
Selection Process of a Mixed Inoculum of Non-Saccharomyces Yeasts Isolated in the D.O.Ca. Rioja

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Isolates
2.2. Genetic Characterization of the Strains at Clonal Level
2.3. SO2 Resistance
2.4. Enzymatic Characterization
2.5. Vinifications
2.5.1. Pure 100 mL Vinifications
2.5.2. Sequential 2000 mL Vinifications
2.6. Microbial Analysis and Implantation Controls of the Seeded Yeasts
2.7. Analysis of Enological Parameters in Wines
2.8. Analysis of Aromatic Compounds
2.9. Analysis of Stilbenes
2.10. Statistical Analysis
3. Results
3.1. Clonal Characterization of Non-Saccharomyces Strains
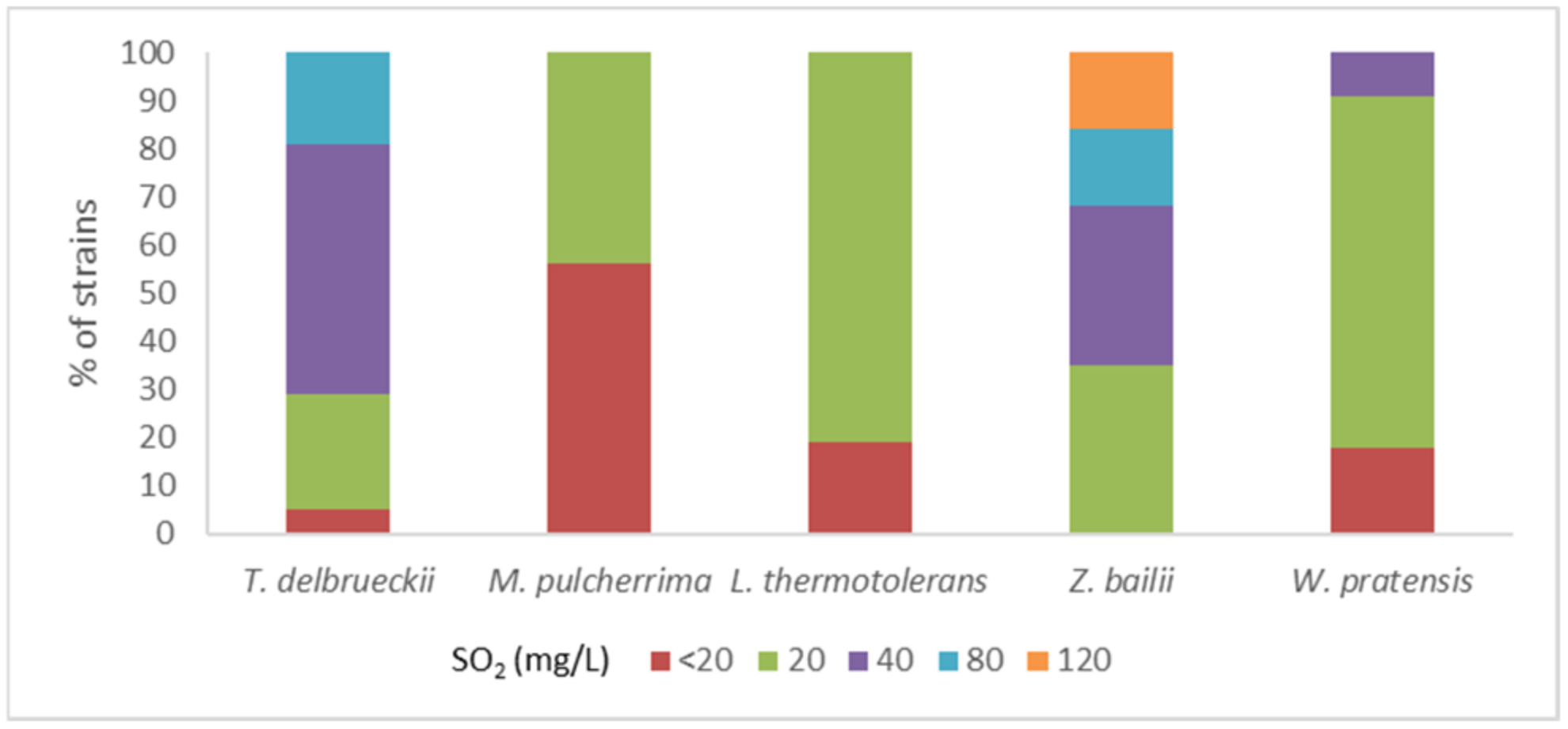
3.2. SO2 Resistance
3.3. Screening of Enzymatic Activities
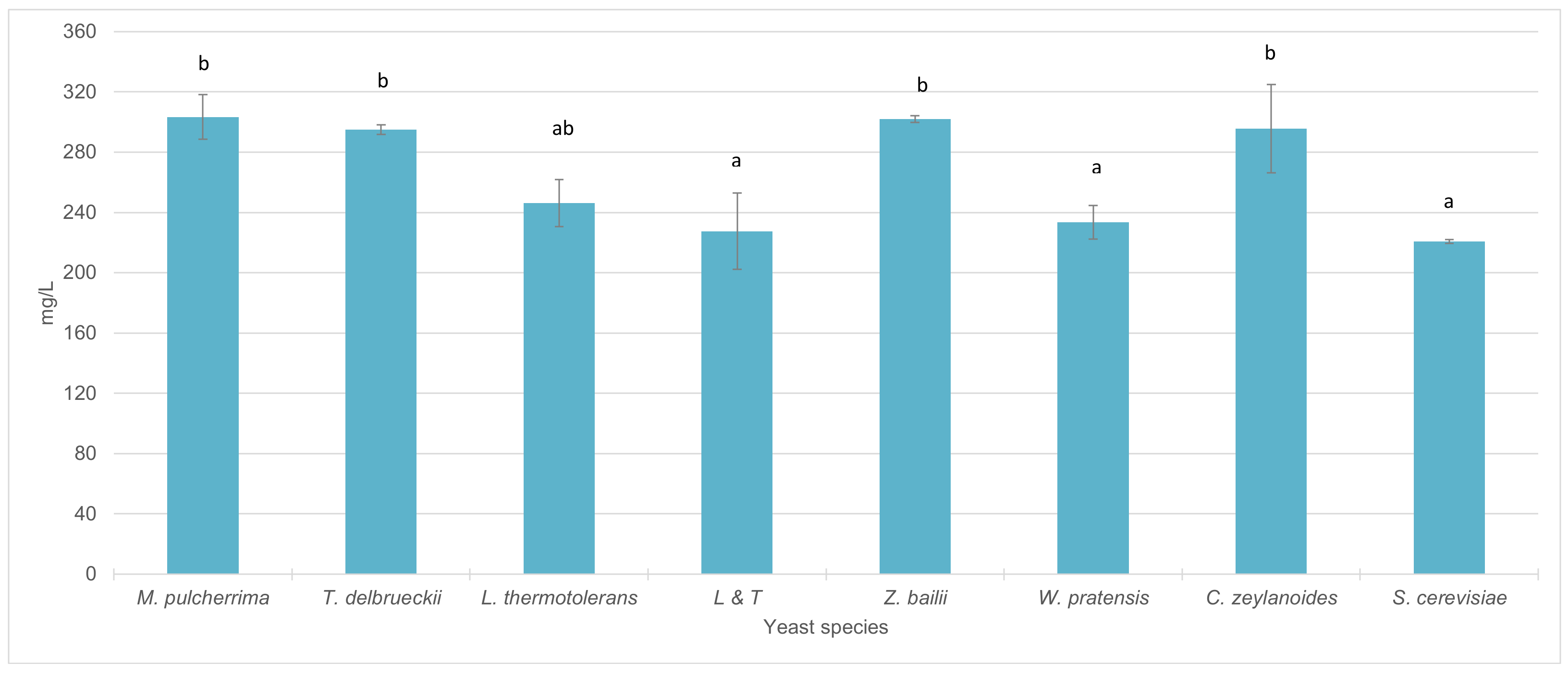
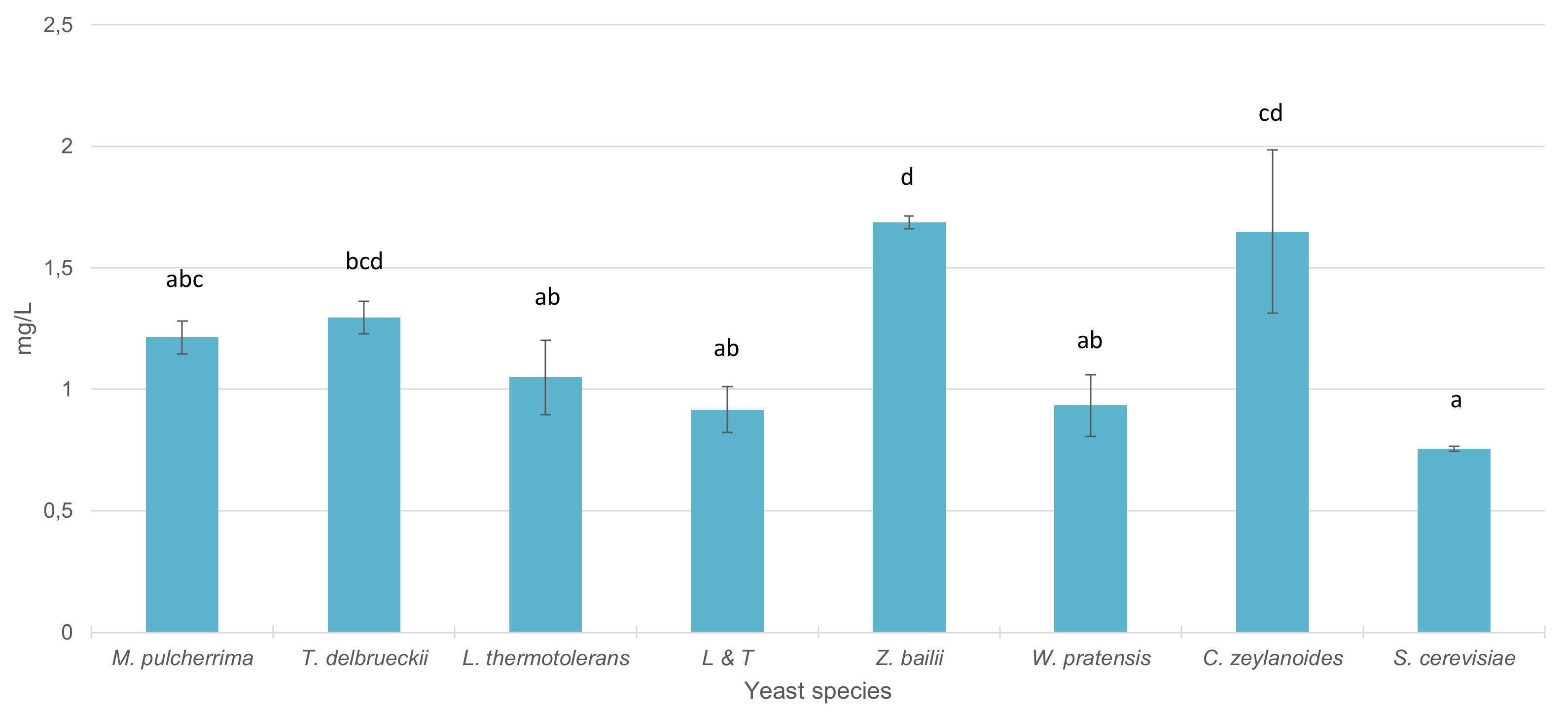
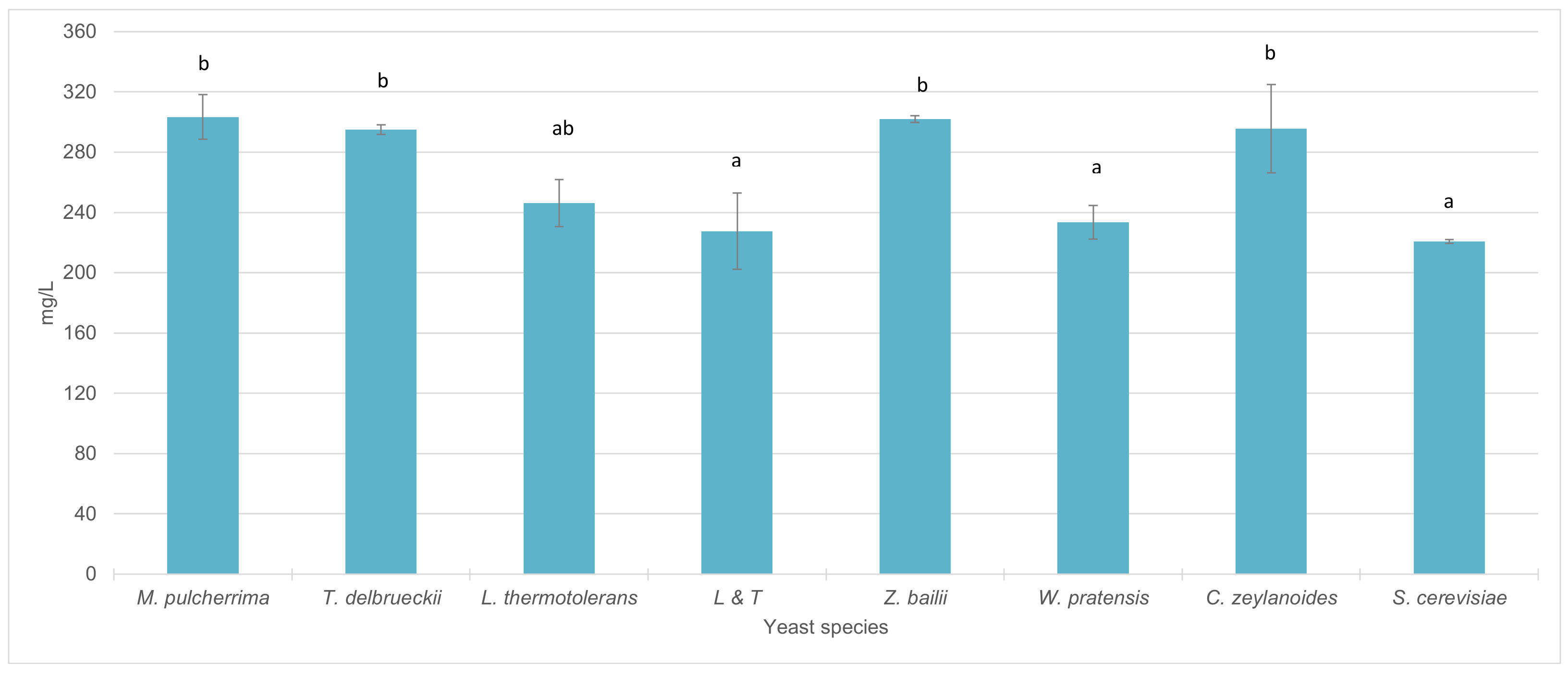
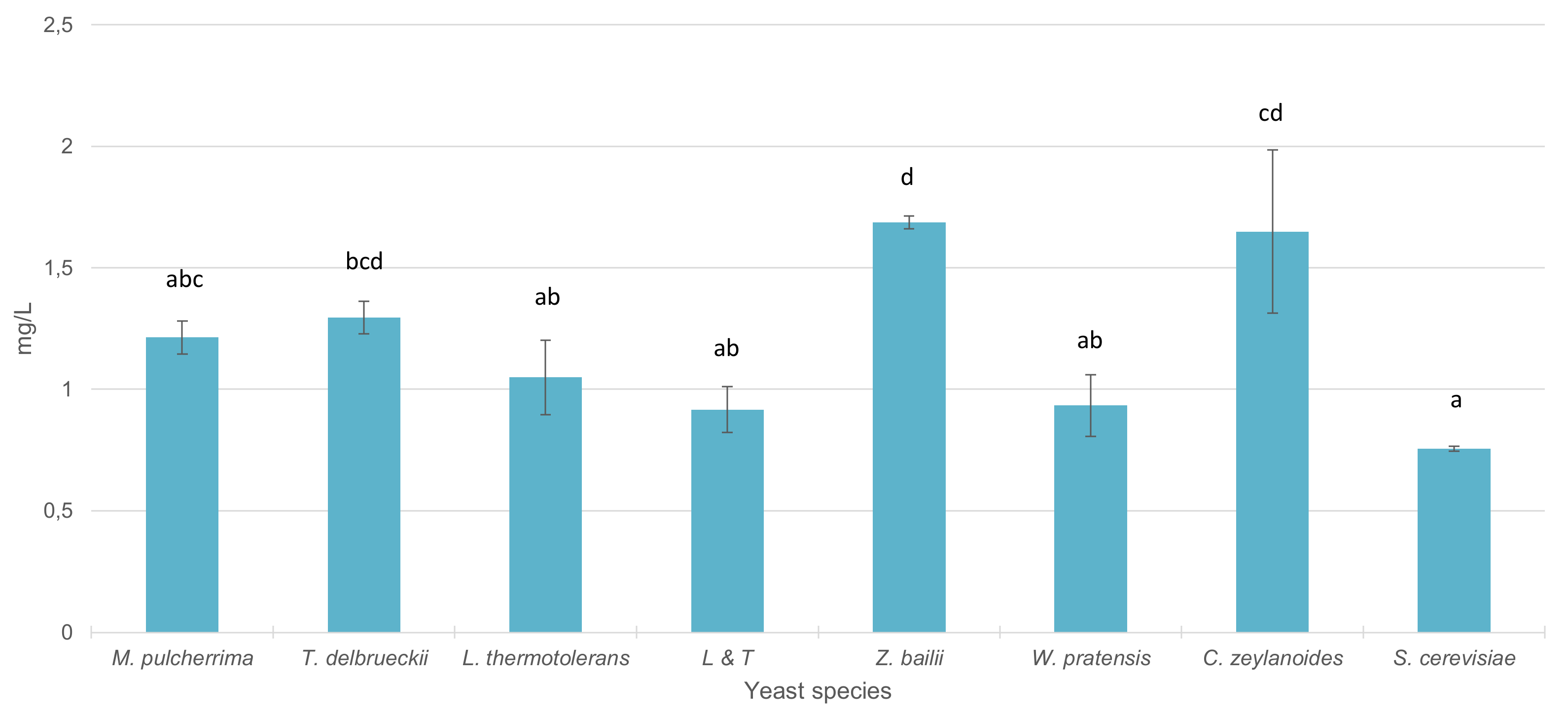
3.4. Fermentative Behavior and Aromatic Compound Production of Non-Saccharomyces Strains in Pure 100 mL Vinifications
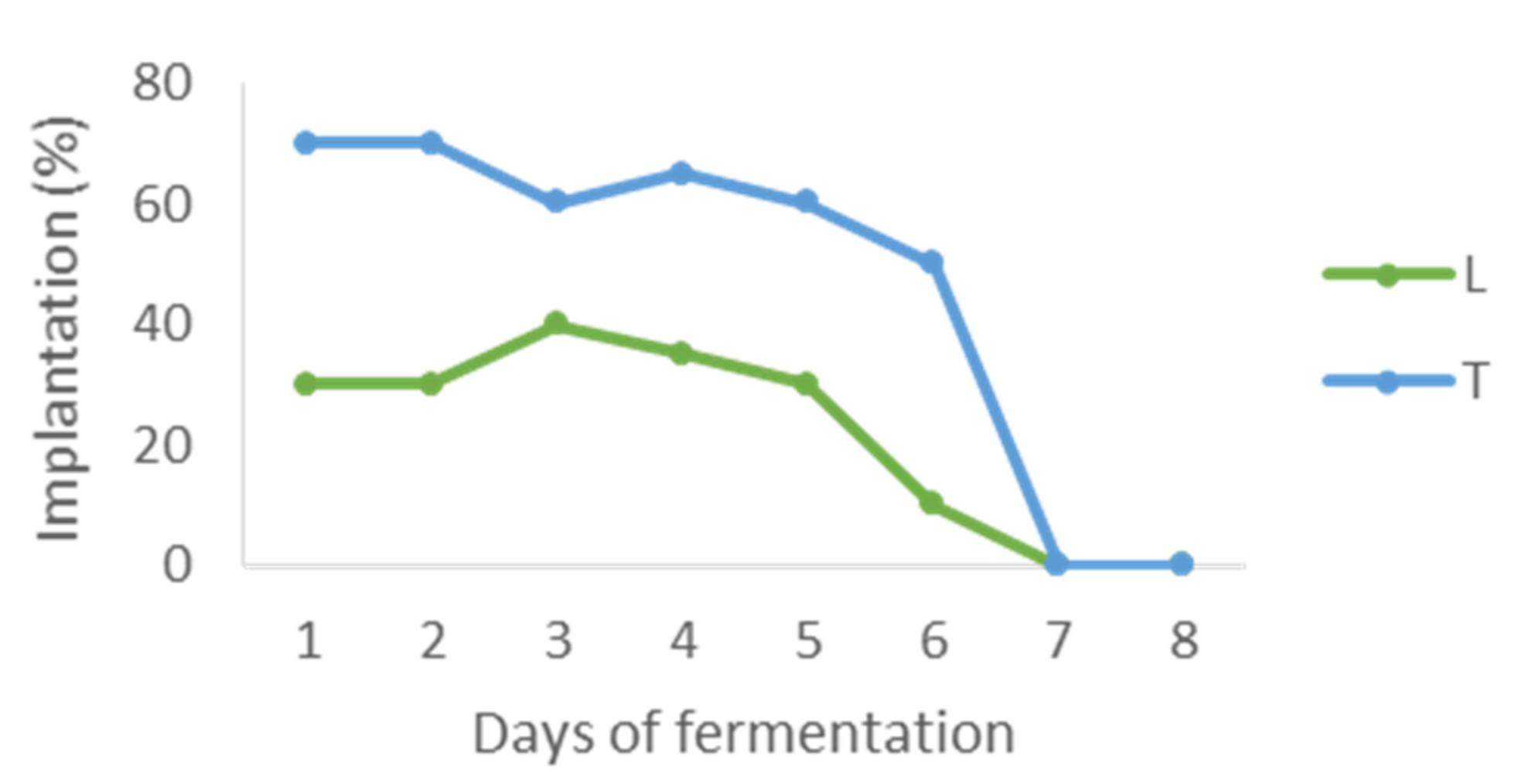
3.5. Influence of Sequential Inoculations with Non-Saccharomyces/S. cerevisiae on Wine Aroma and Composition of Anthocyanins and Stilbenes in 2000 mL Vinifications
4. Discussion
4.1. Clonal Characterization and SO2 Resistance
4.2. Screening of Enzymatic Activities
4.3. Fermentative Behavior and Aromatic Compound Production of Non-Saccharomyces Strains in Pure 100 mL Vinifications
4.4. Influence of Sequential Inoculations with Non-Saccharomyces/S. cerevisiae on Wine Aroma and Composition of Anthocyans and Stilbenes in 2000 mL Vinifications
4.5. Selection of the Inoculum for Red Winemaking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Varela, C.; Siebert, T.; Cozzolino, D.; Rose, L.; McLean, H.; Henschke, P.A. Discovering a chemical basis for differentiating wines made by fermentation with “wild” indigenous and inoculated yeasts: Role of yeast volatile compounds. Aust. J. Grape Wine Res. 2009, 15, 238–248. [Google Scholar] [CrossRef]
- Romano, P.; Capece, A.; Jespersen, L. Taxonomic and Ecological Diversity of Food and Beverage Yeasts; Springer: Berlin/Heidelberg, Germany, 2006; ISBN 4535283230. [Google Scholar]
- Rainieri, S.; Pretorius, I.S. Selection and improvement of wine yeasts. Ann. Microbiol. 2000, 50, 15–31. [Google Scholar]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The Role and Use of Non-Saccharomyces Yeasts in Wine Production. S. Afr. J. Enol. Vitic. 2006, 27, 15–39. [Google Scholar] [CrossRef]
- Rojas, V.; Gil, J.V.; Piñaga, F.; Manzanares, P. Acetate ester formation in wine by mixed cultures in laboratory fermentations. Int. J. Food Microbiol. 2003, 86, 181–188. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces Cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its Importance to Wine Aroma—A Review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Arneborg, N.; Toldam-Andersen, T.B.; Zhang, S.; Petersen, M.A.; Bredie, W.L.P. Impact of sequential co-culture fermentations on flavour characters of Solaris wines. Eur. Food Res. Technol. 2017, 243, 437–445. [Google Scholar] [CrossRef]
- Loira, I.; Morata, A.; Comuzzo, P.; Callejo, M.J.; González, C.; Calderón, F.; Suárez-Lepe, J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 2015, 76, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarède, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii-Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 2008, 122, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderon, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 2015, 99, 1911–1922. [Google Scholar] [CrossRef]
