
Ability of Yeast Metabolic Activity to Reduce Sugars and Stabilize Betalains in Red Beet Juice

 ,
,  , ,
, , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Red Beet Juice
2.2. Sugar Content
2.3. Yeast Strains
2.4. Assimilation Profiles
2.5. Enzymatic Profiles
2.6. Red Beet Juice Fermentation
2.7. Fermentation Efficiency
2.8. Betalain Content
2.9. Sugar Index
2.10. Statistics
3. Results and Discussion
3.1. Sugar Content in Red Beet Juice
3.2. Assimilation Profiles
3.3. Enzymatic Fingerprinting
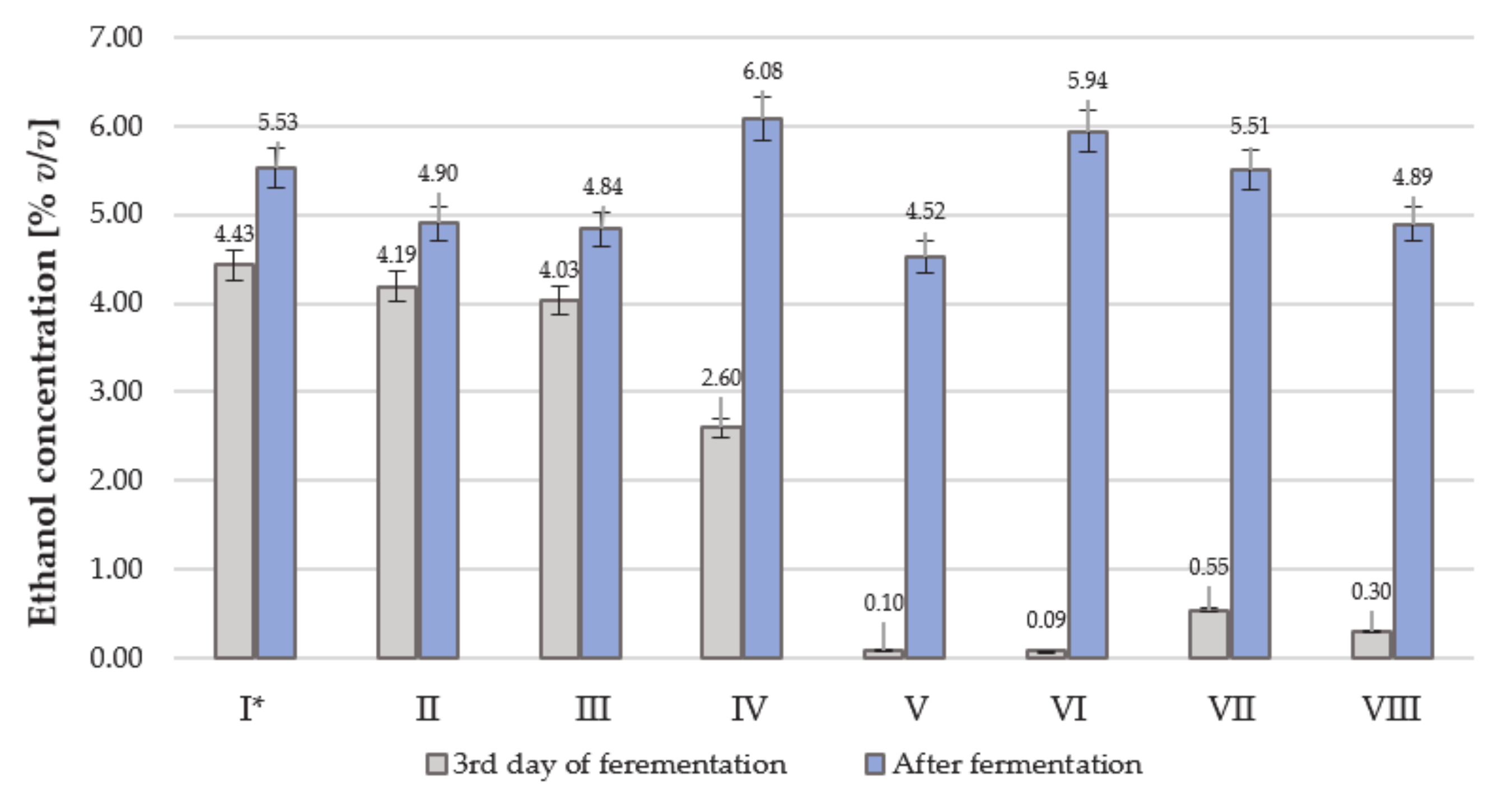
3.4. Red Beet Juice Fermentation
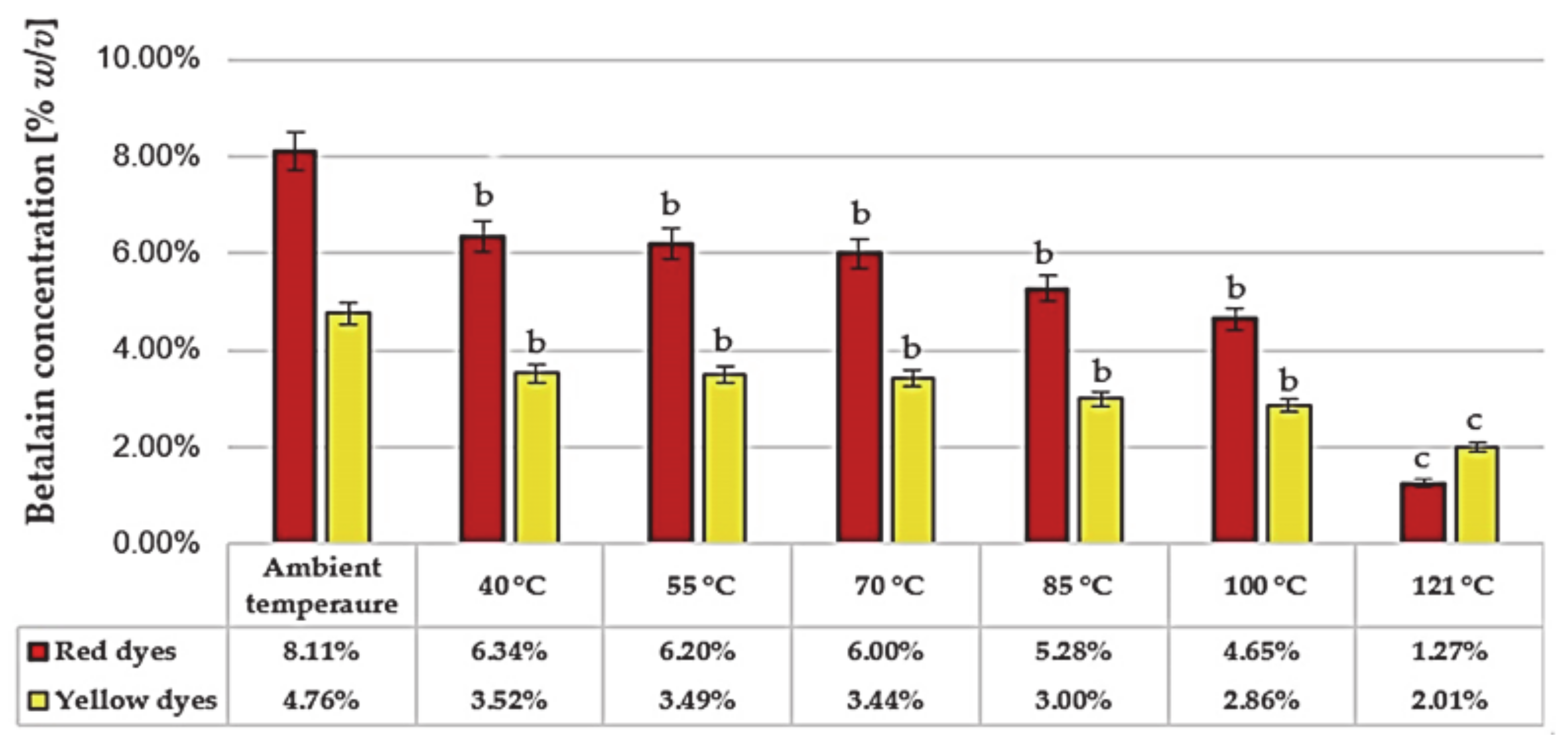
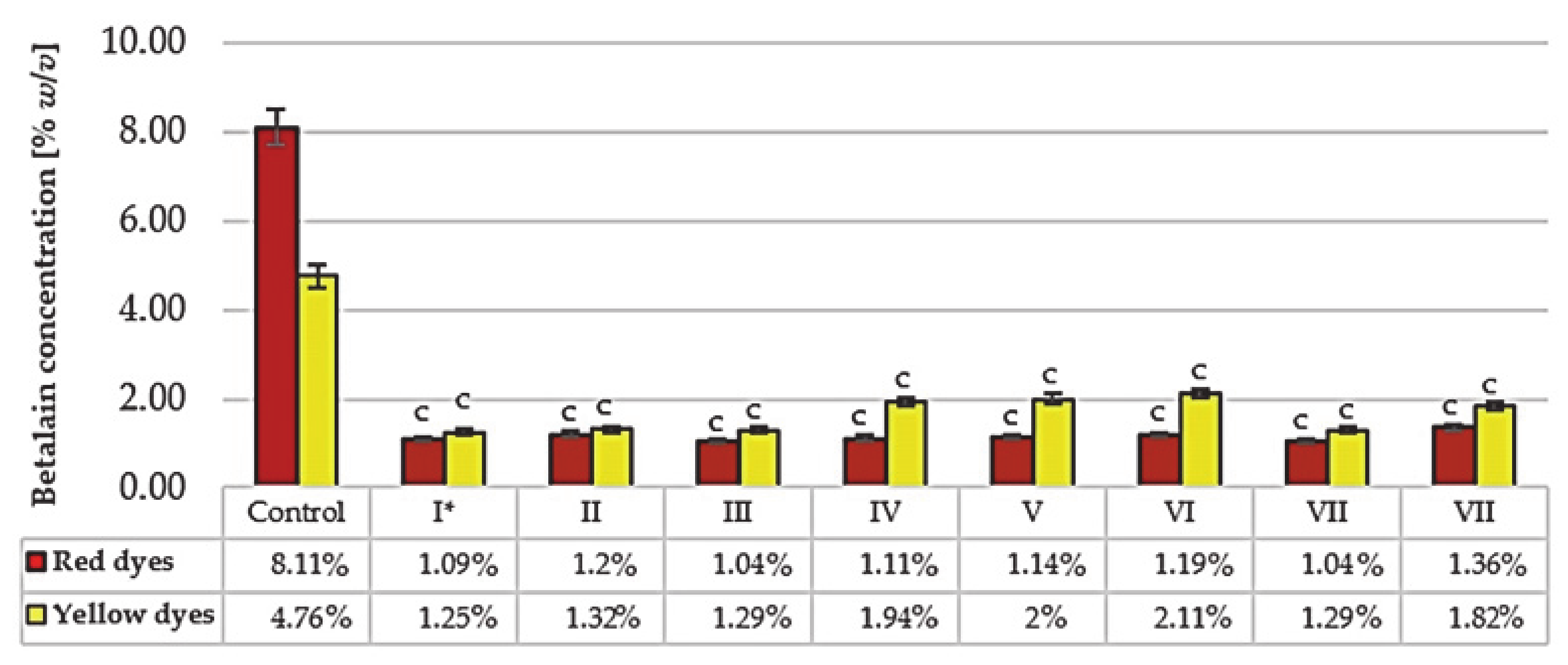
3.5. Control of Betalains and Sugar Index
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Moynihan, P.; Petersen, P.E. Diet, nutrition and the prevention of dental diseases. Public Health Nutr. 2004, 7, 201–226. [Google Scholar] [CrossRef] [PubMed]
- Guldiken, B.; Toydemir, G.; Nur Memis, K.; Okur, S.; Boyacioglu, D.; Capanoglu, E. Home-processed red beetroot (Beta vulgaris L.) products: Changes in antioxidant properties and bioaccessibility. Int. J. Mol. Sci. 2016, 17, 858. [Google Scholar] [CrossRef] [PubMed]
- Doll, S.; Rodier, F.; Willenbrink, J. Accumulation of sucrose in vacuoles isolated from red beet tissue. Planta 1979, 144, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Babarykin, D.; Smirnova, G.; Pundinsh, I.; Vasiljeva, S.; Krumina, G.; Agejchenko, V. Red Beet (Beta vulgaris) Impact on Human Health. J. Biosci. Med. 2019, 07, 61–79. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Singh, A.; Masih, D. Red beetroot: A source of natural colourant and antioxidants: A review. J. Pharmacogn. Phytochem. 2019, 8, 162–166. [Google Scholar]
