.; Zeldes, B.M.; Crosby, J.R.; Conway, J.M.; Otten, J.K.; Lipscomb, G.L.; Schut, G.J.; et al. Biotechnology of Extremely Thermophilic Archaea. FEMS Microbiol. Rev. 2018, 42, 543–578. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
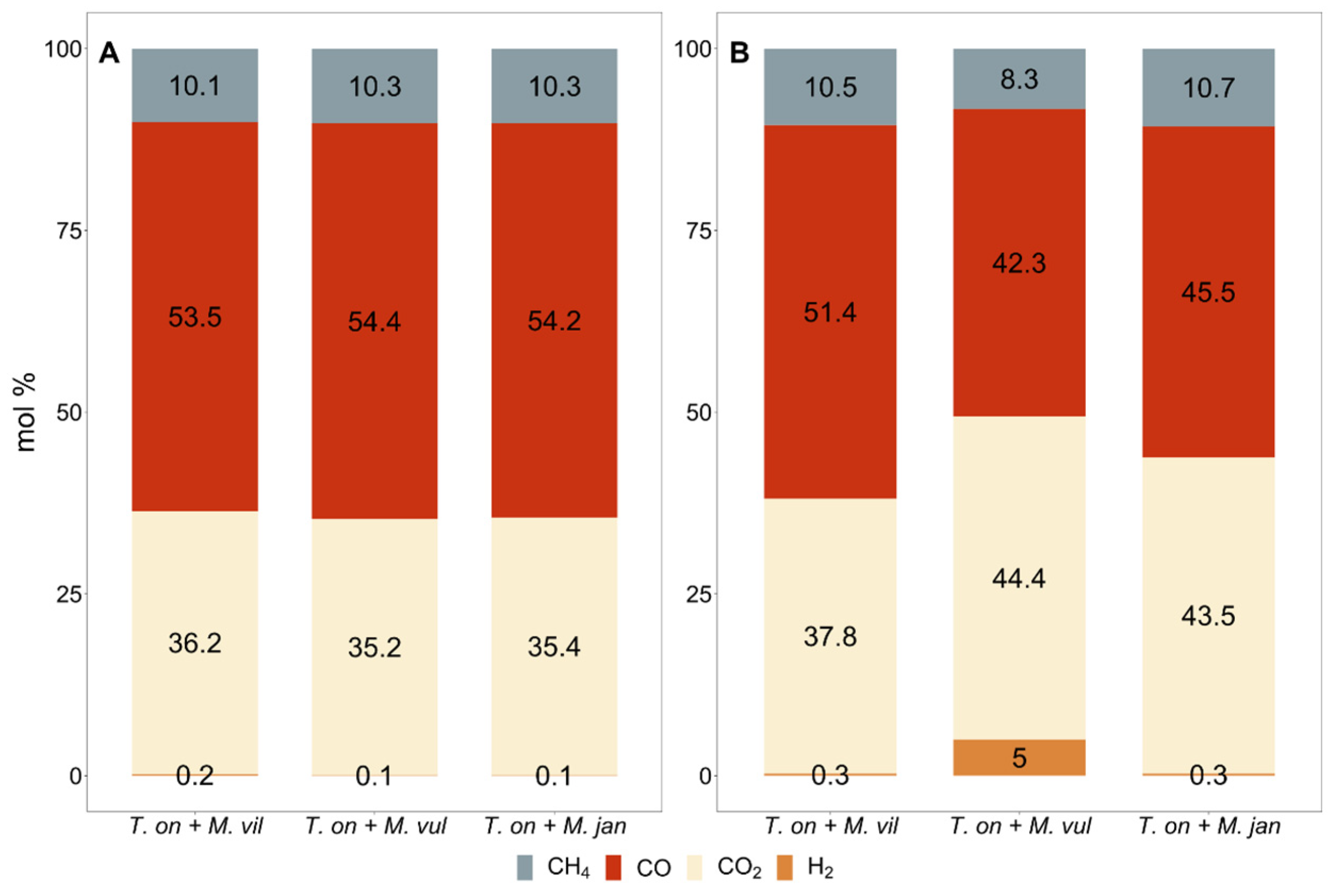{kind=link}
| Co-Culture | MER/mmol L−1 h−1 | HUR/mmol L−1 h−1 | CUR/mmol L−1 h−1 | COUR/mmol L−1 h−1 | CER/mmol L−1 h−1 |
|---|---|---|---|---|---|
| M. villosus + T. onnurineus | 1.4 ± 0.4 | 6.4 ± 1.5 | 1.4 ± 0.4 | 6.4 ± 1.5 | 5.0 ± 1.1 |
| M. vulcanius + T. onnurineus | 1.6 ± 0.3 | 7.0 ± 1.3 | 1.6 ± 0.3 | 7.0 ± 1.3 | 5.4 ± 0.9 |
| M. jannaschii + T. onnurineus | 1.5 ± 0.4 | 6.5 ± 1.5 | 1.5 ± 0.3 | 6.5 ± 1.5 | 5.0 ±1.2 |
| Co-Culture | MER/mmol L−1 h−1 | HUR/mmol L−1 h−1 | CUR/mmol L−1 h−1 | COUR/mmol L−1 h−1 | CER/mmol L−1 h−1 |
|---|---|---|---|---|---|
| M. villosus + T. onnurineus | 2.0 ± 0.3 | 8.3 ± 1.3 | 2.1 ± 0.3 | 8.4 ± 1.3 | 6.3 ± 0.9 |
| M. vulcanius + T. onnurineus | 1.6 ± 0.5 | 10.3 ± 0.2 | 2.6 ± 0.3 | 11.3 ± 0.7 | 8.7 ± 0.5 |
| M. jannaschii + T. onnurineus | 1.8 ± 0.4 | 9.0 ± 1.5 | 1.9 ± 0.5 | 9.1 ± 1.6 | 7.2 ± 1.2 |
| Co-Culture | Gas | MER/mmol L−1 h−1 | CER/mmol L−1 h−1 |
|---|---|---|---|
| M. villosus + T. onnurineus | CO | 2.0 ± 0.3 | 6.3 ± 0.9 |
| T. onnurineus + M. villosus | CO | 1.1 ± 0.1 | 3.7 ± 1.0 |
| M. villosus + T. onnurineus | H2:CO | 1.0 ± 0.4 | 0.0 ± 0.0 |
