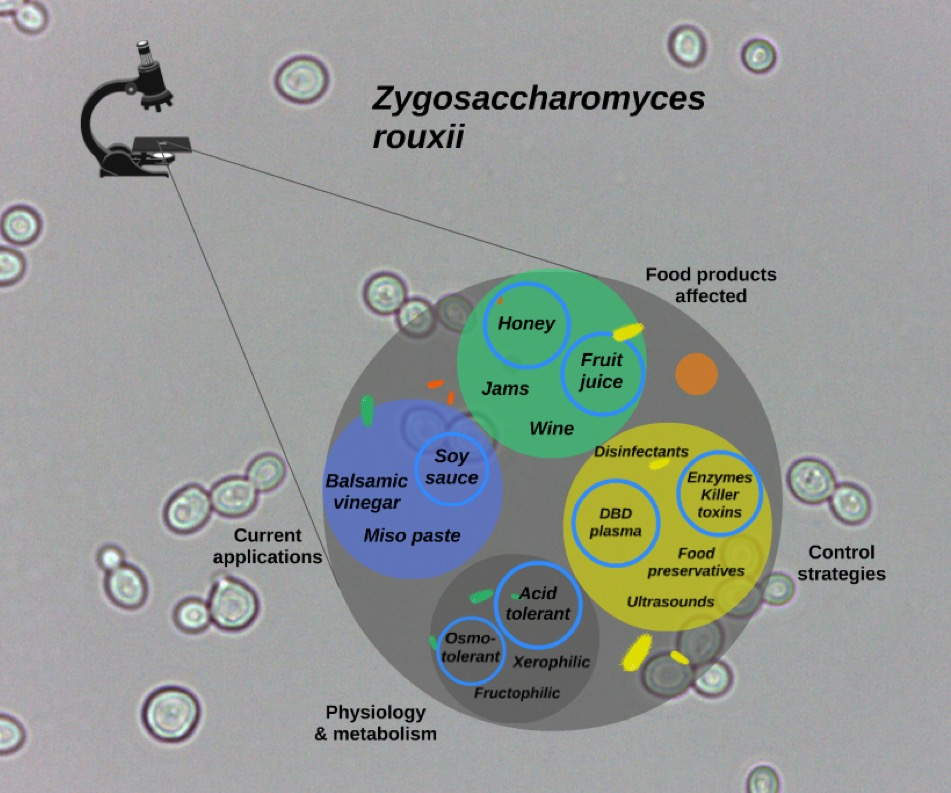
Zygosaccharomyces rouxii: Control Strategies and Applications in Food and Winemaking

 ,
, 
 ,
,  and
and
Abstract
:
1. The Genus Zygosaccharomyces
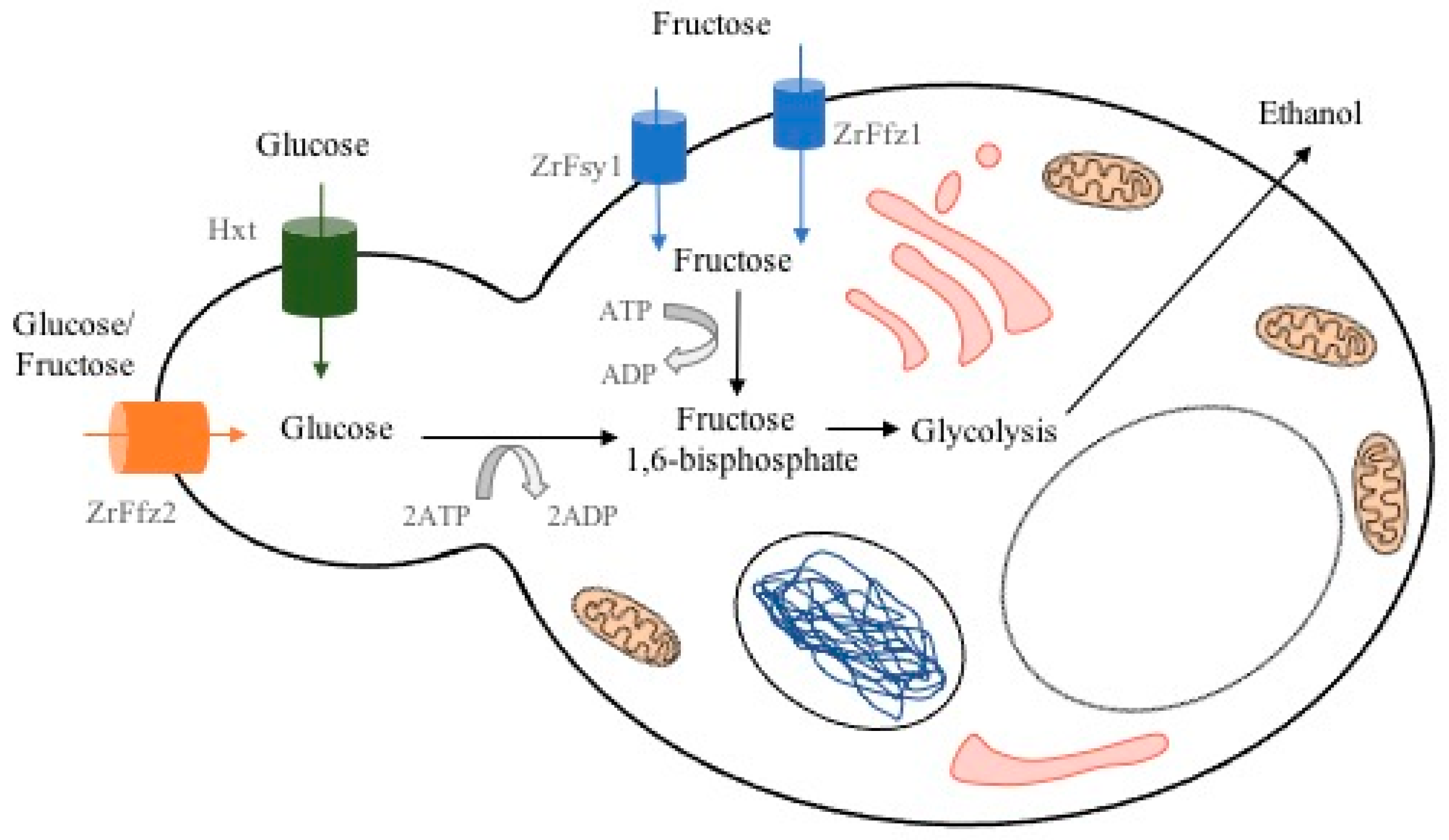
2. Physiology and Metabolism of Z. rouxii
3. Food Spoilage Activity
4. Detection
5. Control Strategies
6. Food Applications
7. Alcohol-Fermentation Applications
8. Conclusions
Author Contributions
Conflicts of Interest
References
- Esteve-Zarzoso, B.; Zorman, T.; Belloch, C.; Quero, A. Molecular Characterisation of the Species of the Genus Zyosaccharomyces. Syst. Appl. Microbiol. 2003, 26, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Hulin, M.; Wheals, A. Rapid Identification of Zygosaccharomyces with Genus-Specific Primers. Int. J. Food Microbiol. 2014, 173, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, Z.; Long, F.; Guo, C.; Niu, C.; Yuan, Y.; Yue, T. Combined Effect of Sugar Content and PH on the Growth of a Wild Strain of Zygosaccharomyces Rouxii and Time for Spoilage in Concentrated Apple Juice. Food Control. 2016, 59, 298–305. [Google Scholar] [CrossRef]
- Romano, P.; Suzzi, G. Higher Alcohol and Acetoin Production by Zygosaccharomyces Wine Yeasts. J. Appl. Bacteriol. 1993, 75, 541–545. [Google Scholar] [CrossRef]
- Steels, H.; James, S.A.; Roberts, I.N.; Stratford, M. Zygosaccharomyces Lentus: A Significant New Osmophilic, Preservative-Resistant Spoilage Yeast, Capable of Growth at Low Temperature. J. Appl. Microbiol. 