- Kayın, N.; Atalay, D.; Türken Akçay, T.; Erge, H.S. Color stability and change in bioactive compounds of red beet juice concentrate stored at different temperatures. J. Food Sci. Technol. 2019, 56, 5097–5106. [Google Scholar] [CrossRef] [PubMed]
- Carle, R.; Schweiggert, R.M. Handbook on Natural Pigments in Food and Beverages, 1st ed.; Woodhead Publishing: Cambridge, UK, 2016. [Google Scholar] [CrossRef]
- Gasztonyi, M.N.; Daood, H.; Hájos, M.T.; Biacs, P. Comparison of red beet (Beta vulgaris var. conditiva) varieties on the basis of their pigment components. Proc. J. Sci. Food Agric. 2001, 81, 932–933. [Google Scholar] [CrossRef]
- Escribano, J.; Cabanes, J.; Jiménez-Atiénzar, M.; Ibañez-Tremolada, M.; Gómez-Pando, L.R.; García-Carmona, F.; Gandía-Herrero, F. Characterization of betalains, saponins and antioxidant power in differently colored quinoa (Chenopodium quinoa) varieties. Food Chem. 2017, 234, 285–294. [Google Scholar] [CrossRef]
- Vasconcellos, J.; Conte-Junior, C.; Silva, D.; Pierucci, A.P.; Paschoalin, V.; Alvares, T.S. Comparison of total antioxidant potential, and total phenolic, nitrate, sugar, and organic acid contents in beetroot juice, chips, powder, and cooked beetroot. Food Sci. Biotechnol. 2016, 25, 79–84. [Google Scholar] [CrossRef]
- World Health Organization. Global Status Report on Non-Communicable Diseases; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- World Health Organization. Sugars Intake for Adults and Children; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- World Health Organization. Taxes on Sugary Drinks: Why Do it? WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Kyung, Y.Y.; Woodams, E.E.; Hang, Y.D. Fermentation of beet juice by beneficial lactic acid bacteria. LWT Food Sci. Technol. 2005, 38, 73–75. [Google Scholar] [CrossRef]
