Syngas Fermentation: A Microbial Conversion Process of Gaseous Substrates to Various Products


Abstract
:
1. Introduction to Syngas Fermentation
1.1. Energy Demand
1.2. Potential Resources
1.2.1. Biomass
1.2.2. Wastes
1.3. Syngas Production
1.4. Microbial Conversion of CO and H2
2. Chemoautotrophic Microbes
2.1. Species and Habitat
2.2. Structure
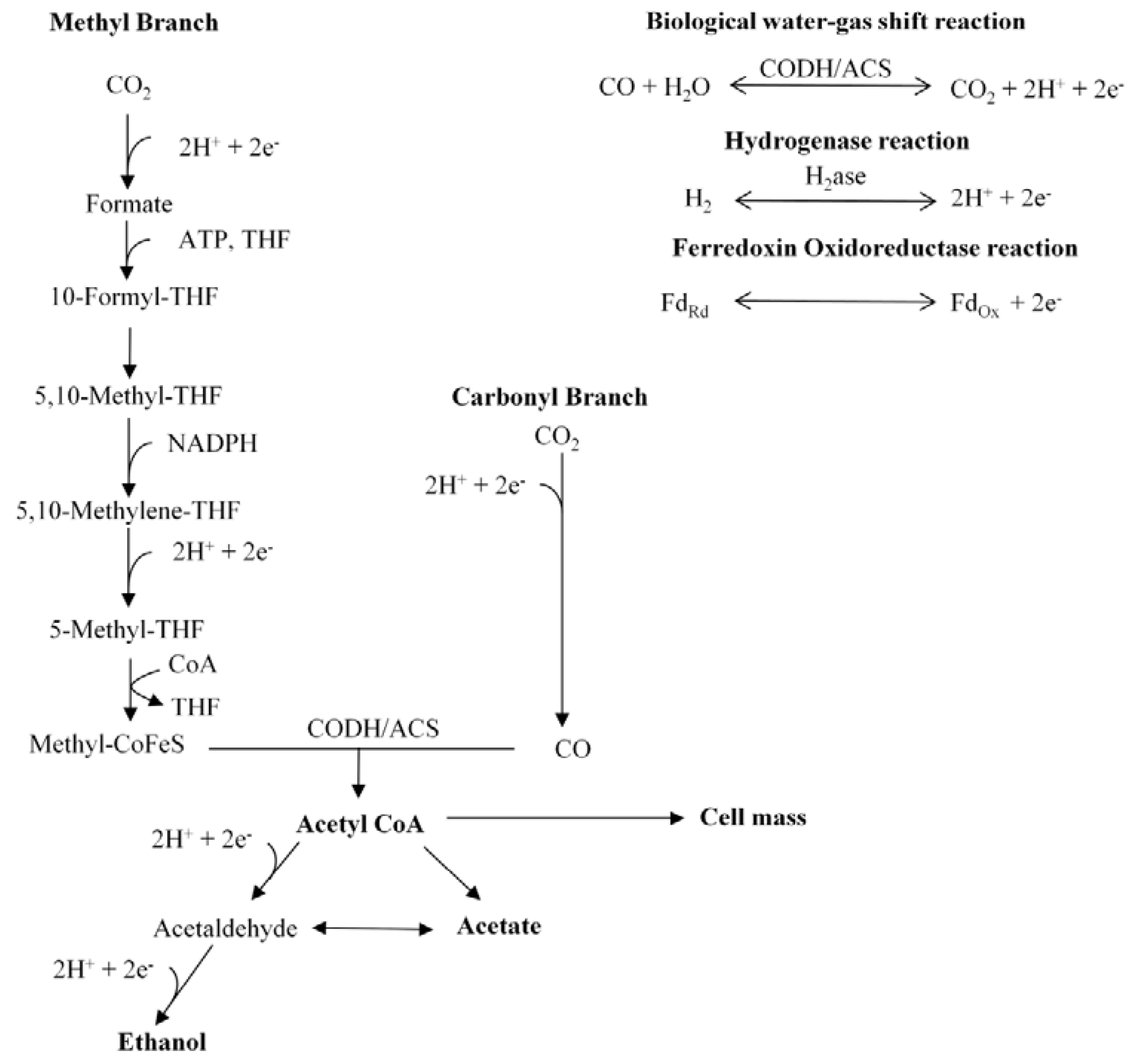
2.3. Pathway
2.3.1. Stoichiometry
2.3.2. Production Reactions
Carbon
Energy
2.3.3. Key Oxidation/Reduction Reactions in the Acetyl-CoA Pathway
2.4. ATP and Cell Growth
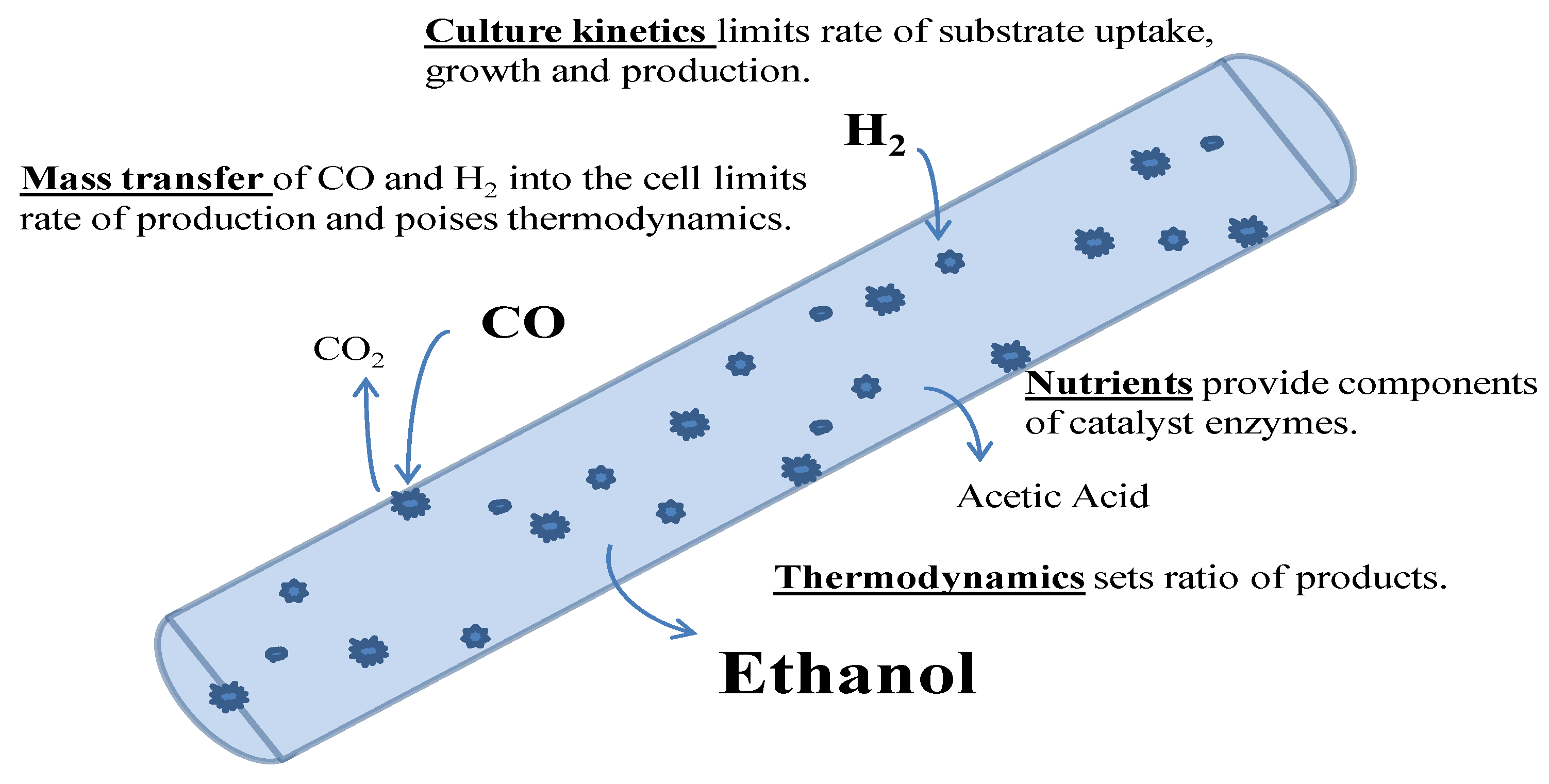
3. Microbial Conversion of Gas Phase Substrates
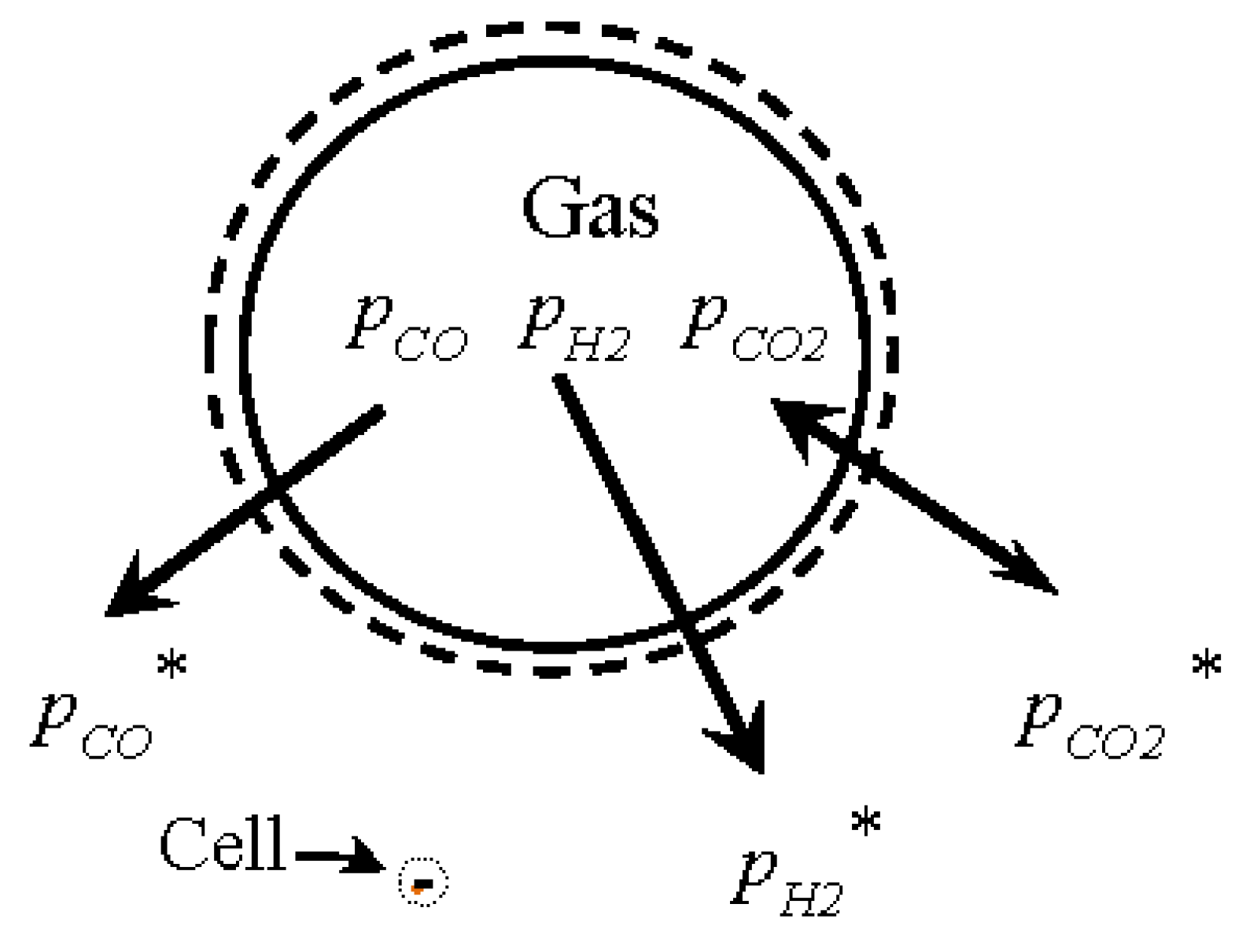
3.1. Gas Solubility
3.2. Transport Theory
3.3. Enzyme Catalyzed Reactions
3.4. Thermodynamics
3.5. Electrochemistry
3.6. Electron Carriers
3.7. Kinetics
3.8. Conceptual Model of Fermentation
3.9. Reactor Design
4. Potential Products
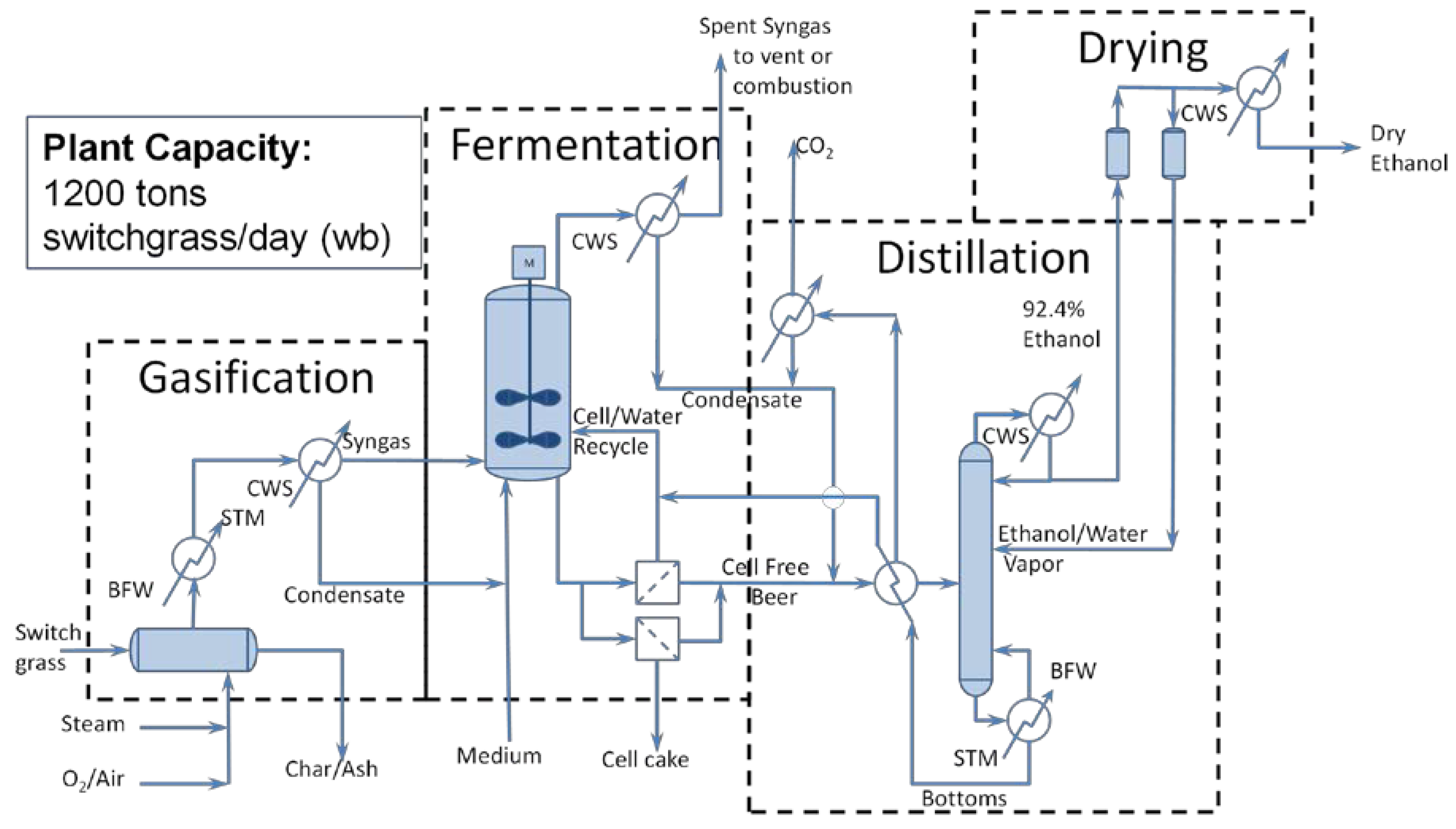
5. Techno-Economic Analysis
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wilkins, M.R.; Atiyeh, H.K. Microbial production of ethanol from carbon monoxide. Curr. Opin. Biotechnol. 2011, 22, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Drake, H.L.; Gossner, A.S.; Daniel, S.L. Old acetogens, new light. Ann. N. Y. Acad. Sci. 2008, 1125, 100–128. [Google Scholar] [CrossRef] [PubMed]
