A Future Place for Saccharomyces Mixtures and Hybrids in Wine Making

Abstract

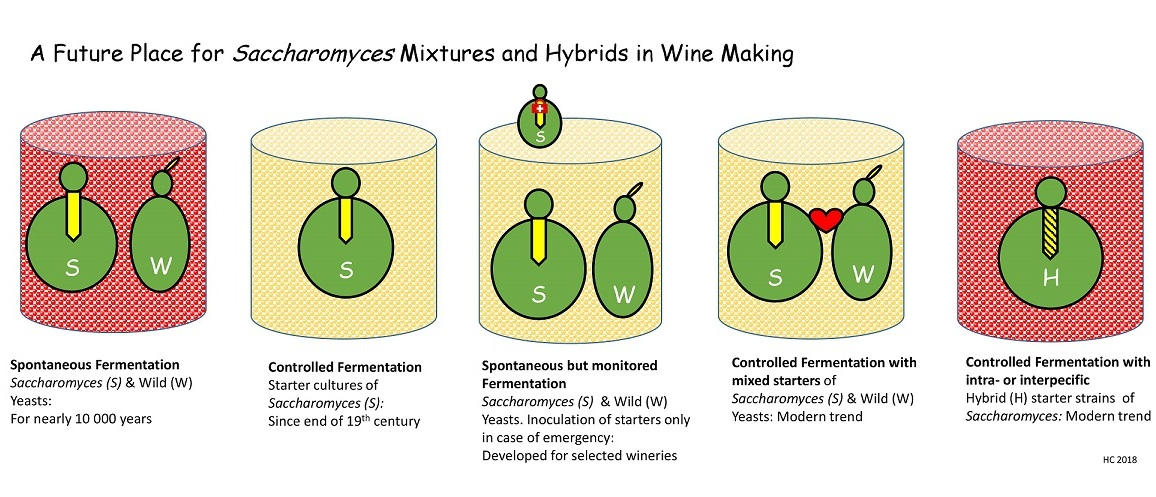{kind=link}
1. Introduction
2. Today’s Principal Procedures and Obstacles of Wine Making
3. Suggested Efforts for More Sophisticated Wine Production
3.1. Application of Wild Yeasts and Yeast Mixtures
3.2. Application of Hybrid Yeasts for Overcoming Fermentation Problems
4. Discussion
Conflicts of Interest
References
- McGovern, P.E. Acient Wine. The Search for the Origins of Viniculture; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar]
- McGovern, P.E.; Zhang, J.; Tang, J. Fermented beverages of pre- and proto-historic China. Proc. Natl. Acad. Sci. USA 2004, 101, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Kupfer, P. Weinstraße vor der Seidenstraße—Weinkulturen zwischen Georgien und China. In Kulturgut Rebe und Wein; König, H., Decker, H., Eds.; Springer Spektrum: Heidelberg, Germany, 2013; pp. 3–17. [Google Scholar]
- Kupfer, P. Der älteste Wein der Menschheit in China: Jiahu und die Suche nach den Ursprüngen der eurasischen Weinkultur. In Wo aber der Wein fehlt, stirbt der Reiz des Lebens; Decker, H., König, H., Zwickel, W., Eds.; Nünnerich-Asmus Verlag: Mainz, Germany, 2015; pp. 12–25. [Google Scholar]
- Gay-Lussac, J.L. Sur l’analyse de l’alcohol et de l’ether sulfurique et sur les produits de la fermentation. Annales de Chimie et de Physique 1815, 95, 311. [Google Scholar]
- Schwann, T. Vorläufige Mitteilung, betreffend Versuche über die Weingährung und Fäulnis. Annalen der Physik und Chemie 1837, 41, 184–193. [Google Scholar] [CrossRef]
- Kützing, F.T. Mikroskopische Untersuchungen über die Hefe und Esssigmutter, nebst mehreren anderen dazu gehörigen vegetabilischen Gebilden. J. Prakt. Chem. 1837, 11, 385–409. [Google Scholar] [CrossRef]
- Cagniard-Latour, C.C. Mémoire sur la fermentation vineuse. Ann. Chim. Phys. 1838, 68, 206–222. [Google Scholar]
- Pasteur, M.L. Animalcules infusoires vivant sans gaz oxygène libre et détermination des fermentation. Comptes Rendus de l’Academie des Sciences 1861, 52, 344–347. [Google Scholar]
- Feldmann, H. Yeast. Molecular and Cell Biology; Wiley-Blackwell: Weinheim, Germany, 2010. [Google Scholar]
- König, H.; Unden, G.; Fröhlich, J. (Eds.) Biology of Microorganisms on Grapes. In Must and in Wine, 2nd ed.; Springer: Heidelberg, Germany, 2017. [Google Scholar]
- Bisson, L.F.; Joseph, C.M.L.; Domizio, P. Yeasts. In Biology of Microorganisms on Grapes, in Must and in Wine, 2nd ed.; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Heidelberg, Germany, 2017; pp. 65–105. [Google Scholar]
- Fugelsang, K.C.; Edwards, C.G. Wine Microbiology. Practical Applications and Procedures; Springer: Heidelberg, Germany, 2007. [Google Scholar]
- Fleet, G.H.; Heard, G.M. Yeasts—Growth during fermentation. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic Publishers: Chur, Switzerland, 1993; pp. 27–54. [Google Scholar]
- König, H.; Berkelmann-Löhnertz, B. Maintenance of wine-associated microorganisms. In Biology of Microorganisms on Grapes, in Must and in Wine, 2nd ed.; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Heidelberg, Germany, 2017; pp. 549–571. [Google Scholar]