1999, 87, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Liu, X.; Li, J.; Liu, S.; Zhang, H.; Bai, Y. Effects of Dielectric Barrier Discharge Plasma on the Inactivation of Zygosaccharomyces Rouxii and Quality of Apple Juice. Food Chem. 2018, 254, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Martorell, P.; Stratford, M.; Steels, H.; Fernández-Espinar, M.; Querol, A. Physiological Characterization of Spoilage Strains of Zygosaccharomyces Bailii and Zygosaccharomyces Rouxii Isolated from High Sugar Environments. Int. J. Food Microbiol. 2007, 114, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Dakal, T.; Solieri, L.; Giudici, P. Adaptive Response and Tolerance to Sugar and Salt Stress in the Food Yeast Zygosaccharomyces Rouxii. Int. J. Food Microbiol. 2014, 185, 140–157. [Google Scholar] [CrossRef] [PubMed]
- Leandro, M.J.; Sychrová, H.; Prista, C.; Loureiro-Dias, M.C. The Osmotolerant Fructophilic Yeast Zygosaccharomyces Rouxii Employs Two Plasma-Membrane Fructose Uptake Systems Belonging to a New Family of Yeast Sugar Transporters. Microbiology 2011, 157, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Pina, C.; Gonçalves, P.; Prista, C.; Loureiro-Dias, M.C. Ffz1, a New Transporter Specific for Fructose from Zygosaccharomyces Bailii. Microbiology 2004, 150, 2429–2433. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.L.; Wolfe, K.H. Recent Allopolyploid Origin of Zygosaccharomyces Rouxii Strain ATCC 42981. Yeast 2008, 25, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Rojo, M.C.; Torres Palazzolo, C.; Cuello, R.; González, M.; Guevara, F.; Ponsone, M.L.; Mercado, L.A.; Martínez, C.; Combina, M. Incidence of Osmophilic Yeasts and Zygosaccharomyces Rouxii during the Production of Concentrate Grape Juices. Food Microbiol. 2017, 64, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Rojo, M.C.; Arroyo López, F.N.; Lerena, M.C.; Mercado, L.; Torres, A.; Combina, M. Effects of pH and Sugar Concentration in Zygosaccharomyces Rouxii Growth and Time for Spoilage in Concentrated Grape Juice at Isothermal and Non-Isothermal Conditions. Food Microbiol. 