- Ramió-Pujol, S.; Ganigué, R.; Bañeras, L.; Colprim, J. How can alcohol production be improved in carboxydotrophic clostridia? Process Biochem. 2015, 50, 1047–1055. [Google Scholar] [CrossRef]
- Singla, A.; Verma, D.; Lal, B.; Sarma, P.M. Enrichment and optimization of anaerobic bacterial mixed culture for conversion of syngas to ethanol. Bioresour. Technol. 2014, 172, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Valle, C.R.; Perales, A.L.V.; Vidal-Barrero, F.; Gomez-Barea, A. Techno-economic assessment of biomass-to-ethanol by indirect fluidized bed gasification: Impact of reforming technologies and comparison with entrained flow gasification. Appl. Energy 2013, 109, 254–266. [Google Scholar] [CrossRef]
- Griffin, D.W.; Schultz, M.A. Fuel and chemical products from biomass syngas: A comparison of gas fermentation to thermochemical conversion routes. Environ. Progress Sustain. Energy 2012, 219–224. [Google Scholar] [CrossRef]
- Energy Information Administration (EIA). Annual Energy Outlook; Department of Energy: Washington, DC, USA, 2012.
- Energy Information Administration (EIA). International Energy Outlook; Department of Energy: Washington, DC, USA, 2013.
- Balan, V.; Chiaramonti, D.; Kumar, S. Review of US and EU initiatives toward development, demonstration, and commercialization of lignocellulosic biofuels. Biofuels Bioprod. Biorefin. 2013, 7, 732–759. [Google Scholar] [CrossRef]
- Energy Information Administration (EIA). Biofuels Issues and Trends; Department of Energy: Washington, DC, USA, 2012.
- Liew, F.M.; Köpke, M.; Simpson, S.D. Gas fermentation for commercial biofuels production. In Liquid, Gaseous and Solid Biofuels—Conversion Techniques; Fang, Z., Ed.; InTech: Rijeka, Croatia, 2013; pp. 125–173. [Google Scholar]
- Energy Information Administration (EIA). Annual Energy Outlook (US) with Projections to 2040; Department of Energy: Washington, DC, USA, 2015.
- Perlack, R.D.; Stokes, B.J. U.S. Billion-Ton Update: Biomass Supply for a Bioenergy and Bioproducts Industry; Department of Energy: Washington, DC, USA, 2011.
- Environmental Protection Agency. Municipal Solid Waste Generation, Recycling, and Disposal in the United States: Facts and Figures for 2012; Environmental Protection Agency: Washington, DC, USA, 2012.
