
Microbiota Diversity During Grape Drying and Spontaneous Fermentations of Vin Santo

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Winemaking
2.2. Microbial Analysis
2.3. Microbial Identification and Typing of Yeast Strains
2.4. Chemical Analysis
2.5. Phenolic Compounds and Ochratoxin Content Determination
2.6. Statistical Analysis
3. Results
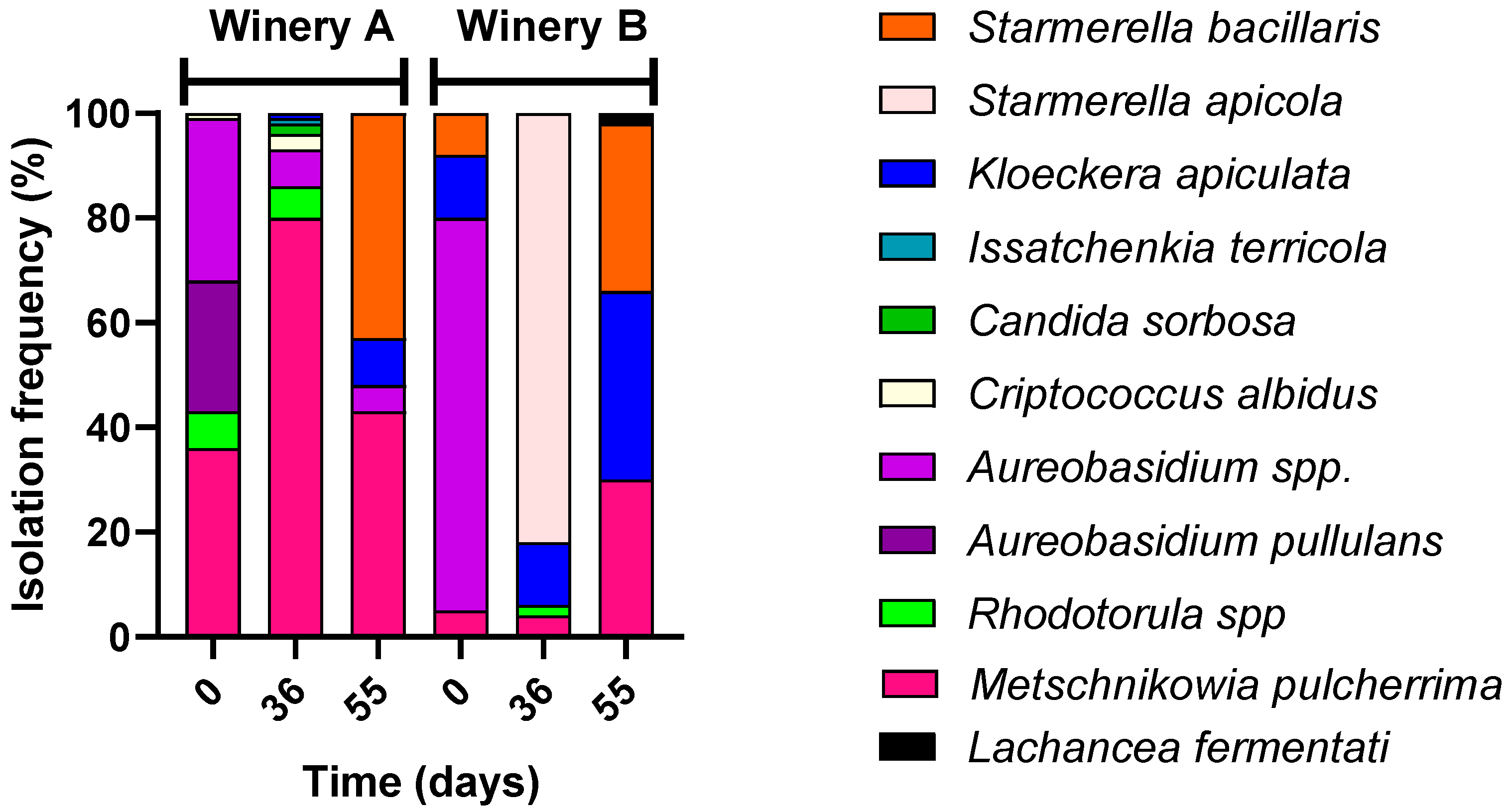
3.1. Microbiota of Grapes During Drying
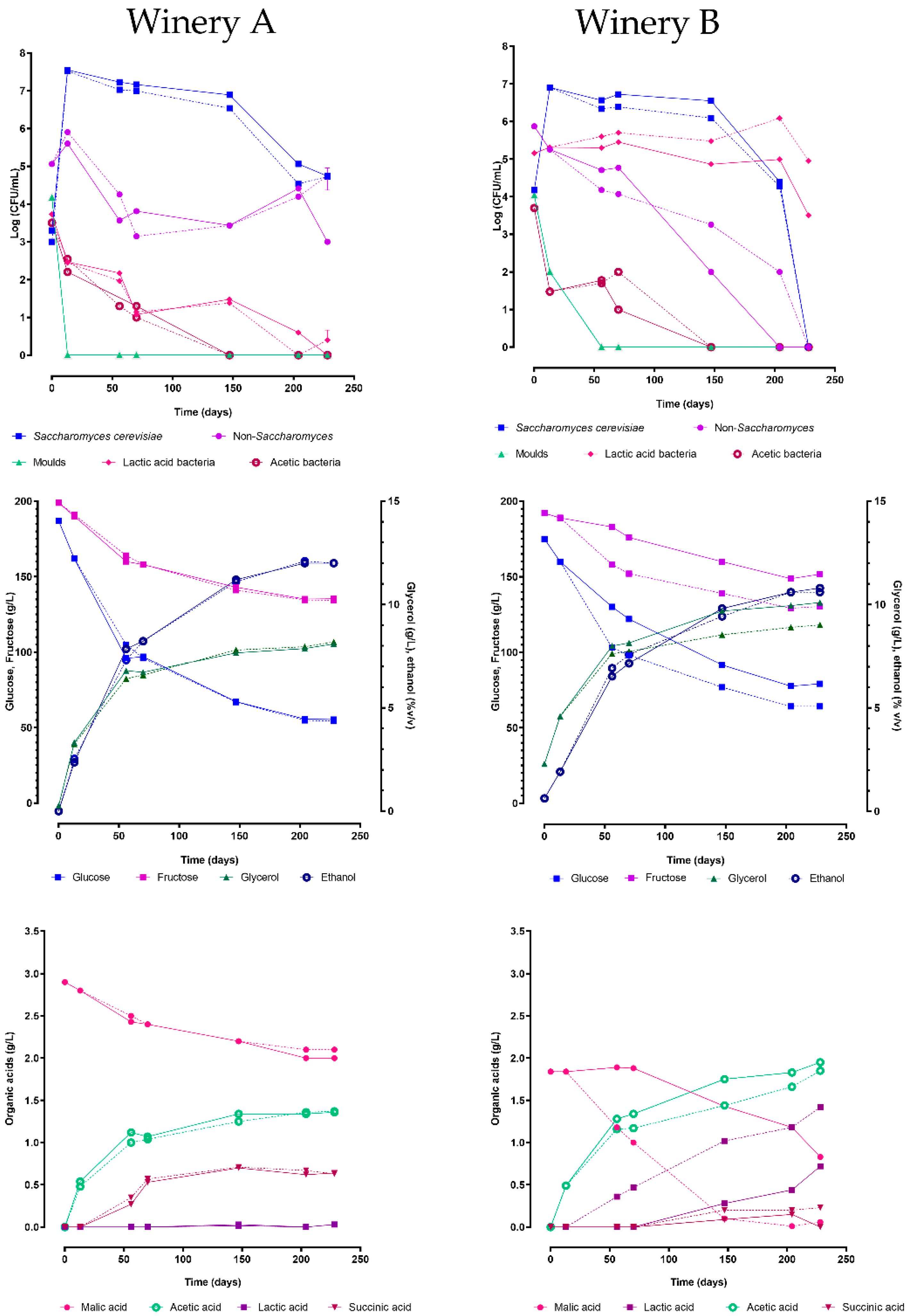
3.2. Microbiota and Chemicals Kinetics During Spontaneous Fermentations
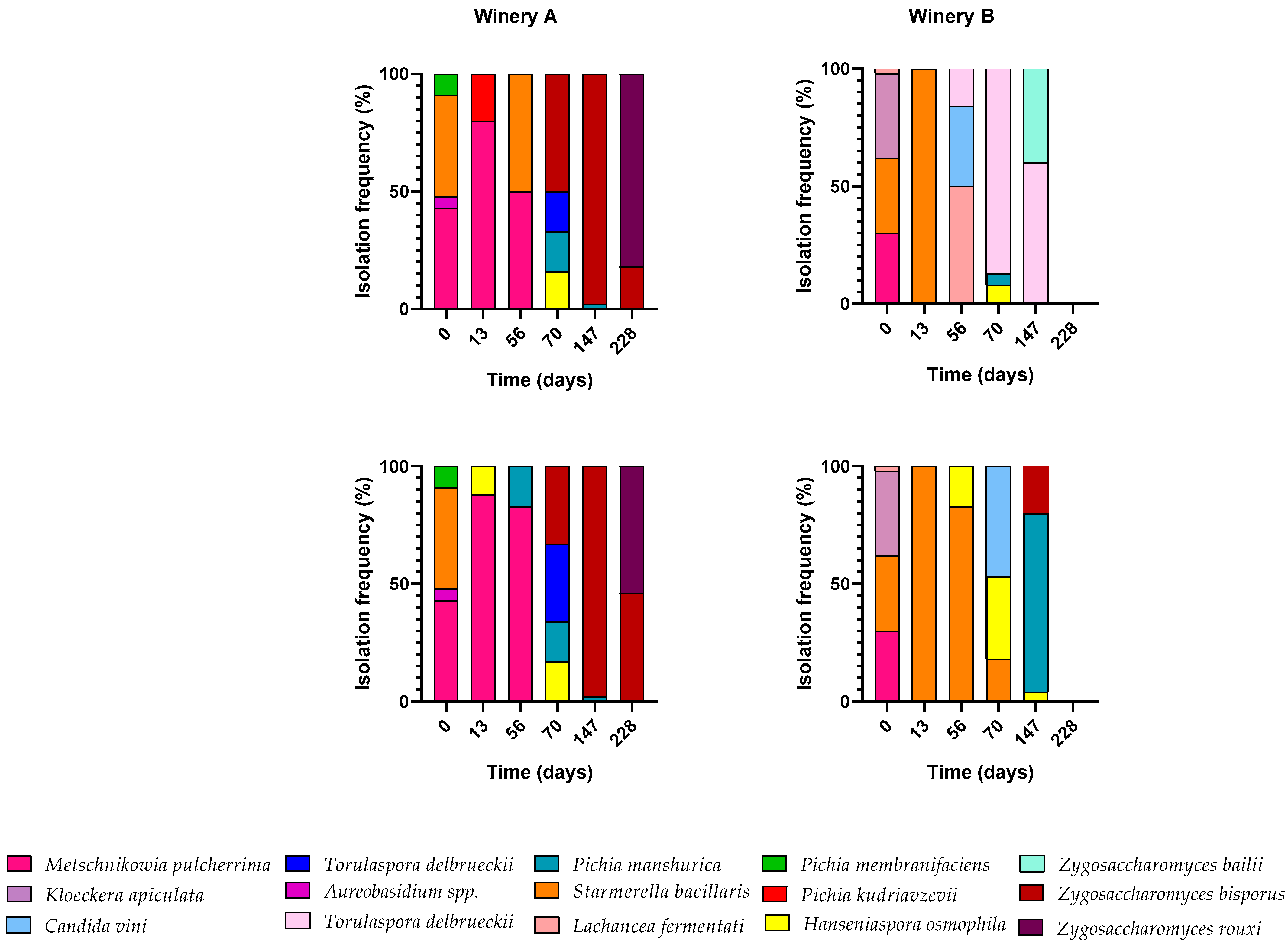
3.3. Biodiversity of Non-Saccharomyces Yeasts During Fermentation
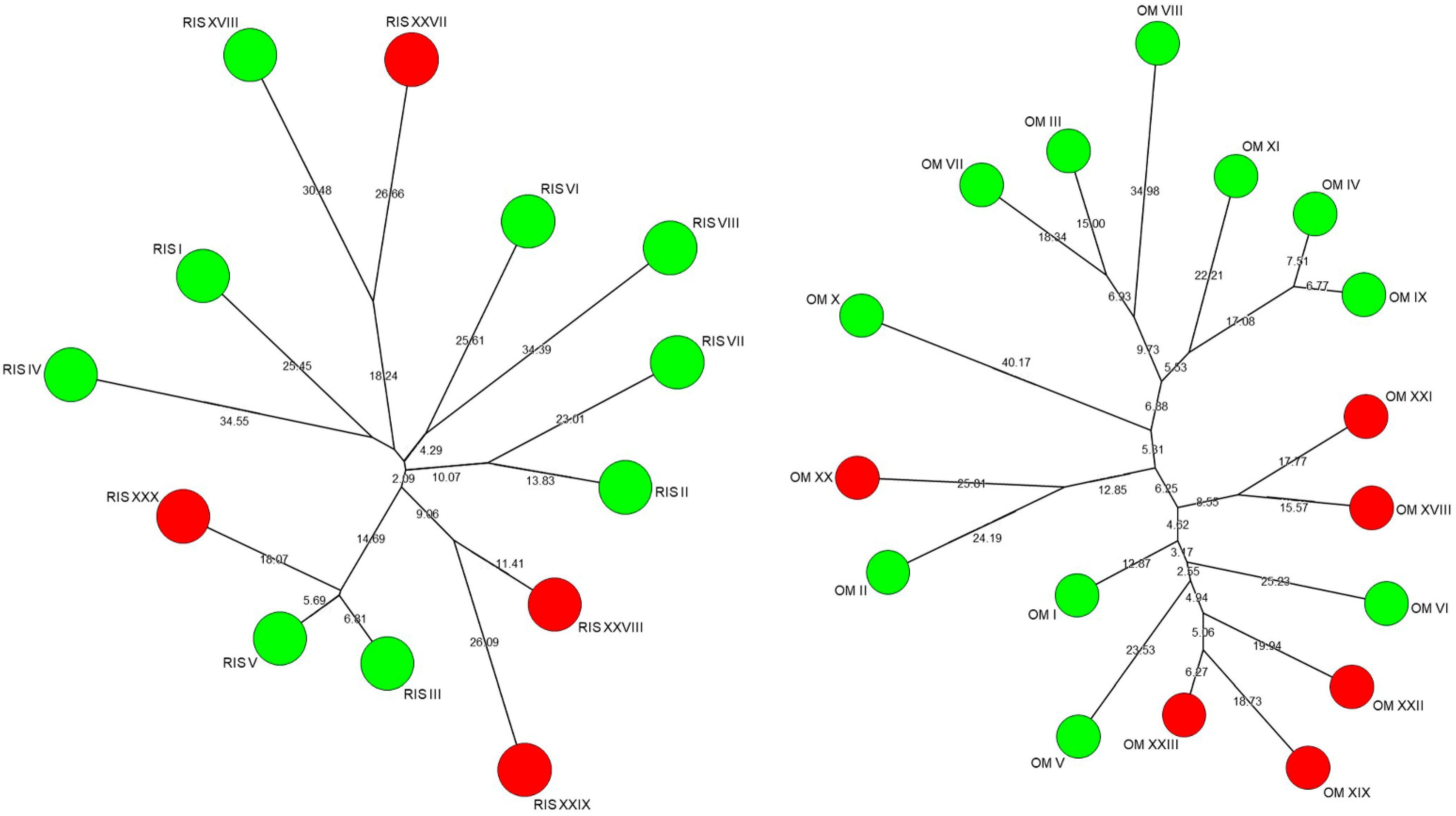
3.4. Intraspecific Biodiversity of Saccharomyces cerevisiae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Filippis, F.; Aponte, M.; Piombino, P.; Lisanti, M.T.; Moio, L.; Ercolini, D.; Blaiotta, G. Influence of microbial communities on the chemical and sensory features of Falanghina sweet passito wines. Food Res. Int. 2019, 120, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; Lencioni, L. Vin santo. In Advances in Food and Nutrition Research; Jackson, R.S., Ed.; Academic Press: Waltham, MA, USA, 2011; Volume 63, pp. 41–100. [Google Scholar]
- Domizio, P.; Manazzu, I.; Ciani, M. Impact of mother sediment on yeast growth, biodiversity, and ethanol production during fermentation of Vinsanto wine. Int. J. Food Microbiol. 2009, 129, 83–87. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EU) No 401/2010 of 7 May 2010 amending and correcting Regulation (EC) No 607/2009 laying down certain detailed rules for the implementation of Council Regulation (EC) No 479/2008 as regards protected designations of origin and geographical indications. traditional terms. labelling and presentation of certain wine sector products. Off. J. Eur. Union 2010, 53, 13–59. [Google Scholar]
- Laureati, M.; Cattaneo, C.; Tateo, F.; Bononi, M. Identification of the Volatile Compounds and Sensory Attributes of Long-Term Aging Vin Santo Wine from Malvasia di Candia Aromatic Grapes. Foods 2020, 9, 1736. [Google Scholar] [CrossRef]
- Tofalo, R.; Chaves-López, C.; Di Fabio, F.; Schirone, M.; Felis, G.E.; Torriani, S.; Paparella, A.; Suzzi, G. Molecular identification and osmotolerant profile of wine yeasts that ferment a high sugar grape must. Int. J. Food Microbiol. 2009, 130, 179–187. [Google Scholar] [CrossRef]
- Kertsch, A.L.; Wagner, J.; Henle, T. Selected Maillard Reaction Products and Their Yeast Metabolites in Commercial Wines. J. Agric. Food Chem. 2023, 71, 12300. [Google Scholar] [CrossRef]
- Romani, C.; Lencioni, L.; Domizio, P. Chemical composition and organoleptic characteristics of Vinsanto wine obtained by using different Saccharomyces strains. Ann. Microbiol. 2009, 59, 76. [Google Scholar]
- Aponte, M.; Blaiotta, G. Selection of an autochthonous Saccharomyces cerevisiae strain for the vinification of “Moscato di Saracena”, a southern Italy (Calabria Region) passito wine. Food Microbiol. 2016, 54, 30–39. [Google Scholar] [CrossRef]