- Renault, P.; Coulon, J.; de Revel, G.; Barbe, J.C.; Bely, M. Increase of fruity aroma during mixed T. delbrueckii/S. cerevisiae wine fermentation is linked to specific esters enhancement. Int. J. Food Microbiol. 2015, 207, 40–48. [Google Scholar] [CrossRef]
- Azzolini, M.; Tosi, E.; Lorenzini, M.; Finato, F.; Zapparoli, G. Contribution to the aroma of white wines by controlled Torulaspora delbrueckii cultures in association with Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2015, 31, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Ruiz, J.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Benito, S.; Santos, A. Influence of Torulaspora delbrueckii in varietal thiol (3-SH and 4-MSP) release in wine sequential fermentations. Int. J. Food Microbiol. 2017, 257, 183–191. [Google Scholar] [CrossRef]
- Kapsopoulou, K.; Mourtzini, A.; Anthoulas, M.; Nerantzis, E. Biological acidification during grape must fermentation using mixed cultures of Kluyveromyces thermotolerans and Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2007, 23, 735–739. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef]
- Vilela, A. Lachancea thermotolerans, the Non-Saccharomyces yeast that reduces the volatile acidity of wines. Fermentation 2018, 4, 56. [Google Scholar] [CrossRef] [Green Version]
- Hranilovic, A.; Gambetta, J.M.; Schmidtke, L.; Boss, P.K.; Grbin, P.R.; Masneuf-Pomarede, I.; Bely, M.; Albertin, W.; Jiranek, V. Oenological traits of Lachancea thermotolerans show signs of domestication and allopatric differentiation. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Ciani, M.; Beco, L.; Comitini, F. Fermentation behaviour and metabolic interactions of multistarter wine yeast fermentations. Int. J. Food Microbiol. 2006, 108, 239–245. [Google Scholar] [CrossRef]
- Divol, B.; Du Toit, M.; Duckitt, E. Surviving in the presence of sulphur dioxide: Strategies developed by wine yeasts. Appl. Microbiol. Biotechnol. 2012, 95, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Krieger, S. Application of yeast and bacteria as starter cultures. In Biology of Microorganisms on Grapes, in Must and in Wine; König, H., Unden, G.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 605–634. [Google Scholar]
- Fernández-González, M.; Di Stefano, R.; Briones, A. Hydrolysis and transformation of terpene glycosides from muscat must by different yeast species terpene glycosides from muscat. Food Microbiol. 2003, 20, 35–41. [Google Scholar] [CrossRef]
- Zoecklein, B.W.; Marcy, J.E.; Williams, J.M.; Jasinski, Y. Effect of native yeasts and selected strains of Saccharomyces cerevisiae on glycosyl glucose, potential volatile terpenes, and selected aglycones of White Riesling (Vitis vinifera L.) wines. J. Food Compos. Anal. 1997, 10, 55–65. [Google Scholar] [CrossRef]
- Strauss, M.L.A.; Jolly, N.P.; Lambrechts, M.G.; Van Rensburg, P. Screening for the production of extracellular hydrolytic enzymes by non-Saccharomyces wine yeasts. J. Appl. Microbiol. 2001, 91, 182–190. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Nieuwoudt, H.H.; Prior, B.A.; Pretorius, L.S.; Bauer, F.F. Glycerol in South African Table Wines: An Assessment of its Relationship to Wine Quality. S. Afr. J. Enol. Vitic. 2017, 23, 22–30. [Google Scholar] [CrossRef]
- Aranda, A.; Matallana, E.; del Olmo, M. Levaduras. Saccharomyces I. Levaduras de primera fermentación. In Microbiología del Vino; Carrascosa, A.V., Muñoz, R., González, R., Eds.; AMV Ediciones: Madrid, Spain, 2005; pp. 273–297. [Google Scholar]
