Impact of Fermentation on Phenolic Compounds and Antioxidant Capacity of Quinoa



Abstract
1. Introduction
2. Fermentation and Phenolic Compounds in Grain-Based Foods
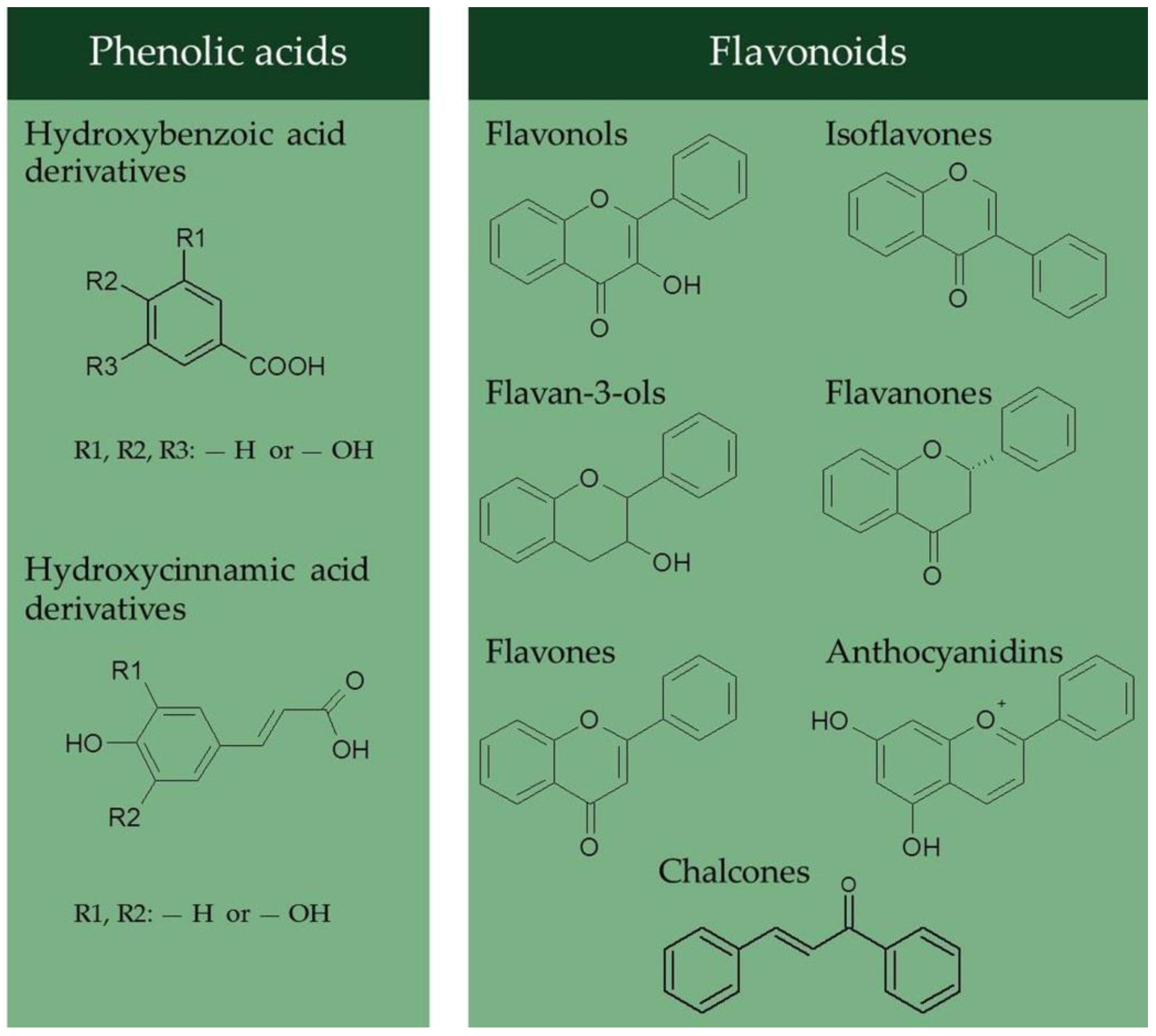
3. Phenolic Compounds in Quinoa
4. Impact of Fermentation on Phenolic Compounds in Quinoa
4.1. Fermentation and Phenolic Compound Content
4.2. Fermentation and Phenolic Compound Profile
5. Impact of Fermentation on Antioxidant Capacity in Quinoa-Based Foods
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Melini, F.; Melini, V.; Luziatelli, F.; Ruzzi, M. Current and Forward-Looking Approaches to Technological and Nutritional Improvements of Gluten-Free Bread with Legume Flours: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1101–1102. [Google Scholar] [CrossRef] [PubMed]
- Bazile, D.; Jacobsen, S.-E.; Verniau, A. The Global Expansion of Quinoa: Trends and Limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [PubMed]
- Adebo, O.A.; Medina-Meza, I.G. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef]
- Melini, F.; Melini, V.; Luziatelli, F.; Ficca, A.G.; Ruzzi, M. Health-promoting components in fermented foods: An up-to-date systematic review. Nutrients 2019, 11, 1189. [Google Scholar] [CrossRef] [PubMed]
- Melini, V.; Melini, F.; Acquistucci, R. Phenolic compounds and bioaccessibility thereof in functional pasta. Antioxidants 2020, 9, 343. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Terefe, N.S. Food Fermentation. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented foods: Definitions and characteristics, impact on the gut microbiota and effects on gastrointestinal health and disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef]
- Gan, R.Y.; Li, H.; Gunaratne, A.; Sui, Z.Q.; Corke, H. Effects of Fermented Edible Seeds and Their Products on Human Health: Bioactive Components and Bioactivities. Compr. Rev. Food Sci. Food Saf. 2017, 16, 489–531. [Google Scholar] [CrossRef]
- Paz, P.C.; Janny, R.J.; Håkansson, Å. Safeguarding of quinoa beverage production by fermentation with Lactobacillus plantarum DSM 9843. Int. J. Food Microbiol. 2020, 324, 108630. [Google Scholar]
- Koch, W. Dietary Polyphenols-Important Non-Nutrients in the Prevention of Chronic Noncommunicable Diseases. Syst. Rev. 2019, 11, 1039. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Varatharajan, V.; Oh, W.Y.; Peng, H. Phenolic compounds in agri-food by-products, their bioavailability and health effects. J. Food Bioact. 2019, 5, 57–119. [Google Scholar] [CrossRef]
- Melini, V.; Melini, F. Functional components and anti-nutritional factors in gluten-free grains: A focus on quinoa seeds. Foods 2021. in accepted. [Google Scholar]
- Mhada, M.; Metougui, M.L.; El Hazzam, K.; El Kacimi, K.; Yasri, A. Variations of Saponins, Minerals and Total Phenolic Compounds Due to Processing and Cooking of Quinoa (Chenopodium quinoa Willd.) Seeds. Foods 2020, 9, 660. [Google Scholar] [CrossRef] [PubMed]
- Saad-Allah, K.M.; Youssef, M.S. Phytochemical and genetic characterization of five quinoa (Chenopodium quinoa Willd.) genotypes introduced to Egypt. Physiol. Mol. Biol. Plants 2018, 24, 617–629. [Google Scholar] [CrossRef]
- Han, Y.; Chi, J.; Zhang, M.; Zhang, R.; Fan, S.; Dong, L.; Huang, F.; Liu, L. Changes in saponins, phenolics and antioxidant activity of quinoa (Chenopodium quinoa willd) during milling process. LWT 2019, 114, 108381. [Google Scholar]
