
Citric Acid Influences the Dynamics of the Fermentation Quality, Protease Activity and Microbial Community of Mulberry Leaf Silage



Abstract
1. Introduction
2. Materials and Methods
2.1. Pre-Ensiled Material and Silage Preparation
2.2. Microorganism and Fermentation Parameter Analyses
2.3. Protease Activity Analysis
2.4. Analysis of Microbial Communities
2.5. Statistical Analysis
3. Results and Discussion
3.1. Characteristics of Pre-Ensiled ML
3.2. The Fermentation Parameters, Organic Acids and Microbial Population of ML Silages
3.3. The Protein Fraction of ML Silages
3.4. The Protease Activities of ML Silage
3.5. Bacterial Community of ML Silage
3.5.1. The Dynamic Variation and Alpha Diversity of the Bacterial Community
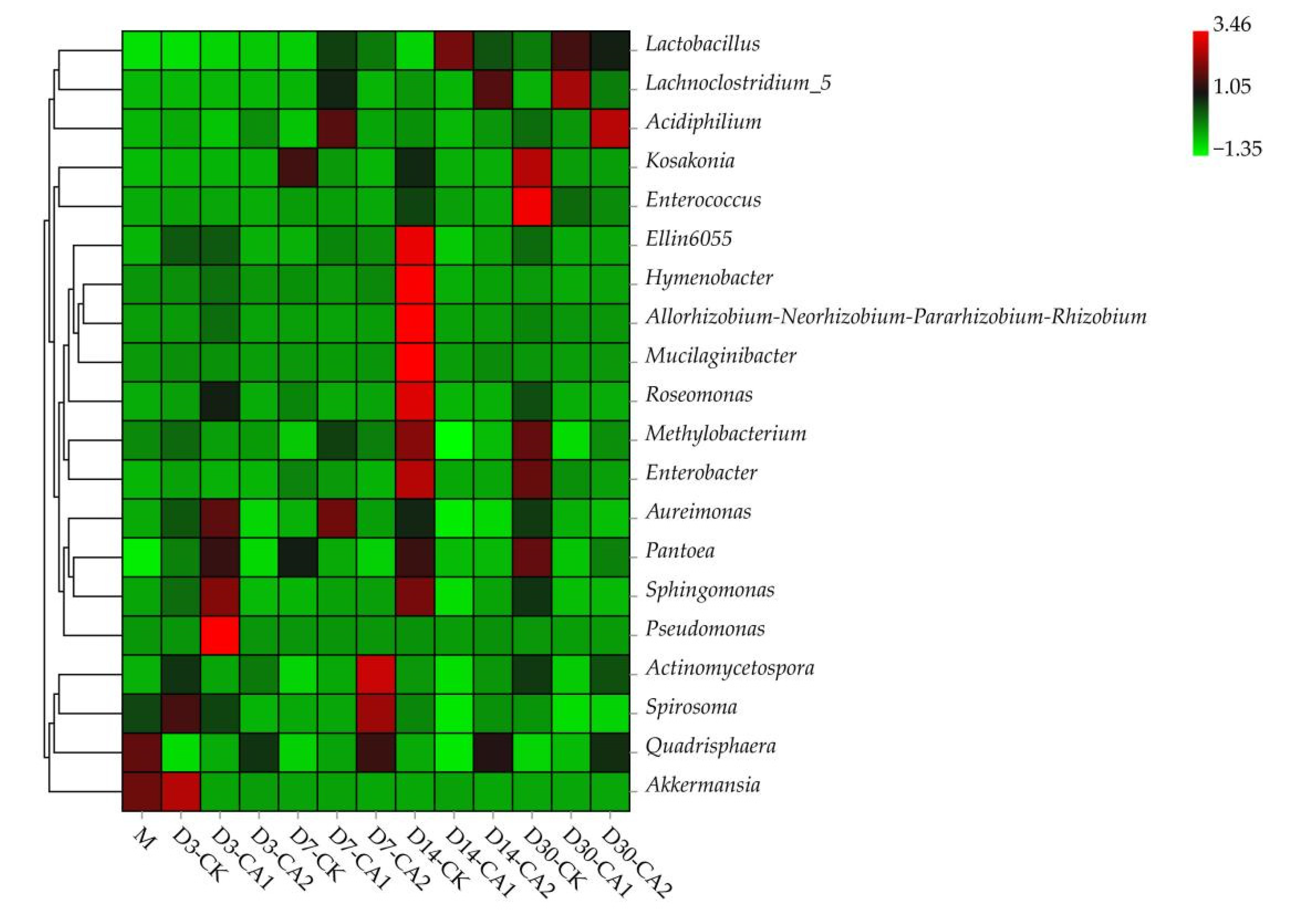
3.5.2. The Changes in the Dynamics of Relative Abundance among Bacterial Communities
3.5.3. 16S rDNA Gene-Predicted Functional Profiles
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brief Introduction of Cocoon Silk Industry Operation in China. Available online: http://www.mofcom.gov.cn/article/tongjiziliao/sjtj/jsc/201904/20190402852178.shtml (accessed on 12 April 2019).