- Renault, P.; Miot-Sertier, C.; Marullo, P.; Hernández-Orte, P.; Lagarrigue, L.; Lonvaud-Funel, A.; Bely, M. Genetic characterization and phenotypic variability in Torulaspora delbrueckii species: Potential applications in the wine industry. Int. J. Food Microbiol. 2009, 134, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Zott, K.; Thibon, C.; Bely, M.; Lonvaud-Funel, A.; Dubourdieu, D.; Masneuf-Pomarede, I. The grape must non-Saccharomyces microbial community: Impact on volatile thiol release. Int. J. Food Microbiol. 2011, 151, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Schvarczová, E.; Štefániková, J.; Jankura, E.; Kolek, E. Selection of autochthonous Saccharomyces cerevisiae strains for production of typical Pinot Gris wines. J. Food Nutr. Res. 2017, 56, 389–397. [Google Scholar]
- Querol, A.; Huerta, T.; Barrio, E.; Ramón, D. Dry Yeast Strain For Use in Fermentation of Alicante Wines: Selection and DNA Patterns. J. Food Sci. 1992, 57, 183–185. [Google Scholar] [CrossRef]
- Rodríguez, M.E.; Infante, J.J.; Molina, M.; Domínguez, M.; Rebordinos, L.; Cantoral, J.M. Genomic characterization and selection of wine yeast to conduct industrial fermentations of a white wine produced in a SW Spain winery. J. Appl. Microbiol. 2010, 108, 1292–1302. [Google Scholar] [CrossRef]
- Bae, S.; Fleet, G.H.; Heard, G.M. Lactic acid bacteria associated with wine grapes from several Australian vineyards. J. Appl. Microbiol. 2006, 100, 712–727. [Google Scholar] [CrossRef]
- Lopes, C.A.; Rodríguez, M.E.; Sangorrín, M.; Querol, A.; Caballero, A.C. Patagonian wines: The selection of an indigenous yeast starter. J. Ind. Microbiol. Biotechnol. 2007, 34, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Tristezza, M.; Fantastico, L.; Vetrano, C.; Bleve, G.; Corallo, D.; Mita, G.; Grieco, F. Molecular and technological characterization of Saccharomyces cerevisiae strains isolated from natural fermentation of susumaniello grape must in Apulia, Southern Italy. Int. J. Microbiol. 2014, 2014, 897428. [Google Scholar] [CrossRef] [PubMed]
- Ocón, E.; Garijo, P.; Sanz, S.; Olarte, C.; López, R.; Santamaría, P.; Gutiérrez, A.R. Screening of yeast mycoflora in winery air samples and their risk of wine contamination. Food Control 2013, 34, 261–267. [Google Scholar] [CrossRef]
- Ocón, E.; Gutiérrez, A.R.; Garijo, P.; Tenorio, C.; López, I.; López, R.; Santamaría, P. Quantitative and qualitative analysis of non-Saccharomyces yeasts in spontaneous alcoholic fermentations. Eur. Food Res. Technol. 2010, 230, 885–891. [Google Scholar] [CrossRef]
- Ocón, E.; Gutiérrez, A.R.; Garijo, P.; López, R.; Santamaría, P. Presence of non-Saccharomyces yeasts in cellar equipment and grape juice during harvest time. Food Microbiol. 2010, 27, 1023–1027. [Google Scholar] [CrossRef]
- González-Arenzana, L.; Garijo, P.; Berlanas, C.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Genetic and phenotypic intraspecific variability of non-Saccharomyces yeasts populations from La Rioja winegrowing region (Spain). J. Appl. Microbiol. 2017, 122, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Escribano, R.; González-Arenzana, L.; Garijo, P.; Berlanas, C.; López-Alfaro, I.; López, R.; Gutiérrez, A.R.; Santamaría, P. Screening of enzymatic activities within different enological non-Saccharomyces yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef] [Green Version]
- Sampaio, T.L.; Kennedy, J.A.; Vasconcelos, M.C. Use of microscale fermentations in grape and wine research. Am. J. Enol. Vitic. 2007, 58, 534–539. [Google Scholar]
- López, I.; Torres, C.; Ruiz-Larrea, F. Genetic typification by pulsed-field gel electrophoresis (PFGE) and randomly amplified polymorphic DNA (RAPD) of wild Lactobacillus plantarum and Oenococcus oeni wine strains. Eur. Food Res. Technol. 2008, 227, 547–555. [Google Scholar] [CrossRef]
