
Staged Fermentation with a Saccharomyces cerevisiae Consortium and Monascus purpureus for Sustainable Valorization of Persimmon into High-Quality Wine


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Persimmon Juice Preparation
2.3. Strain Activation and Inoculum Preparation
2.4. Single-Factor Optimization of Mixed Yeast Fermentation for Alcohol Production
2.4.1. Determination of Optimal Yeast Composition Ratio
2.4.2. Optimization of Yeast Inoculation Ratio
2.4.3. Effect of Initial Sugar Concentration
2.4.4. Effect of pH
2.4.5. Effect of Temperature
2.4.6. Effect of Fermentation Duration
2.5. Multivariate Optimization of Mixed Yeast Fermentation Using Box–Behnken Design
2.6. Single-Factor Optimization of Monascus purpureus Fermentation for Esterification
2.6.1. Effect of Monascus purpureus Inoculation Ratio
2.6.2. Effect of Initial Sugar Concentration
2.6.3. Effect of pH
2.6.4. Effect of Temperature
2.6.5. Effect of Fermentation Duration
2.7. Multivariate Optimization of Monascus purpureus Fermentation Using Box–Behnken Design
2.8. Sensory Evaluation and Aroma Profiling
2.9. Staged Fermentation Protocol
3. Results
3.1. Synergistic Effects of Mixed Yeast Composition Ratios
3.2. Single-Factor Optimization of Mixed Yeast Fermentation for Alcohol Production
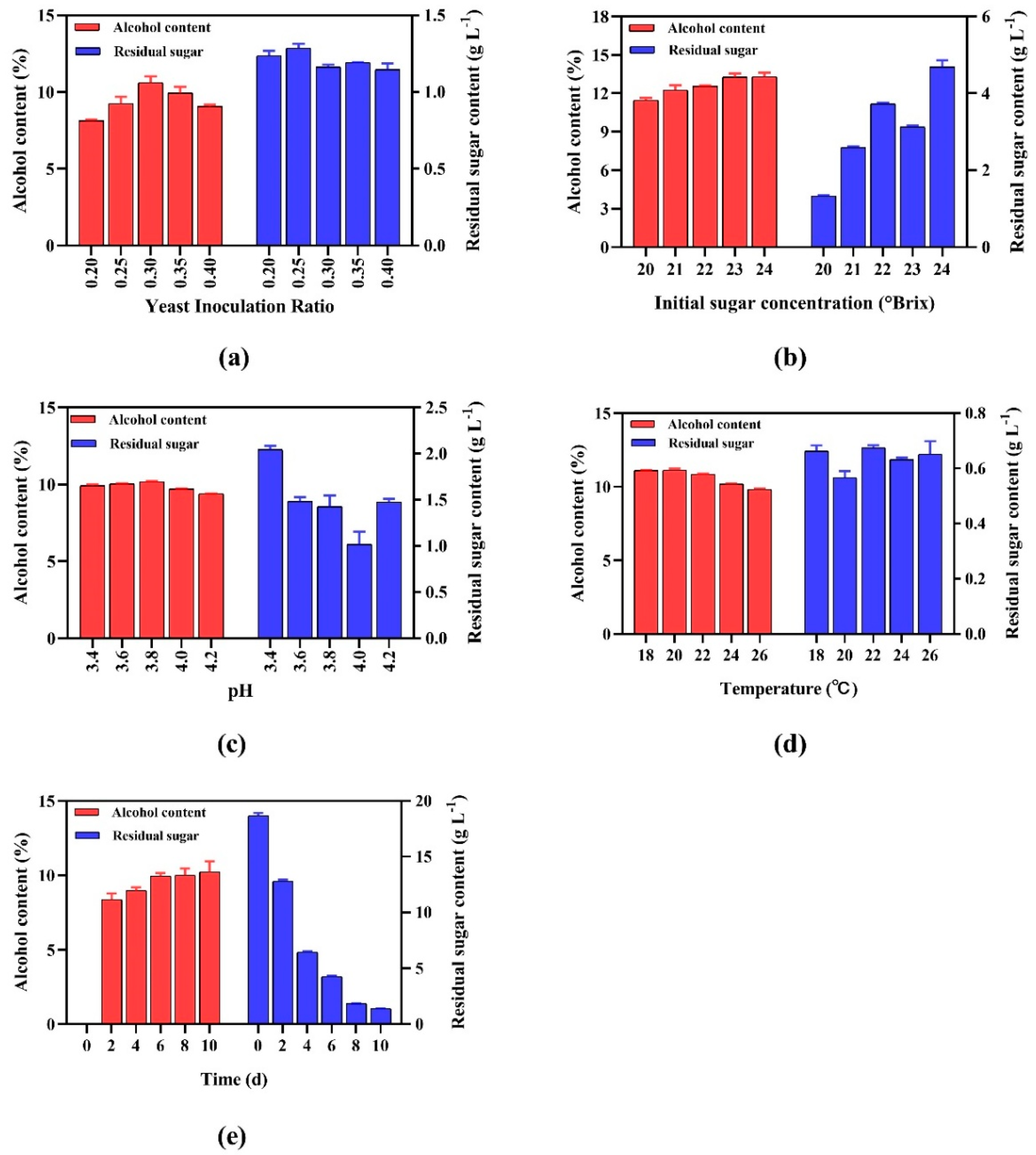
3.2.1. Yeast Inoculation Ratio
3.2.2. Initial Sugar Concentration
3.2.3. pH Dependence
3.2.4. Temperature Effects
3.2.5. Fermentation Duration
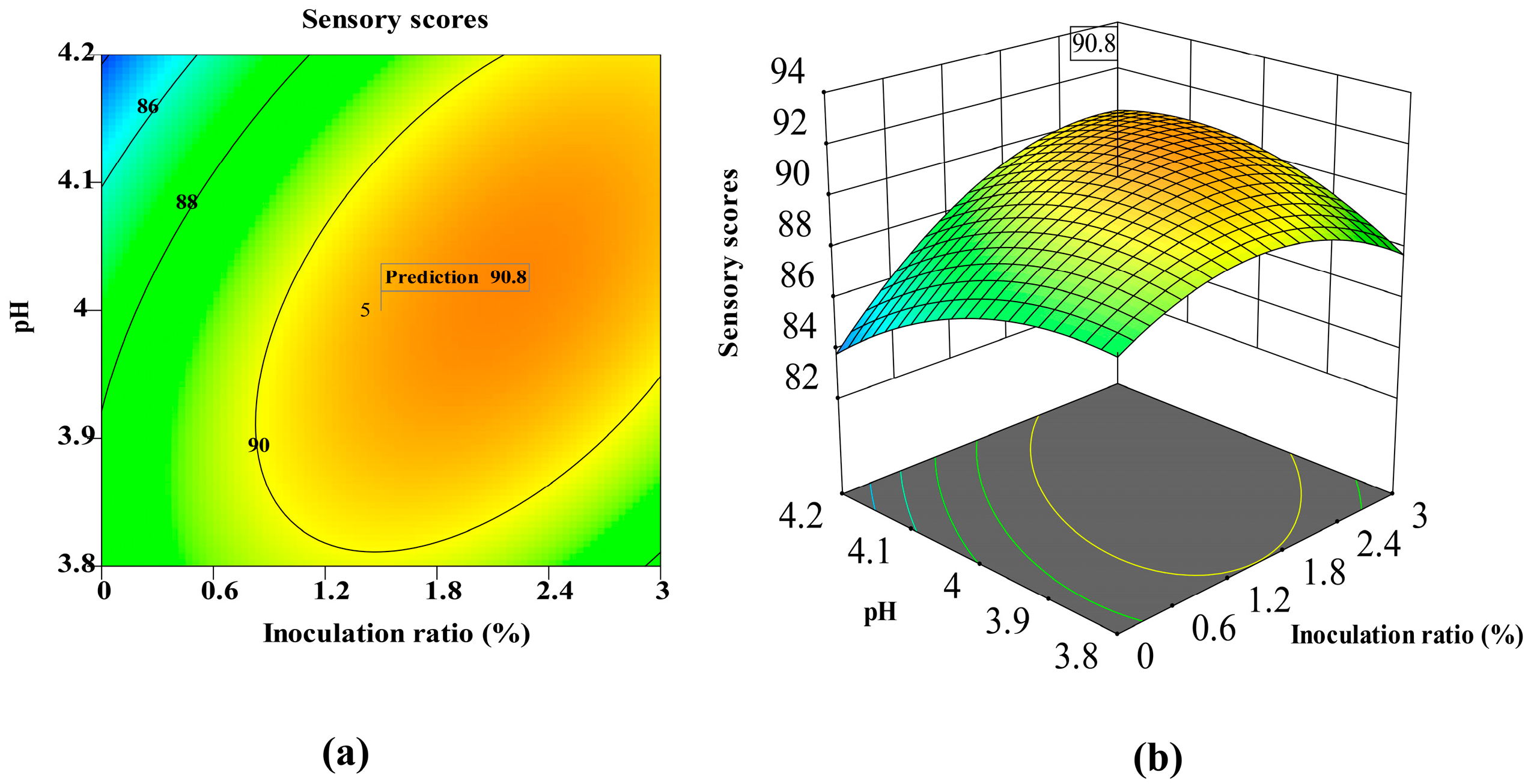