- Sawicki, T.; Wiczkowski, W. The effects of boiling and fermentation on betalain profiles and antioxidant capacities of red beetroot products. Food Chem. 2018, 259, 292–303. [Google Scholar] [CrossRef]
- Jafar, N.B.; Ghaleb, Z.T.; Fadhil, Z.H. Production of fermented red beet juice using probiotic lactobacilli bacteria. Ann. Trop. Med. Public Health 2019, 22, 73–75. [Google Scholar] [CrossRef]
- Garcia, C.; Guerin, M.; Souidi, K.; Remize, F. Lactic fermented fruit or vegetable juices: Past, present and future. Beverages 2020, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Kregiel, D. Succinate Dehydrogenase of Saccharomyces cerevisiae—The Unique Enzyme of TCA Cycle—Current Knowledge and New Perspectives. In Dehydrogenases, 1st ed.; Canuto, R.A., Ed.; INTECH: Rijeka, Croatia, 2012; pp. 211–234. [Google Scholar] [CrossRef] [Green Version]
- Imura, M.; Nitta, K.; Iwakiri, R.; Matsuda, F.; Shimizu, H.; Fukusaki, E. Comparison of metabolic profiles of yeasts based on the difference of the Crabtree positive and negative. J. Biosci. Bioeng. 2020, 129, 52–58. [Google Scholar] [CrossRef]
- Joshi, V.K.; Sharma, S.; Devi, M.P. Influence of different yeast strains on fermentation behaviour, physico-chemical and sensory qualities of plum wine. Indian J. Nat. Prod. Resour. 2009, 8, 445–451. [Google Scholar]
- Modelska, M.; Berlowska, J.; Kregiel, D.; Cieciura, W.; Antolak, H.; Tomaszewska, J.; Binczarski, M.; Szubiakiewicz, E.; Witonska, I.A. Concept for recycling waste biomass from the sugar industry for chemical and biotechnological purposes. Molecules 2017, 22, 1544. [Google Scholar] [CrossRef] [Green Version]
- Polish Standard PN-90/A-75101/07. Przetwory Owocowe i Warzywne -- Przygotowanie Próbek i Metody Badań Fizykochemicznych. Oznaczanie Zawartości Cukrów i Ekstraktu Bezcukrowego. Available online: https://www.pkn.pl/en/polish-standard (accessed on 9 May 2021). (In Polish).