| T. onnurineus + M. villosus | H2:CO | 1.0 ± 0.3 | 0.0 ± 0.0 |
| M. villosus + T. onnurineus | Art. syngas | 1.5 ± 0.5 | 0.9 ± 0.6 |
| T. onnurineus + M. villosus | Art. syngas | 0.9 ± 0.3 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zipperle, A.; Reischl, B.; Schmider, T.; Stadlbauer, M.; Kushkevych, I.; Pruckner, C.; Vítězová, M.; Rittmann, S.K.-M.R. Biomethanation of Carbon Monoxide by Hyperthermophilic Artificial Archaeal Co-Cultures. Fermentation 2021, 7, 276. https://doi.org/10.3390/fermentation7040276
Zipperle A, Reischl B, Schmider T, Stadlbauer M, Kushkevych I, Pruckner C, Vítězová M, Rittmann SK-MR. Biomethanation of Carbon Monoxide by Hyperthermophilic Artificial Archaeal Co-Cultures. Fermentation. 2021; 7(4):276. https://doi.org/10.3390/fermentation7040276
Chicago/Turabian StyleZipperle, Aaron, Barbara Reischl, Tilman Schmider, Michael Stadlbauer, Ivan Kushkevych, Christian Pruckner, Monika Vítězová, and Simon K.-M. R. Rittmann. 2021. "Biomethanation of Carbon Monoxide by Hyperthermophilic Artificial Archaeal Co-Cultures" Fermentation 7, no. 4: 276. https://doi.org/10.3390/fermentation7040276
APA StyleZipperle, A., Reischl, B., Schmider, T., Stadlbauer, M., Kushkevych, I., Pruckner, C., Vítězová, M., & Rittmann, S. K.-M. R. (2021). Biomethanation of Carbon Monoxide by Hyperthermophilic Artificial Archaeal Co-Cultures. Fermentation, 7(4), 276. https://doi.org/10.3390/fermentation7040276