- Lim, J.G.; Park, H.; Yoon, K.S. Analysis of saponin composition and comparison of the antioxidant activity of various parts of the quinoa plant (Chenopodium quinoa Willd.). Food Sci. Nutr. 2020, 8, 694–702. [Google Scholar] [CrossRef]
- Mattila, P.H.; Pihlava, J.M.; Hellström, J.; Nurmi, M.; Eurola, M.; Mäkinen, S.; Jalava, T.; Pihlanto, A. Contents of phytochemicals and antinutritional factors in commercial protein-rich plant products. Food Qual. Saf. 2018, 2, 213–219. [Google Scholar] [CrossRef]
- Multari, S.; Marsol-Vall, A.; Keskitalo, M.; Yang, B.; Suomela, J.P. Effects of different drying temperatures on the content of phenolic compounds and carotenoids in quinoa seeds (Chenopodium quinoa) from Finland. J. Food Compos. Anal. 2018, 72, 75–82. [Google Scholar] [CrossRef]
- Abderrahim, F.; Huanatico, E.; Segura, R.; Arribas, S.; Carmen Gonzalez, M.; Condezo-Hoyos, L. Physical features, phenolic compounds, betalains and total antioxidant capacity of coloured quinoa seeds (Chenopodium quinoa Willd.) from Peruvian Altiplano. Food Chem. 2015, 183, 83–90. [Google Scholar] [CrossRef]
- Vega-Gálvez, A.; Zura, L.; Lutz, M.; Jagus, R.; Victoria Agüero, M.; Pastén, A.; Di Scala, K.; Uribe, E. Assessment of dietary fiber, isoflavones and phenolic compounds with antioxidant and antimicrobial properties of quinoa (Chenopodium quinoa Willd.). Chil. J. Agric. Anim. Sci. 2018, 34, 1–11. [Google Scholar] [CrossRef]
- Nickel, J.; Spanier, L.P.; Botelho, F.T.; Gularte, M.A.; Helbig, E. Effect of different types of processing on the total phenolic compound content, antioxidant capacity, and saponin content of Chenopodium quinoa Willd grains. Food Chem. 2016, 209, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Melini, V.; Panfili, G.; Fratianni, A.; Acquistucci, R. Bioactive compounds in rice on Italian market: Pigmented varieties as a source of carotenoids, total phenolic compounds and anthocyanins, before and after cooking. Food Chem. 2019, 277, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, C.; Ji, Y.; Lin, J.; Chen, X.; Qi, B. Improvement of nutritional value, bioactivity and volatile constituents of quinoa seeds by fermentation with Lactobacillus casei. J. Cereal Sci. 2018, 84, 83–89. [Google Scholar] [CrossRef]
- Rocchetti, G.; Miragoli, F.; Zacconi, C.; Lucini, L.; Rebecchi, A. Impact of cooking and fermentation by lactic acid bacteria on phenolic profile of quinoa and buckwheat seeds. Food Res. Int. 2019, 119, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.N.; Guo, S.; Zhang, S.W. Effects of solid-state fermentation on the nutritional components and antioxidant properties from quinoa. Emir. J. Food Agric. 2019, 31, 39–45. [Google Scholar] [CrossRef]
- Hur, J.; Nguyen, T.T.H.; Park, N.; Kim, J.; Kim, D. Characterization of quinoa (Chenopodium quinoa) fermented by Rhizopus oligosporus and its bioactive properties. AMB Express 2018, 8, 143. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Stodolak, B.; Gómez-Caravaca, A.M.; Mickowska, B.; Martin-Garcia, B.; Byczyński, Ł. Mould starter selection for extended solid-state fermentation of quinoa. LWT 2019, 99, 231–237. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Duliński, R.; Stodolak, B.; Mickowska, B.; Wikiera, A. Prolonged tempe-type fermentation in order to improve bioactive potential and nutritional parameters of quinoa seeds. J. Cereal Sci. 2016, 71, 116–121. [Google Scholar] [CrossRef]
