Novel Non-Cerevisiae Saccharomyces Yeast Species Used in Beer and Alcoholic Beverage Fermentations



Abstract
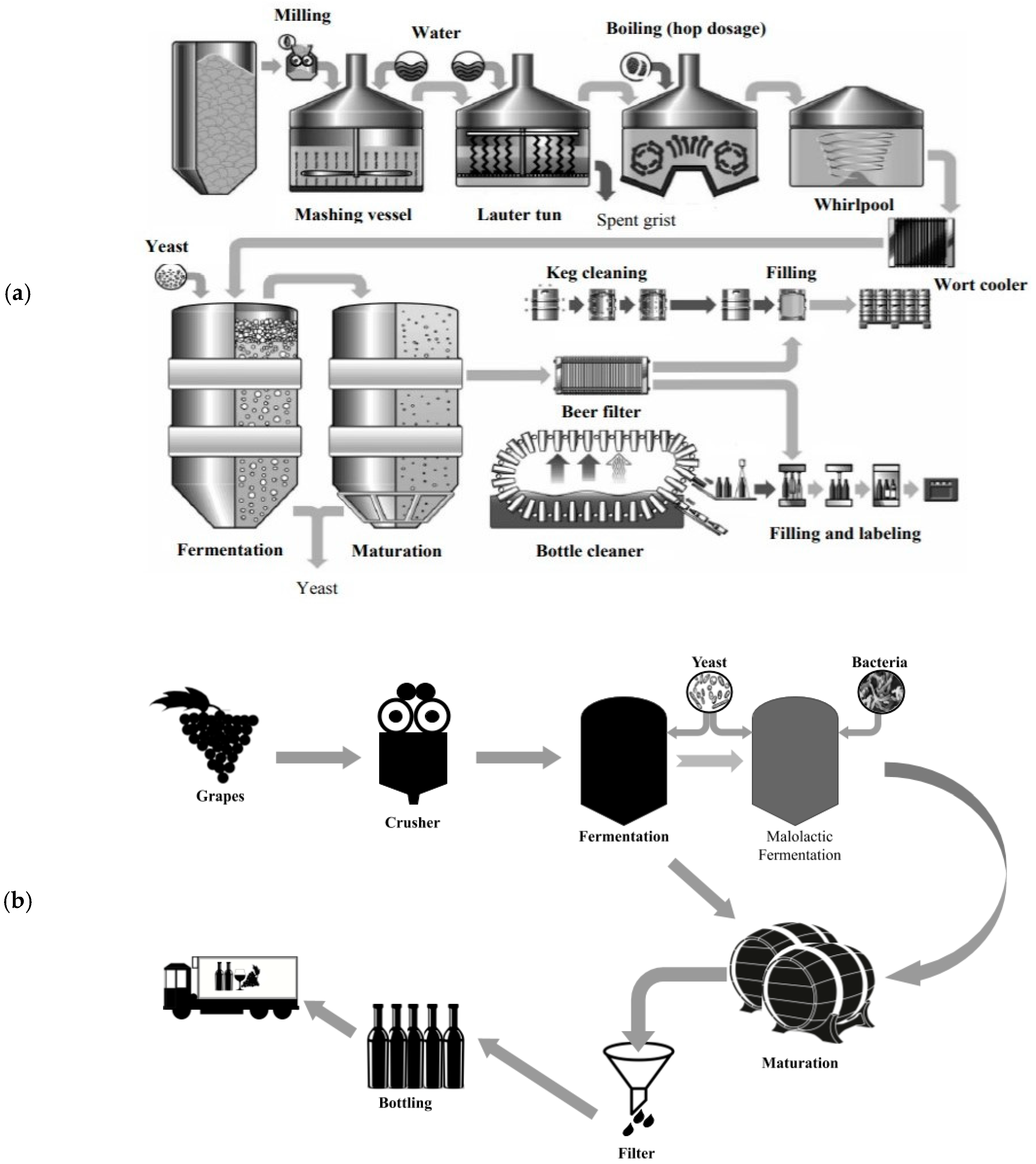
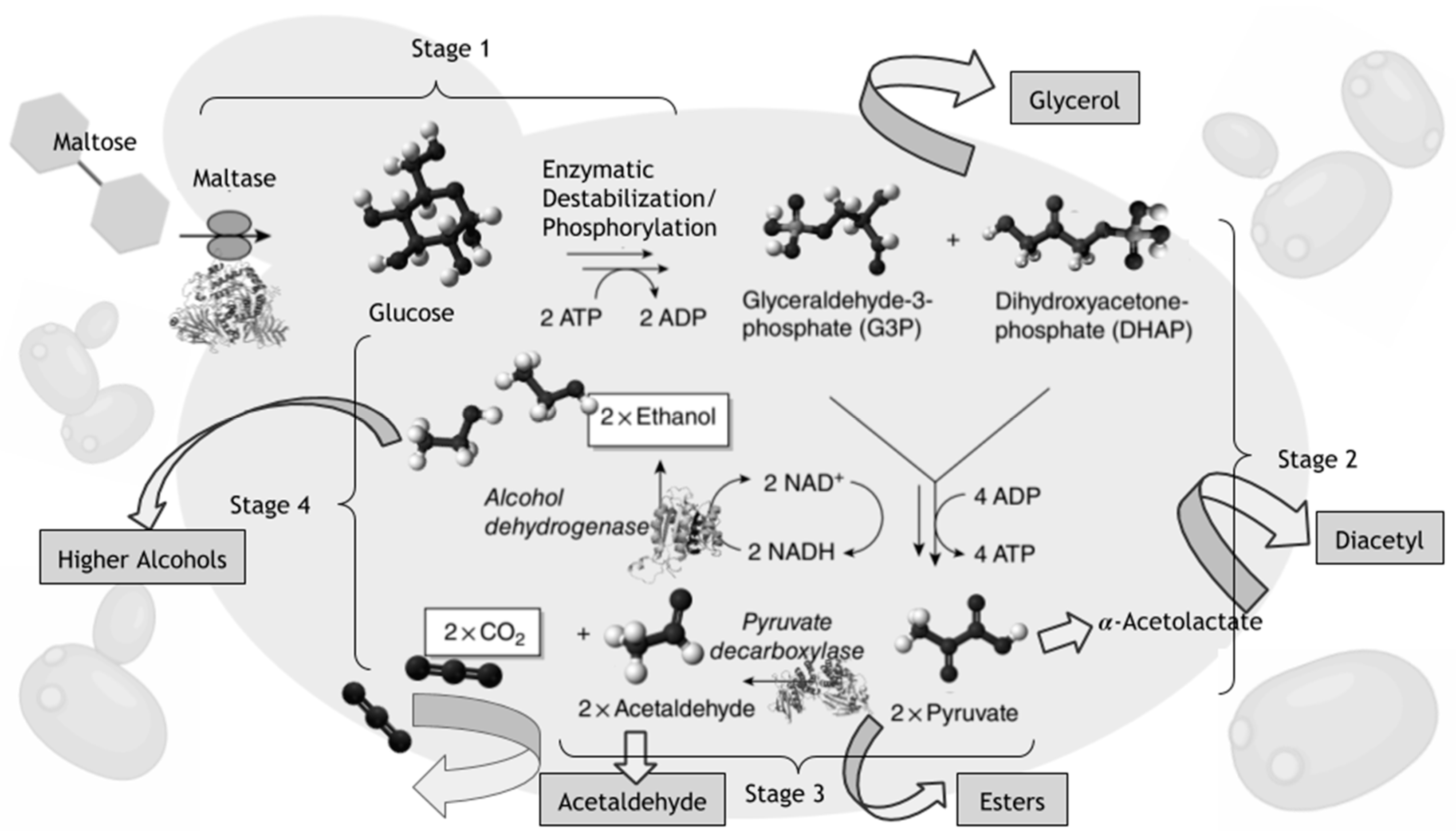
1. Introduction
2. Saccharomyces Species Diversity
3. Saccharomyces kudriavzevii
4. Saccharomyces paradoxus
5. Saccharomyces mikatae
6. Saccharomyces uvarum
7. Saccharomyces bayanus
8. Other Saccharomyces spp. Used in Alcoholic Fermentation
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Legras, J.-L.; Merdinoglu, D.; Cornuet, J.-M.; Karst, F. Bread, beer and wine: Saccharomyces cerevisiae diversity reflects human history. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef]
- Meussdoerffer, F.G. A comprehensive history of beer brewing. In Handbook of Brewing; Elinger, H.M., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2009; pp. 1–42. ISBN 9783527316748. [Google Scholar]
- Perruchini, E.; Glatz, C.; Hald, M.M.; Casana, J.; Toney, J.L. Revealing invisible brews: A new approach to the chemical identification of ancient beer. J. Archaeol. Sci. 2018, 100, 176–190. [Google Scholar] [CrossRef]
- Liu, L.; Wang, J.; Rosenberg, D.; Zhao, H.; Lengyel, G.; Nadel, D. Fermented beverage and food storage in 13,000 y-old stone mortars at Raqefet Cave, Israel: Investigating Natufian ritual feasting. J. Archaeol. Sci. Rep. 2018, 21, 783–793. [Google Scholar] [CrossRef]
- Bamforth, C.W. The Horace Brown Medal. Forever in focus: Researches in malting and brewing sciences. J. Inst. Brew. 2020. [Google Scholar] [CrossRef]
- Balling, K. Die Sacharometrische Bierprobe; Borrosch & André: Prague, Czechoslavakia, 1842. [Google Scholar]
- Joule, J.P. On the calorific effects of magneto-electricity, and on the mechanical value of heat. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1843, 23, 435–443. [Google Scholar] [CrossRef]
- Pasteur, L. Études sur le vin: Ses Maladies, Causes qui les Provoquent, Procédés Nouveaux Pour le Conserver et Pour le Vieillir, 2nd ed.; Imprimerie Impériale: Paris, France, 1866. [Google Scholar]
- Bamforth, C.W. Scientific Principles of Malting and Brewing; American Society of Brewing Chemists, Ed.; American Society of Brewing Chemists: St. Paul, MN, USA, 2006; pp. 1–246. ISBN 1-881696-08-1. [Google Scholar]
- Ziliak, S.T. Retrospectives: Guinnessometrics: The economic foundation of “student’s”t. J. Econ. Perspect. 2008, 22, 199–216. [Google Scholar] [CrossRef]
- Sörensen, S.P.L. Enzymstudien. II. Mitteilung. Über die Messung und die Bedeutung der Wasserstoffionenkoncentration bei enzymatischen Prozessen. Biochem. Z. 1909, 21, 131–304. [Google Scholar]
- Bamforth, C. Beer: Tap Into The Art and Science of Brewing, 3rd ed.; Oxford University Press: Oxford, UK, 2009; p. 239. ISBN 978-0-19-530542-5. [Google Scholar]
- Wunderlich, S.; Zürcher, A.; Back, W. Enrichment of xanthohumol in the brewing process. Mol. Nutr. Food Res. 2005, 49, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, P.J.; Carvalho, D.O.; Cruz, J.M.; Guido, L.F.; Barros, A.A. Fundamentals and health benefits of xanthohumol, a natural product derived from hops and beer. Nat. Prod. Commun. 2009, 4, 591–610. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Dong, J.; Yin, H.; Zhao, Y.; Chen, R.; Wan, X.; Chen, P.; Hou, X.; Liu, J.; Chen, L. Wort composition and its impact on the flavour-active higher alcohol and ester formation of beer—A review. J. Inst. Brew. 2014. [Google Scholar] [CrossRef]
- Waterhouse, A.L.; Sacks, G.L.; Jeffery, D.W. Outline of wine production. In Understanding Wine Chemistry; John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 159–171. ISBN 9781118627808. [Google Scholar]