2014, 38, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Hosono, K. Effect of Salt Stress on Lipid Composition and Membrane Fluidity of the Salttolerant Yeast Zygosaccharomyces Rouxii. J. Gen. Microbiol. 1992, 138, 91–96. [Google Scholar] [CrossRef]
- Wang, H.; Hu, Z.; Long, F.; Guo, C.; Yuan, Y.; Yue, T. Detection of Zygosaccharomyces rouxii and Candida tropicals in a High Sugar Medium by a Metal Oxide Sensor-Based Electronic Nose and Comparison with Test Panel Evaluation. J. Food Prot. 2015, 78, 2052–2063. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G. Yeast Spoilage of Foods and Beverages. In The Yeasts; Elsevier: Amsterdam, The Netherlands, 2010; pp. 53–63. [Google Scholar]
- Deak, T.; Beuchat, L.R. Yeasts Associated with Fruit Juice Concentrates. J. Food Prot. 1993, 56, 777–782. [Google Scholar] [CrossRef]
- Combina, M.; Daguerre, C.; Massera, A.; Mercado, L.; Sturm, M.E.; Ganga, A.; Martinez, C. Yeast Identification in Grape Juice Concentrates from Argentina. Lett. Appl. Microbiol. 2007, 46, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, Z.; Long, F.; Guo, C.; Yuan, Y.; Yue, T. Early Detection of Zygosaccharomyces Rouxii—Spawned Spoilage in Apple Juice by Electronic Nose Combined with Chemometrics. Int. J. Food Microbiol. 2016, 217, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, K.; Arakawa, E.; Kita, J.I.; Aoyama, Y.; Manome, Y.; Ikeda, K.; Yamamoto, K. Detection of Aeromonas Hydrophila in Liquid Media by Volatile Production Similarity Patterns, Using a FF-2A Electronic Nose. Sensors 2013, 13, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, E.; Falasconi, M.; Concina, I.; Mantero, G.; Bianchi, F.; Mattarozzi, M.; Musci, M.; Sberveglieri, G. Electronic Nose and Alicyclobacillus spp. Spoilage of Fruit Juices: An Emerging Diagnostic Tool. Food Control 2010, 21, 1374–1382. [Google Scholar] [CrossRef]