- The Grain and Feed Trade Association (GAFTA) Method 10.1. Available online: https://www.gafta.com/write/MediaUploads/Contracts/2018/METHOD_10.1_SUGAR_-_LUFF_SCHOORL_METHOD.pdf (accessed on 9 May 2021).
- Chwil, M.; Kostryco, M. Bioactive compounds and antioxidant activity of Rubus idaeus L. leaves. Acta Sci. Pol. Hortorum Cultus 2018, 17, 135–147. [Google Scholar] [CrossRef]
- Ghanbari, R.; Rezaie, S.; Noorbakhsh, F.; Khaniki, G.J.; Soleimani, M.; Aghaee, E.M. Biocontrol effect of Kluyveromyces lactis on aflatoxin expression and production in Aspergillus parasiticus. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef]
- Papini, M.; Nookaew, I.; Uhlén, M.; Nielsen, J. Scheffersomyces stipitis: A comparative systems biology study with the Crabtree positive yeast Saccharomyces cerevisiae. Microb. Cell Fact. 2012, 11, 136. [Google Scholar] [CrossRef] [Green Version]
- Pawlikowska, E.; James, S.A.; Breierova, E.; Antolak, H.; Kregiel, D. Biocontrol capability of local Metschnikowia sp. isolates. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2019, 112, 1425–1445. [Google Scholar] [CrossRef] [Green Version]
- Mickelsen, P.A.; McCarthy, L.R.; Propst, M.A. Further modifications of the auxanographic method for identification of yeasts. J. Clin. Microbiol. 1977, 5, 5. [Google Scholar] [CrossRef]
- Šavel, J.; Kosin, P.; Broz, A.; Sigler, K. Convenient Monitoring of Brewery Fermentation Course by Refractometry. Kvas. Prum. 2009, 55, 94–99. [Google Scholar] [CrossRef]
- OIV-MA-BS-06. Density of Alcohols and Alcohlic Beverages Method for Determining Electronic Densimetry (Principle Based on Measuring the Period of Oscillation). Available online: https://www.oiv.int/public/medias/2667/oiv-ma-bs-06.pdf (accessed on 13 June 2021).