- Fleet, G.H. Growth of yeasts during wine fermentations. J. Wine Res. 1990, 1, 211–223. [Google Scholar] [CrossRef]
- Tronchoni, J.; Gamero, A.; Arroyo-López, F.N.; Barrio, E.; Querol, A. Differences in the glucose and fructose consumption profiles in diverse Saccharomyces wine species and their hybrids during grape juice fermentation. Int. J. Food Microbiol. 2009, 134, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Moritz, E.R.; Morris, G.H. A Text-Book of the Science of Brewing; Spon: London, UK, 1891. [Google Scholar]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410.e16. [Google Scholar] [CrossRef] [PubMed]
- Fay, J.C.; Liu, P.; Ong, G.T.; Dunham, M.J.; Cromie, G.A.; Jeffery, E.W.; Ludlow, C.L.; Dudley, A.M. A polyploid admixed origin of beer yeasts derived from European and Asian wine populations. PLoS Biol. 2019, 17, e3000147. [Google Scholar] [CrossRef] [PubMed]
- Olesen, K.; Felding, T.; Gjermansen, C.; Hansen, J. The dynamics of the brewing yeast transcriptome during a production-scale lager beer fermentation. Fems Yeast Res. 2002, 2, 563–573. [Google Scholar] [CrossRef]
- Denby, C.M.; Li, R.A.; Vu, V.T.; Costello, Z.; Lin, W.; Chan, L.J.G.; Williams, J.; Donaldson, B.; Bamforth, C.W.; Petzold, C.J.; et al. Industrial brewing yeast engineered for the production of primary flavor determinants in hopped beer. Nat. Commun. 2018, 9, 965. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Hiralal, L.; Mokoena, M.P.; Pillay, B. Flavour-active volatile compounds in beer: Production, regulation and control. J. Inst. Brew. 2017, 123, 13–23. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Saerens, S.M.G.; Pretorius, I.S. The development of yeast strains as tools for adjusting the flavor of fermented beverages to market specifications. In Biotechnology in Flavor Production; Havkin-Frenkel, D., Belanger, F.C., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2008; pp. 1–55. ISBN 9781405156493. [Google Scholar]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Glycolysis is an Energy-Conversion Pathway in Many Organisms. 2002. Available online: https://www.ncbi.nlm.nih.gov/books/NBK22593/ (accessed on 4 May 2020).
- Aranda, A.; Orozco, H.; Picazo, C.; Matallana, E. Yeast life span and its impact on food fermentations. Fermentation 2019, 5, 37. [Google Scholar] [CrossRef]
- Dunham, M.J.; Badrane, H.; Ferea, T.; Adams, J.; Brown, P.O.; Rosenzweig, F.; Botstein, D. Characteristic genome rearrangements in experimental evolution of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2002, 99, 16144–16149. [Google Scholar] [CrossRef]
- Gresham, D.; Desai, M.M.; Tucker, C.M.; Jenq, H.T.; Pai, D.A.; Ward, A.; DeSevo, C.G.; Botstein, D.; Dunham, M.J. The repertoire and dynamics of evolutionary adaptations to controlled nutrient-limited environments in yeast. PLoS Genet. 2008, 4, e1000303. [Google Scholar] [CrossRef] [PubMed]
- Scannell, D.R.; Zill, O.A.; Rokas, A.; Payen, C.; Dunham, M.J.; Eisen, M.B.; Rine, J.; Johnston, M.; Hittinger, C.T. The Awesome Power of Yeast Evolutionary Genetics: New Genome Sequences and Strain Resources for the Saccharomyces sensu stricto Genus. G3 (Bethesda) 2011, 1, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Large, C.R.L.; Hanson, N.A.; Tsouris, A.; Abou Saada, O.; Koonthongkaew, J.; Toyokawa, Y.; Schmidlin, T.; Moreno-Habel, D.A.; McConnellogue, H.; Preiss, R.; et al. Genomic stability and adaptation of beer brewing yeasts during serial repitching in the brewery. BioRxiv 2020. [Google Scholar] [CrossRef]