- Zhang, J.; Yue, T.; Yuan, Y. Alicyclobacillus Contamination in the Production Line of Kiwi Products in China. PLoS ONE 2013, 8, e67704. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Manzanares, N.; Huertas-Pérez, J.F.; Gámiz-Gracia, L.; García-Campaña, A.M. Simple and Efficient Methodology to Determine Mycotoxins in Cereal Syrups. Food Chem. 2015, 177, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Pearson, B.M.; McKee, R.A. Rapid Identification of Saccharomyces cerevisiae, Zygosaccharomyces bailii and Zygosaccharomyces rouxii. Int. J. Food Microbiol. 1992, 16, 63–67. [Google Scholar] [CrossRef]
- Kwon, H.; Yeo, E.; Hahn, S.; Bae, S.; Kim, D.; Byun, M. Cloning and Characterization of Genes Encoding Trehalose-6-Phosphate Synthase (TPS1) and Trehalose-6-Phosphate Phosphatase (TPS2) from Zygosaccharomyces rouxii. FEMS Yeast Res. 2003, 3, 433–440. [Google Scholar] [CrossRef]
- Iwaki, T.; Higashida, Y.; Tsuji, H.; Tamai, Y.; Watanabe, Y. Characterization of a Second Gene (ZSOD22) of Na+/H+ Antiporter from Salt-Tolerant Yeast Zygosaccharomyces rouxii and Functional Expression of ZSOD2 and ZSOD22 in Saccharomyces cerevisiae. Yeast 1998, 14, 1167–1174. [Google Scholar] [CrossRef]
- Jaramillo, M.C.; Huttener, M.; Alvarez, J.M.; Homs-Corbera, A.; Samitier, J.; Torrens, E.; Juárez, A. Dielectrophoresis Chips Improve PCR Detection of the Food-Spoiling Yeast Zygosaccharomyces rouxii in Apple Juice. Electrophoresis 2015, 36, 1471–1478. [Google Scholar] [CrossRef] [PubMed]