- Kurtzman, C.; Fell, J.W. The Yeasts: A Taxonomic Study, 4th ed.; Elsevier Science: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Commission Regulation. Laying Down Certain Detailed Rules for Implementing Council Regulation (EC) No 479/2008 as Regards the Categories of Grapevine Products, Oenological Practices and the Applicable Restrictions; (EC) No 606/2009 of 10 July 2009; European Comission: Brussels, Belgium, 2009; pp. 1–59. [Google Scholar]
- Somers, T.C.; Evans, M.E. Wine quality: Correlations with colour density and anthocyanin equilibria in a group of young red wines. J. Sci. Food Agric. 1974, 25, 1369–1379. [Google Scholar] [CrossRef]
- Ortega, C.; López, R.; Cacho, J.; Ferreira, V. Fast analysis of important wine volatile compounds. J. Chromatogr. A 2001, 923, 205–214. [Google Scholar] [CrossRef]
- López, R.; López-Alfaro, I.; Gutiérrez, A.R.; Tenorio, C.; Garijo, P.; González-Arenzana, L.; Santamaría, P. Malolactic fermentation of Tempranillo wine: Contribution of the lactic acid bacteria inoculation to sensory quality and chemical composition. Int. J. Food Sci. Technol. 2011, 46, 2373–2381. [Google Scholar] [CrossRef]
- Portu, J.; López-Alfaro, I.; Gómez-Alonso, S.; López, R.; Garde-Cerdán, T. Changes on grape phenolic composition induced by grapevine foliar applications of phenylalanine and urea. Food Chem. 2015, 180, 171–180. [Google Scholar] [CrossRef]
- Portu, J.; López, R.; Ewald, P.; Santamaría, P.; Winterhalter, P.; Garde-Cerdán, T. Evaluation of Grenache, Graciano and Tempranillo grape stilbene content after field applications of elicitors and nitrogen compounds. J. Sci. Food Agric. 2018, 98, 1856–1862. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, A.R.; Santamaría, P.; González-Arenzana, L.; Garijo, M.P.; Berlanas, C.; López-Alfaro, I.; López, R. SO2 tolerance in non-Saccharomyces yeasts. In Proceedings of the 39th World Congress of Vine and Wine, Bento Gonçalves, Brazil, 24–28 October 2016. [Google Scholar]
- Trinh, T.T.T.; Woon, W.Y.; Yu, B.; Curran, P.; Liu, S.Q. Effect of L-isoleucine and L-phenylalanine addition on aroma compound formation during longan juice fermentation by a co-culture of Saccharomyces cerevisiae and Williopsis saturnus. S. Afr. J. Enol. Vitic. 2010, 31, 116–124. [Google Scholar] [CrossRef]
- Santamaría, P.; López, R.; Portu, J.; González-Arenzana, L.; López-Alfaro, I.; Gutiérrez, A.R.; Garde-Cerdán, T. Role of phenylalanine in viticulture and enology. In Phenylalanine. Dietary, Sources, Functions and Health Effects; Elliamson, D., Ed.; Nova Publisher: New York, NY, USA, 2015; pp. 49–70. [Google Scholar]
- Dizy, M.; Bisson, L.F. Proteolytic activity of yeast strains during grape juice fermentation. Am. J. Enol. Vitic. 2000, 51, 155–167. [Google Scholar]
- Escribano, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aromatic compound production and fermentative behaviour within different non-Saccharomyces species and clones. J. Appl. Microbiol. 2018, 124, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Rapp, A.; Versini, G. Influence of nitrogen compounds in grapes on aroma compounds of wines. Dev. Food Sci. 1995, 37, 1659–1694. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud-Funel, A. Handbook of Enology; John Willey and Son: West Sussex, UK, 2007; Volume 2. [Google Scholar]
- Romano, P.; Suzzi, G. Origin and production of acetoin during wine yeast fermentation. Appl. Environ. Microbiol. 1996, 62, 309–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escribano-Viana, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aroma evolution throughout alcoholic fermentation sequentially inoculated with non- Saccharomyces/Saccharomyces yeasts. Food Res. Int. 2018, 112, 17–24. [Google Scholar] [CrossRef]