- Mensour, N.A.; Margaritis, A.; Briens, C.L.; Pilkington, H.; Russell, I. Application of immobilized yeast cells in the brewing industry. In Immobilized Cells—Basics and Applications; Progress in Biotechnology; Elsevier: Amsterdam, The Netherlands, 1996; Volume 11, pp. 661–671. ISBN 9780444819840. [Google Scholar]
- Marongiu, A.; Zara, G.; Legras, J.-L.; Del Caro, A.; Mascia, I.; Fadda, C.; Budroni, M. Novel starters for old processes: Use of Saccharomyces cerevisiae strains isolated from artisanal sourdough for craft beer production at a brewery scale. J. Ind. Microbiol. Biotechnol. 2015, 42, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Canonico, L.; Comitini, F.; Ciani, M. Metschnikowia pulcherrima Selected Strain for Ethanol Reduction in Wine: Influence of Cell Immobilization and Aeration Condition. Foods 2019, 8, 378. [Google Scholar] [CrossRef]
- Canonico, L.; Agarbati, A.; Comitini, F.; Ciani, M. Torulaspora delbrueckii in the brewing process: A new approach to enhance bioflavour and to reduce ethanol content. Food Microbiol. 2016, 56, 45–51. [Google Scholar] [CrossRef]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef]
- Michel, M.; Meier-Dörnberg, T.; Jacob, F.; Methner, F.-J.; Wagner, R.S.; Hutzler, M. Review: Pure non- Saccharomyces starter cultures for beer fermentation with a focus on secondary metabolites and practical applications. J. Inst. Brew. 2016, 122, 569–587. [Google Scholar] [CrossRef]
- Basso, R.F.; Alcarde, A.R.; Portugal, C.B. Could non-Saccharomyces yeasts contribute on innovative brewing fermentations? Food Res. Int. 2016, 86, 112–120. [Google Scholar] [CrossRef]
- Alonso-Del-Real, J.; Lairón-Peris, M.; Barrio, E.; Querol, A. Effect of Temperature on the Prevalence of Saccharomyces Non cerevisiae Species against a S. cerevisiae Wine Strain in Wine Fermentation: Competition, Physiological Fitness, and Influence in Final Wine Composition. Front. Microbiol. 2017, 8, 150. [Google Scholar] [CrossRef]
- Pérez-Torrado, R.; Barrio, E.; Querol, A. Alternative yeasts for winemaking: Saccharomyces non-cerevisiae and its hybrids. Crit. Rev. Food Sci. Nutr. 2018, 58, 1780–1790. [Google Scholar] [CrossRef] [PubMed]
- Kodama, Y.; Kielland-Brandt, M.C.; Hansen, J. Lager brewing yeast. In Comparative Genomics; Sunnerhagen, P., Piskur, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 145–164. ISBN 978-3-540-31480-6. [Google Scholar]
- Rainieri, S.; Kodama, Y.; Kaneko, Y.; Mikata, K.; Nakao, Y.; Ashikari, T. Pure and mixed genetic lines of Saccharomyces bayanus and Saccharomyces pastorianus and their contribution to the lager brewing strain genome. Appl. Environ. Microbiol. 2006, 72, 3968–3974. [Google Scholar] [CrossRef]
- Gibson, B.; Liti, G. Saccharomyces pastorianus: Genomic insights inspiring innovation for industry. Yeast 2015, 32, 17–27. [Google Scholar] [CrossRef] [PubMed]
- White, C.; Zainasheff, J. Yeast: The Practical Guide To Beer Fermentation; Brewers Publications: Boulder, CO, USA, 2010; p. 300. ISBN 978-0937381960. [Google Scholar]
- Mortimer, R.K. Evolution and variation of the yeast (Saccharomyces) genome. Genome Res. 2000, 10, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Naumov, G.I.; James, S.A.; Naumova, E.S.; Louis, E.J.; Roberts, I.N. Three new species in the Saccharomyces sensu stricto complex: Saccharomyces cariocanus, Saccharomyces kudriavzevii and Saccharomyces mikatae. Int. J. Syst. Evol. Microbiol. 2000, 50, 1931–1942. [Google Scholar] [CrossRef]
- Borneman, A.R.; Pretorius, I.S. Genomic insights into the Saccharomyces sensu stricto complex. Genetics 2015, 199, 281–291. [Google Scholar] [CrossRef]