- De Filippis, F.; La Storia, A.; Blaiotta, G. Monitoring the mycobiota during Greco di Tufo and Anglianico wine fermentation by 18S rRNA gene sequencing. Food Microbiol. 2017, 63, 117–122. [Google Scholar] [CrossRef]
- Englezos, V.; Giacosa, S.; Rantsiou, K.; Rolle, L.; Cocolin, L. Starmerella bacillaris in winemaking: Opportunities and risks. Curr. Opin. Food Sci. 2017, 17, 30–35. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Serafino, G.; Di Gianvito, P.; Giacosa, S.; Škrab, D.; Cocolin, L.; Englezos, V.; Rantsiou, K. Survey of the yeast ecology of dehydrated grapes and strain selection for wine fermentation. Food Res. Int. 2023, 170, 113005. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, D.; Blanco, P. Preliminary Study on Yeasts Associated with the Production of “Tostado”—A Traditional Sweet Wine from Galicia (NW Spain). Front. Biosci. (Elite Ed.) 2024, 16, 10. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Bonesi, M.; Di Lecce, G.; Boselli, E.; Tundis, R.; Pugliese, A.; Menichini, F.; Frega, N.G. Phenolics, Aroma Profile, and In Vitro Antioxidant Activity of Italian Dessert Passito Wine from Saracena (Italy). Food Sci. 2013, 78, C703–C708. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.M.; Inglis, D.L.; Pickering, G.J. Sensorial and volatile analysis of wines made from partially dehydrated grapes: An Ontario case study. J. Food Qual. 2020, 2020, 8861185. [Google Scholar] [CrossRef]
- Guerrini, S.; Barbato, D.; Mangani, S.; Ganucci, D.; Buscioni, G.; Galli, V.; Triossi, A.; Granchi, L. Management of in-Amphora “Trebbiano Toscano” Wine Production: Selection of Indigenous Saccharomyces cerevisiae Strains and Influence on the Phenolic and Sensory Profile. Foods 2023, 12, 2372. [Google Scholar] [CrossRef]