- Carciochi, R.A.; Galván-D’Alessandro, L.; Vandendriessche, P.; Chollet, S. Effect of Germination and Fermentation Process on the Antioxidant Compounds of Quinoa Seeds. Plant Foods Hum. Nutr. 2016, 71, 361–367. [Google Scholar] [CrossRef]
- Ayyash, M.; Johnson, S.K.; Liu, S.Q.; Mesmari, N.; Dahmani, S.; Al Dhaheri, A.S.; Kizhakkayil, J. In vitro investigation of bioactivities of solid-state fermented lupin, quinoa and wheat using Lactobacillus spp. Food Chem. 2019, 275, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, A.; Verni, M.; Montemurro, M.; Coda, R.; Gobbetti, M.; Rizzello, C.G. Use of fermented quinoa flour for pasta making and evaluation of the technological and nutritional features. LWT Food Sci. Technol. 2017, 78, 215–221. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Lorusso, A.; Montemurro, M.; Gobbetti, M. Use of sourdough made with quinoa (Chenopodium quinoa) flour and autochthonous selected lactic acid bacteria for enhancing the nutritional, textural and sensory features of white bread. Food Microbiol. 2016, 56, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Montemurro, M.; Pontonio, E.; Gobbetti, M.; Rizzello, C.G. Investigation of the nutritional, functional and technological effects of the sourdough fermentation of sprouted flours. Int. J. Food Microbiol. 2019, 302, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Chiş, M.S.; Păucean, A.; Man, S.M.; Vodnar, D.C.; Teleky, B.-E.; Pop, C.R.; Stan, L.; Borsai, O.; Kadar, C.B.; Urcan, A.C.; et al. Quinoa Sourdough Fermented with Lactobacillus plantarum ATCC 8014 Designed for Gluten-Free Muffins—A Powerful Tool to Enhance Bioactive Compounds. Appl. Sci. 2020, 10, 7140. [Google Scholar] [CrossRef]
- Soltani, M.; Hekmat, S.; Ahmadi, L. Microbial and sensory evaluation of probiotic yoghurt supplemented with cereal/pseudo-cereal grains and legumes. Int. J. Dairy Technol. 2018, 71, 141–148. [Google Scholar] [CrossRef]
- Lorusso, A.; Coda, R.; Montemurro, M.; Rizzello, C. Use of Selected Lactic Acid Bacteria and Quinoa Flour for Manufacturing Novel Yogurt-Like Beverages. Foods 2018, 7, 51. [Google Scholar] [CrossRef]
- Karovičová, J.; Kohajdová, Z.; Minarovičová, L.; Lauková, M.; Greifová, M.; Greif, G.; Hojerová, J. Utilisation of quinoa for development of fermented beverages. Potravin. Slovak J. Food Sci. 2020, 14, 465–472. [Google Scholar] [CrossRef]
- Shanker, K.S.; Kishore, K.H.; Kanjilal, S.; Misra, S.; Murty, U.S.N.; Prasad, R.B.N. Biotransformation of ferulic acid to acetovanillone using Rhizopus oryzae. Biocatal. Biotransform. 2007, 25, 109–112. [Google Scholar] [CrossRef]


{kind=link}
| Raw Material | Fermented Quinoa-Based Food | Microorganism(s) | Phenolic Content in Raw Quinoa/Quinoa-Based Food | Phenolic Content in Fermented Food | Reference |
|---|---|---|---|---|---|
| Quinoa seeds | Fermented quinoa seeds | Lactobacillus casei CICC 20995 | TPC (free): 205 1 TPC (bound): 1180 1 | TPC (free): 910 1 TPC (bound): 743 1 | Li et al. [25] |
| Quinoa cooked and acidified seeds | Fermented quinoa seeds | Lactobacillus paracasei A1 2.6 strain and/or Pediococcus pentosaceus GS·B strain | TPC (raw): 65.7 1 TPC (cooked): 70.3 1 TPC (cooked+acidified): 60.8 1 | TPC (L. paracasei): 59.6 1 TPC (P. pentosaceus): 70.9 1 TPC (L. paracasei + P. pentosaceus): 74.8 1 | Rocchetti et al. [26] |