- Naumov, G. Genetic identification of biological species in the Saccharomyces sensu stricto complex. J. Ind. Microbiol. Biotechnol. 1996, 17, 295–302. [Google Scholar] [CrossRef]
- Bing, J.; Han, P.-J.; Liu, W.-Q.; Wang, Q.-M.; Bai, F.-Y. Evidence for a Far East Asian origin of lager beer yeast. Curr. Biol. 2014, 24, R380-1. [Google Scholar] [CrossRef]
- Fay, J.C. Tapping into yeast diversity. Mol. Ecol. 2012, 21, 5387–5389. [Google Scholar] [CrossRef]
- Sampaio, J.P.; Gonçalves, P. Natural populations of Saccharomyces kudriavzevii in Portugal are associated with oak bark and are sympatric with S. cerevisiae and S. paradoxus. Appl. Environ. Microbiol. 2008, 74, 2144–2152. [Google Scholar] [CrossRef]
- Erny, C.; Raoult, P.; Alais, A.; Butterlin, G.; Delobel, P.; Matei-Radoi, F.; Casaregola, S.; Legras, J.L. Ecological success of a group of Saccharomyces cerevisiae/Saccharomyces kudriavzevii hybrids in the northern european wine-making environment. Appl. Environ. Microbiol. 2012, 78, 3256–3265. [Google Scholar] [CrossRef] [PubMed]
- Peris, D.; Pérez-Torrado, R.; Hittinger, C.T.; Barrio, E.; Querol, A. On the origins and industrial applications of Saccharomyces cerevisiae × Saccharomyces kudriavzevii hybrids. Yeast 2018, 35, 51–69. [Google Scholar] [CrossRef]
- Kellis, M.; Patterson, N.; Endrizzi, M.; Birren, B.; Lander, E.S. Sequencing and comparison of yeast species to identify genes and regulatory elements. Nature 2003, 423, 241–254. [Google Scholar] [CrossRef] [PubMed]
- González, S.S.; Barrio, E.; Querol, A. Molecular characterization of new natural hybrids of Saccharomyces cerevisiae and S. kudriavzevii in brewing. Appl. Environ. Microbiol. 2008, 74, 2314–2320. [Google Scholar] [CrossRef] [PubMed]
- Scholtes, C.; Nizet, S.; Collin, S. Guaiacol and 4-methylphenol as specific markers of torrefied malts. Fate of volatile phenols in special beers through aging. J. Agric. Food Chem. 2014, 62, 9522–9528. [Google Scholar] [CrossRef] [PubMed]
- Henriques, D.; Alonso-Del-Real, J.; Querol, A.; Balsa-Canto, E. Saccharomyces cerevisiae and S. kudriavzevii Synthetic Wine Fermentation Performance Dissected by Predictive Modeling. Front. Microbiol. 2018, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Gallone, B.; Steensels, J.; Mertens, S.; Dzialo, M.C.; Gordon, J.L.; Wauters, R.; Theßeling, F.A.; Bellinazzo, F.; Saels, V.; Herrera-Malaver, B.; et al. Interspecific hybridization facilitates niche adaptation in beer yeast. Nat. Ecol. Evol. 2019, 3, 1562–1575. [Google Scholar] [CrossRef]
- Groth, C.; Hansen, J.; Piskur, J. A natural chimeric yeast containing genetic material from three species. Int. J. Syst. Bacteriol. 1999, 49, 1933–1938. [Google Scholar] [CrossRef]
- Masneuf, I.; Hansen, J.; Groth, C.; Piskur, J.; Dubourdieu, D. New hybrids between Saccharomyces sensu stricto yeast species found among wine and cider production strains. Appl. Environ. Microbiol. 1998, 64, 3887–3892. [Google Scholar] [CrossRef]
- Belloch, C.; Orlic, S.; Barrio, E.; Querol, A. Fermentative stress adaptation of hybrids within the Saccharomyces sensu stricto complex. Int. J. Food Microbiol. 2008, 122, 188–195. [Google Scholar] [CrossRef]
- Salvadó, Z.; Arroyo-López, F.N.; Guillamón, J.M.; Salazar, G.; Querol, A.; Barrio, E. Temperature adaptation markedly determines evolution within the genus Saccharomyces. Appl. Environ. Microbiol. 2011, 77, 2292–2302. [Google Scholar] [CrossRef]
- VIN 7. Available online: http://www.oenobrands.com/en/our-brands/anchor/new-world-wine-yeasts/product-data-sheets/vin-7 (accessed on 27 April 2020).