- Granchi, L.; Bosco, M.; Messini, A.; Vincenzini, M. Rapid detection and quantification of yeast species during spontaneous wine fermentation by PCR–RFLP analysis of the rDNA ITS region. J. Appl. Microbiol. 1999, 87, 949–956. [Google Scholar] [CrossRef]
- Legras, J.L.; Karst, F. Optimisation of interdelta analysis for Saccharomyces cerevisiae strain characterisation. FEMS Microbiol. Lett. 2003, 221, 249–255. [Google Scholar] [CrossRef]
- Shannon, S.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Champaign, IL, USA, 1963. [Google Scholar]
- Mari, E.; Guerrini, S.; Granchi, L.; Vincenzini, M. Yeast microbiota in the olive oil extractive process: A three-year study at an industrial scale. World J. Microbiol. Biotechnol. 2016, 32, 93–103. [Google Scholar] [CrossRef]
- Sultan, Y.; Magan, N.; Medina, A. Comparison of five different C18 HPLC analytical columns for the analysis of ochratoxin A in different matrices. J. Chromatogr. B 2014, 971, 89–93. [Google Scholar] [CrossRef]
- Saitou, N.; Masatoshi, N. The Neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Rantsiou, K.; Campolongo, S.; Alessandria, V.; Rolle, L.; Torchio, F.; Cocolin, L. Yeast populations associated with grapes during withering and their fate during alcoholic fermentation of high-sugar must. Aust. J. Grape Wine Res. 2013, 19, 40–46. [Google Scholar] [CrossRef]