- Wang, Y.; Chen, X.; Wang, C.; He, L.; Zhou, W.; Yang, F.; Qing, Z. The bacterial community and fermentation quality of mulberry (Morus alba) leaf silage with or without Lactobacillus casei and sucrose. Bioresour. Technol. 2019, 293, 122059. [Google Scholar] [CrossRef]
- Singh, B.; Makkar, H. The potential of mulberry foliage as feed supplement in India. Fao Anim. Prod. Health Pap. 2002, 1, 139–156. [Google Scholar]
- Ouyang, J.; Wang, M.; Hou, Q.; Feng, D.; Pi, Y.; Zhao, W. Effects of Dietary Mulberry Leaf Powder in Concentrate on the Rumen Fermentation and Ruminal Epithelium in Fattening Hu Sheep. Animals 2019, 9, 218. [Google Scholar] [CrossRef]
- Król, E.; Jeszka-Skowron, M.; Krejpcio, Z.; Flaczyk, E.; Wójciak, R.W. The Effects of Supplementary Mulberry Leaf (Morus alba) Extracts on the Trace Element Status (Fe, Zn and Cu) in Relation to Diabetes Management and Antioxidant Indices in Diabetic Rats. Biol. Trace Element Res. 2016, 174, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Lee, S.-M.; Lee, J.E.; Kim, J.-H. Anti-inflammatory activity of mulberry leaf extract through inhibition of NF-κB. J. Funct. Foods 2013, 5, 178–186. [Google Scholar] [CrossRef]
- He, L.; Chen, N.; Lv, H.; Wang, C.; Zhou, W.; Chen, X.; Zhang, Q. Gallic acid influencing fermentation quality, nitrogen distribution and bacterial community of high-moisture mulberry leaves and stylo silage. Bioresour. Technol. 2020, 295, 122255. [Google Scholar] [CrossRef]
- He, L.; Wang, C.; Xing, Y.; Zhou, W.; Pian, R.; Yang, F.; Chen, X.; Zhang, Q. Dynamics of proteolysis, protease activity and bacterial community of Neolamarckia cadamba leaves silage and the effects of formic acid and Lactobacillus farciminis. Bioresour. Technol. 2019, 294, 122127. [Google Scholar] [CrossRef]
- Nozawa, S.R.; Rigoli, I.C.; Thedei, G., Jr.; Rossi, A. Mind the buffering capacity of citric acid. Fungal Genet. Rep. 1995, 42, 56. [Google Scholar] [CrossRef]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef]
- Soccol, C.R.; Luciana, P.S.V.; Rodrigues, C.; Pandey, A. New perspectives for citric acid production and application. Food Technol. Biotech. 2006, 44, 141–149. [Google Scholar] [CrossRef]
- Muck, R. Factors Influencing Silage Quality and Their Implications for Management. J. Dairy Sci. 1988, 71, 2992–3002. [Google Scholar] [CrossRef]
- Lv, H.; Pian, R.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Effects of citric acid on fermentation characteristics and bacterial diversity of Amomum villosum silage. Bioresour. Technol. 2020, 307, 123290. [Google Scholar] [CrossRef]
- Ke, W.; Ding, W.; Xu, D.; Ding, L.; Zhang, P.; Li, F.; Guo, X. Effects of addition of malic or citric acids on fermentation quality and chemical characteristics of alfalfa silage. J. Dairy Sci. 2017, 100, 8958–8966. [Google Scholar] [CrossRef]