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The Role and Use of Non-Saccharomyces Yeasts in Wine Production. SAJEV 2017, 27. [Google Scholar] [CrossRef]
- Martini, A.V.; Martini, A. A.V.; Martini, A. A proposal for correct nomenclature of the domesticated species of the genus saccharomyces. In Biotechnology Applications in Beverage Production; Cantarelli, C., Lanzarini, G., Eds.; Springer: Dordrecht, The Netherlands, 1989; pp. 1–16. ISBN 978-94-009-1113-0. [Google Scholar]
- Sniegowski, P.D.; Dombrowski, P.G.; Fingerman, E. Saccharomyces cerevisiae and Saccharomyces paradoxus coexist in a natural woodland site in North America and display different levels of reproductive isolation from European conspecifics. Fems Yeast Res. 2002, 1, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Hyma, K.E.; Fay, J.C. Mixing of vineyard and oak-tree ecotypes of Saccharomyces cerevisiae in North American vineyards. Mol. Ecol. 2013, 22, 2917–2930. [Google Scholar] [CrossRef]
- Charron, G.; Leducq, J.-B.; Bertin, C.; Dubé, A.K.; Landry, C.R. Exploring the northern limit of the distribution of Saccharomyces cerevisiae and Saccharomyces paradoxus in North America. Fems Yeast Res. 2014, 14, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Liti, G.; Carter, D.M.; Moses, A.M.; Warringer, J.; Parts, L.; James, S.A.; Davey, R.P.; Roberts, I.N.; Burt, A.; Koufopanou, V.; et al. Population genomics of domestic and wild yeasts. Nature 2009, 458, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Roop, J.I.; Brem, R.B. Rare variants in hypermutable genes underlie common morphology and growth traits in wild Saccharomyces paradoxus. Genetics 2013, 195, 513–525. [Google Scholar] [CrossRef]
- Redzepović, S.; Orlić, S.; Sikora, S.; Majdak, A.; Pretorius, I.S. Identification and characterization of Saccharomyces cerevisiae and Saccharomyces paradoxus strains isolated from Croatian vineyards. Lett. Appl. Microbiol. 2002, 35, 305–310. [Google Scholar] [CrossRef]
- Orlic, S.; Redzepovic, S.; Jeromel, A.; Herjavec, S.; Iacumin, L. Influence of indigenous Saccharomyces paradoxus strains on Chardonnay wine fermentation aroma. Int. J. Food Sci. Technol. 2007, 42, 95–101. [Google Scholar] [CrossRef]
- Majdak, A.; Herjavec, S.; Orlic, S.; Redzepovic, S.; Mirosevic, N. Comparison of Wine Aroma Compounds Produced by Saccharomyces paradoxus and Saccharomyces cerevisiae Strains. Food Technol. Biotechnol. 2002, 40, 103–109. [Google Scholar]
- Naumova, E.; Korshunova, I.; Jespersen, L.; Naumov, G. Molecular genetic identification of sensu stricto strains from African sorghum beer. Fems Yeast Res. 2003, 3, 177–184. [Google Scholar] [CrossRef]
- Sweeney, J.; Kuehne, H.; Sniegowski, P. Sympatric natural and populations have different thermal growth profiles. Fems Yeast Res. 2004, 4, 521–525. [Google Scholar] [CrossRef]
- Gonçalves, P.; Valério, E.; Correia, C.; de Almeida, J.M.G.C.F.; Sampaio, J.P. Evidence for divergent evolution of growth temperature preference in sympatric Saccharomyces species. PLoS ONE 2011, 6, e20739. [Google Scholar] [CrossRef]
- Rollero, S.; Bloem, A.; Ortiz-Julien, A.; Camarasa, C.; Divol, B. Fermentation performances and aroma production of non-conventional wine yeasts are influenced by nitrogen preferences. Fems Yeast Res. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Dunn, B.; Richter, C.; Kvitek, D.J.; Pugh, T.; Sherlock, G. Analysis of the Saccharomyces cerevisiae pan-genome reveals a pool of copy number variants distributed in diverse yeast strains from differing industrial environments. Genome Res. 2012, 22, 908–924. [Google Scholar] [CrossRef]
- Bellon, J.R.; Schmid, F.; Capone, D.L.; Dunn, B.L.; Chambers, P.J. Introducing a new breed of wine yeast: Interspecific hybridisation between a commercial Saccharomyces cerevisiae wine yeast and Saccharomyces mikatae. PLoS ONE 2013, 8, e62053. [Google Scholar] [CrossRef] [PubMed]
- Bellon, J.; Schmidt, S.; Solomon, M. Case study: Development of Saccharomyces cerevisiae × Saccharomyces mikatae wine yeast hybrids and their potential to deliver alternative wine styles. AWRI Tech. Rev. 2019, 241, 6–11. [Google Scholar]
- Pulvirenti, A.; Nguyen, H.; Caggia, C.; Giudici, P.; Rainieri, S.; Zambonelli, C. Saccharomyces uvarum, a proper species within Saccharomyces sensu stricto. FEMS Microbiol. Lett. 2000, 192, 191–196. [Google Scholar] [CrossRef]
- Nguyen, H.-V.; Gaillardin, C. Evolutionary relationships between the former species Saccharomyces uvarum and the hybrids Saccharomyces bayanus and Saccharomyces pastorianus; reinstatement of Saccharomyces uvarum (Beijerinck) as a distinct species. FEMS Yeast Res. 2005, 5, 471–483. [Google Scholar] [CrossRef]
- Rainieri, S.; Zambonelli, C.; Hallsworth, J.E.; Pulvirenti, A.; Giudici, P. Saccharomyces uvarum, a distinct group within Saccharomyces sensu stricto. FEMS Microbiol. Lett. 1999, 177, 177–185. [Google Scholar] [CrossRef]