3.3. Multivariate Optimization of Mixed Yeast Fermentation Using Box–Behnken Design
3.4. Single-Factor Optimization of Monascus purpureus Fermentation for Esterification
3.4.1. Inoculation Ratio
3.4.2. Initial Sugar Concentration
3.4.3. pH Dependence
3.4.4. Temperature Effects
3.4.5. Fermentation Duration
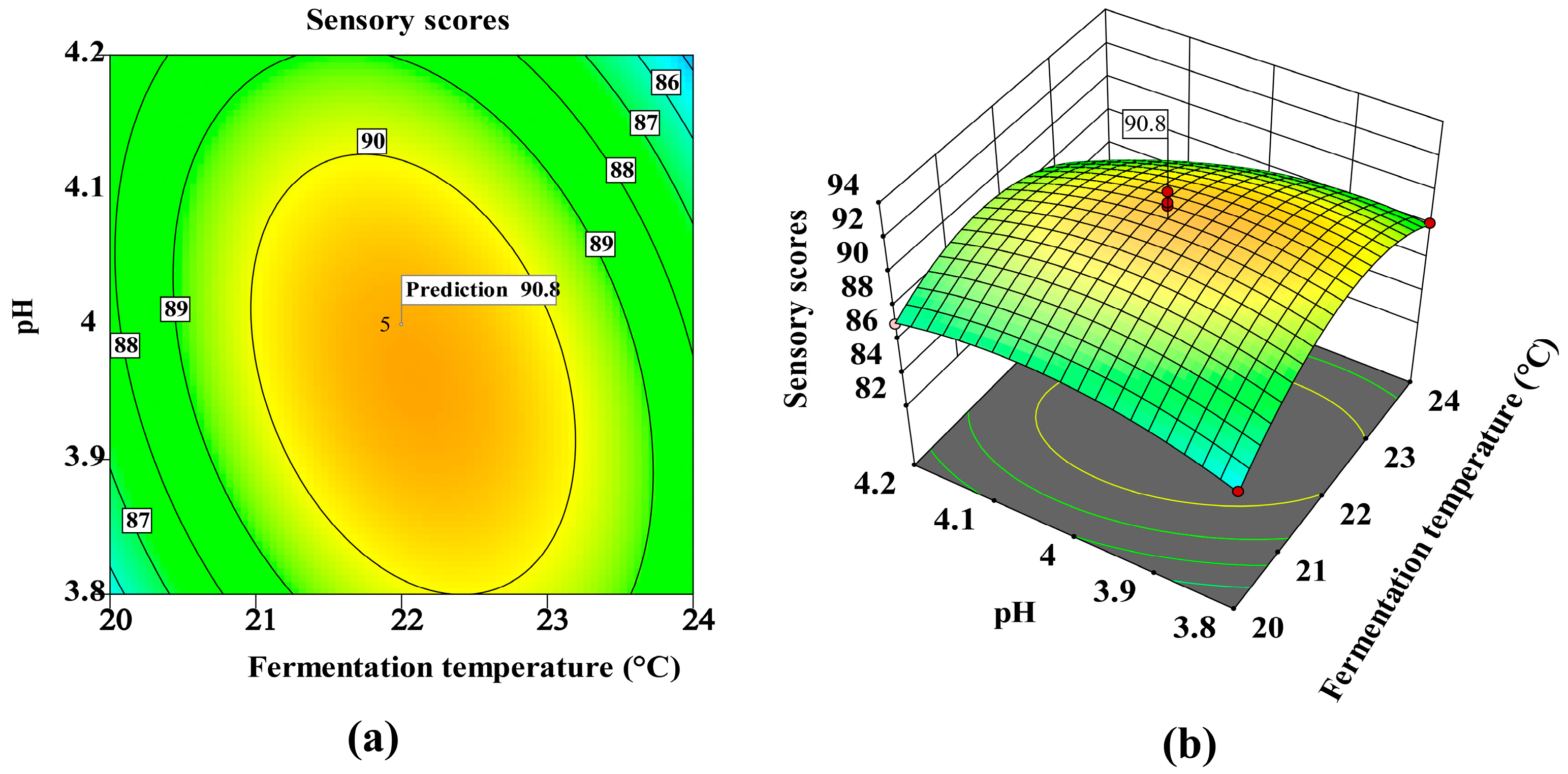
3.5. Multivariate Optimization of Monascus purpureus Fermentation Using Box–Behnken Design
3.6. Sensory Evaluation and Aroma Profile of Persimmon Wine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GC–MS | Gas chromatography–mass spectrometry |
| RSM | Response surface methodology |
| SPME | Solid phase microextraction |
| MSF | Multi-strain fermentation |
References
- Matheus, J.R.V.; Andrade, C.J.; de Miyahira, R.F.; Fai, A.E.C. Persimmon (Diospyros kaki L.): Chemical properties, bioactive compounds and potential use in the development of new products-A Review. Food Rev. Int. 2020, 38, 384–401. [Google Scholar] [CrossRef]
- M. González, C.; Hernando, I.; Moraga, G. In vitro and in vivo digestion of persimmon and derived products: A review. Foods 2021, 10, 3083. [Google Scholar] [CrossRef] [PubMed]
- Gea-Botella, S.; Agulló, L.; Martí, N.; Martínez-Madrid, M.C.; Lizama, V.; Martín-Bermudo, F.; Berná, G.; Saura, D.; Valero, M. Carotenoids from persimmon juice processing. Food Res. Int. 2021, 141, 109882. [Google Scholar] [CrossRef] [PubMed]