- König, H.; Fröhlich, J. Lactic acid bacteria. In Biology of Microorganisms on Grapes, in Must and in Wine, 2nd ed.; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Heidelberg, Germany, 2017; pp. 3–41. [Google Scholar]
- Guillamón, J.M.; Mas, A. Acetic acid bacteria. In Biology of Microorganisms on Grapes, in Must and in Wine, 2nd ed.; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Heidelberg, Germany, 2017; pp. 43–64. [Google Scholar]
- Mendes-Ferreira, A.; lí del Olmo, M.; Garcia-Martínez, J.; Pérez-Ortin, J. Functional genomics in wine yeast: DNA arrays and next generation sequencing. In Biology of Microorganisms on Grapes, in Must and in Wine, 2nd ed.; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Heidelberg, Germany, 2017; pp. 573–604. [Google Scholar]
- Fröhlich, J.; König, H.; Claus, H. Molecular methods for the identification of wine microorganisms and yeast development. In Biology of Microorganisms on Grapes, in Must and in Wine, 2nd ed.; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Heidelberg, Germany, 2017; pp. 517–547. [Google Scholar]
- Morrison-Whittle, P.; Goddard, M.R. From vineyard to winery: A source map of microbial diversity driving wine fermentation. Environ. Microbiol. 2018, 20, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Borneman, A.R.; Pretorius, I.S.; Chambers, P.J. Comparative genomics: a revolutionary tool for wine yeast strain development. Curr. Opin. Biotechnol. 2013, 24, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Bisson, L.F. Stuck and sluggish fermentations. Am. J. Enol. Vitic. 1999, 50, 1–13. [Google Scholar]
- Bisson, L.F.; Butzke, C.E. Diagnosis and rectification of stuck and sluggish fermentations. Am. J. Enol. Vitic. 2000, 51, 168–177. [Google Scholar]
- Heinisch, J.J.; Rodicio, R. Stress responses in wine yeast. In Biology of Microorganisms on Grapes, in Must and in Wine, 2nd ed.; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Heidelberg, Germany, 2017; pp. 377–395. [Google Scholar]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology. The Microbiology of Wine and Vinifications, 2nd ed.; John Wiley & Sons Ltd.: Chichester, UK, 2006; Volume 1, pp. 79–113. [Google Scholar]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial resources and enological significance: Opportunities and benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Mas, A.; Guillamón, J.M.; Beltran, G. Non-Conventional Yeast in the Wine Industry. Front. Media 2016, 7, 1494. [Google Scholar] [CrossRef]
- Visser, W.; Scheffers, W.A.; Batenburg-van der Vegte, A.H.; van Dijken, J.P. Oxygen requirements of yeasts. Appl. Environ. Microbiol. 1990, 56, 3785–3792. [Google Scholar] [PubMed]
- Hanl, L.; Sommer, P.; Arneborg, N. The effect of decreasing oxygen feed rates on growth and metabolism of Torulaspora delbrueckii. Appl. Microbiol. Biotechnol. 2005, 67, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Mauricio, J.C.; Millán, C.; Ortega, J.M. Influence of oxygen on the biosynthesis of cellular fatty acids, sterols and phospholipids during alcoholic fermentation by Saccharomyces cerevisiae and Torulaspora delbrueckii. World J. Microbiol. Biotechnol. 1998, 14, 405–410. [Google Scholar] [CrossRef]