- Nilsson, T. Studies into the pigments in beetroot (Beta vulgaris L. ssp. vulgaris var. rubra L.). Lantbr. Ann. 1970, 36, 179–219. [Google Scholar]
- Slavov, A.; Karagyozov, V.; Denev, P.; Kratchanova, M.; Kratchanov, C. Antioxidant activity of red beet juices obtained after microwave and thermal pretreatments. Czech. J. Food Sci. 2013, 31, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Richardson, K. Preliminary evaluation of the leaf and root nutrient composition of a fresh market beetroot variety. J. Plant. Nutr. 2014, 20, 408–420. [Google Scholar]
- Chhikara, N.; Kushwaha, K.; Sharma, P.; Gat, Y.; Panghal, A. Bioactive compounds of beetroot and utilization in food processing industry: A critical review. Food Chem. 2019, 272, 192–200. [Google Scholar] [CrossRef]
- Dai, Z.; Huang, M.; Chen, Y.; Siewers, V.; Nielsen, J. Global rewiring of cellular metabolism renders Saccharomyces cerevisiae Crabtree negative. Nat. Commun. 2018, 9, 3059. [Google Scholar] [CrossRef] [Green Version]
- Ji Ho, C.; Hwan Yeo, S.; Park, J.-H.; Choi, H.S.; Gang, J.-E.; In Kim, S.; Tae Jeong, S.; Ra Kim, S. Isolation of aromatic yeasts (non-Saccharomyces cerevisiae) from Korean traditional nuruks; and identification of fermentation characteristics. Agric. Sci. 2013, 4, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.A. Biotechnology of non-Saccharomyces yeasts-the basidiomycetes. Appl. Microbiol. Biotechnol. 2013, 97, 7563–7577. [Google Scholar] [CrossRef]
- Lu, G.; Fellman, J.K.; Edwards, C.G.; Mattinson, D.S.; Navazio, J. Quantitative determination of geosmin in red beets (Beta vulgaris L.) using headspace solid-phase microextraction. J. Agric. Food Chem. 2003, 51, 1021–1025. [Google Scholar] [CrossRef]
- Su, E.; Xia, T.; Gao, L.; Dai, Q.; Zhang, Z. Immobilization of β-glucosidase and its aroma-increasing effect on tea beverage. Food Bioprod. Process. 2010, 88, 83–89. [Google Scholar] [CrossRef]
- Zhang, X.B.; Du, X.F. Effects of exogenous enzymatic treatment during processing on the sensory quality of summer tieguanyin oolong tea from the Chinese Anxi county. Food Technol. Biotechnol. 2015, 53, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Fang, K.; Ni, H.; Li, T.; Li, L.J.; Li, Q.B.; Chen, F. Aroma enhancement of instant green tea infusion using β-glucosidase and β-xylosidase. Food Chem. 2020, 315, 126287. [Google Scholar] [CrossRef] [PubMed]
- Dodor, D.E.; Tabatabai, M.A. Arylamidase activity as an index of nitrogen mineralization in soils. Commun. Soil Sci. Plant. Anal. 2007, 38, 2197–2207. [Google Scholar] [CrossRef]
- Pennacchia, C.; Blaiotta, G.; Pepe, O.; Villani, F. Isolation of Saccharomyces cerevisiae strains from different food matrices and their preliminary selection for a potential use as probiotics. J. Appl. Microbiol. 2008, 105, 1919–1928. [Google Scholar] [CrossRef]