- Yan, Y.; Li, X.; Guan, H.; Huang, L.; Ma, X.; Peng, Y.; Li, Z.; Nie, G.; Zhou, J.; Yang, W.; et al. Microbial community and fermentation characteristic of Italian ryegrass silage prepared with corn stover and lactic acid bacteria. Bioresour. Technol. 2019, 279, 166–173. [Google Scholar] [CrossRef]
- Broderick, G.; Kang, J. Automated Simultaneous Determination of Ammonia and Total Amino Acids in Ruminal Fluid and In Vitro Media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, Z.; Wang, X.; Tian, J. Effects of inoculants and environmental temperature on fermentation quality and bacterial diversity of alfalfa silage. Anim. Sci. J. 2018, 89, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhou, H.; Yu, Z.; Zhang, Y. Changes in the distribution of nitrogen and plant enzymatic activity during ensilage of lucerne treated with different additives. Grass Forage Sci. 2007, 62, 35–43. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Li, D.X.; Wang, X.K.; Lin, Y.L.; Zhang, Q.; Chen, X.Y.; Yang, F.Y. Fermentation dynamics and diversity of bacterial community in four typical woody forages. Ann. Microbiol. 2019, 69, 233–240. [Google Scholar] [CrossRef]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef]
- He, L.; Lv, H.; Chen, N.; Wang, C.; Zhou, W.; Chen, X.; Zhang, Q. Improving fermentation, protein preservation and antioxidant activity of Moringa oleifera leaves silage with gallic acid and tannin acid. Bioresour. Technol. 2020, 297, 122390. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.J.; Moser, L.E. Plant Factors Affecting Forage Quality. In Forage Quality, Evaluation, and Utilization; Acsess Publication: Madison, WI, USA, 1994; pp. 115–154. [Google Scholar] [CrossRef]
- Cai, Y.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of Lactobacillus spp. from an Inoculant and of Weissella and Leuconostoc spp. from Forage Crops on Silage Fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [PubMed]
- Borreani, G.; Tabacco, E.; Schmidt, R.; Holmes, B.; Muck, R. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [PubMed]
- Trabi, E.B.; Yuan, X.; Li, J.; Dong, Z.; Shah, A.A.; Shao, T. Effect of Glucose and Lactic Acid Bacteria on the Fermentation Quality, Chemical Compositions and in vitro Digestibility of Mulberry (Morus Alba) Leaf Silage. Pak. J. Zool. 2017, 49. [Google Scholar] [CrossRef]
- Ranjit, N.; Kung, L. The Effect of Lactobacillus buchneri, Lactobacillus plantarum, or a Chemical Preservative on the Fermentation and Aerobic Stability of Corn Silage. J. Dairy Sci. 2000, 83, 526–535. [Google Scholar] [CrossRef]
- Muck, R.; Nadeau, E.; McAllister, T.A.; Contreras-Govea, F.; Santos, M.; Kung, L. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Mohan, A.; Pohlman, F. Role of organic acids and peroxyacetic acid as antimicrobial intervention for controlling Escherichia coli O157:H7 on beef trimmings. LWT Food Sci. Technol. 2016, 65, 868–873. [Google Scholar] [CrossRef]