- Nadai, C.; Fernandes Lemos, W.J.; Favaron, F.; Giacomini, A.; Corich, V. Biocontrol activity of Starmerella bacillaris yeast against blue mold disease on apple fruit and its effect on cider fermentation. PLoS ONE 2018, 13, e0204350. [Google Scholar] [CrossRef]
- Englezos, V.; Jolly, N.; Di Gianvito, P.; Rantsiou, K.; Cocolin, L. Microbial interactions in winemaking: Ecological aspects and effect on wine quality. Trends Food Sci. Technol. 2022, 126, 99–113. [Google Scholar] [CrossRef]
- Erasmus, D.J.; van der Merwe, G.K.; van Vuuren, H.J. Genome-wide expression analyses: Metabolic adaptation of Saccharomyces cerevisiae to high sugar stress. FEMS Yeast Res. 2003, 3, 375–399. [Google Scholar] [CrossRef]
- Kimani, B.G.; Kerekes, E.B.; Szebenyi, C.; Krisch, J.; Vágvölgyi, C.; Papp, T.; Takó, M. In Vitro Activity of Selected Phenolic Compounds against Planktonic and Biofilm Cells of Food-Contaminating Yeasts. Foods 2021, 10, 1652. [Google Scholar] [CrossRef]
- Pastorkova, E.; Zakova, T.; Landa, P.; Novakova, J.; Vadlejch, J.; Kokoska, L. Growth inhibitory effect of grape phenolics against wine spoilage yeasts and acetic acid bacteria. Int. J. Food Microbiol. 2013, 161, 209–213. [Google Scholar] [CrossRef]
- Sabel, A.; Bredefeld, S.; Schlander, M.; Claus, H. Wine Phenolic Compounds: Antimicrobial Properties against Yeasts, Lactic Acid and Acetic Acid Bacteria. Beverages 2017, 3, 29. [Google Scholar] [CrossRef]
- Sun, X.; Liu, L.; Zhao, Y.; Ma, T.; Zhao, F.; Huang, W.; Zhan, J. Effect of copper stress on growth characteristics and fermentation properties of Saccharomyces cerevisiae and the pathway of copper adsorption during wine fermentation. Food Chem. 2016, 192, 43–52. [Google Scholar] [CrossRef]