- Frisón, L.N.; Chiericatti, C.A.; Aríngoli, E.E.; Basílico, J.C.; Basílico, M.Z. Effect of Different Sanitizers against Zygosaccharomyces Rouxii. J. Food Sci. Technol. 2014, 52, 4619–4624. [Google Scholar] [CrossRef] [PubMed]
- Rojo, M.C.; Arroyo López, F.N.; Lerena, M.C.; Mercado, L.; Torres, A.; Combina, M. Evaluation of Different Chemical Preservatives to Control Zygosaccharomyces Rouxii Growth in High Sugar Culture Media. Food Control 2014, 50, 349–355. [Google Scholar] [CrossRef]
- Kusumegi, K.; Yoshida, H.; Tomiyama, S. Inhibitory Effects of Acetic Acid on Respiration and Growth of Zygosaccharomyces Rouxii. J. Ferment. Bioeng. 1998, 85, 213–217. [Google Scholar] [CrossRef]
- Kirimli, S.; Kunduhoglu, B. Inactivation of Zygosaccharomyces Rouxii Using Ultrasound at Different Temperatures, PH and Water Activity Conditions. Ital. J. Food Sci. 2016, 28, 64–72. [Google Scholar]
- Escott, C.; Loira, I.; Morata, A.; Bañuelos, M.; Suárez-Lepe, J. Wine Spoilage Yeasts: Control Strategy. In Yeast-Industrial Applications; Morata, A., Loira, I., Eds.; InTech: Rijeka, Croatia, 2017; pp. 89–116. [Google Scholar]
- Alonso, A.; Belda, I.; Santos, A.; Navascués, E.; Marquina, D. Advances in the Control of the Spoilage Caused by Zygosaccharomyces Species on Sweet Wines and Concentrated Grape Musts. Food Control 2015, 51, 129–134. [Google Scholar] [CrossRef]
- Enrique, M.; Ibáñez, A.; Marcos, J.F.; Yuste, M.; Martínez, M.; Vallés, S.; Manzanares, P. β-Glucanases as a Tool for the Control of Wine Spoilage Yeasts. J. Food Sci. 2010, 75, M41–M45. [Google Scholar] [CrossRef] [PubMed]
- Usseglio-Tomasset, L. Properties and Use of Sulphur Dioxide. Food Addit. Contam. 1992, 9, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Warth, A.D. Resistance of Yeast Species to Benzoic and Sorbic Acids and to Sulfur Dioxide. J. Food Prot. 1985, 48, 564–569. [Google Scholar] [CrossRef]