- Atsonios, K.; Christodoulou, C.; Koytsoumpa, E.I.; Panopoulos, K.D.; Kakaras, E. Plant design aspects of catalytic biosyngas conversion to higher alcohols. Biomass Bioenergy 2013, 53, 54–64. [Google Scholar] [CrossRef]
- Ahmed, A.; Cateni, B.G.; Huhnke, R.L.; Lewis, R.S. Effects of biomass-generated producer gas constituents on cell growth, product distribution and hydrogenase activity of Clostridium carboxidivorans P7(T). Biomass Bioenergy 2006, 30, 665–672. [Google Scholar] [CrossRef]
- Xu, D.; Tree, D.R.; Lewis, R.S. The effects of syngas impurities on syngas fermentation to liquid fuels. Biomass Bioenergy 2011, 35, 2690–2696. [Google Scholar] [CrossRef]
- Woolcock, P.J.; Brown, R.C. A review of cleaning technologies for biomass-derived syngas. Biomass Bioenergy 2013, 52, 54–84. [Google Scholar] [CrossRef]
- Ferry, J.G.; House, C.H. The stepwise evolution of early life driven by energy conservation. Mol. Biol. Evol. 2006, 23, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Fischer, F.; Lieske, R.; Winzer, K. Biological gas reactions II concerning the formation of acetic acid in the biological conversion of carbon oxide and carbonic acid with hydrogen to methane. Biochem. Z. 1932, 245, 2–12. [Google Scholar]
- Wieringa, K.T. Over het verwijnenvan waterstofen koolzuur onder anaerobe voorwaarden. Antonie Leeuwenhoek 1936, 3, 263–273. [Google Scholar] [CrossRef]
- Collins, M.D.; Lawson, P.A.; Willems, A.; Cordoba, J.J.; Fernandezgarayzabal, J.; Garcia, P.; Cai, J.; Hippe, H.; Farrow, J.A.E. Thephylogeny of the genus Clostridium—Proposal of five new genera and eleven new species combinations. Int. J. Syst. Bacteriol. 1994, 44, 812–826. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, F.E. A new type of glucose fermentation by Clostridium thermoaceticum n. sp. J. Bacteriol. 1942, 43, 700–715. [Google Scholar]
- Ragsdale, S.W. Enzymology of the Wood-Ljungdahl pathway of acetogenesis. Ann. N. Y. Acad. Sci. 2008, 1125, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Barik, S.; Prieto, S.; Harrison, S.B.; Clausen, E.C.; Gaddy, J.L. Biological Production of Alcohols from Coal through Indirect Liquefaction. Appl. Biochem. Biotechnol. 1988, 18, 363–378. [Google Scholar] [CrossRef]
- Tanner, R.S.; Miller, L.M.; Yang, D. Clostridium ljungdahlii sp. nov., an Acetogenic Species in Clostridial rRNA Homology Group I. Int. J. Syste. Bacteriol. 1993, 43, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Worden, R.M.; Grethlein, A.J.; Jain, M.K.; Datta, R. Production of butanol and ethanol from synthesis gas via fermentation. Fuel 1991, 70, 615–619. [Google Scholar] [CrossRef]
- Abrini, J.; Naveau, H.; Nyns, E.-J. Clostridium autoethanogenum, sp. nov., an anaerobic bacterium that produces ethanol from carbon monoxide. Arch. Microbiol. 1994, 161, 345–351. [Google Scholar] [CrossRef]
- Liou, J.S.-C.; Balkwill, D.L.; Drake, G.R.; Tanner, R.S. Clostridium carboxidivorans sp. nov., a solvent-producing clostridium isolated from an agricultural settling lagoon, and reclassification of the acetogen Clostridium scatologenes strain SL1 as Clostridium drakei sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 2085–2091. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.R.; Atiyeh, H.K.; Tanner, R.S.; Torres, J.R.; Saxena, J.; Wilkins, M.R.; Huhnke, R.L. Butanol and hexanol production in Clostridium carboxidivorans syngas fermentation: Medium development and culture techniques. Bioresour. Technol. 2015, 190, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Saxena, J. Development of an optimized and cost-effective medium for ethanol production by Clostridium strain P11. In Botany and Microbiology; University of Oklahoma: Norman, OK, USA, 2008; p. 131. [Google Scholar]
- Allen, T.D.; Caldwell, M.E.; Lawson, P.A.; Huhnke, R.L.; Tanner, R.S. Alkalibaculum bacchi gen. nov., sp. nov., a CO-oxidizing, ethanol-producing acetogen isolated from livestock-impacted soil. Int. J. Syst. Evol. Microbiol. 2010, 60, 2483–2489. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Atiyeh, H.K.; Tanner, R.S.; Wilkins, M.R.; Huhnke, R.L. Fermentative production of ethanol from syngas using novel moderately alkaliphilic strains of Alkalibaculum bacchi. Bioresour. Technol. 2012, 104, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Atiyeh, H.K.; Stevenson, B.S.; Tanner, R.S.; Wilkins, M.R.; Huhnke, R.L. Continuous syngas fermentation for the production of ethanol, n-propanol and n-butanol. Bioresour. Technol. 2014, 151, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Atiyeh, H.K.; Stevenson, B.S.; Tanner, R.S.; Wilkins, M.R.; Huhnke, R.L. Mixed culture syngas fermentation and conversion of carboxylic acids into alcohols. Bioresour. Technol. 2014, 152, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Fu, B.; Zhang, L.; Liu, H. Bioconversion of H2/CO2 by acetogen enriched cultures for acetate and ethanol production: The impact of pH. World J. Microbiol. Biotechnol. 2015, 31, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Tanner, R.S. Cultivation of Bacteria and Fungi. In Manual of Environmental Microbiology, 3rd ed.; ASM Press: Washington, DC, USA, 2007; pp. 69–78. [Google Scholar]
- Maddipati, P.; Atiyeh, H.K.; Bellmer, D.D.; Huhnke, R.L. Ethanol production from syngas by Clostridium strain P11 using corn steep liquor as a nutrient replacement to yeast extract. Bioresour. Technol. 2011, 102, 6494–6501. [Google Scholar] [CrossRef] [PubMed]