- Capece, A.; Fiore, C.; Maraz, A.; Romano, P. Molecular and technological approaches to evaluate strain biodiversity in Hanseniaspora uvarum of wine origin. J. Appl. Microbiol. 2005, 98, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, D.; Bauer, F.F. Exploring the phenotypic space of non-Saccharomyces wine yeast biodiversity. Food Microbiol. 2016, 55, 32–46. [Google Scholar] [CrossRef]
- Fernández, M.; Úbeda, J.F.; Briones, A.I. Typing of non-Saccharomyces yeasts with enzymatic activities of interest in wine-making. Int. J. Food Microbiol. 2000, 59, 29–36. [Google Scholar] [CrossRef]
- Ferreira, A.M.; Clímaco, M.C.; Faia, A.M. The role of non-Saccharomyces species in releasing glycosidic bound fraction of grape aroma components—A preliminary study. J. Appl. Microbiol. 2001, 91, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Maturano, Y.P.; Rodríguez Assaf, L.A.; Toro, M.E.; Nally, M.C.; Vallejo, M.; Castellanos de Figueroa, L.I.; Combina, M.; Vazquez, F. Multi-enzyme production by pure and mixed cultures of Saccharomyces and non-Saccharomyces yeasts during wine fermentation. Int. J. Food Microbiol. 2012, 155, 43–50. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Palomero, F.; Benito, S. Combine use of selected Schizosaccharomyces pombe and Lachancea thermotolerans yeast strains as an alternative to the traditional malolactic fermentation in red wine production. Molecules 2015, 20, 9510–9523. [Google Scholar] [CrossRef] [Green Version]
- Taing, O.; Taing, K. Production of malic and succinic acids by sugar-tolerant yeast Zygosaccharomyces rouxii. Eur. Food Res. Technol. 2007, 224, 343–347. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masneuf-Pomarede, I.; Bely, M.; Marullo, P.; Albertin, W. The genetics of non-conventional wine yeasts: Current knowledge and future challenges. Front. Microbiol. 2016, 6. [Google Scholar] [CrossRef]
- Clemente-Jimenez, J.M.; Mingorance-Cazorla, L.; Martínez-Rodríguez, S.; Las Heras-Vázquez, F.J.; Rodríguez-Vico, F. Molecular characterization and oenological properties of wine yeasts isolated during spontaneous fermentation of six varieties of grape must. Food Microbiol. 2004, 21, 149–155. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Prior, K.J.; Bauer, F.F.; Divol, B. The utilisation of nitrogenous compounds by commercial non-Saccharomyces yeasts associated with wine. Food Microbiol. 2019, 79, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, M.E.; Lopes, C.A.; Barbagelata, R.J.; Barda, N.B.; Caballero, A.C. Influence of Candida pulcherrima Patagonian strain on alcoholic fermentation behaviour and wine aroma. Int. J. Food Microbiol. 2010, 138, 19–25. [Google Scholar] [CrossRef]
- Ciani, M.; Maccarelli, F. Oenological properties of non-Saccharomyces yeasts associated with wine-making. World J. Microbiol. Biotechnol. 1998, 14, 199–203. [Google Scholar] [CrossRef]
- van Breda, V.; Jolly, N.; van Wyk, J. Characterisation of commercial and natural Torulaspora delbrueckii wine yeast strains. Int. J. Food Microbiol. 2013, 163, 80–88. [Google Scholar] [CrossRef]
- Del Fresno, J.M.; Morata, A.; Loira, I.; Bañuelos, M.A.; Escott, C.; Benito, S.; González Chamorro, C.; Suárez-Lepe, J.A. Use of non-Saccharomyces in single-culture, mixed and sequential fermentation to improve red wine quality. Eur. Food Res. Technol. 2017, 243, 2175–2185. [Google Scholar] [CrossRef]