| Quinoa seeds | Fermented quinoa seeds | Agaricus bisporus AS2796 | TPC: ≈ 70 1 | TPC (7th day): ≈ 73 1 TPC (14th day): ≈ 81 1 TPC (21th day): ≈ 110 1 TPC (28th day): ≈ 135 1 TPC (35th day): ≈ 138 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa seeds | Fomitiporia yanbeiensis G1 | TPC: ≈ 70 1 | TPC (7th day): ≈ 68 1 TPC (14th day): ≈ 79 1 TPC (21th day): ≈ 110 1 TPC (28th day): ≈ 138 1 TPC (35th day): ≈ 130 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa seeds | Helvella lacunosa X1 | TPC: ≈ 70 1 | TPC (7th day): ≈ 69 1 TPC (14th day): ≈ 79 1 TPC (21th day): ≈ 110 1 TPC (28th day): ≈ 138 1 TPC (35th day): ≈130 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa seeds | Rhizopus microspores var. oligosporus | TPC: 4.1 1 TFC: 1.3 5 | TPC (3 days): 7.4 1 TPC (5 days): 8.0 1 TFC (3 days): 1.6 5 TFC (5 days): 1.9 5 | Hur et al. [28] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Aspergillus oryzae | TP (black, raw): 0.47 2 TP (black, pre-cooked): 0.31 2 TP (red, raw): 0.47 2 TP (red, pre-cooked): 0.31 2 | TP (black, 2 days): 0.53 2 TP (black, 4 days): 0.52 2 TP (black, 6 days): 0.56 2 TP (red, 2 days): 0.47 2 TP (red, 4 days): 0.52 2 TP (red, 6 days): 0.60 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Neurospora intermedia | TP (black, raw): 0.47 2 TP (black, pre-cooked): 0.31 2 TP (red, raw): 0.47 2 TP (red, pre-cooked): 0.31 2 | TP (black, 3 days): 0.54 2 TP (black, 5 days): 0.54 2 TP (red, 3 days): 0.51 2 TP (red, 5 days): 0.53 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Rhizopus oligosporus | TP (black, raw): 0.47 2 TP (black, pre-cooked): 0.31 2 TP (red, raw): 0.47 2 TP (red, pre-cooked): 0.31 2 | TP (black, 2 days): 0.77 2 TP (black, 4 days): 1.82 2 TP (black, 6 days): 1.96 2 TP (red, 2 days): 0.50 2 TP (red, 4 days): 0.84 2 TP (red, 6 days): 0.89 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds (white, red, black varieties) | Fermented quinoa seeds | R. oligosporus ATCC 64063 strain | TPC (white, cooked): 2.76 1 TPC (red, cooked): 2.73 1 TPC (black, cooked): 3.28 1 | TPC (white, fermented 30 h): 8.71 1 TPC (white, fermented 40 h): 9.12 1 TPC (red, fermented 30 h): 7.22 1 TPC (red, fermented 40 h): 7.08 1 TPC (black, fermented 30 h): 7.65 1 TPC (black, fermented 40 h): 8.46 1 | Starzyńska-Janiszewska et al. [30] |
| Quinoa seeds | Fermented quinoa seeds | Saccharomyces cerevisiae NBRC 2375 and NBRC 1951 | TPC: 39.3 1 | TPC (NBRC 2375): ≈ 57 1 TPC (NBRC 1951): ≈ 55 1 | Carciochi et al. [31] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | Lactobacillus plantarum DSM2648 | TPC (0 h): ≈ 0.13 1 TPC (24 h): ≈ 0.11 1 TPC (48 h): ≈ 0.14 1 TPC (72 h): ≈ 0.18 1 | TPC (0 h): ≈ 0.16 1 TPC (24 h): ≈ 0.36 1 TPC (48 h): ≈ 0.34 1 TPC (72 h): ≈ 0.39 1 | Ayyash et al. [32] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | L. plantarum KX881779 | TPC (0 h): ≈ 0.13 1 TPC (24 h): ≈ 0.11 1 TPC (48 h): ≈ 0.14 1 TPC (72 h): ≈ 0.18 1 | TPC (0 h): ≈ 0.10 1 TPC (24 h): ≈ 0.20 1 TPC (48 h): ≈ 0.28 1 TPC (72 h): ≈ 0.37 1 | Ayyash et al. [32] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | Lactobacillus reuteri KX88177 | TPC (0 h): ≈ 0.13 1 TPC (24 h): ≈ 0.11 1 TPC (48 h): ≈ 0.14 1 TPC (72 h): ≈ 0.18 1 | TPC (0 h): ≈ 0.11 1 TPC (24 h): ≈ 0.17 1 TPC (48 h): ≈ 0.36 1 TPC (72 h): ≈ 0.45 1 | Ayyash et al. [32] |