- Naumova, E.S.; Naumov, G.I.; Masneuf-Pomarède, I.; Aigle, M.; Dubourdieu, D. Molecular genetic study of introgression between Saccharomyces bayanus and S. cerevisiae. Yeast 2005, 22, 1099–1115. [Google Scholar] [CrossRef] [PubMed]
- Demuyter, C.; Lollier, M.; Legras, J.L.; Le Jeune, C. Predominance of Saccharomyces uvarum during spontaneous alcoholic fermentation, for three consecutive years, in an Alsatian winery. J. Appl. Microbiol. 2004, 97, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Tosi, E.; Azzolini, M.; Guzzo, F.; Zapparoli, G. Evidence of different fermentation behaviours of two indigenous strains of Saccharomyces cerevisiae and Saccharomyces uvarum isolated from Amarone wine. J. Appl. Microbiol. 2009, 107, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.E.; Pérez-Través, L.; Sangorrín, M.P.; Barrio, E.; Querol, A.; Lopes, C.A. Saccharomyces uvarum is responsible for the traditional fermentation of apple chicha in Patagonia. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.E.; Pérez-Través, L.; Sangorrín, M.P.; Barrio, E.; Lopes, C.A. Saccharomyces eubayanus and Saccharomyces uvarum associated with the fermentation of Araucaria araucana seeds in Patagonia. FEMS Yeast Res. 2014, 14, 948–965. [Google Scholar] [CrossRef] [PubMed]
- Beijerinck, M.W. Über Regeneration der Sporenbildung bei Alkoholhefen, wo diese Function im Verschwinden begriffen ist. Cent. Bakteriol. Parasitenkd. 1898, 4, 721–730. [Google Scholar]
- Krogerus, K.; Preiss, R.; Gibson, B. A Unique Saccharomyces cerevisiae × Saccharomyces uvarum Hybrid Isolated from Norwegian Farmhouse Beer: Characterization and Reconstruction. Front. Microbiol. 2018, 9, 2253. [Google Scholar] [CrossRef]
- Nguyen, H.-V.; Boekhout, T. Characterization of Saccharomyces uvarum (Beijerinck, 1898) and related hybrids: Assessment of molecular markers that predict the parent and hybrid genomes and a proposal to name yeast hybrids. Fems Yeast Res. 2017, 17. [Google Scholar] [CrossRef]
- Krogerus, K.; Gibson, B. A re-evaluation of diastatic Saccharomyces cerevisiae strains and their role in brewing. Appl. Microbiol. Biotechnol. 2020, 104, 3745–3756. [Google Scholar] [CrossRef]
- Smukowski Heil, C.S.; Large, C.R.L.; Patterson, K.; Hickey, A.S.-M.; Yeh, C.-L.C.; Dunham, M.J. Temperature preference can bias parental genome retention during hybrid evolution. PLoS Genet. 2019, 15, e1008383. [Google Scholar] [CrossRef]
- Morgan, S.C.; Haggerty, J.J.; Jiranek, V.; Durall, D.M. Competition between Saccharomyces cerevisiae and Saccharomyces uvarum in Controlled Chardonnay Wine Fermentations. Am. J. Enol. Vitic. 2020. [Google Scholar] [CrossRef]
- Feuillat, M. Use of Yeasts in Burgandy and In Other Regions: Fermentation and Ading on Lees. In Yeast’s Contribution to the Sensory Profile of Wine: Maintaining Typicity and Biodiversity in the Context of Globalization; Lallemand, S.A.S., Ed.; Entretiens Scientifiques: Wiesbaden, Germany, 2005; Volume XVII, pp. 27–32. [Google Scholar]
- Le Jeune, C.; Lollier, M.; Demuyter, C.; Erny, C.; Legras, J.-L.; Aigle, M.; Masneuf-Pomarède, I. Characterization of natural hybrids of Saccharomyces cerevisiae and Saccharomyces bayanus var. uvarum. FEMS Yeast Res. 2007, 7, 540–549. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Available Microbial Strains—The Australian Wine Research Institute. Available online: https://www.awri.com.au/research_and_development/wine-microorganism/winemaking-yeast-and-bacterial-strains/ (accessed on 3 May 2020).
- Caudy, A.A.; Guan, Y.; Jia, Y.; Hansen, C.; DeSevo, C.; Hayes, A.P.; Agee, J.; Alvarez-Dominguez, J.R.; Arellano, H.; Barrett, D.; et al. A new system for comparative functional genomics of Saccharomyces yeasts. Genetics 2013, 195, 275–287. [Google Scholar] [CrossRef]
- Vaughan Martini, A.; Kurtzman, C.P. Deoxyribonucleic Acid Relatedness among Species of the Genus Saccharomyces Sensu Stricto. Int. J. Syst. Bacteriol. 1985, 35, 508–511. [Google Scholar] [CrossRef]
- Nguyen, H.-V.; Legras, J.-L.; Neuvéglise, C.; Gaillardin, C. Deciphering the hybridisation history leading to the Lager lineage based on the mosaic genomes of Saccharomyces bayanus strains NBRC1948 and CBS380. PLoS ONE 2011, 6, e25821. [Google Scholar] [CrossRef] [PubMed]
- Libkind, D.; Hittinger, C.T.; Valério, E.; Gonçalves, C.; Dover, J.; Johnston, M.; Gonçalves, P.; Sampaio, J.P. Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 14539–14544. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, P.M.C. Microbe Domestication and the Identification of the Wild Genetic Stock of Wine Yeasts. Proc. Natl. Acad. Sci. USA 2016. Available online: http://hdl.handle.net/10362/19871 (accessed on 24 November 2020).