- Omidiran, M.O.; Baiyewu, R.A.; Ademola, I.T.; Fakorede, O.C.; Toyinbo, E.O.; Adekunle, E.A. Phytochemical Analysis, Nutritional Composition and Antimicrobial Activities of White Mulberry (Morus alba). Pak. J. Nutr. 2012, 11, 456–460. [Google Scholar] [CrossRef][Green Version]
- Mcdonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage; Chalcombe Publication: Marlow Bucks, UK, 1991. [Google Scholar]
- Ke, W.; Ding, W.; Xu, D.; Shah, M.N.; Zhang, P.; Guo, X. Influences of addition of malic acid or citric acid, Lactobacillus plantarum and their mixtures on fermentation quality, proteolysis and fatty acid composition of ensiled alfalfa. Arch. Anim. Nutr. 2018, 72, 492–502. [Google Scholar] [CrossRef]
- Yuan, X.; Wen, A.; Desta, S.T.; Dong, Z.; Shao, T. Effects of four short-chain fatty acids or salts on the dynamics of nitrogen transformations and intrinsic protease activity of alfalfa silage. J. Sci. Food Agric. 2017, 97, 2759–2766. [Google Scholar] [CrossRef]
- Helander, I.M.; Mattila-Sandholm, T. Fluorometric assessment of Gram-negative bacterial permeabilization. J. Appl. Microbiol. 2000, 88, 213–219. [Google Scholar] [CrossRef]
- Ni, K.; Minh, T.T.T.; Tu, T.T.M.; Tsuruta, T.; Pang, H.; Nishino, N. Comparative microbiota assessment of wilted Italian ryegrass, whole crop corn, and wilted alfalfa silage using denaturing gradient gel electrophoresis and next-generation sequencing. Appl. Microbiol. Biotechnol. 2017, 101, 1385–1394. [Google Scholar] [CrossRef]
- Yang, L.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 2018, 275, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, O.; Ogunade, I.; Weinberg, Z.; Adesogan, A. Silage review: Foodborne pathogens in silage and their mitigation by silage additives. J. Dairy Sci. 2018, 101, 4132–4142. [Google Scholar] [CrossRef] [PubMed]
- Knief, C.; Delmotte, N.; Chaffron, S.; Stark, M.; Innerebner, G.; Wassmann, R.; von Mering, C.; A Vorholt, J. Metaproteogenomic analysis of microbial communities in the phyllosphere and rhizosphere of rice. ISME J. 2011, 6, 1378–1390. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yao, D.; Li, D.; Lin, Y.; Bureenok, S.; Ni, K.; Yang, F. Effects of Lactic Acid Bacteria Isolated from Rumen Fluid and Feces of Dairy Cows on Fermentation Quality, Microbial Community, and in vitro Digestibility of Alfalfa Silage. Front. Microbiol. 2020, 10, 2998. [Google Scholar] [CrossRef]
- Ogunade, I.; Jiang, Y.; Cervantes, A.P.; Kim, D.; de Oliveira, A.S.; Vyas, D.; Weinberg, Z.; Jeong, K.; Adesogan, A. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157:H7 and silage additives. J. Dairy Sci. 2018, 101, 2048–2059. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, C.M.; Camejo, P.; Oshlag, J.Z.; Noguera, D.R. Ammonia-oxidizing microbial communities in reactors with efficient nitrification at low-dissolved oxygen. Water Res. 2015, 70, 38–51. [Google Scholar] [CrossRef]