- Saxena, J.; Tanner, R.S. Optimization of a corn steep medium for production of ethanol from synthesis gas fermentation by Clostridium ragsdalei. World J. Microbiol. Biotechnol. 2012, 28, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Kundiyana, D.K.; Huhnke, R.L.; Maddipati, P.; Atiyeh, H.K.; Wilkins, M.R. Feasibility of incorporating cotton seed extract in Clostridium strain P11 fermentation medium during synthesis gas fermentation. Bioresour. Technol. 2010, 101, 9673–9680. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.R.; Klasson, K.T.; Clausen, E.C.; Gaddy, J.L. Biological production of ethanol from coal synthesis gas - Medium development studies. Appl. Biochem. Biotechnol. 1993, 39, 559–571. [Google Scholar] [CrossRef]
- Cramer, W.A.; Knaff, D.B. Energy transduction in biological membranes: A textbook of bioenergetics. In Springer Advanced Texts in Chemistry; edn Springer Study; Springer-Verlag: New York, NY, USA, 1991. [Google Scholar]
- Roberts, J.R.; Lu, W.P.; Ragsdale, S.W. Acetyl-coenzyme A synthesis from methyltetrahydrofolate, CO, and coenzyme A by enzymes purified from Clostridium thermoaceticum: Attainment of in vivo rates and identification of rate-limiting steps. J. Bacteriol. 1992, 174, 4667–4676. [Google Scholar] [CrossRef] [PubMed]
- Ljungdahl, L.G. The Autotrophic Pathway of Acetate Synthesis in Acetogenic Bacteria. Annu. Rev. Microbiol. 1986, 40, 415–450. [Google Scholar] [CrossRef] [PubMed]
- Ragsdale, S.W.; Lindahl, P.A.; Münck, E. Mössbauer, EPR, and optical studies of the corrinoid/iron-sulfur protein involved in the synthesis of acetyl coenzyme A by Clostridium thermoaceticum. J. Biol. Chem. 1987, 262, 14289–14297. [Google Scholar] [PubMed]
- Yamamoto, I.; Saiki, T.; Liu, S.M.; Ljungdahl, L.G. Purification and properties of NADP-dependent formate dehydrogenase from Clostridium thermoaceticum, a tungsten-selenium-iron protein. J. Biol. Chem. 1983, 258, 1826–1832. [Google Scholar] [PubMed]
- Mejillano, M.R.; Jahansouz, H.; Matsunaga, T.O.; Kenyon, G.L.; Himes, R.H. Formation and utilization of formyl phosphate by N10-formyltetrahydrofolate synthetase: Evidence for formyl phosphate as an intermediate in the reaction. Biochemistry 1989, 28, 5136–5145. [Google Scholar] [CrossRef] [PubMed]
- Sun, A.Y.; Ljungdahl, L.; Wood, H.G. Total synthesis of acetate from CO2 II. Purification and properties of formyltetrahydrofolate synthetase from Clostridium thermoaceticum. J. Bacteriol. 1969, 98, 842–844. [Google Scholar] [PubMed]
- Ljungdahl, L.G.; O’Brien, W.E.; Moore, M.R.; Liu, M.-T. Methylenetetrahydrofolate dehydrogenase from Clostridium formicoaceticum and methylenetetrahydrofolate dehydrogenase, methenyltetrahydrofolate cyclohydrolase (combined) from Clostridium thermoaceticum. In Methods in Enzymology; Donald, B., McCormick, L.D.W., Eds.; Academic Press: Olrando, FL, USA, 1980; pp. 599–609. [Google Scholar]
- Park, E.Y.; Clark, J.E.; de Vartanian, D.V.; Ljungdahl, L.G. 5,10-methylenetetrahydrofolate Reductases: Iron-sulfur-zinc flavoproteins. In Chemistry and Biochemistry of flavoenzymes; Müller, F., Ed.; CRC Press: Boca Raton, FL, USA, 1991; pp. 389–400. [Google Scholar]
- Lu, W.P.; Harder, S.R.; Ragsdale, S.W. Controlled potential enzymology of methyl transfer reactions involved in acetyl-CoA synthesis by CO dehydrogenase and the corrinoid/iron-sulfur protein from Clostridium thermoaceticum. J. Biol. Chem. 1990, 265, 3124–3133. [Google Scholar] [PubMed]
- Drake, H.L.; Hu, S.I.; Wood, H.G. Purification of five components from Clostridium thermoaceticum which catalyze synthesis of acetate from pyruvate and methyltetrahydrofolate. Properties of phosphotransacetylase. J. Biol. Chem. 1981, 256, 11137–11144. [Google Scholar] [PubMed]
- White, H.; Strobl, G.; Feicht, R.; Simon, H. Carboxylic acid reductase: A new tungsten enzyme catalyses the reduction of non-activated carboxylic acids to aldehydes. Eur. J. Biochem. 1989, 184, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Fraisse, L.; Simon, H. Observations on the reduction of non-activated carboxylates by Clostridium formicoaceticum with carbon monoxide or formate and the Influence of various viologens. Arch. Microbiol. 1988, 150, 381–386. [Google Scholar] [CrossRef]
- Phillips, J.R.; Clausen, E.C.; Gaddy, J.L. Synthesis gas as substrate for the biological production of fuels and chemicals. Appl. Biochem. Biotechnol. 1994, 45, 145–157. [Google Scholar] [CrossRef]
- Latif, H.; Zeidan, A.A.; Nielsen, A.T.; Zengler, K. Trash to treasure: Production of biofuels and commodity chemicals via syngas fermenting microorganisms. Curr. Opin. Biotechnol. 2014, 27, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.; Martin, M.E.; Tappel, R.C.; Heijstra, B.D.; Mihalcea, C.; Köpke, M. Gas fermentation—A flexible platform for commercial scale production of low-carbon-fuels and chemicals from waste and renewable feedstocks. Front. Microbiol. 2016, 7, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Hurst, K.M.; Lewis, R.S. Carbon monoxide partial pressure effects on the metabolic process of syngas fermentation. Biochem. Eng. J. 2010, 48, 159–165. [Google Scholar] [CrossRef]
- Vega, J.L.; Prieto, S.; Elmore, B.B.; Clausen, E.C.; Gaddy, J.L. The Biological Production of Ethanol from Synthesis Gas. Appl. Biochem. Biotechnol. 1989, 20, 781–797. [Google Scholar] [CrossRef]
- Ukpong, M.N.; Atiyeh, H.K.; de Lorme, M.J.M.; Liu, K.; Zhu, X.; Tanner, R.S.; Wilkins, M.R.; Stevenson, B.S. Physiological response of Clostridium carboxidivorans during conversion of synthesis gas to solvents in a gas-fed bioreactor. Biotechnol. Bioeng. 2012, 109, 2720–2728. [Google Scholar] [CrossRef] [PubMed]