- Jansen, M.; Veurink, J.; Euverink, G. Growth of the Salt-Tolerant Yeast Zygosaccharomyces Rouxii in Microtiter Plates: Effects of NaCl, PH and Temperature on Growth and Fusel Alcohol Production From. FEMS Yeast Res. 2003, 3, 313–318. [Google Scholar] [PubMed]
- Solieri, L.; Cassanelli, S.; Giudici, P. A New Putative Zygosaccharomyces Yeast Species Isolated from Traditional Balsamic Vinegar. Yeast 2007, 24, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Solieri, L.; Giudici, P. Yeasts Associated to Traditional Balsamic Vinegar: Ecological and Technological Features. Int. J. Food Microbiol. 2008, 125, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Hecquet, L.; Sancelme, M.; Bolte, J.; Demuynck, C. Biosynthesis of 4-Hydroxy-2,5-Dimethyl-3(2H)-Furanone by Zygosaccharomyces Rouxii. J. Agric. Food Chem. 1996, 44, 1357–1360. [Google Scholar] [CrossRef]
- Saha, B.C.; Sakakibara, Y.; Cotta, M.A. Production of D-Arabitol by a Newly Isolated Zygosaccharomyces Rouxii. J. Ind. Microbiol. Biotechnol. 2007, 34, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, P.; Sabu, A.; Pandey, A.; Szakacs, G.; Soccol, C.R. Extra-Cellular l-Glutaminase Production by Zygosaccharomyces Rouxii under Solid-State Fermentation. Process Biochem. 2002, 38, 307–312. [Google Scholar] [CrossRef]
- Ok, T. Method of Utilization of Zygosaccharomyces Rouxii. U.S. Patent US20030219456A1, 21 May 2002. [Google Scholar]