- Li, L.; Yuan, Z.; Sun, Y.; Kong, X.; Dong, P.; Zhang, J. A reused method for molasses-processed wastewater: Effect on silage quality and anaerobic digestion performance of Pennisetum purpereum. Bioresour. Technol. 2017, 241, 1003–1011. [Google Scholar] [CrossRef]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef]
- He, L.; Wang, C.; Xing, Y.; Zhou, W.; Pian, R.; Chen, X.; Zhang, Q. Ensiling characteristics, proteolysis and bacterial community of high-moisture corn stalk and stylo silage prepared with Bauhinia variegate flower. Bioresour. Technol. 2020, 296, 122336. [Google Scholar] [CrossRef]
- Han, X.Y.; Pham, A.S.; Tarrand, J.J.; Rolston, K.V.; Helsel, L.O.; Levett, P.N. Bacteriologic Characterization of 36 Strains of Roseomonas Species and Proposal of Roseomonas mucosasp nov and Roseomonas gilardii subsp rosea subsp nov. Am. J. Clin. Pathol. 2003, 120, 256–264. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Mulberry Leaves ± SEM |
|---|---|
| Dry matter (g/kg FM) | 374 ± 12.8 |
| Crude protein (g/kg DM) | 139 ± 1.9 |
| Neutral detergent fiber (g/kg DM) | 440 ± 35.7 |
| Acid detergent fiber (g/kg DM) | 353 ± 24.8 |
| Water soluble carbohydrates (g/kg DM) | 114 ± 3.2 |
| Lactic acid bacteria (log10 cfu/g FM) | 5.72 ± 0.1 |
| Coliform bacteria (log10 cfu/g FM) | 5.21 ± 0.3 |
| Yeasts (log10 cfu/g FM) | <2.00 |
| Molds (log10 cfu/g FM) | <2.00 |
| Item | Treatments | Ensiling Days | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| 3 | 7 | 14 | 30 | D | T | D × T | |||
| Crude protein (g/kg DM) | CK | 134 A,b | 147 A,a | 154 A,a | 153 A,a | 8.57 | 0.000 | 0.000 | 0.011 |
| CA1 | 136 A,a | 138 A,B,a | 137 B,a | 146 A,B,a | |||||
| CA2 | 133 A,b,c | 130 B,c | 143 B,a | 139 B,a,b | |||||
| True protein (g/kg TN) | CK | 878 A,a | 792 A,b | 679 B,c | 620 B,d | 80.1 | 0.000 | 0.679 | 0.000 |
| CA1 | 816 B,a | 748 B,b | 752 A,b | 625 A,B,c | |||||
| CA2 | 808 B,a | 764 A,B,b | 678 B,c | 690 A,c | |||||
| Non-protein nitrogen (g/kg TN) | CK | 122 B,d | 208 B,c | 321 A,b | 380 A,a | 80.1 | 0.000 | 0.679 | 0.000 |
| CA1 | 184 A,c | 252 A,b | 248 B,b | 375 A,B,a | |||||
| CA2 | 196 A,c | 236 A,b | 322 A,a | 310 B,a | |||||
| Ammonia-N (g/kg TN) | CK | 4.40 A | 7.38 A | 14.6 A | 18.9 A | 5.15 | 0.000 | 0.000 | 0.000 |
| CA1 | 3.71 B | 4.57 B | 7.58 B | 10.8 B | |||||
| CA2 | 1.76 B | 2.45 C | 3.95 C | 6.07 C | |||||
| Item | Treatments | Ensiling Days | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| 3 | 7 | 14 | 30 | D | T | D×T | |||
| Carboxypeptidase (units/h/DM) | CK | 70.9 A,b | 66.9 A,b | 80.8 A,a | 83.1 A,a | 7.33 | 0.003 | 0.000 | 0.021 |
| CA1 | 69.2 A,a | 74.8 B,a | 76.5 A,a | 80.8 A,a | |||||
| CA2 | 66.9 A,a | 66.9 A,a | 66.2 B,a | 65.9 B,a | |||||
| Aminopeptidase (units/h/DM) | CK | 86.5 A,a | 83.4 A,a | 23.9 A,b | 17.4 A,b | 28.2 | 0.000 | 0.000 | 0.000 |