- Hosseininejad, S.; González, C.M.; Hernando, I.; Moraga, G. Valorization of persimmon fruit through the development of new food products. Front. Food Sci. Technol. 2022, 2, 914952. [Google Scholar] [CrossRef]
- Kim, Y.M.; Lubinska-Szczygel, M.; Park, Y.S.; Deutsch, J.; Ezra, A.; Luksrikul, P.; Shafreen, R.M.B.; Gorinstein, S. Characterization of bioactivity of selective molecules in fruit wines by FTIR and NMR spectroscopies, fluorescence and docking calculations. Molecules 2023, 28, 6036. [Google Scholar] [CrossRef]
- An, X.; Wang, Z.; Li, J.; Nie, X.; Liu, K.; Zhang, Y.; Zhao, Z.; Chitrakar, B.; Ao, C. Analysis of flavor-related compounds in fermented persimmon beverages stored at different temperatures. Lwt 2022, 163, 113524. [Google Scholar] [CrossRef]
- Wei, J.; Li, Y.; Liu, Y.; Liu, S.; Yang, X.; Wang, X. Process optimization for production of persimmon wine with lower methanol. Foods 2024, 13, 748. [Google Scholar] [CrossRef]
- Lao-Martil, D.; Verhagen, K.J.; Schmitz, J.P.; Teusink, B.; Wahl, S.A.; van Riel, N.A. Kinetic modeling of Saccharomyces cerevisiae central carbon metabolism: Achievements, limitations, and opportunities. Metabolites 2022, 12, 74. [Google Scholar] [CrossRef]
- García-Barón, S.E.; Carmona-Escutia, R.P.; Herrera-López, E.J.; Leyva-Trinidad, D.A.; Gschaedler-Mathis, A. Consumers’ drivers of perception and preference of fermented food products and beverages: A systematic review. Foods 2025, 14, 713. [Google Scholar] [CrossRef]
- Niyomvong, N.; Trakunjae, C.; Boondaeng, A. Fermentation characteristics and aromatic profiles of plum wines produced with Hanseniaspora thailandica Zal1 and common wine yeasts. Molecules 2023, 28, 3009. [Google Scholar] [CrossRef]