- Granchi, L.; Ganucci, D.; Buscioni, G.; Mangani, S.; Guerrini, S. The biodiversity of Saccharomyces cerevisiae in spontaneous wine fermentation: The occurrence and persistence of winery-strains. Fermentation 2019, 5, 86. [Google Scholar] [CrossRef]
- Berbegal, C.; Spano, G.; Tristezza, M.; Grieco, F.; Capozzi, V. Microbial Resources and Innovation in the Wine Production Sector. S. Afr. J. Enol. Vitic. 2017, 38, 156–166. [Google Scholar] [CrossRef]
- Tufariello, M.; Fragasso, M.; Pico, J.; Panighel, A.; Castellarin, S.D.; Flamini, R.; Grieco, F. Influence of Non-Saccharomyces on Wine Chemistry: A Focus on Aroma-Related Compounds. Molecules 2021, 26, 644. [Google Scholar] [CrossRef] [PubMed]
- Torres-Guardado, R.; Rozès, N.; Esteve-Zarzoso, B.; Reguant, C.; Bordons, A. Succinic acid production by wine yeasts and the influence of GABA and glutamic acid. Int. Microbiol. 2024, 27, 505–512. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Winery A-100 L | Winery A-50 L | Winery B-100 L | Winery B-50 L | |
|---|---|---|---|---|
| Polyphenols (mg/L) | ||||
| gallic acid | 15.3 ± 0.8 a | 20.4 ± 1.0 a | 36.3 ± 1.8 b | 35.1 ± 0.2 b |
| protocatechuic acid | 13.4 ± 0.7 a | 15.2 ± 0.8 a | 25.1 ± 0.3 b | 21.7 ± 1 b |
| caftaric acid | 7.9 ± 0.4 a | 8.2 ± 0.4 a | 17.1 ± 0.9 b | 18.3 ± 0.9 b |
| catechin | 12.7 ± 0.6 a | 12.8 ± 0.6 a | 14.2 ± 0.7 a | 8.7 ± 0.4 c |
| vanillic acid | 16.8 ± 0.8 a | 19.0 ± 0.9 a | 25.2 ± 1.26 b | 17.0 ± 0.8 a |
| caffeic acid | 4.6 ± 02 a | 4.5 ± 02 a | 17.2 ± 09 b | 18.4 ± 09 b |