| Quinoa flour | Fermented quinoa pasta | L. plantarum T6B10 Lactobacillus rossiae T0A16 | TPC (semolina pasta): 2.21 3 TPC (quinoa pasta): 3.02 3 | TPC (fermented quinoa pasta): 4.06 3 | Lorusso et al. [33] |
| Quinoa flour | Quinoa sourdough | L. plantarum T6B10 L. rossiae T0A16 | TPC: 3.65 3 | TPC: 8.78 3 | Rizzello et al. [34] |
| Quinoa raw flour | Quinoa sourdough | L. rossiae LB5 L. plantarum 1A7 Lactobacillus sanfranciscensis DE9 | TPC: 3.28 3 | TPC: 5.79 3 | Montemurro et al. [35] |
| Quinoa sprouted flour | Quinoa sourdough | L. rossiae LB5 L. plantarum 1A7 L. sanfranciscensis DE9 | TPC: 5.44 3 | TPC: 6.63 3 | Montemurro et al. [35] |
| Quinoa wholemeal flour | Quinoa sourdough | L. plantarum ATCC 8014 | TFC: 997 4 | TFC (sourdough, 24 h fermentation): 757 4 TFC (muffin): 1317 4 | Chiş et al. [36] |
| Quinoa flour | Yogurt-like beverages | Lactobacillus rhamnosus SP1 L. plantarum T6B10 Weissella confusa DSM 20194 | TPC (B-SP1): 5.3 3 TPC (B-T6B10): 5.2 3 TPC (B-DSM 20194): 4.0 3 | TPC (B-SP1): 5.8 3 TPC (B-T6B10): 8.4 3 TPC (B-DSM 20194): 5.9 3 TPC (B-SP1, 20-day storage): 9.6 3 TPC (B-T6B10, 20-day storage): 9.3 3 TPC (B-DSM 20194, 20-day storage): 7.9 3 | Lorusso et al. [37] |
| Quinoa flour | Fermented quinoa beverages | Commercial probiotic culture including Bifidobacterium sp., Lactobacillus acidophilus, and Streptococcus thermophilus | TPC: 142.37 1 | TPC: 180.33 1 | Karovičová et al. [38] |
| Phenolic Compound | Food Matrix | Content in Raw Quinoa | Content in Fermented Quinoa | Microorganism(s) | Reference |
|---|---|---|---|---|---|
| Hydroxybenzoic acid derivatives | |||||
| Gallic acid | Quinoa seeds | 0.01 mg kg−1 | 2.37 mg kg−1 (3-day fermentation) 0.84 mg kg−1 (5-day fermentation) | Rhizopus microspores var. oligosporus | Hur et al. [28] |
| Hydroxybenzoic acid (bound form) | Red and black quinoa seeds | 1.890 mg 100 g−1 (black) 5.530 mg 100 g−1 (red) | 1.680 mg 100 g−1 (black) 1.775 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| p-Hydroxybenzoic acid | Quinoa seeds | 100% | 831% area relative to raw grain | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| p-Hydroxybenzoic acid | Quinoa seeds | 100% | 868% area relative to raw grain | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Protocatechuic acid | White quinoa seeds | 0.60 µg g−1 dm (cooked) | 1.94 µg g−1 dm (30 h) 3.69 µg g−1 dm (40 h) | Rhizopus oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Protocatechuic acid | Red quinoa seeds | 16.09 µg g−1 dm (cooked) | 28.40 µg g−1 dm (30 h) 29.19 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Protocatechuic acid | Black quinoa seeds | 20.4 µg g−1 dm (cooked) | 28.48 µg g−1 dm (30 h) 34.45 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Vanillic acid (free form) | Black and red quinoa seeds | 0.222 mg 100 g−1 (black) 0.762 mg 100 g−1 (red) | 0.482 mg 100 g−1 (black) 0.690 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Vanillic acid (bound form) | Black and red quinoa seeds | 0.990 mg 100 g−1 (black) 0.195 mg 100 g−1 (red) | 1.125 mg 100 g−1 (black) 1.215 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Vanillic acid | White quinoa seeds | 22.02 µg g−1 dm (cooked) | 42.05 µg