- Brandam, C.; Lai, Q.P.; Julien-Ortiz, A.; Taillandier, P. Influence of oxygen on alcoholic fermentation by a wine strain of Torulaspory delbrueckii: Kinetics and carbon mass balance. Biosci. Biotechnol. Biochem. 2013, 77, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulou, M.; Lonvaud-Funel, A.; Dols-Lafargue, M. Polysaccharide production by grapes must and wine microorganisms. In Biology of Microorganisms on Grapes, in Must and in Wine, 2nd ed.; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Heidelberg, Germany, 2017; pp. 293–314. [Google Scholar]
- Sanz, L.M.; Martínez-Castro, I. Carbohydrates. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, M.C., Eds.; Springer: Heidelberg, Germany, 2010; pp. 231–248. [Google Scholar]
- Santos-Buelga, C.; de Freitas, V. Influence of phenolics on wine organoleptic properties. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, M.C., Eds.; Springer: Heidelberg, Germany, 2010; pp. 529–570. [Google Scholar]
- Sabel, A.; Claus, H.; Martens, S.; König, H. Wickerhamomyces anomalus AS1: A new strain with potential to improve wine aroma. Ann. Microbiol. 2014, 64, 483–491. [Google Scholar] [CrossRef]
- Schwentke, J.; Sabel, A.; Petri, A.; König, H.; Claus, H. The yeast Wickerhamomyces anomalus AS1 secretes a multifunctional exo-ß-1,3-glucanase with implications for winemaking. Yeast 2014, 31, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, H.H.; Großmann, M. Mikrobiologie des Weines. 4. aktualisierte Auflage; Ulmer Verlag: Stuttgart, Germany, 2010. [Google Scholar]
- Christ, E.; Kowalczyk, M.; Zuchowska, M.; Claus, H.; Löwenstein, R.; Szopinska-Morawska, A.; Renaut, J.; König, H. An exemplary model study for overcoming stuck fermentation during spontaneous fermentation with the aid of a Saccharomyces triple hybrid. J. Agric. Sci. 2015, 7, 18–34. [Google Scholar] [CrossRef]
- Szopinska, A.; Christ, E.; Planchon, S.; König, H.; Evers, D.; Renaut, J. Stuck at work? Quantitative proteomics of environmental wine yeast strains reveals the natural mechanism of overcoming stuck fermentation. Proteomics 2016, 16, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Blättel, V.; Petri, A.; Rabenstein, A.; Kuever, J.; König, H. Differentiation of species of the genus Saccharomyces using biomolecular fingerprinting methods. Appl. Microbiol. Biotechnol. 2013, 97, 4597–4606. [Google Scholar] [CrossRef] [PubMed]
- Jiranek, A.; Langridge, P.; Henschke, P.A. Amino acid and ammonium utilization by Saccharomyces cerevisiae wine yeasts from a chemically defined medium. Am. J. Enol. Vitic. 1995, 46, 75–83. [Google Scholar]
- Zuchowska, M.; König, H.; Claus, H. Allelic variants of hexose transporter Hxt3p und hexokinase Hxk1p/Hxk2p in strains of Saccharomyces cerevisiae und interspecies hybrids. Yeast 2015, 32, 657–669. [Google Scholar] [CrossRef] [PubMed]
- González, S.S.; Barrio, E.; Gafner, J.; Querol, A. Natural hybrids from Saccharomyces cerevisiae, Saccharomyces bayanus and Saccharomyces kudriavzevii in wine fermentations. FEMS Yeast Res. 2006, 6, 1221–1234. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, J.; Richards, K.; Niederer, H.; Lee, S.; Rod Dunbar, P.; Gardner, R. A homozygous diploidsubset of commercial wine yeast strains. Antonie van Leeuwenhoek 2006, 89, 27–37. [Google Scholar] [CrossRef] [PubMed]