- Choi, K.T.; Lee, S.H.; Kim, Y.J.; Choi, J.S.; Lee, S.B. Improvement of volatile aromatic compound levels and sensory quality of distilled soju derived from Saccharomyces cerevisiae and Wickerhamomyces anomalus co-fermentation. Food Chem. X 2024, 22, 101368. [Google Scholar] [CrossRef] [PubMed]
- De Gioia, M.; Russo, P.; De Simone, N.; Grieco, F.; Spano, G.; Capozzi, V.; Fragasso, M. Interactions among relevant non-Saccharomyces, Saccharomyces, and lactic acid bacteria species of the wine microbial consortium: Towards advances in antagonistic phenomena and biocontrol potential. Appl. Sci. 2022, 12, 12760. [Google Scholar] [CrossRef]
- Du, Y.; Shim, S.; Wang, L.; Gao, X.; Fu, X. Impact of Monascus purpureus combined with Lactobacillus plantarum and Saccharomyces cerevisiae fermentation on nutritional and flavor characteristics of Pyropia yezoensis. Food Chem. 2025, 472, 142973. [Google Scholar] [CrossRef] [PubMed]
- Rong, Y.; Yu, X.; Hong, K. Synergistic effect enhances aromatic profile in beer brewing through mixed-culture fermentation of Pichia kluyveri and Saccharomyces cerevisiae var. diastaticus. Fermentation 2025, 11, 148. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, J.; Chen, Q.; Yang, C. Effects of main nutrient sources on improving monascus pigments and saccharifying power of Monascus purpureus in submerged fermentation. Fermentation 2023, 9, 696. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, C.; Ablimit, A.; Chen, M.; Sun, Q.; Dong, H.; Liu, W.; Huang, A.; Zhang, B.; Hu, W.; et al. Development of red grapefruit jam fermented with Monascus purpureus: Effect of naringenin on Monascus purpureus growth and metabolism. Food Biosci. 2025, 65, 106061. [Google Scholar] [CrossRef]
- Arun, K.B.; Anoopkumar, A.N.; Sindhu, R.; Binod, P.; Aneesh, E.M.; Madhavan, A.; Awasthi, M.K. Synthetic biology for sustainable food ingredients production: Recent trends. Syst. Microbiol. Biomanuf. 2023, 3, 137–149. [Google Scholar] [CrossRef]
- Cosme, F.; Inês, A.; Vilela, A. Microbial and commercial enzymes applied in the beverage production process. Fermentation 2023, 9, 385. [Google Scholar] [CrossRef]
- Lan, L.; Cao, Y.; Yuan, J.; Feng, R.; Pan, H.; Mao, X.; Ji, S.; Hu, Q.; Zhou, H. A comprehensive investigation of lipid profile during the solid-state fermentation of rice by Monascus purpureus. Foods 2025, 14, 537. [Google Scholar] [CrossRef]
- Tong, S.; Li, W.; Rao, Y.; Xiao, Y.; Yan, Y.; Guo, W.; Lü, X.; Sun, J.; Ai, L.; Ni, L. Microbiomics and metabolomics insights into the microbial regulation on the formation of flavor components in the traditional fermentation process of Chinese Hongqu aged vinegar. Food Sci. Hum. Well. 2024, 13, 2765–2778. [Google Scholar] [CrossRef]
- GB/T 5009.7-2016; National Food Safety Standard—Determination of Reducing Sugar in Foods. National Standards of the People’s Republic of China: Beijing, China, 2016.