g−1 dm (30 h) 44.68 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Vanillic acid | Red quinoa seeds | 10.07 µg g−1 dm (cooked) | 43.60 µg g−1 dm (30 h) 46.21 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Vanillic acid | Black quinoa seeds | 12.08 µg g−1 dm (cooked) | 23.10 µg g−1 dm (30 h) 20.86 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Vanillic acid | Quinoa seeds | 1.3 mg kg−1 | 1.55 mg kg−1 (3-day fermentation) 1.83 mg kg−1 (5-day fermentation) | Rhizopus microspores var. oligosporus | Hur et al. [28] |
| Vanillic acid | Quinoa seeds | 100% | 142% area relative to raw grain | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| Vanillic acid | Quinoa seeds | 100% | 343% area relative to raw grain | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Hydroxycinnamic acid derivatives | |||||
| Caffeic acid (bound form) | Black and red quinoa seeds | 0.105 mg 100 g−1 (black) 0.030 mg 100 g−1 (red) | 0.160 mg 100 g−1 (black) 0.034 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Chlorogenic acid | Quinoa seeds | 0.002 mg kg−1 | 0.03 mg kg−1 (3-day fermentation) 0.002 mg kg−1 (5-day fermentation) | Rhizopus microspores var. oligosporus | Hur et al. [28] |
| Ferulic acid (free form) | Black and red quinoa seeds | 0.650 mg 100 g−1 (black) 0.888 mg 100 g−1 (red) | 0.230 mg 100 g−1 (black) 0.273 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Ferulic acid (bound form) | Black and red quinoa seeds | 0.16 mg 100 g−1 (black) 0.125 mg 100 g−1 (red) | 0.13 mg 100 g−1 (black) 0.082 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Ferulic acid | Quinoa seeds | 100% | 235% area relative to raw grain | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| Ferulic acid | Quinoa seeds | 100% | 61% area relative to raw grain | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Hydroxycinnamic acid (free form) | Black and red quinoa seeds | 12.48 mg 100 g−1 (black) 3.620 mg 100 g−1 (red) | 1.282 mg 100 g−1 (black) 0.670 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| p-Coumaric acid (bound form) | Black and red quinoa seeds | 1.015 mg 100 g−1 (black) 0.025 mg 100 g−1 (red) | 0.720 mg 100 g−1 (black) 0.014 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| p-Coumaric acid | Quinoa seeds | 100% | 1002% area relative to raw grain | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| p-Coumaric acid | Quinoa seeds | 100% | 813% area relative to raw grain | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Flavonoids | |||||
| Catechin (bound form) | Black and red quinoa seeds | 0.205 mg 100 g−1 (black) 0.055 mg 100 g−1 (red) | 0.165 mg 100 g−1 (black) 0.130 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Kaempferol (free form) | Black and red quinoa seeds | 0.061 mg 100 g−1 (black) 0.047 mg 100 g−1 (red) | n.q. (black) 0.102 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Kaempferol (bound form) | Black and red quinoa seeds | n.q. (black) n.q. (red) | n.q. (black) n.q. (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Kaempferol | Quinoa seeds | 100% | n.d. | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| Kaempferol | Quinoa seeds | 100% | n.d. | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Quercetin (free form) | Black and red quinoa seeds | 0.082 mg 100 g−1 (black) 0.633 mg 100 g−1 (red) | n.q. (black) 0.187 