- González, S.S.; Barrio, E.; Gafner, J.; Querol, A. Natural hybrids from Saccharomyces cerevisiae, Saccharomyces bayanus and Saccharomyces kudriavzevii in wine fermentations. FEMS Yeast Res. 2006, 6, 1221–1234. [Google Scholar] [CrossRef]
- Wendland, J. Lager yeast comes of age. Eukaryot. Cell 2014, 13, 1256–1265. [Google Scholar] [CrossRef]
- Sicard, D.; Legras, J.-L. Bread, beer and wine: Yeast domestication in the Saccharomyces sensu stricto complex. C. R. Biol. 2011, 334, 229–236. [Google Scholar] [CrossRef]
- Ugliano, M.; Fedrizzi, B.; Siebert, T.; Travis, B.; Magno, F.; Versini, G.; Henschke, P.A. Effect of nitrogen supplementation and Saccharomyces species on hydrogen sulfide and other volatile sulfur compounds in shiraz fermentation and wine. J. Agric. Food Chem. 2009, 57, 4948–4955. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Espinar, M. Study of the authenticity of commercial wine yeast strains by molecular techniques. Int. J. Food Microbiol. 2001, 70, 1–10. [Google Scholar] [CrossRef]
- Hoff, J.W. Molecular Typing of Wine Yeasts: Evaluation of Typing Techniques and Establishment of a Database. 2012. Available online: https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.933.7961&rep=rep1&type=pdf (accessed on 1 May 2020).
- Nakao, Y.; Kanamori, T.; Itoh, T.; Kodama, Y.; Rainieri, S.; Nakamura, N.; Shimonaga, T.; Hattori, M.; Ashikari, T. Genome sequence of the lager brewing yeast, an interspecies hybrid. DNA Res. 2009, 16, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Cordente, A.G.; Curtin, C.D.; Varela, C.; Pretorius, I.S. Flavour-active wine yeasts. Appl. Microbiol. Biotechnol. 2012, 96, 601–618. [Google Scholar] [CrossRef]
- Hisamoto, M.; Furuya, T.; Yanagida, A.; Okuda, T.; Sato, M. Phenolic off Flavor Characterization of Commercially Available Wine Yeasts and Selection of the Yeast for Koshu Winemaking. J. ASEV Jpn. 2010, 21, 112–117. [Google Scholar]
- Ward, H.M. V. the ginger-beer plant, and the organisms composing it: A contribution to the study of fermentation-yeasts and bacteria. Philos. Trans. R. Soc. Lond. B 1892, 183, 125–197. [Google Scholar] [CrossRef]
- Pidoux, M. The microbial flora of sugary kefir grain (the gingerbeer plant): Biosynthesis of the grain fromLactobacillus hilgardii producing a polysaccharide gel. World J. Microbiol. Biotechnol. 1989, 5, 223–238. [Google Scholar] [CrossRef]
- Naumov, G.I.; Naumova, E.S.; Masneuf-Pomarède, I. Genetic identification of new biological species Saccharomyces arboricolus Wang et Bai. Antonie Van Leeuwenhoek 2010, 98, 1–7. [Google Scholar] [CrossRef]
- Wang, S.-A.; Bai, F.-Y. Saccharomyces arboricolus sp. nov., a yeast species from tree bark. Int. J. Syst. Evol. Microbiol. 2008, 58, 510–514. [Google Scholar] [CrossRef]
- Winans, M.J.; Yamamoto, Y.; Fujimaru, Y.; Kusaba, Y.; Gallagher, J.E.G.; Kitagaki, H. Saccharomyces arboricola and Its Hybrids’ Propensity for Sake Production: Interspecific Hybrids Reveal Increased Fermentation Abilities and a Mosaic Metabolic Profile. Fermentation 2020, 6, 14. [Google Scholar] [CrossRef]
- Naseeb, S.; James, S.A.; Alsammar, H.; Michaels, C.J.; Gini, B.; Nueno-Palop, C.; Bond, C.J.; McGhie, H.; Roberts, I.N.; Delneri, D. Suppl. Saccharomyces jurei sp. nov., isolation and genetic identification of a novel yeast species from Quercus robur. Int. J. Syst. Evol. Microbiol. 2017, 67, 2046–2052. [Google Scholar] [CrossRef] [PubMed]
- Liti, G.; Barton, D.B.H.; Louis, E.J. Sequence diversity, reproductive isolation and species concepts in Saccharomyces. Genetics 2006, 174, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, R.; Almeida, P.; Safar, S.V.B.; Santos, R.O.; Morais, P.B.; Nielly-Thibault, L.; Leducq, J.-B.; Landry, C.R.; Gonçalves, P.; Rosa, C.A.; et al. Evidence of Natural Hybridization in Brazilian Wild Lineages of Saccharomyces cerevisiae. Genome Biol. Evol. 2016, 8, 317–329. [Google Scholar] [CrossRef] [PubMed]
- de Paula, B.P.; Chávez, D.W.H.; Lemos Junior, W.J.F.; Guerra, A.F.; Corrêa, M.F.D.; Pereira, K.S.; Coelho, M.A.Z. Growth Parameters and Survivability of Saccharomyces boulardii for Probiotic Alcoholic Beverages Development. Front. Microbiol. 2019, 10, 2092. [Google Scholar] [CrossRef] [PubMed]
- Capece, A.; Romaniello, R.; Pietrafesa, A.; Siesto, G.; Pietrafesa, R.; Zambuto, M.; Romano, P. Use of Saccharomyces cerevisiae var. boulardii in co-fermentations with S. cerevisiae for the production of craft beers with potential healthy value-added. Int. J. Food Microbiol. 2018, 284, 22–30. [Google Scholar] [CrossRef]
- Mulero-Cerezo, J.; Briz-Redón, Á.; Serrano-Aroca, Á. Saccharomyces cerevisiae var. boulardii: Valuable probiotic starter for craft beer production. Appl. Sci. 2019, 9, 3250. [Google Scholar] [CrossRef]