- Mujdeci, G.N.; Ozbas, Z.Y. Technological and enzymatic characterization of the yeasts isolated from natural fermentation media of Gemlik olives. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef]
- Lee, S.B.; Park, H.D. Isolation and investigation of potential non-Saccharomyces yeasts to improve the volatile terpene compounds in Korean muscat bailey a wine. Microorganisms 2020, 8, 1552. [Google Scholar] [CrossRef]
- Fonseca, G.G.; Heinzle, E.; Wittmann, C.; Gombert, A.K. The yeast Kluyveromyces marxianus and its biotechnological potential. Appl. Microbiol. Biotechnol. 2008, 79, 339–354. [Google Scholar] [CrossRef]
- Borren, E.; Tian, B. The Important Contribution of Non-Saccharomyces Yeasts to the Aroma Complexity of Wine: A Review. Foods 2020, 10, 13. [Google Scholar] [CrossRef]
- Nurcholis, M.; Lertwattanasakul, N.; Rodrussamee, N.; Kosaka, T.; Murata, M.; Yamada, M. Integration of comprehensive data and biotechnological tools for industrial applications of Kluyveromyces marxianus. Appl. Microbiol. Biotechnol. 2020, 104, 475–488. [Google Scholar] [CrossRef]
- Rajkumar, A.S.; Morrissey, J.P. Rational engineering of Kluyveromyces marxianus to create a chassis for the production of aromatic products. Microb. Cell Fact. 2020, 19, 2–7. [Google Scholar] [CrossRef]
- Kregiel, D. Health safety of soft drinks: Contents, containers, and microorganisms. Biomed Res. Int. 2015. [Google Scholar] [CrossRef] [Green Version]
- Brattinga, C.; de Kok, P.M.T.; Bult, J.H.F. Sugar reduction in flavoured beverages: The robustness of aroma-induced sweetness enhancement. Proc. Flavour Sci. 2018, 199–206. [Google Scholar] [CrossRef]
- Askew, K. Sugar Reduction through Smell: ‘Aromas Can Be Used to Produce Long-Lasting Sweetness-Enhancing Effects’. Food Navigator. 2020. Available online: https://www.foodnavigator.com/Article/2020/01/21/Sugar-reduction-through-smell-Aromas-can-be-used-to-produce-long-lasting-sweetness-enhancing-effects (accessed on 5 May 2021).
- Pawlikowska, E.; Kolesińska, B.; Nowacka, M.; Kregiel, D. A New Approach to Producing High Yields of Pulcherrimin from Metschnikowia Yeasts. Fermentation 2020, 6, 114. [Google Scholar] [CrossRef]
- Sipiczki, M. Metschnikowia strains isolated from botrytized grapes antagonize fungal and bacterial growth by iron depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef] [Green Version]
- Sipiczki, M. Metschnikowia pulcherrima and related pulcherrimin-producing yeasts: Fuzzy species boundaries and complex antimicrobial antagonism. Microorganisms 2020, 8, 1029. [Google Scholar] [CrossRef]
- Kregiel, D. Physiology and metabolism of Crabtree-negative yeast Debaryomyces occidentalis. Food Chem. Biotechnol. 2008, 72, 35–44. [Google Scholar]
- Kregiel, D.; Berlowska, J.; Ambroziak, W. Growth and metabolic activity of conventional and non-conventional yeasts immobilized in foamed alginate. Enzym. Microb. Technol. 2013, 53, 229–234. [Google Scholar] [CrossRef]