- Cao, X.; Hou, L.; Lu, M.; Wang, C.; Zeng, B. Genome Shuffling of Zygosaccharomyces Rouxii to Accelerate and Enhance the Flavour Formation of Soy Sauce. J. Sci. Food Agric. 2010, 90, 281–285. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, G.; Turchetti, B.; Sileoni, V.; Marconi, O.; Perretti, G. Screening of New Strains of Saccharomycodes ludwigii and Zygosaccharomyces rouxii to Produce Low-Alcohol Beer. J. Inst. Brew. 2015, 121, 113–121. [Google Scholar] [CrossRef]
- Romano, P.; Suzzi, G. MINIREVIEW Origin and Production of Acetoin during Wine Yeast Fermentation. Appl. Environ. Microbiol. 1996, 62, 309–315. [Google Scholar] [PubMed]
- Romano, P.; Suzzi, G.; Zironi, R.; Comi, G. Biometric Study of Acetoin Production in Hanseniaspora guilliermondii and Kloeckera apiculata. Appl. Environ. Microbiol. 1993, 59, 1838–1841. [Google Scholar] [PubMed]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The Application of Non-Saccharomyces Yeast in Fermentations with Limited Aeration as a Strategy for the Production of Wine with Reduced Alcohol Content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage Yeasts in the Wine Industry. Int. J. Food Microbiol. 2003, 86, 23–50. [Google Scholar] [CrossRef]
- Romano, P.; Suzzi, G. Potential Use for Zygosaccharomyces Species in Winemaking. J. Wine Res. 1993, 4, 87–94. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a Future for Non-Saccharomyces Yeasts: Selection of Putative Spoilage Wine Strains to Be Used in Association with Saccharomyces cerevisiae for Grape Juice Fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Nor Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
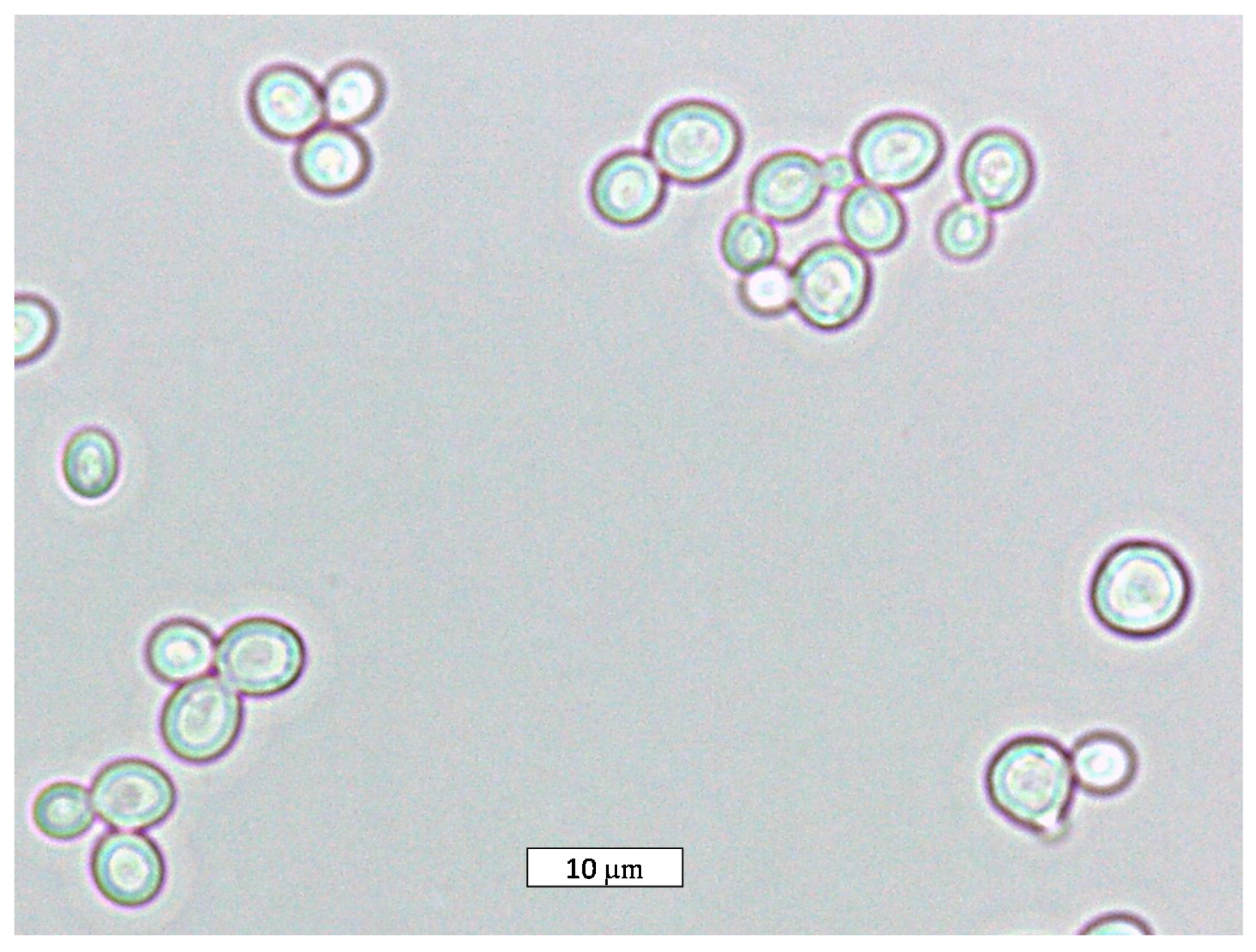


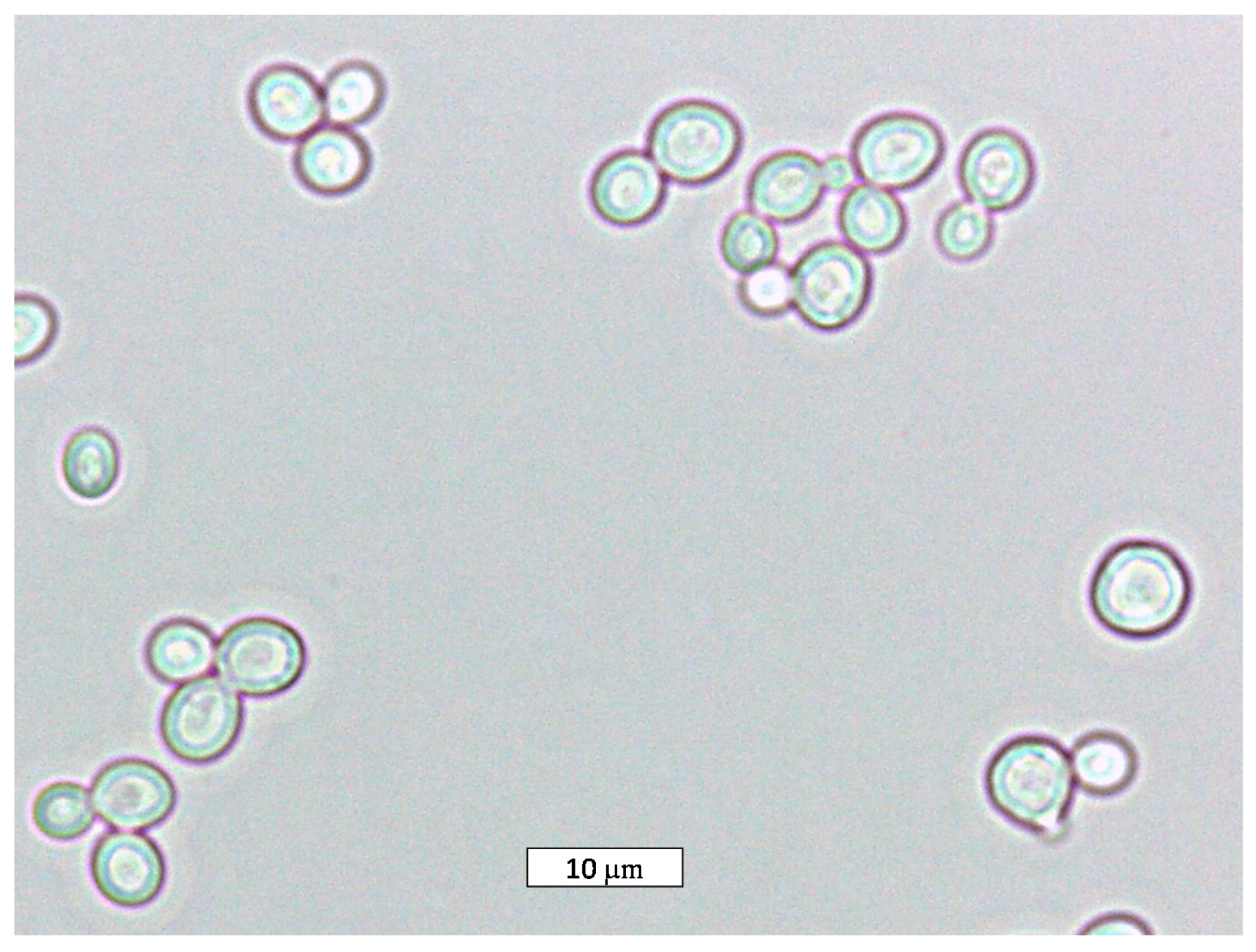
{kind=link}
{kind=link}
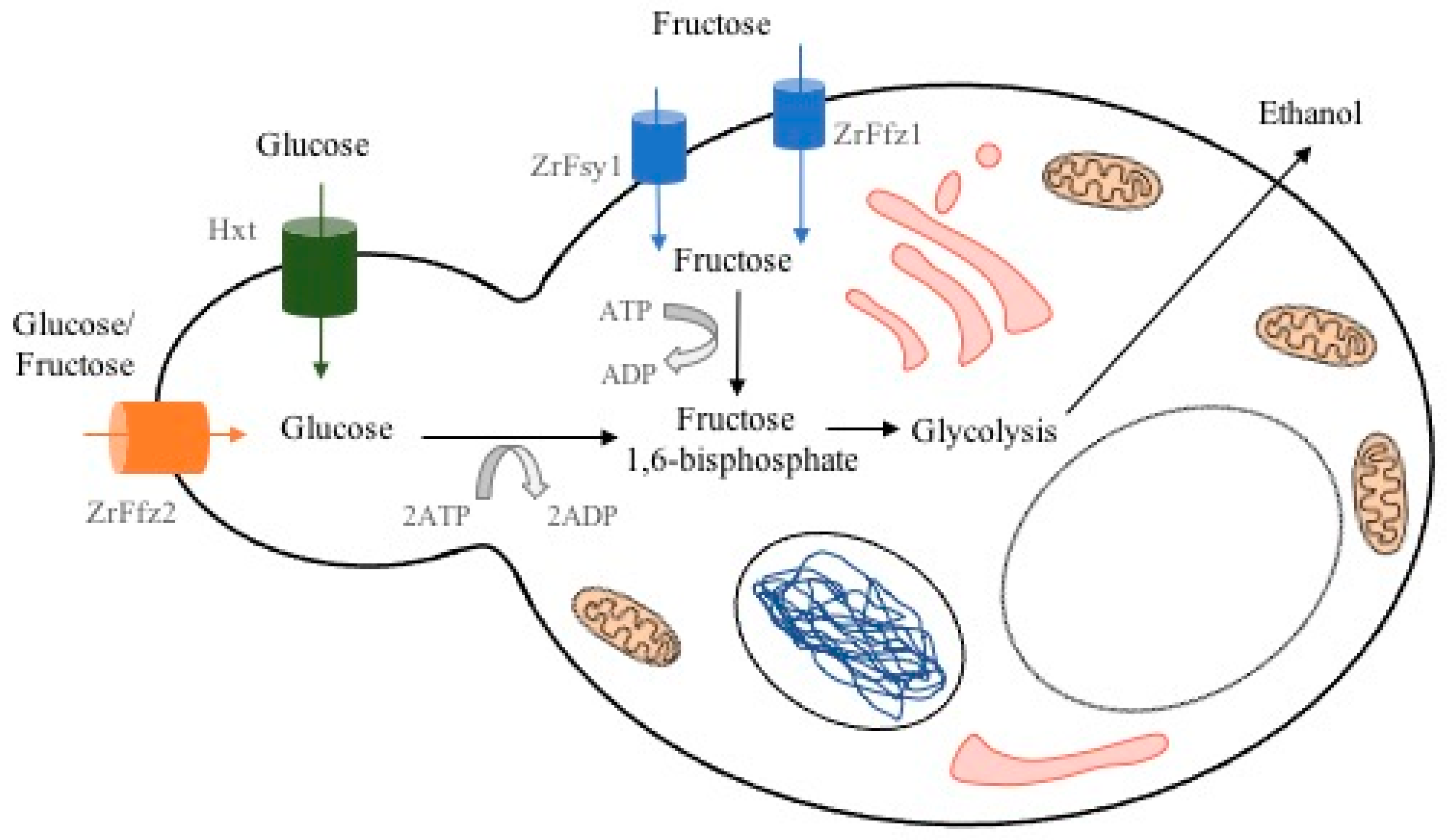{kind=link}
{kind=link}