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Quercetin (bound form) | Black and red quinoa seeds | 0.022 mg 100 g−1 (black) 0.043 mg 100 g−1 (red) | 0.061 mg 100 g−1 (black) 0.080 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Quercetin | Quinoa seeds | 100% | n.d. | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| Quercetin | Quinoa seeds | 100% | n.d. | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Rutin (free form) | Black and red quinoa seeds | 0.326 mg 100 g−1 (black) 0.580 mg 100 g−1 (red) | 0.054 mg 100 g−1 (black) 0.309 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Rutin | White quinoa seeds | 84.34 µg g−1 dm (cooked) | 107.09 µg g−1 dm (30 h) 104.68 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Rutin | Red quinoa seeds | 57.70 µg g−1 dm (cooked) | 235.11 µg g−1 dm (30 h) 229.98 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Rutin | Black quinoa seeds | 39.53 µg g−1 dm (cooked) | 56.37 µg g−1 dm (30 h) 55.52 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Raw Material | Fermented Quinoa-Based Food | Microorganism(s) | Antioxidant Capacity in Raw Quinoa/Quinoa-Based Food | Antioxidant Capacity in Fermented Food | Reference |
|---|---|---|---|---|---|
| Quinoa seeds | Fermented quinoa seeds | Lactobacillus casei CICC 20995 | IC50: 6.65 5 | IC50: 3.43 5 | Li et al. [25] |
| Quinoa cooked and acidified seeds | Quinoa fermented seeds | Lactobacillus paracasei A1 2.6 strain and/or Pediococcus pentosaceus GS·B strain | FRAP (raw): 15.4 4 FRAP (cooked): n.d. FRAP (acidified): n.d. ORAC (raw): 45.84 2 ORAC (cooked): 79.45 2 ORAC (acidified): 66.88 2 | FRAP (L. paracasei): n.d. FRAP (P. pentosaceus): n.d. FRAP (L. paracasei + P. pentosaceus): n.d. ORAC (L. paracasei): 59.68 2 ORAC (P. pentosaceus): 70.11 2 ORAC (L. paracasei + P. pentosaceus): 68.55 2 | Rocchetti et al. [26] |
| Quinoa seeds | Fermented quinoa | Helvella lacunosa X1 | DPPH ≈ 47 1 | DPPH (7th day): ≈ 45 1 DPPH (14th day): ≈ 51 1 DPPH (21th day): ≈ 23 1 DPPH (28th day): ≈ 9 1 DPPH (35th day): ≈ 8 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa | Agaricus bisporus AS2796 | DPPH ≈ 47 1 | DPPH (7th day): ≈ 45 1 DPPH (14th day): ≈ 53 1 DPPH (21th day): ≈ 22 1 DPPH (28th day): ≈ 9 1 DPPH (35th day): ≈ 8 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa | Fomitiporia yanbeiensis G1 | DPPH ≈ 47 1 | DPPH (7th day): ≈ 45 1 DPPH (14th day): ≈ 53 1 DPPH (21th day): ≈ 22 1 DPPH (28th day): ≈ 9 1 DPPH (35th day): ≈ 8 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa seeds | Rhizopus microspores var. oligosporus | DPPH: 3.6 5 | DPPH (3 days): 3.4 5 DPPH (5 days): 2.3 5 | Hur et al. [28] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Rhizopus oligosporus | ABTS (black): 49.20 2 ABTS (red): 42.22 2 | ABTS (black, 2 days): 84.38 2 ABTS (black, 4 days): 185.06 2 ABTS (black, 6 days): 227.71 2 ABTS (red, 2 days): 55.68 2 ABTS (red, 4 days): 82.37 2 ABTS (red, 6 days): 105.01 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Aspergillus oryzae | ABTS (black): 49.20 2 ABTS (red): 42.22 2 | ABTS (black, 2 days): 50.13 2 ABTS (black, 4 days): 47.91 2 ABTS (black, 6 days): 48.95 2 ABTS (red, 2 days): 42.40 2 ABTS (red, 4 days): 49.00 2 ABTS (red, 6 days): 53.13 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Neurospora intermedia | ABTS (black): 49.20 2 ABTS (red): 42.22 2 | ABTS (black, 3 days): 53.70 2 ABTS (black, 5 days):56.15 2 ABTS (red, 3 days): 52.23 2 ABTS (red, 5 days): 54.93 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds | Fermented quinoa seeds | S. cerevisiae NBRC 2375 S. cerevisiae NBRC 1951 | DPPH: 2.60 2 ABTS: 3.50 2 FRAP: 1.20 2 | DPPH (NBRC 2375): ≈ 3.70 2 ABTS (NBRC 2375): ≈ 4.40 2 FRAP (NBRC 2375): ≈ 1.90 2 DPPH (NBRC 1951): ≈ 3.50 2 ABTS (NBRC 1951): ≈ 4.50 2 FRAP (NBRC 1951): ≈ 185 2 | Carciochi et al. [31] |