- GB 5009.3-2016; National Food Safety Standard—Determination of Moisture Content in Foods. National Standards of the People’s Republic of China: Beijing, China, 2016.
- GB/T 10468-1989; Fruit and Vegetable Products—Determination of pH. State Technical Supervision Bureau: Beijing, China, 1989.
- NY/T 1600-2008; Determination of Tannin Content in Fruit, Vegetable and Derived Product. Spectrophotometry Method. National Standards of the People’s Republic of China: Beijing, China, 2008.
- GB/T 15038-2006; Analytical Methods of Wine and Fruit Wine. National Standards of the People’s Republic of China: Beijing, China, 2006.
- ISO 11035:1994; Sensory Analysis—Identification and Selection of Descriptors for Establishing a Sensory Profile by a Multidimensional Approach. ISO: Geneva, Switzerland, 1994.
- Gong, Z.; Wu, Z.; Yang, Q.; Liu, J.; Jiao, P.; Tang, C. Influences of lactic acid bacteria strains on the flavor profiles, metabolites and quality characteristics of red yeast rice produced by solid-state fermentation. Food Res. Int. 2024, 197, 115172. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wang, M.; Ding, Y.; Ding, Y.; Zhou, X. Flavor and quality enhancement of miiuy croaker (Miichthys miiuy) surimi by co-fermentation with Monascus purpureus and Actinomucor elegans. Food Biosci. 2025, 67, 106231. [Google Scholar] [CrossRef]
- Zhang, J.; Li, M.; Li, L.; Liu, Y.; Gu, T.; Wang, J.; Gao, M. Evaluation of differences in volatile flavor compounds between liquid-state and solid-state fermented Tartary buckwheat by Monascus purpureus. Food Chem. 2025, 464, 141662. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Xi, B.; Lu, L. Strategies to enhance production of metabolites in microbial co-culture systems. Bioresour. Technol. 2024, 406, 131049. [Google Scholar] [CrossRef]
- Anantayanon, J.; Chamkhuy, W.; Rattanaphan, N.; Panchanawaporn, S.; Laoteng, K.; Jeennor, S. Enhancing bioactive cordycepin production via precision fermentation with an engineered Aspergillus oryzae. Fermentation 2025, 11, 32. [Google Scholar] [CrossRef]
- Živković, N.M.; Čakar, U.D.; Petrović, A.V. Effects of spontaneous and inoculated fermentation on the total phenolic content and antioxidant activity of Cabernet Sauvignon wines and fermented pomace. Food Feed Res. 2024, 51, 119–129. [Google Scholar] [CrossRef]
- Zhang, J.X.; Liu, X.L.; Wang, L.; Fang, Z. Two-stage process production of microbial lipid by co-fermentation of glucose and N-acetylglucosamine from food wastes with Cryptococcus curvatus. Bioresour. Technol. 2023, 387, 129685. [Google Scholar] [CrossRef]