| Total polyphenols index (TPI) | 31.19 ± 0.20 ab | 32.48 ± 0.10 a | 37.62 ± 0.35 c | 29.73 ± 0.44 b |
| Cu2+ (mg/L) | 2.13 ± 0.10 a | 1.35 ± 0.07 b | 0.53 ± 0.03 c | 0.37 ± 0.02 c |
| Degree of browning (Abs 420) | 0.1032 ± 0.0030 a | 0.1072 ± 0.0040 a | 0.2370 ± 0.0035 b | 0.1804 ± 0.0025 c |
| H Index | E Index | |
|---|---|---|
| Winery A 100 L | 2.28 | 0.55 |
| Winery A 50 L | 2.26 | 0.55 |
| Winery B 100 L | 2.17 | 0.53 |
| Winery B 50 L | 2.65 | 0.65 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbato, D.; Galli, V.; Mangani, S.; Mari, E.; Buscioni, G.; Granchi, L.; Guerrini, S. Microbiota Diversity During Grape Drying and Spontaneous Fermentations of Vin Santo. Fermentation 2025, 11, 310. https://doi.org/10.3390/fermentation11060310
Barbato D, Galli V, Mangani S, Mari E, Buscioni G, Granchi L, Guerrini S. Microbiota Diversity During Grape Drying and Spontaneous Fermentations of Vin Santo. Fermentation. 2025; 11(6):310. https://doi.org/10.3390/fermentation11060310
Chicago/Turabian StyleBarbato, Damiano, Viola Galli, Silvia Mangani, Eleonora Mari, Giacomo Buscioni, Lisa Granchi, and Simona Guerrini. 2025. "Microbiota Diversity During Grape Drying and Spontaneous Fermentations of Vin Santo" Fermentation 11, no. 6: 310. https://doi.org/10.3390/fermentation11060310
APA StyleBarbato, D., Galli, V., Mangani, S., Mari, E., Buscioni, G., Granchi, L., & Guerrini, S. (2025). Microbiota Diversity During Grape Drying and Spontaneous Fermentations of Vin Santo. Fermentation, 11(6), 310. https://doi.org/10.3390/fermentation11060310