| Quinoa flour | Fermented quinoa pasta | L. plantarum T6B10 L. rossiae T0A16 | DPPH (semolina pasta): 14 3 DPPH (quinoa pasta): 26 3 | DPPH: 35 3 | Lorusso et al. [33] |
| Quinoa flour | Quinoa sourdough | L. plantarum T6B10 L. rossiae T0A16 | DPPH: 41.7 3 | DPPH: 71.8 3 | Rizzello et al. [34] |
| Quinoa raw flour | Quinoa sourdough | L. rossiae LB5 L. plantarum 1A7 L. sanfranciscensis DE9 | DPPH: 81 3 | DPPH: 90 3 | Montemurro et al. [35] |
| Quinoa sprouted flour | Quinoa sourdough | L. rossiae LB5 L. plantarum 1A7 L. sanfranciscensis DE9 | DPPH: 94 3 | DPPH: 97 3 | Montemurro et al. [35] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | L. plantarum DSM2648 | DPPH (0 h): ≈ 29 3 DPPH (24 h): ≈ 28 3 DPPH (48 h): ≈ 30 3 DPPH (72 h): ≈ 27 3 | DPPH (0 h): ≈ 27 3 DPPH (24 h): ≈ 68 3 DPPH (48 h): ≈ 66 3 DPPH (72 h): ≈ 64 3 | Ayyash et al. [32] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | L. reuteri KX88177 | DPPH (0 h): ≈ 29 3 DPPH (24 h): ≈ 28 3 DPPH (48 h): ≈ 30 3 DPPH (72 h): ≈ 27 3 | DPPH (0 h): ≈ 32 3 DPPH (24 h): ≈ 75 3 DPPH (48 h): ≈ 58 3 DPPH (72 h): ≈ 73 3 | Ayyash et al. [32] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | L. plantarum KX881779 | DPPH (0 h): ≈ 29 3 DPPH (24 h): ≈ 28 3 DPPH (48 h): ≈ 30 3 DPPH (72 h): ≈ 27 3 | DPPH (0 h): ≈ 28 3 DPPH (24 h): ≈ 69 3 DPPH (48 h): ≈ 68 3 DPPH (72 h): ≈ 63 3 | Ayyash et al. [32] |
| Quinoa flour | Yogurt-like beverages | L. rhamnosus SP1 L. plantarum T6B10 W. confusa DSM 20194 | DPPH (B-SP1): 25 3 DPPH (B-T6B10):24 3 DPPH (B-DMS 20194): 29 3 | DPPH (B-SP1, after fermentation): 32 3 DPPH (B-T6B10, after fermentation): 37 3 DPPH (B-DMS 20194, after fermentation): 32 3 DPPH (B-SP1, after storage): 49 3 DPPH (B-T6B10, after storage): 44 3 DPPH (B-DMS 20194, after storage): 38 3 | Lorusso et al. [37] |
| Quinoa flour | Fermented quinoa-based beverages | Commercial probiotic culture including Bifidobacterium sp., Lactobacillus acidophilus, and Streptococcus thermophilus | DPPH: 60.31 3 | DPPH: 69.20 3 | Karovičová et al. [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melini, F.; Melini, V. Impact of Fermentation on Phenolic Compounds and Antioxidant Capacity of Quinoa. Fermentation 2021, 7, 20. https://doi.org/10.3390/fermentation7010020
Melini F, Melini V. Impact of Fermentation on Phenolic Compounds and Antioxidant Capacity of Quinoa. Fermentation. 2021; 7(1):20. https://doi.org/10.3390/fermentation7010020
Chicago/Turabian StyleMelini, Francesca, and Valentina Melini. 2021. "Impact of Fermentation on Phenolic Compounds and Antioxidant Capacity of Quinoa" Fermentation 7, no. 1: 20. https://doi.org/10.3390/fermentation7010020
APA StyleMelini, F., & Melini, V. (2021). Impact of Fermentation on Phenolic Compounds and Antioxidant Capacity of Quinoa. Fermentation, 7(1), 20. https://doi.org/10.3390/fermentation7010020