- Chen, K.; Escott, C.; Loira, I.; del Fresno, J.M.; Morata, A.; Tesfaye, W.; Calderon, F.; Suárez-Lepe, J.A.; Han, S.; Benito, S. Use of non-Saccharomyces yeasts and oenological tannin in red winemaking: Influence on colour, aroma and sensorial properties of young wines. Food Microbiol. 2018, 69, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Conchillo, L.B.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Selection and use of pectinolytic yeasts for improving clarification and phenolic extraction in winemaking. Int. J. Food Microbiol. 2016, 223, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Clare, S.S.; Skurray, G.R.; Shalliker, R.A. Effect of yeast strain selection on the concentration of cis- and trans-resveratrol and resveratrol glucoside isomers in wine. Aust. J. Grape Wine Res. 2005, 11, 9–14. [Google Scholar] [CrossRef]
- Kostadinović, S.; Wilkens, A.; Stefova, M.; Ivanova, V.; Vojnoski, B.; Mirhosseini, H.; Winterhalter, P. Stilbene levels and antioxidant activity of Vranec and Merlot wines from Macedonia: Effect of variety and enological practices. Food Chem. 2012, 135, 3003–3009. [Google Scholar] [CrossRef]
- Gaensly, F.; Agustini, B.C.; da Silva, G.A.; Picheth, G.; Bonfim, T.M.B. Autochthonous yeasts with β-glucosidase activity increase resveratrol concentration during the alcoholic fermentation of Vitis labrusca grape must. J. Funct. Foods 2015, 19, 288–295. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Profiles | N | Enzymatic Activities | ||||||
|---|---|---|---|---|---|---|---|---|
| Esterase | Esterase-Lipase | Lipase | LeucineA | ValineA | CystineA | β-Glucosidase | ||
| Candida spp. (C) | ||||||||
| CA 1 | 5 | + | + | − | + | + | − | − |
| CA 2 | 3 | + | + | − | + | − | − | − |
| CA 3 | 1 | − | + | − | + | + | − | − |
| CA 4 | 1 | − | + | − | + | + | − | + |
| CA 5 | 2 | + | + | − | + | + | − | + |
| Cryptococcus spp. (CC) | ||||||||
| CCA 1 | 2 | + | + | − | + | − | − | + |
| CCA 2 | 1 | + | + | − | + | − | − | + |
| CCA 3 | 2 | + | + | − | + | − | − | − |
| Debaryomyces hansenii (D) | ||||||||
| DA 1 | 1 | − | − | − | − | − | − | − |
| DA 2 | 1 | + | + | − | − | − | − | − |
| DA 3 | 1 | + | + | − | − | − | − | − |
| Lachancea thermotolerans (L) | ||||||||
| LA 1 | 11 | + | + | − | + | + | − | − |
| LA 2 | 1 | + | + | − | + | − | − | − |
| LA 3 | 1 | − | + | − | + | + | − | − |
| LA 4 | 1 | + | + | + | + | + | − | − |
| LA 5 | 1 | + | + | + | + | + | − | + |
| LA 6 | 1 | + | + | − | + | + | + | − |
| Metschnikowia pulcherrima (M) | ||||||||
| MA 1 | 5 | + | + | − | + | − | − | − |
| MA 2 | 2 | + | + | − | + | − | − | + |
| MA 3 | 1 | + | + | − | − | + | − | − |
| MA 4 | 5 | + | + | − | + | + | − | + |
| MA 5 | 2 | + | + | − | + | − | − | + |
| MA 6 | 1 | + | + | − | + | + | − | + |
| Pichia kluyveri (PK) | ||||||||
| PKA 1 | 4 | + | + | − | + | − | − | − |
| Sporodiobulus salmonicolor (SP) | ||||||||
| SPA 1 | 4 | + | + | − | + | + | − | − |
| SPA 2 | 2 | + | + | + | + | + | − | − |
| Torulaspora delbrueckii (T) | ||||||||
| TA 1 | 1 | + | + | − | − | − | − | − |
| TA 2 | 5 | − | − | − | − | − | − | − |
| TA 3 | 8 | + | + | − | + | − | − | − |
| TA 4 | 3 | − | − | − | + | − | − | − |
| TA 5 | 1 | + | + | − | + | − | + | − |
| Williopsis pratensis (W) | ||||||||
| WA 1 | 9 | + | + | − | + | + | + | − |
| WA 2 | 1 | + | + | − | + | + | − | − |
| Zygosaccharomyces bailii (Z) | ||||||||
| ZA 1 | 1 | + | + | − | + | − | − | − |
| ZA 2 | 1 | + | − | − | + | − | − | − |
| ZA 3 | 1 | + | + | − | + | − | − | + |
| ZA 4 | 2 | − | + | − | + | − | − | − |
| ZA 5 | 2 | + | + | − | + | − | − | − |
| Species/Profiles | N | Enzymatic Activities | ||||||
|---|---|---|---|---|---|---|---|---|
| Pectinase | Cellulase | Xilanase | Glucanase | LeucineA | ValineA | CystineA | ||