- Berłowska, J.; Binczarski, M.; Dziugan, P.; Wilkowska, A.; Kręgiel, D.; Witońska, I. Sugar Beet Pulp as a Source of Valuable Biotechnological Products. Adv. Biotechnol. Food Ind. 2018, 14, 359–392. [Google Scholar] [CrossRef]
- Mroczek, A.; Kapusta, I.; Janda, B.; Janiszowska, W. Triterpene saponin content in the roots of red beet (Beta vulgaris L.) cultivars. J. Agric. Food Chem. 2012, 60, 12387–12402. [Google Scholar] [CrossRef]
- Prakash, B.; Kumar, A.; Singh, P.P.; Songachan, L.S. Antimicrobial and antioxidant properties of phytochemicals: Current status and future perspective. In Functional and Preservative Properties of Phytochemicals; Prakash, B., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 1–45. [Google Scholar] [CrossRef]
- Rahimi, P.; Abedimanesh, S.; Mesbah-Namin, S.A.; Ostadrahimi, A. Betalains, the nature-inspired pigments, in health and diseases. Crit. Rev. Food Sci. Nutr. 2019, 59, 2949–2978. [Google Scholar] [CrossRef]
- Panghal, A.; Virkar, K.; Kumar, V.; Dhull, S.B.; Gat, Y.; Chhikara, N. Development of probiotic beetroot drink. Curr. Res. Nutr. Food Sci. 2017, 5. [Google Scholar] [CrossRef]
- Herbach, K.M.; Stintzing, F.C.; Carle, R. Betalain Stability and Degradation—Structural and Chromatic Aspects. J. Food. Sci. 2006, 71, 41–50. [Google Scholar] [CrossRef]
- Mekoue Nguela, J.; Vernhet, A.; Sieczkowski, N.; Brillouet, J.M. Interactions of Condensed Tannins with Saccharomyces cerevisiae Yeast Cells and Cell Walls: Tannin Location by Microscopy. J. Agric. Food Chem. 2015, 63, 7539–7545. [Google Scholar] [CrossRef]
- Mekoue Nguela, J.; Sieczkowski, N.; Roi, S.; Vernhet, A. Sorption of grape proanthocyanidins and wine polyphenols by yeasts, inactivated yeasts, and yeast cell walls. J. Agric. Food Chem. 2015, 63, 660–670. [Google Scholar] [CrossRef]
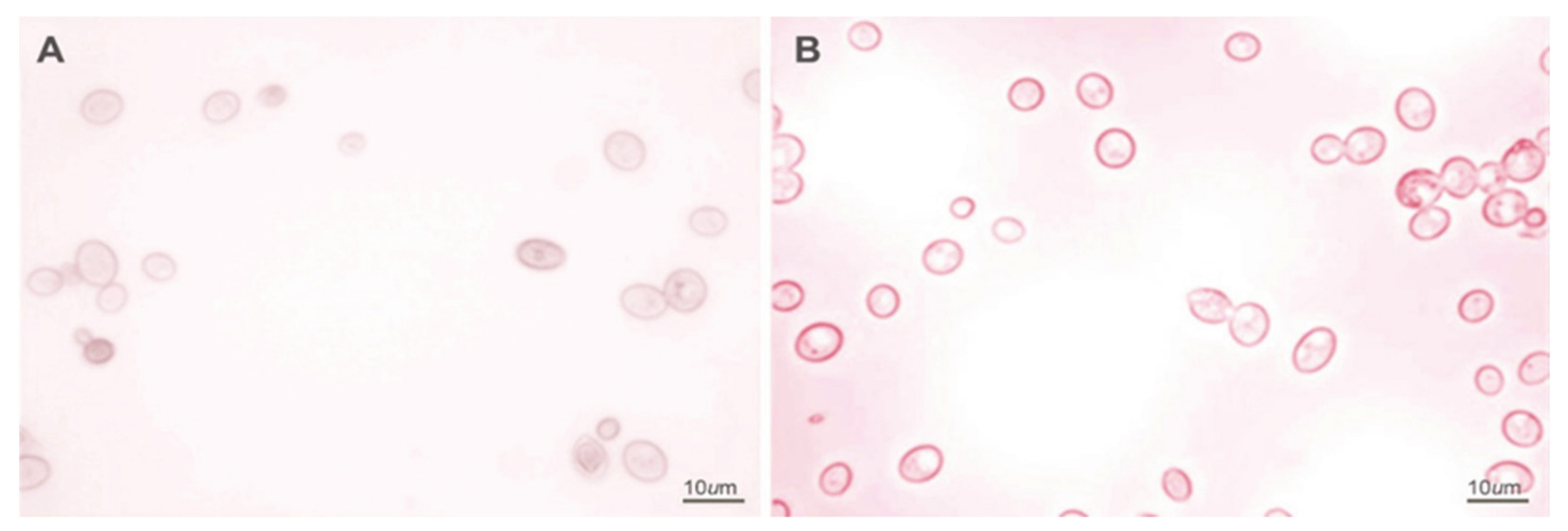


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Collection Number / Manufacturer | No. |
|---|---|---|
| Saccharomyces cerevisiae TT (brewery) | LOCK**0105 | I |
| Saccharomyces cerevisiae Tokay (winery) | LOCK0204 | II |
| Saccharomyces cerevisiae Ethanol Red*(distillery) | Leaf/Lessaffre* | III |
| Saccharomyces cerevisiae Lalvin*(winery) | ICV K1-V1116 Lallemand* | IV |
| Kluyveromyces marxianus | NCYC***179 | V |
| Kluyveromyces lactis | LOCK0028 | VI |
| Scheffersomycesstipitis | NCYC1541 | VII |
| Metschnikowia pulcherrima | NCYC747 | VIII |
| Carbohydrate [g/L] | |||||
|---|---|---|---|---|---|
| Saccharose | Fructose | Glucose | Rhamnose | Raffinose | Xylose |
| 86.2 ± 7.5 | 33.8 ± 0.2 | 17.6 ± 0.1 | 8.8 ± 0.6 | 4.3 ± 0.3 | 3.0 ± 0.2 |
| Strain | Glucose | Saccharose | Fructose | Arabinose | Rhamnose | Raffinose | Xylose |
|---|---|---|---|---|---|---|---|
| I* | +** | + | + | + | - | + | + |
| II | + | + | + | + | + | + | + |
| III | + | + | + | + | + | + | - |
| IV | + | + | + | + | + | + | - |
| V | + | + | + | + | + | - | - |