- Xin, Y.; Qiao, M. Towards microbial consortia in fermented foods for metabolic engineering and synthetic biology. Food Res. Int. 2025, 201, 115677. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, S.; Wang, H.; Wu, Q.; Guo, W.; Ni, L.; Lv, X. Metagenomic and metabolomic profiling reveals the differences of flavor quality between Hongqu rice wines fermented with Gutian qu and Wuyi qu. Foods 2024, 13, 3114. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, Z.; Liang, X.; Tu, C.; Khalifa, I.; Wang, C.; Zhu, Y.; Chen, H.; Hu, L.; Li, C. Unlocking the potential of persimmons: A comprehensive review on emerging technologies for post-harvest challenges, processing innovations, and prospective applications. Food Chem. 2024, 459, 140344. [Google Scholar] [CrossRef]
- García-García, J.C.; G-García, M.E.; Mauricio, J.C.; Moreno, J.; García-Martínez, T. Evaluation of the protein profile of a Saccharomyces cerevisiae strain immobilized in biocapsules for use in fermented foods. Foods 2024, 13, 3871. [Google Scholar] [CrossRef]
- Wang, B.; Duan, Y.; Wang, C.; Liu, C.; Wang, J.; Jia, J.; Wu, Q. Combined volatile compounds and non-targeted metabolomics analysis reveals variation in flavour characteristics, metabolic profiles and bioactivity of mulberry leaves after Monascus purpureus fermentation. J. Sci. Food Agric. 2024, 104, 3294–3305. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition Ratio EC:Aq:F33 | CO2 Weight Loss (g) | Residual Sugar Concentration (g L−1) |
|---|---|---|
| 1/0/0 | 6.39 ± 0.17 | 1.51 ± 0.08 |
| 0/1/0 | 6.25 ± 0.16 | 1.58 ± 0.08 |
| 0/0/1 | 6.20 ± 0.20 | 1.65 ± 0.10 |
| 7/4/4 | 8.85 ± 0.16 | 0.64 ± 0.14 |
| 4/4/7 | 10.99 ± 0.34 | 0.62 ± 0.01 |
| 4/7/4 | 9.71 ± 0.27 | 1.01 ± 0.02 |
| 1/2/2 | 7.73 ± 0.30 | 0.64 ± 0.03 |
| 2/1/2 | 9.12 ± 0.17 | 0.63 ± 0.12 |
| 2/2/1 | 8.96 ± 0.27 | 0.67 ± 0.01 |
| 1/1/3 | 8.14 ± 0.36 | 0.77 ± 0.03 |
| 1/3/1 | 9.01 ± 0.07 | 0.84 ± 0.04 |
| 3/1/1 | 10.06 ± 0.15 | 0.65 ± 0.04 |
| 1/1/1 | 10.03 ± 0.12 | 0.67 ± 0.02 |
| Source | Sum of Squares | df | Mean Square | F-value | p-Value | Significant |
|---|---|---|---|---|---|---|
| Model | 14.49 | 9 | 1.61 | 12.97 | 0.00014 | ** |
| A—Initial sugar concentration | 7.51 | 1 | 7.51 | 60.52 | 0.0001 | ** |
| B—Inoculation ratio | 0.5243 | 1 | 0.5243 | 4.22 | 0.0789 | |
| C—Temperature | 2.74 | 1 | 2.74 | 22.06 | 0.0022 | ** |
| AB | 0.2153 | 1 | 0.2153 | 1.73 | 0.2293 | |
| AC | 2.06 | 1 | 2.06 | 16.61 | 0.0047 | ** |
| BC | 0.0001 | 1 | 0.0001 | 0.0005 | 0.9825 | |
| A2 | 0.1070 | 1 | 0.1070 | 0.8619 | 0.3841 | |
| B2 | 0.4349 | 1 | 0.4349 | 3.50 | 0.1034 | |
| C2 | 0.7691 | 1 | 0.7691 | 6.20 | 0.0416 | ** |
| Residual | 0.8688 | 7 | 0.1241 | |||
| Lack of Fit | 0.6314 | 3 | 0.2105 | 3.55 | 0.1265 | |
| Pure Error | 0.2374 | 4 | 0.0593 | |||
| Cor Total | 15.36 | 16 | ||||
| R2 | 0.9434 |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value | Significant |
|---|---|---|---|---|---|---|
| Model | 108.82 | 9 | 12.09 | 9.37 | 0.0038 | ** |