| Candida spp. (C) | ||||||||
| CC 1 | 3 | + | − | − | − | + | + | − |
| CC 2 | 3 | + | − | − | − | + | − | − |
| CC 3 | 6 | − | − | − | − | + | + | − |
| Cryptococcus spp. (CC) | ||||||||
| CCC 1 | 2 | − | + | − | + | + | − | − |
| CCC 2 | 1 | + | + | − | + | + | − | − |
| CCC 3 | 2 | + | − | − | − | + | − | − |
| Debaryomyces hansenii (D) | ||||||||
| DC 1 | 1 | + | − | − | + | − | − | − |
| DC 2 | 1 | + | + | − | + | − | − | − |
| DC 3 | 1 | + | − | − | − | − | − | − |
| Lachancea thermotolerans (L) | ||||||||
| LC 1 | 9 | − | + | + | + | + | + | − |
| LC 2 | 2 | − | + | − | + | + | + | − |
| LC 3 | 1 | − | − | − | + | + | + | − |
| LC 4 | 2 | − | − | − | − | + | + | − |
| LC 5 | 1 | + | − | − | − | + | + | − |
| LC 6 | 1 | + | + | + | + | + | + | + |
| Metschnikowia pulcherrima (M) | ||||||||
| MC 1 | 5 | + | − | − | − | + | − | − |
| MC 2 | 2 | + | + | − | − | − | − | − |
| MC 3 | 2 | + | − | − | − | + | + | − |
| MC 4 | 1 | + | + | − | − | + | − | − |
| MC 5 | 2 | − | + | − | − | + | + | − |
| MC 6 | 1 | − | − | − | + | + | + | − |
| Pichia kluyveri (PK) | ||||||||
| PKC 1 | 4 | + | − | − | − | + | − | − |
| Sporodiobulus salmonicolor (SP) | ||||||||
| SPC 1 | 4 | + | − | − | − | + | + | − |
| SPC 2 | 2 | − | − | − | − | + | + | − |
| Torulaspora delbrueckii (T) | ||||||||
| TC 1 | 5 | + | − | − | − | − | − | − |
| TC 2 | 1 | − | − | − | − | − | − | − |
| TC 3 | 2 | − | − | − | − | + | − | − |
| TC 4 | 2 | + | + | + | − | + | − | − |
| TC 5 | 7 | + | − | − | − | + | − | − |
| TC 6 | 1 | + | − | − | − | + | − | + |
| Williopsis pratensis (W) | ||||||||
| WC 1 | 9 | − | − | − | − | + | + | + |
| WC 2 | 1 | − | − | − | − | + | − | − |
| Zygosaccharomyces bailii (Z) | ||||||||
| ZC 1 | 2 | + | + | − | − | + | − | − |
| ZC 2 | 3 | + | − | − | − | + | − | − |
| ZC 3 | 2 | − | − | − | − | + | − | − |
| Parameters | Yeast Species | ||||||
|---|---|---|---|---|---|---|---|
| D. hansenii | C. zeylanoides | M. pulcherrima | T. delbrueckii | L. thermotolerans | Z. bailii | S. cerevisiae | |
| Residual sugars (g/L) | 201.0 e | 151.1 d | 111.2 cd | 47.2 b | 75.6 bc | 96.6 c | 0.08 a |
| % sugar (72 h) | 00.0 a | 10.5 ab | 15.5 b | 24.5 bc | 23.0 b | 12.0 ab | 39.5 c |
| Ethanol (% v/v) | 0.11 a | 2.99 b | 5.31 c | 9.83 e | 8.22 de | 6.33 cd | 12.60 f |
| Glycerol (g/L) | 0.55 a | 3.10 b | 4.80 de | 4.05 c | 4.65 d | 5.60 f | 5.20 ef |
| pH | 3.54 b | 3.45 b | 3.46 b | 3.47 b | 3.26 a | 3.25 a | 3.43 b |
| Total acidity (g/L) | 2.30 a | 3.30 b | 3.50 b | 3.25 b | 5.70 d | 4.40 c | 3.55 b |
| Volatile acidity (g/L) | 0.12 ab | 0.41 e | 0.05 a | 0.22 c | 0.26 cd | 0.34 de | 0.20 bc |
| Acetaldehyde (mg/L) | 0.00 a | 45.0 b | 11.5 a | 7.50 a | 17.5 a | 21.5 ab | 19.5 a |
| Malic acid (g/L) | 0.90 bc | 0.95 c | 0.90 bc | 0.71 a | 0.74 ab | 1.26 d | 0.69 a |
| Lactic acid (g/L) | 0.13 a | 0.13 a | 0.12 a | 0.11 a | 3.81 b | 0.09 a | 0.09 a |
| Parameters | Yeast Species | ||||||
|---|---|---|---|---|---|---|---|
| D. hansenii | C. zeylanoides | M. pulcherrima | T. delbrueckii | L. thermotolerans | Z. bailii | S. cerevisiae | |
| ∑ higher alcohols | 11.2 a | 62.0 ab | 272 e | 178 cd | 215 cde | 130 bc | 264 de |
| ∑ acetates | 0.00 a | 0.05 a | 0.21 a | 0.09 a | 0.20 a | 0.19 a | 1.10 b |
| ∑ esters | 0.01 a | 0.03 a | 0.30 bc | 0.46 d | 0.36 cd | 0.20 b | 0.71 e |
| ∑ Acids | 0.79 ab | 0.96 ab | 0.67 a | 3.57 c | 2.20 b | 1.10 ab | 9.10 d |
| Ethyl acetate | 0.22 a | 2.30 a | 269 c | 33.4 ab | 51.7 b | 31.1 ab | 22.7 ab |
| Acetoin | 69.7 ab | 272 d | 184 c | 4.50 a | 99.3 b | 23.4 ab | 5.20 a |
| Dyacetil | 0.04 a | 0.38 a | 2.10 b | 0.30 a | 0.40 a | 0.10 a | 0.05 a |