| CA1 | 59.6 B,a | 65.0 A,a | 18.8 A,B,b | 14.7 A,b | |||||
| CA2 | 23.6 C,a | 17.7 B,a,b | 14.75 C,b | 13.5 A,b | |||||
| Acid protease (units/h/DM) | CK | 51.3 A,a | 47.1 B,a | 50.5 A,a | 36.6 A,b | 6.76 | 0.000 | 0.018 | 0.360 |
| CA1 | 55.2 A,a | 53.9 A,a | 53.5 A,a | 41.3 A,b | |||||
| CA2 | 49.7 A,a,b | 55.2 A,a | 54.4 A,a,b | 45.8 A,b | |||||
| Item | True Protein | Non-Protein Nitrogen | Ammonia-N | Carboxypeptidase | Aminopeptidase | Acid Protease |
|---|---|---|---|---|---|---|
| True protein | 1 | |||||
| Non-protein nitrogen | −1.00 | 1 | ||||
| 0.00 | ||||||
| Ammonia-N | −0.65 | 0.65 | 1 | |||
| 0.06 | 0.06 | |||||
| Carboxypeptidase | −0.81 | 0.81 | 0.80 | 1 | ||
| <0.01 | <0.01 | <0.01 | ||||
| Aminopeptidase | −0.51 | 0.51 | 0.56 | 0.62 | 1 | |
| 0.16 | 0.16 | 0.11 | 0.08 | |||
| Acid protease | 0.56 | −0.56 | −0.74 | −0.53 | 0.06 | 1 |
| 0.11 | 0.11 | 0.02 | 0.14 | 0.87 |
| Items | Treatments | Ensiling Days | |||
|---|---|---|---|---|---|
| 3 | 7 | 14 | 30 | ||
| Sobs | CK | 1605 | 1339 | 1384 | 1373 |
| CA1 | 1477 | 1402 | 1102 | 1305 | |
| CA2 | 1376 | 1499 | 1301 | 1337 | |
| Ace | CK | 2512 | 2242 | 2263 | 2151 |
| CA1 | 2384 | 2348 | 1951 | 2097 | |
| CA2 | 2159 | 2417 | 2152 | 2241 | |
| Chao1 | CK | 2483 | 2268 | 2204 | 2115 |
| CA1 | 2308 | 2294 | 1964 | 2082 | |
| CA2 | 2095 | 2453 | 2160 | 2168 | |
| Shannon | CK | 3.00 | 3.18 | 5.02 | 5.22 |
| CA1 | 2.97 | 4.32 | 4.36 | 4.62 | |
| CA2 | 2.62 | 3.59 | 3.68 | 4.13 | |
| Simpson | CK | 0.50 | 0.56 | 0.89 | 0.93 |
| CA1 | 0.51 | 0.79 | 0.86 | 0.86 | |
| CA2 | 0.45 | 0.66 | 0.71 | 0.76 | |
| Coverage | CK | 0.99 | 0.99 | 0.99 | 0.99 |
| CA1 | 0.99 | 0.99 | 0.99 | 0.99 | |
| CA2 | 0.99 | 0.99 | 0.99 | 0.99 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, D.; Zhou, W.; Guo, X.; Zheng, M.; Chen, X.; Zhang, Q. Citric Acid Influences the Dynamics of the Fermentation Quality, Protease Activity and Microbial Community of Mulberry Leaf Silage. Fermentation 2021, 7, 185. https://doi.org/10.3390/fermentation7030185
Chen D, Zhou W, Guo X, Zheng M, Chen X, Zhang Q. Citric Acid Influences the Dynamics of the Fermentation Quality, Protease Activity and Microbial Community of Mulberry Leaf Silage. Fermentation. 2021; 7(3):185. https://doi.org/10.3390/fermentation7030185
Chicago/Turabian StyleChen, Dekui, Wei Zhou, Xiang Guo, Mingyang Zheng, Xiaoyang Chen, and Qing Zhang. 2021. "Citric Acid Influences the Dynamics of the Fermentation Quality, Protease Activity and Microbial Community of Mulberry Leaf Silage" Fermentation 7, no. 3: 185. https://doi.org/10.3390/fermentation7030185
APA StyleChen, D., Zhou, W., Guo, X., Zheng, M., Chen, X., & Zhang, Q. (2021). Citric Acid Influences the Dynamics of the Fermentation Quality, Protease Activity and Microbial Community of Mulberry Leaf Silage. Fermentation, 7(3), 185. https://doi.org/10.3390/fermentation7030185