- Ezeji, T.; Milne, C.; Price, N.D.; Blaschek, H.P. Achievements and perspectives to overcome the poor solvent resistance in acetone and butanol-producing microorganisms. Appl. Microbiol. Biotechnol. 2010, 85, 1697–1712. [Google Scholar] [CrossRef] [PubMed]
- Köpke, M.; Mihalcea, C.; Bromley, J.C.; Simpson, S.D. Fermentative production of ethanol from carbon monoxide. Curr. Opin. Biotechnol. 2011, 22, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Seravalli, J.; Ragsdale, S.W. Channeling of carbon monoxide during anaerobic carbon dioxide fixation. Biochemistry 2000, 39, 1274–1277. [Google Scholar] [CrossRef] [PubMed]
- Isom, C.E.; Nanny, M.A.; Tanner, R.S. Improved conversion efficiencies for n-fatty acid reduction to primary alcohols by the solventogenic acetogen “Clostridium ragsdalei”. J. Ind. Microbiol. Biotechnol. 2015, 42, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, T.; Wood, H.G. Interaction of ferredoxin with carbon monoxide dehydrogenase from Clostridium thermoaceticum. J. Biol. Chem. 1992, 267, 897–900. [Google Scholar] [PubMed]
- Buckel, W.; Thauer, R.K. Energy conservation via electron bifurcating ferredoxin reduction and proton/Na+ translocating ferredoxin oxidation. Biochim. Biophys. Acta-Bioenerg. 2013, 1827, 94–113. [Google Scholar] [CrossRef] [PubMed]
- Poehlein, A.; Schmidt, S.; Kaster, A.K.; Goenrich, M.; Vollmers, J.; Thurmer, A.; Bertsch, J.; Schuchmann, K.; Voigt, B.; Hecker, M.; et al. An ancient pathway combining carbon dioxide fixation with the generation and utilization of a sodium ion gradient for ATP synthesis. PLoS ONE 2012, 7, e33439. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, P.L.; Zhang, T.; Dar, S.A.; Leang, C.; Lovley, D.R. The Rnf complex of Clostridium ljungdahlii is a proton-translocating ferredoxin:NAD+ oxidoreductase essential for autotrophic growth. mBio 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Schuchmann, K.; Muller, V. Autotrophy at the thermodynamic limit of life: A model for energy conservation in acetogenic bacteria. Nat. Rev. Microbiol. 2014, 12, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Hugenholtz, J.; Ivey, D.M.; Ljungdahl, L.G. Carbon monoxide-driven electron transport in Clostridium thermoautotrophicum membranes. J. Bacteriol. 1987, 169, 5845–5847. [Google Scholar] [CrossRef] [PubMed]
- Thauer, R.K.; Jungermann, K.; Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Microbiol. Mol. Biol. Rev. 1977, 41, 100–180. [Google Scholar]
- Das, A.; Ljungdahl, L.G. Composition and primary structure of the F1F0 ATP synthase from the obligately anaerobic bacterium Clostridium thermoaceticum. J. Bacteriol. 1997, 179, 3746–3755. [Google Scholar] [CrossRef] [PubMed]
- Ivey, D.M.; Ljungdahl, L.G. Purification and characterization of the F1-ATPase from Clostridium thermoaceticum. J. Bacteriol. 1986, 165, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, R.K.; Scanlon, J.A.B.; Al-Shawi, M.K. The rotary mechanism of the ATP synthase. Arch. Biochem. Biophys. 2008, 476, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Von Ballmoos, C.; Cook, G.M.; Dimroth, P. Unique rotary ATP synthase and its biological diversity. Annu. Rev. Biophys. 2008, 37, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Ragsdale, S.W. Enzymology of the acetyl-CoA pathway of CO2 fixation. Critical Rev. Biochem. Mol. Biol. 1991, 26, 261–300. [Google Scholar] [CrossRef] [PubMed]
- Hougen, O.A.; Watson, K.M.; Ragatz, R.A. Chemical Process Principles; Wiley: New York, NY, USA, 1954. [Google Scholar]
- Bird, R.B.; Stewart, W.E.; Lightfoot, E.N. Transport Phenomena, 2nd ed.; Wiley: New York, NY, USA, 2002. [Google Scholar]
- Charpentier, J.-C. Mass-transfer rates in gas-liquid absorbers and reactors. In Advances in Chemical Engineering; Drew, T.B., Cokelet, G.R., Hooper, J.W., Vermeulen, T, Eds.; Academic Press: Olrando, FL, USA, 1981; pp. 1–133. [Google Scholar]
- Bailey, J.E.; Ollis, D.F. Biochemical Engineering Fundamentals; McGraw-Hill: New York, NY, USA, 1986. [Google Scholar]
- McCabe, W.L.; Smith, J.C. Unit Operations of Chemical Engineering; McGraw-Hill: New York, NY, USA, 1976. [Google Scholar]
- Munasinghe, P.C.; Khanal, S.K. Syngas fermentation to biofuel: Evaluation of carbon monoxide mass transfer coefficient (kLa) in different reactor configurations. Biotechnol. Prog. 2010, 26, 1616–1621. [Google Scholar] [CrossRef] [PubMed]
- Klasson, K.T.; Ackerson, M.D.; Clausen, E.C.; Gaddy, J.L. Mass transport in bioreactors for coal synthesis gas fermentation. Abstr. Pap. Am. Chem. Soc. 1992, 1924–1930. [Google Scholar]
- Shen, Y.W.; Brown, R.; Wen, Z.Y. Enhancing mass transfer and ethanol production in syngas fermentation of Clostridium carboxidivorans P7 through a monolithic biofilm reactor. Appl. Energy 2014, 136, 68–76. [Google Scholar] [CrossRef]
- Vega, J.L.; Antorrena, G.M.; Clausen, E.C.; Gaddy, J.L. Study of gaseous substrate fermentations: Carbon monoxide conversion to acetate. 2. continuous culture. Biotechnol. Bioeng. 1989, 34, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Brown, R.; Wen, Z. Syngas fermentation of Clostridium carboxidivoran P7 in a hollow fiber membrane biofilm reactor: evaluating the mass transfer coefficient and ethanol production performance. Biochem. Eng. J. 2014, 85, 21–29. [Google Scholar] [CrossRef]
- Orgill, J.J.; Atiyeh, H.K.; Devarapalli, M.; Phillips, J.R.; Lewis, R.S.; Hunke, R.L. A comparison of mass transfer coefficients between trickle bed, hollow fiber membrane and stirred tank reactors. Bioresour. Technol. 2013, 133, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, D.G.; Ferguson, S.J. Bioenergetics; Academic Press: Olrando, FL, USA, 2002. [Google Scholar]