| Parameters | Yeast Strains | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T18 | T19 | M28 | M29 | L54 | L57 | LT1 | LT2 | Z71 | W87 | C342 | SC | |
| 1 Propanol | 40.3 cde | 37.7 bcd | 32.3 abc | 30.7 ab | 48.6 e | 44.5 de | 34.7 abc | 41.4 cde | 32.3 abc | 30.0 ab | 26.7 a | 28.1 a |
| 1 butanol | 1.06 bc | 1.26 c | 1.03 abc | 0.92 abc | 1.01 abc | 1.00 abc | 0.99 abc | 1.04 abc | 0.81 ab | 0.87 ab | 0.96 abc | 0.69 a |
| Isobutanol | 50.6 bc | 45.1 abc | 53.1 bc | 47.8 abc | 59.7 c | 59.8 c | 43.3 abc | 44.5 abc | 38.0 ab | 36.6 ab | 37.0 ab | 31.5 a |
| 2 phenyl ethanol | 98.9 c | 81.2 bc | 65.9 ab | 64.1 ab | 54.8 a | 63.0 ab | 54.1 a | 62.1 ab | 52.9 a | 48.2 a | 53.6 a | 47.8 a |
| 1 hexanol | 1.86 ab | 1.78 ab | 1.86 ab | 1.57 ab | 2.11 b | 1.93 ab | 1.86 ab | 1.97 ab | 1.83 ab | 1.56 ab | 1.45 a | 1.82 ab |
| Benzylic alcohol | 0.28 b | 0.27 ab | 0.23 ab | 0.26 ab | 0.26 ab | 0.23 ab | 0.22 ab | 0.18 a | 0.19 ab | 0.20 ab | 0.24 ab | 0.20 ab |
| Methionol | 4.44 e | 3.49 cde | 4.03 de | 3.40 bcde | 1.57 a | 2.49 abcd | 2.30 abcd | 2.61 abcd | 2.21 abc | 1.75 abc | 1.90 abc | 1.72 ab |
| ∑ higher alcohols | 197 d | 171 bcd | 158 abcd | 149 abcd | 169 bcd | 173 cd | 138 abc | 154 abcd | 128 abc | 119 ab | 122 abc | 112 a |
| Isoamyl acetate | 2.24 ab | 1.56 ab | 2.21 ab | 1.79 ab | 1.36 a | 2.08 ab | 2.14 ab | 2.65 ab | 2.55 ab | 3.27 b | 2.50 ab | 2.13 ab |
| 2 phenylethyl acetate | 0.30 bc | 0.19 abc | 0.17 ab | 0.16 abc | 0.14 a | 0.22 abc | 0.19 abc | 0.25 abc | 0.24 abc | 0.33 c | 0.19 abc | 0.13 a |
| ∑ acetates | 2.64 ab | 1.75 ab | 2.38 ab | 2.95 ab | 1.50 a | 2.30 ab | 2.33 ab | 2.90 ab | 2.80 ab | 3.60 b | 2.69 ab | 2.26 ab |
| Ethyl propionate | 0.31 d | 0.26 d | 0.11 ab | 0.10 ab | 0.11 ab | 0.14 bc | 0.16 bc | 0.18 c | 0.12 abc | 0.11 ab | 0.10 ab | 0.07 a |
| Octanoic acid | 1.47 abc | 1.28 ab | 2.04 abc | 1.72 abc | 0.88 a | 1.37 abc | 1.76 abc | 1.75 abc | 2.16 bc | 2.55 bc | 2.39 bc | 2.57 c |
| Ethyl lactate | 1.65 ab | 1.46 ab | 2.65 bcd | 2.50 bc | 16.5 g | 10.6 f | 4.25 e | 3.88 cde | 2.83 bcde | 3.24 cde | 4.11 de | 0.48 a |
| Acetoin | 9.84 cd | 11.3 d | 2.66 a | 2.83 a | 17.0 e | 15.6 e | 8.75 cd | 7.55 bc | 4.72 ab | 3.93 a | 4.30 ab | 2.37 a |
| Dyacetil | 2.46 e | 2.33 de | 1.37 abc | 1.12 abc | 3.46 f | 1.68 cd | 1.45 bc | 1.11 abc | 0.88 ab | 0.83 ab | 0.97 ab | 0.69 a |
| Butirolactone | 1.78 c | 1.57 abc | 1.36 abc | 1.18 a | 1.62 abc | 1.67 abc | 1.58 abc | 1.75 bc | 1.52 abc | 1.47 abc | 1.40 abc | 1.20 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escribano-Viana, R.; González-Arenzana, L.; Garijo, P.; López, R.; Santamaría, P.; Gutiérrez, A.R. Selection Process of a Mixed Inoculum of Non-Saccharomyces Yeasts Isolated in the D.O.Ca. Rioja. Fermentation 2021, 7, 148. https://doi.org/10.3390/fermentation7030148
Escribano-Viana R, González-Arenzana L, Garijo P, López R, Santamaría P, Gutiérrez AR. Selection Process of a Mixed Inoculum of Non-Saccharomyces Yeasts Isolated in the D.O.Ca. Rioja. Fermentation. 2021; 7(3):148. https://doi.org/10.3390/fermentation7030148
Chicago/Turabian StyleEscribano-Viana, Rocío, Lucía González-Arenzana, Patrocinio Garijo, Rosa López, Pilar Santamaría, and Ana Rosa Gutiérrez. 2021. "Selection Process of a Mixed Inoculum of Non-Saccharomyces Yeasts Isolated in the D.O.Ca. Rioja" Fermentation 7, no. 3: 148. https://doi.org/10.3390/fermentation7030148
APA StyleEscribano-Viana, R., González-Arenzana, L., Garijo, P., López, R., Santamaría, P., & Gutiérrez, A. R. (2021). Selection Process of a Mixed Inoculum of Non-Saccharomyces Yeasts Isolated in the D.O.Ca. Rioja. Fermentation, 7(3), 148. https://doi.org/10.3390/fermentation7030148