| VI | + | + | + | + | + | - | - |
| VII | + | + | + | + | + | + | + |
| VIII | + | + | + | - | + | - | - |
| Enzymes | Yeast Strain | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Classes | Specific Activity | I* | II | III | IV | V | VI | VII | VIII |
| Proteases | Leucine arylamidase | 5** | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| Valine arylamidase | 4 | 4 | 4 | 3 | 5 | 4 | 3 | 2 | |
| Cystine arylamidase | 0 | 4 | 4 | 3 | 4 | 4 | 3 | 1 | |
| Esterases | Esterase C4 | 3 | 4 | 2 | 3 | 5 | 3 | 4 | 3 |
| Esterase C8 | 3 | 4 | 2 | 3 | 4 | 4 | 3 | 3 | |
| Phosphatases | Alkaline phosphatase | 3 | 4 | 0 | 4 | 5 | 2 | 0 | 1 |
| Acid phosphatase | 5 | 5 | 5 | 5 | 5 | 5 | 3 | 2 | |
| Naphtol-AS-BI-phosphohydrolase | 4 | 4 | 3 | 5 | 5 | 3 | 3 | 4 | |
| α-Glucosidase | 3 | 3 | 2 | 0 | 0 | 5 | 2 | 5 | |
| 𝛽-Glucosidase | 0 | 0 | 3 | 0 | 4 | 5 | 0 | 3 | |
| Yeast Strain | Control | I* | II | III | IV | V | VI | VII | VIII |
|---|---|---|---|---|---|---|---|---|---|
| Extract Content [°Bx] | 15.01 ± 0.31 | 6.55 ± 0.23 | 6.18 ± 0.34 | 6.64 ± 0.53 | 6.60 ± 0.46 | 6.36 ± 0.43 | 6.58 ± 0.17 | 6.81 ± 0.46 | 7.54 ± 0.67 |
| Day of Completed Fermentation | 0 | 3 | 5 | 4 | 15 | 13 | 17 | 13 | 56 |
| Yeast Strain | Control | I* | II | III | IV | V | VI | VII | VIII |
|---|---|---|---|---|---|---|---|---|---|
| Sugar Index | 0.814 ± 0.023 | 0.062 ± 0.031 | 0.078 ± 0.022 | 0.072 ± 0.011 | 0.044 ± 0.023 | 0.090 ± 0.011 | 0.059 ± 0.009 | 0.041 ± 0.013 | 0.065 ± 0.012 |
| Yeast Strain | Control Sample | I* | II | III |
|---|---|---|---|---|
| Sugar Index | 0.814 ± 0.023 | 0.019 ± 0.009 | 0.033 ± 0.011 | 0.024 ± 0.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dygas, D.; Nowak, S.; Olszewska, J.; Szymańska, M.; Mroczyńska-Florczak, M.; Berłowska, J.; Dziugan, P.; Kręgiel, D. Ability of Yeast Metabolic Activity to Reduce Sugars and Stabilize Betalains in Red Beet Juice. Fermentation 2021, 7, 105. https://doi.org/10.3390/fermentation7030105
Dygas D, Nowak S, Olszewska J, Szymańska M, Mroczyńska-Florczak M, Berłowska J, Dziugan P, Kręgiel D. Ability of Yeast Metabolic Activity to Reduce Sugars and Stabilize Betalains in Red Beet Juice. Fermentation. 2021; 7(3):105. https://doi.org/10.3390/fermentation7030105
Chicago/Turabian StyleDygas, Dawid, Szymon Nowak, Joanna Olszewska, Monika Szymańska, Marta Mroczyńska-Florczak, Joanna Berłowska, Piotr Dziugan, and Dorota Kręgiel. 2021. "Ability of Yeast Metabolic Activity to Reduce Sugars and Stabilize Betalains in Red Beet Juice" Fermentation 7, no. 3: 105. https://doi.org/10.3390/fermentation7030105
APA StyleDygas, D., Nowak, S., Olszewska, J., Szymańska, M., Mroczyńska-Florczak, M., Berłowska, J., Dziugan, P., & Kręgiel, D. (2021). Ability of Yeast Metabolic Activity to Reduce Sugars and Stabilize Betalains in Red Beet Juice. Fermentation, 7(3), 105. https://doi.org/10.3390/fermentation7030105