- González, S.S.; Barrio, E.; Querol, A. Molecular characterization of new natural hybrids between Saccharomyces cerevisiae and Saccharomyces kudriavzevii from brewing. Appl. Environ. Microbiol. 2008, 74, 2314–2320. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.A.; Barrio, E.; Querol, A. Natural hybrids of S. cerevisiae × S. kudriavzevii share alleles with European wild populations of Saccharomyces kudriavzevii. FEMS Yeast Res. 2010, 10, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Naumov, G.I.; James, S.A.; Naumova, E.S.; Louis, E.J.; Roberts, I.N. Three new species in the Saccharomyces sensu stricto complex: Saccharomyces cariocanus, Saccharomyces kudriavzevii and Saccharomyces mikatae. Int. J. System. Evol. Microbiol. 2000, 50, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, J.P.; Gonçalves, P. Natural populations of Saccharomyces kudriavzevii in Portugal are associated with oak bark and sympatric with S. cerevisiae and S. paradoxus. Appl. Environ. Microbiol. 2008, 74, 2144–2152. [Google Scholar] [CrossRef] [PubMed]
- Hirschhäuser, S.; Fröhlich, J.; Gneipel, A.; Schönig, I.; König, H. Fast protocols for the 5S rDNA and ITS-2 based identification of Oenococcus oeni. FEMS Lett. 2005, 244, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, P.; Herr, P.; Fischer, U.; König, H. Molecular identification of lactic acid bacteria occurring in must and wine. South Afr. J. Enol. Vitic. 2011, 32, 300–309. [Google Scholar] [CrossRef]
- Röder, C.; König, H.; Fröhlich, J. Species-specific identification of Dekkera/Brettanomyces yeasts by fluorescently labelled rDNA probes targeting the 26S rRNA. FEMS Yeast Res. 2007, 7, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Petri, A.; Pfannebecker, J.; Fröhlich, J.; König, H. Fast identification of wine related lactic acid bacteria by multiplex PCR. Food Microbiol. 2013, 33, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Chambers, P.J.; Pretorius, I.S. Fermenting knowledge: The history of winemaking, science and yeast research. EMBO Rep. 2010, 11, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of wine making. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Borneman, A.R.; Desany, B.A.; Riches, D.; Affourtit, J.P.; Forgan, A.H.; Pretorius, I.S.; Egholm, M.; Chambers, P.J. The genome sequence of the wine yeast VIN7 reveals an allotriploid hybrid genome with Saccharomyces cerevisiae and Saccharomyces kudriavzevii origins. FEMS Yeast Res. 2012, 12, 88–96. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
König, H.; Claus, H. A Future Place for Saccharomyces Mixtures and Hybrids in Wine Making. Fermentation 2018, 4, 67. https://doi.org/10.3390/fermentation4030067
König H, Claus H. A Future Place for Saccharomyces Mixtures and Hybrids in Wine Making. Fermentation. 2018; 4(3):67. https://doi.org/10.3390/fermentation4030067
Chicago/Turabian StyleKönig, Helmut, and Harald Claus. 2018. "A Future Place for Saccharomyces Mixtures and Hybrids in Wine Making" Fermentation 4, no. 3: 67. https://doi.org/10.3390/fermentation4030067
APA StyleKönig, H., & Claus, H. (2018). A Future Place for Saccharomyces Mixtures and Hybrids in Wine Making. Fermentation, 4(3), 67. https://doi.org/10.3390/fermentation4030067