| Yeast Strain | Ferulic Acid | p-Coumaric Acid | Potassium Sorbate | Sodium Benzoate | Sorbic Acid | Benzoic Acid | Acetic Acid | Cinnamic Acid | Ethanol | SO2 | DMDC | Peracetic Acid | Sodium Hypochlorite | H2O2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (mg/L) | (mg/L) | (mg/L) | (mg/L) | (mg/L) | (mg/L) | (g/L) | (mg/L) | (mg/L) | (mg/L) | (mg/L) | (mg/L) | (mg/L) | (mg/L) | |
| CECT 12003 1 | - | - | - | - | 320 | > 439.6 | 6.6 | > 446.7 | 73.7 | 217 | 227.9 | 247.2 | 204.7 | 459 |
| CECT 12004 1 | - | - | - | - | 314 | 372 | 6.4 | 369.6 | 78.3 | 262 | 227.9 | 247.2 | 189.8 | 544 |
| MC8, MC9, MC10 2 | 229 | 707 | 28.5 | 33.8 | - | - | - | - | - | - | 6.03 | - | - | - |
| R1 3 | - | - | - | - | - | - | 26 | - | - | - | - | - | - | - |
| Z. rouxii4 | - | - | - | - | - | - | - | - | - | - | - | 169.6 | 55.1 | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escott, C.; Del Fresno, J.M.; Loira, I.; Morata, A.; Suárez-Lepe, J.A. Zygosaccharomyces rouxii: Control Strategies and Applications in Food and Winemaking. Fermentation 2018, 4, 69. https://doi.org/10.3390/fermentation4030069
Escott C, Del Fresno JM, Loira I, Morata A, Suárez-Lepe JA. Zygosaccharomyces rouxii: Control Strategies and Applications in Food and Winemaking. Fermentation. 2018; 4(3):69. https://doi.org/10.3390/fermentation4030069
Chicago/Turabian StyleEscott, Carlos, Juan Manuel Del Fresno, Iris Loira, Antonio Morata, and José Antonio Suárez-Lepe. 2018. "Zygosaccharomyces rouxii: Control Strategies and Applications in Food and Winemaking" Fermentation 4, no. 3: 69. https://doi.org/10.3390/fermentation4030069
APA StyleEscott, C., Del Fresno, J. M., Loira, I., Morata, A., & Suárez-Lepe, J. A. (2018). Zygosaccharomyces rouxii: Control Strategies and Applications in Food and Winemaking. Fermentation, 4(3), 69. https://doi.org/10.3390/fermentation4030069