- Hu, P.; Bowen, S.H.; Lewis, R.S. A thermodynamic analysis of electron production during syngas fermentation. Bioresour. Technol. 2011, 102, 8071–8076. [Google Scholar] [CrossRef] [PubMed]
- Lehninger, A.L. Principles of Biochemistry; Worth Publishers: New York, NY, USA, 1982. [Google Scholar]
- Bar-Even, A. Does acetogenesis really require especially low reduction potential? Biochim. Biophys. Acta 2013, 1827, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Shuler, M.L.; Kargi, F. Bioprocess Engineering Basic Concepts; Prentice Hall PTR: Upper Saddle River, NJ, USA, 2002. [Google Scholar]
- Chen, J.; Gomez, J.A.; Hoffner, K.; Barton, P.I.; Henson, M.A. Metabolic modeling of synthesis gas fermentation in bubble column reactors. Biotechnol. Biofuels 2015, 8, 1–12. [Google Scholar] [CrossRef]
- Krivoruchko, A.; Zhang, Y.; Siewers, V.; Chen, Y.; Nielsen, J. Microbial acetyl-CoA metabolism and metabolic engineering. Metab. Eng. 2015, 28, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Bredwell, M.D.; Srivastava, P.; Worden, R.M. Reactor design issues for synthesis-gas fermentations. Biotechnol. Prog. 1999, 15, 834–844. [Google Scholar] [CrossRef] [PubMed]
- Rogers, P.; Chen, J.-S.; Zidwick, M.J. Organic acid and solvent production. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 511–755. [Google Scholar]
- Straub, M.; Demler, M.; Weuster-Botz, D.; Durre, P. Selective enhancement of autotrophic acetate production with genetically modified Acetobacterium woodii. J. Biotechnol. 2014, 178, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Klasson, K.T.; Ackerson, C.M.D.; Clausen, E.C.; Gaddy, J.L. Biological conversion of synthesis gas into fuels. Int. J. Hydrogen Energy 1992, 17, 281–288. [Google Scholar] [CrossRef]
- Atiyeh, H.K.; Phillips, J.R.; Huhnke, R.L. Fermentation Control for Optimization of Syngas Utilization. WO2016077778 A1, 2016. [Google Scholar]
- Kundiyana, D.K.; Huhnke, R.L.; Wilkins, M.R. Effect of nutrient limitation and two-stage continuous fermentor design on productivities during “Clostridium ragsdalei” syngas fermentation. Bioresour. Technol. 2011, 102, 6058–6064. [Google Scholar] [CrossRef] [PubMed]
- Devarapalli, M.; Atiyeh, H.K.; Phillips, J.R.; Lewis, R.S.; Huhnke, R.L. Ethanol production during semi-continuous syngas fermentation in a trickle bed reactor using Clostridium ragsdalei. Bioresour. Technol. 2016, 209, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Devarapalli, M.; Lewis, R.S.; Atiyeh, H.K. Continuous ethanol production from synthesis gas by Clostridium ragsdalei in a trickle-bed reactor. Fermentation 2017, 3, 1–13. [Google Scholar] [CrossRef]
- Richter, H.; Martin, M.E.; Angenent, L.T. A two-stage continuous fermentation system for conversion of syngas into ethanol. Energies 2013, 6, 3987–4000. [Google Scholar] [CrossRef]
- Nguyen, T.T.N.; Belliere-Baca, V.; Rey, P.; Millet, J.M.M. Efficient catalysts for simultaneous dehydration of light alcohols in gas phase. Catal. Sci. Technol. 2015, 5, 3576–3584. [Google Scholar] [CrossRef]
- Kopke, M.; Mihalcea, C.; Liew, F.M.; Tizard, J.H.; Ali, M.S.; Conolly, J.J.; Al-Sinawi, B.; Simpson, S.D. 2,3-Butanediol production by acetogenic bacteria, an alternative route to chemical synthesis, using industrial waste gas. Appl. Environ. Microbiol. 2011, 77, 5467–5475. [Google Scholar] [CrossRef] [PubMed]
- Dürre, P. Butanol formation from gaseous substrates. FEMS Microbiol. Lett. 2016, 363, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Chakraborty, S.; Kumar, A.; Woolston, B.; Liu, H.; Emerson, D.; Stephanopoulos, G. Integrated bioprocess for conversion of gaseous substrates to liquids. Proc. Natl. Acad. Sci. USA 2016, 113, 3773–3778. [Google Scholar] [CrossRef] [PubMed]
- Pakpour, F.; Najafpour, G.; Tabatabaei, M.; Tohidfar, M.; Younesi, H. Biohydrogen production from CO-rich syngas via a locally isolated Rhodopseudomonas palustris PT. Bioprocess Biosyst. Eng. 2014, 37, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Youngsukkasem, S.; Chandolias, K.; Taherzadeh, M.J. Rapid bio-methanation of syngas in a reverse membrane bioreactor: Membrane encased microorganisms. Bioresour. Technol. 2015, 178, 334–340. [Google Scholar] [CrossRef] [PubMed]
- EIA: 4-Week average U.S. Oxygenate Plant Production of Fuel Tthanol (Thousand Barrels Per Day); Energy Department of Energy Information Administration: Washington, DC, USA, 2017. [Google Scholar]
- Renewable Fuels Association (RFA). World Fuel Ethanol Production; Renewable Fuels Association: Washington, DC, USA, 2016. [Google Scholar]
- Nasdaq. Ethanol Futures. 2017. Available online: http://www.nasdaq.com/markets/ethanol.aspx (accessed on 27 April 2017).
- USDA NASS, Quick Stats Database. USDA ERS—U.S. Bioenergy Statistics: Fuel Ethanol, Corn and Gasoline Prices; Service UER. USDA Economic Research Service: Washington, DC, USA, 2017. [Google Scholar]
- Klein-Marcuschamer, D.; Oleskowicz-Popiel, P.; Simmons, B.A.; Blanch, H.W. Technoeconomic analysis of biofuels: A wiki-based platform for lignocellulosic biorefineries. Biomass Bioenergy 2010, 34, 1914–1921. [Google Scholar] [CrossRef]
- Humbird, D.; Davis, R.; Tao, L.; Kinchin, C.; Hsu, D.; Aden, A.; Schoen, P.; Lukas, J.; Olthof, B.; Worley, M. Process Design and Economics for Biochemical Conversion of Lignocellulosic Biomass to Ethanol: Dilute-Acid Pretreatment and Enzymatic Hydrolysis of Corn Stover; National Renewable Energy Laboratory (NREL): Golden, CO, USA, 2011.