- Van der Aa Kühle, A.; Jespersen, L. The taxonomic position of Saccharomyces boulardii as evaluated by sequence analysis of the D1/D2 domain of 26S rDNA, the ITS1-5.8S rDNA-ITS2 region and the mitochondrial cytochrome-c oxidase II gene. Syst. Appl. Microbiol. 2003, 26, 564–571. [Google Scholar] [CrossRef]
- Ramírez-Cota, G.Y.; López-Villegas, E.O.; Jiménez-Aparicio, A.R.; Hernández-Sánchez, H. Modeling the Ethanol Tolerance of the Probiotic Yeast Saccharomyces cerevisiae var. boulardii CNCM I-745 for its Possible Use in a Functional Beer. Probiotics Antimicrob. Proteins 2020. [Google Scholar] [CrossRef]
- Sampaolesi, S.; Gamba, R.R.; De Antoni, G.L.; León Peláez, Á.M. Potentiality of yeasts obtained as beer fermentation residue to be used as probiotics. LWT 2019, 113, 108251. [Google Scholar] [CrossRef]
- Sakai, K.; Fukui, S.; Yabuuchi, S.; Aoyagi, S.; Tsumura, Y. Expression of the saccharomyces diastaticus sta1 gene in brewing yeasts. ASBCJ 1989, 47, 87–91. [Google Scholar] [CrossRef]
- Meier-Dörnberg, T.; Kory, O.I.; Jacob, F.; Michel, M.; Hutzler, M. Saccharomyces cerevisiae variety diastaticus friend or foe?-spoilage potential and brewing ability of different Saccharomyces cerevisiae variety diastaticus yeast isolates by genetic, phenotypic and physiological characterization. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, H.; Yamamoto, H.; Shibano, Y.; Amaya, N.; Saeki, T. Rapid methods for detecting Saccharomyces diastaticus, a beer spoilage yeast, using the polymerase chain reaction. ASBCJ 1998, 56, 58–63. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Year | Scientist | Employment | Discovery |
|---|---|---|---|
| 1762 | Michael Combrune | Brewer’s Company Middlesex | using a thermometer for analysis [2] |
| 1769 | James Baverstock | family brewery | using a hydrometer for analysis [2] |
| 1833 | Anselme Payen and Jean-François Perzoz | École Centrale Paris | discovered diastase enzyme and cellulose while working with barley [2] |
| 1843 | Karl J.N. Balling | Polytechnic in Prague | invents the balling saccharimeter [6] |
| 1843 | James Joule and Lord Kelvin | family brewery | create temperature scale and first law of thermodynamics [7] |
| 1857 | Louis Pasteur | University of Lille | microbes are responsible for fermentation [8] |
| 1860 | P.E. Marcellin Berthelot | Collège de France | discovered invertase in Saccharomyces [2] |
| 1873 | Carl von Linde | Spaten Brewery | invented the refrigeration cycle [2] |
| 1883 | Johan Kjeldahl | Carlsberg Brewery | develops method for protein quantification [9] |
| 1888 | Emil Christian Hansen | Carlsberg Brewery | first isolation of pure yeast strain [9] |
| 1908 | William Sealy Gosset | Guinness Brewery | invents the statistical t-test for students [10] |
| 1909 | Søren Sørenson | Carlsberg Brewery | creates pH scale based on H+ ion concentration [11] |
| S. kudriavzevii | S. paradoxus | S. uvarum | S. mikatae | S. bayanus | |
|---|---|---|---|---|---|
| Fermentation Of: | |||||
| Maltose | + | + | + | + | + |
| Melibiose | − | − | + | + | −,+ |
| Dextrins (STA1) | −,+ | − | −,+ | − | − |
| Ethanol Tolerant | + | + | + | + | + |
| Characteristics: | |||||
| Attenuation | moderate | low-moderate | moderate | moderate | moderate |
| Flocculation | moderate-high | moderate | high | moderate | moderate |
| Growth at 10 °C | + | + | + | + | + |
| Growth at 25 °C | + | + | + | + | + |
| Growth at 37 °C | − | − | + | − | + |
| POF | + | u | − | u | + |
| Region of Origin | Western Europe | Northeastern Europe | Scandinavia | Japan | Europe |
| Isolated From | Oak tree bark | Oak sap | Fruit/Seeds | Soil/Leaves | Insects/Leaves |
| Type Strain | NCYC 2889T | DBVPG 6411 | DBVPG 6173 | NCYC 2888T | CBS 380 |
| CommercialAvailability | Anchor Vin7 | Anchor Exotics SPH | AWRI 1176 & 1375 | AB Biotek/AWRI 2526 | Lalvin S6U |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruner, J.; Fox, G. Novel Non-Cerevisiae Saccharomyces Yeast Species Used in Beer and Alcoholic Beverage Fermentations. Fermentation 2020, 6, 116. https://doi.org/10.3390/fermentation6040116
Bruner J, Fox G. Novel Non-Cerevisiae Saccharomyces Yeast Species Used in Beer and Alcoholic Beverage Fermentations. Fermentation. 2020; 6(4):116. https://doi.org/10.3390/fermentation6040116
Chicago/Turabian StyleBruner, James, and Glen Fox. 2020. "Novel Non-Cerevisiae Saccharomyces Yeast Species Used in Beer and Alcoholic Beverage Fermentations" Fermentation 6, no. 4: 116. https://doi.org/10.3390/fermentation6040116
APA StyleBruner, J., & Fox, G. (2020). Novel Non-Cerevisiae Saccharomyces Yeast Species Used in Beer and Alcoholic Beverage Fermentations. Fermentation, 6(4), 116. https://doi.org/10.3390/fermentation6040116