- Dwivedi, P.; Alavalapati, J.R.; Lal, P. Cellulosic ethanol production in the United States: Conversion technologies, current production status, economics, and emerging developments. Energy Sustain. Dev. 2009, 13, 174–182. [Google Scholar] [CrossRef]
- Piccolo, C.; Bezzo, F. A techno-economic comparison between two technologies for bioethanol production from lignocellulose. Biomass Bioenergy 2009, 33, 478–491. [Google Scholar] [CrossRef]
- Anex, R.P.; Aden, A.; Kazi, F.K.; Fortman, J.; Swanson, R.M.; Wright, M.M.; Satrio, J.A.; Brown, R.C.; Daugaard, D.E.; Platon, A. Techno-economic comparison of biomass-to-transportation fuels via pyrolysis, gasification, and biochemical pathways. Fuel 2010, 89, S29–S35. [Google Scholar] [CrossRef]
- Dutta, A.; Talmadge, M.; Hensley, J.; Worley, M.; Dudgeon, D.; Barton, D.; Groenendijk, P.; Ferrari, D.; Stears, B.; Searcy, E. Techno-economics for conversion of lignocellulosic biomass to ethanol by indirect gasification and mixed alcohol synthesis. Environ. Progress Sustain. Energy 2012, 31, 182–190. [Google Scholar] [CrossRef]
- Spath, P.L.; Dayton, D.C. Preliminary Screening-Technical and Economic Assessment of Synthesis Gas to Fuels and Chemicals with Emphasis on the Potential for Biomass-Derived Syngas. 2003. Available online: http://oai.dtic.mil/oai/oai?verb=getRecord&metadataPrefix=html&identifier=ADA436529 (accessed on 27 April 2017).
- Brown, T.R. A techno-economic review of thermochemical cellulosic biofuel pathways. Bioresour. Technol. 2015, 178, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Kazi, F.K.; Fortman, J.A.; Anex, R.P.; Hsu, D.D.; Aden, A.; Dutta, A.; Kothandaraman, G. Techno-economic comparison of process technologies for biochemical ethanol production from corn stover. Fuel 2010, 89, S20–S28. [Google Scholar] [CrossRef]
- Tanner, R.S. Production of ethanol from synthesis gas. In Bioenergy; Wall, J.D., Harwood, C.S., Demain, A.L., Eds.; American Society of Microbiology: Washington, DC, USA, 2008; pp. 147–151. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Reaction | Reference |
|---|---|---|
| Carbon Monoxide dehydrogenase | [43] | |
| Hydrogenase | [44] | |
| Ferredoxin oxidoreductase | [45] | |
| Formate dehydrogenase | [46] | |
| Formate kinase | [47] | |
| Formyl THF synthetase 1 | [48] | |
| Methenyl THF cyclohydrolase | [44] | |
| Methylene THF dehydrogenase | [49] | |
| Methylene THF reductase | [50] | |
| Methyl transferase | [44] | |
| Corrinoid-Iron-Sulfur protein | [51] | |
| Acetyl-CoA synthase | [24] | |
| Phosphotransacetylase | [52] | |
| Acetate kinase | [52] | |
| Aldehyde dehydrogenase | [53] | |
| Alcohol dehydrogenase | [54] |
| Products | Reaction | ΔG° kJ/mol | |
|---|---|---|---|
| Acetic Acid | (1) | −154.6 | |
| (2) | −134.5 | ||
| (3) | −114.5 | ||
| (4) | −94.4 | ||
| (5) | −74.3 | ||
| Ethanol | (6) | −217.4 | |
| (7) | −197.3 | ||
| (8) | −177.3 | ||
| (9) | −157.2 | ||
| (10) | −137.1 | ||
| (11) | −117.1 | ||
| (12) | −97.0 | ||
| Acetic Acid | (13) | −94.4 | |
| Ethanol | (14) | −117.1 | |
| Half Cell Reduction | ΔGr° (kJ/mol) | E° (mV) | ne | ΔmH | ΔGr°′ (kJ/mol) | E°′ (mV) | Πprod/Πreact |
|---|---|---|---|---|---|---|---|
| 2H+ + 2e− ↔ H2(g) | 0 | 0 | 2 | −2 | 79.90 | −414 | pH2 |
| CO2 + 2H+ + 2e− ↔ CO(g) + H2O | 20.03 | −104 | 2 | −2 | 99.93 | −518 | pCO/pCO2 |
| CH3COOH + 2H+ + 2e− ↔ CH3CHO + H2O | −7.67 | 40 | 2 | −2 | 72.23 | −374 | CAld/CHA |
| CH3CHO + 2H+ + 2e− ↔ CH3CH2OH | −41.85 | 217 | 2 | −2 | 38.05 | −197 | CEt/CAld |
| NAD+ + H+ + 2e− ↔ NADH | 21.80 | −113 | 2 | −1 | 61.75 | −320 | CNADH/CNAD+ |
| FdOx + 2e− ↔ FdRd | 81.05 | −420 | 2 | 0 | 81.05 | −420 | CFdr/CFdo |
| Gas | H (kPa L/mol) | Di,W (m2/s) |
|---|---|---|
| CO | 121,561 | 2.50 × 10−9 |
| H2 | 140,262 | 6.24 × 10−9 |
| CO2 | 4240 | 2.69 × 10−9 |
| O2 | 101,300 | 3.25 × 10−9 |
| Process a | Plant Size (MTPD) b | Feedstock Cost ($/mT) | Ethanol Yield (L/mT) | TCI c (M$) | MESP d ($/L) | Reference |
|---|---|---|---|---|---|---|
| GF | 2206 | 38.70 | 289 | NR e | 0.44 | [120] |
| 2030 | 80.13 | 204 | 575 | 1.32 f | [117] | |
| 2030 | 80.13 | 282 | NR | 1.07 g | ||
| GMA | 2140 | 88.74 | 236 | 578 | 0.86 | [5] |
| 2000 | 78.06 | 350 | 593 | 0.62 | [119] | |
| EHF | 2000 | 74.17 | 330 | 509 | 0.65 | [115] |
| 2030 | 80.13 | 310 | 301 | 1.01 h | [117] | |
| 2000 | 95.45 | 289 | 432 | 1.03 i | [122] | |
| 2000 | 95.45 | 250 | 444 | 1.11 j |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phillips, J.R.; Huhnke, R.L.; Atiyeh, H.K. Syngas Fermentation: A Microbial Conversion Process of Gaseous Substrates to Various Products. Fermentation 2017, 3, 28. https://doi.org/10.3390/fermentation3020028
Phillips JR, Huhnke RL, Atiyeh HK. Syngas Fermentation: A Microbial Conversion Process of Gaseous Substrates to Various Products. Fermentation. 2017; 3(2):28. https://doi.org/10.3390/fermentation3020028
Chicago/Turabian StylePhillips, John R., Raymond L. Huhnke, and Hasan K. Atiyeh. 2017. "Syngas Fermentation: A Microbial Conversion Process of Gaseous Substrates to Various Products" Fermentation 3, no. 2: 28. https://doi.org/10.3390/fermentation3020028
APA StylePhillips, J. R., Huhnke, R. L., & Atiyeh, H. K. (2017). Syngas Fermentation: A Microbial Conversion Process of Gaseous Substrates to Various Products. Fermentation, 3(2), 28. https://doi.org/10.3390/fermentation3020028