| A—Inoculation ratio | 18.61 | 1 | 18.61 | 14.41 | 0.0067 | ** |
| B—Temperature | 0.0012 | 1 | 0.0012 | 0.0010 | 0.9760 | |
| C-pH | 1.71 | 1 | 1.71 | 1.33 | 0.2874 | |
| AB | 3.61 | 1 | 3.61 | 2.80 | 0.1384 | |
| AC | 11.56 | 1 | 11.56 | 8.95 | 0.0202 | * |
| BC | 6.00 | 1 | 6.00 | 4.65 | 0.0680 | |
| A2 | 15.40 | 1 | 15.40 | 11.93 | 0.0106 | * |
| B2 | 37.58 | 1 | 37.58 | 29.11 | 0.0010 | ** |
| C2 | 8.11 | 1 | 8.11 | 6.28 | 0.0407 | * |
| Residual | 9.04 | 7 | 1.29 | |||
| Lack of Fit | 0.4375 | 3 | 0.1458 | 0.0678 | 0.9741 | |
| Pure Error | 8.60 | 4 | 2.15 | |||
| Cor Total | 117.86 | 16 |
| No | Compounds | Fermented Methods (Area, %) | Odor Description | ||||
|---|---|---|---|---|---|---|---|
| MSF | EC | Aq | F33 | Mixed Yeast | |||
| 1 | Phenylethyl alcohol | 73.08 | 28.64 | 33.62 | 42.31 | 47.06 | Rose-like fragrance |
| 2 | 4-Hydroxyphenethyl alcohol | 0.23 | 0.04 | 0.06 | 0.19 | Sweet floral-fruity aroma | |
| 3 | 1-Decanol | 0.16 | Waxy, sweet, floral, fruity aroma | ||||
| 4 | Decyl methanol | 0.12 | 0.05 | 0.02 | 0.02 | 0.06 | Mild sweet rose-like scent |
| 5 | 1-Decanol | 0.16 | Waxy, sweet, floral, fruity aroma | ||||
| 6 | Terpinen-4-ol | 0.43 | Spicy note | ||||
| 7 | Nonanediol | 0.68 | Rosy, waxy, fruity lipid aroma | ||||
| 8 | Lauryl alcohol | 0.12 | Tuberose, violet-like scent | ||||
| 9 | Geranylgeraniol | 0.28 | Blooming rose aroma | ||||
| 10 | Ethyl 2-methylacetoacetate | 0.12 | Fruity aroma | ||||
| 11 | Isoamyl propionate | 0.14 | Fresh sweet fruity aroma | ||||
| 12 | 3,4-Dimethylbenzaldehyde | 2.99 | 0.48 | 0.59 | 0.54 | 0.12 | |
| 13 | Pentadecanal | 0.47 | Spicy note | ||||
| 14 | Hexadecanal | 0.11 | Weak floral, waxy aroma | ||||
| 15 | 8-Nonen-2-one | 0.21 | |||||
| 16 | Ketones (Total) | 0.39 | 0.17 | 0.15 | 0.08 | ||
| 17 | Phenethyl acetate | 1.72 | 1.36 | 6.96 | 2.37 | 4.73 | Rose, honey-like fragrance |
| 18 | Ethyl octanoate | 0.71 | 1.08 | 0.94 | Brandy-like aroma | ||
| 19 | Decanal | 0.05 | Citrus peel-like odor | ||||
| 20 | Ethyl decanoate | 0.54 | 0.42 | 0.75 | 1.08 | Alcoholic, sweet, floral notes | |
| 21 | 2,3-Butanediol | 0.11 | 0.16 | 0.07 | Fruity, creamy notes | ||
| 22 | 2,4-Di-tert-butylphenol | 3.17 | 6.07 | 5.38 | plastic-like or medicinal off-odors | ||
| 23 | Citronellol | 0.29 | 0.10 | 0.19 | Fresh green rose-like aroma | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Sun, Y.; Chen, R.; Xu, R.; Li, Q.; Liu, X.; Li, L.; Zhao, T. Staged Fermentation with a Saccharomyces cerevisiae Consortium and Monascus purpureus for Sustainable Valorization of Persimmon into High-Quality Wine. Fermentation 2025, 11, 278. https://doi.org/10.3390/fermentation11050278
Du Y, Sun Y, Chen R, Xu R, Li Q, Liu X, Li L, Zhao T. Staged Fermentation with a Saccharomyces cerevisiae Consortium and Monascus purpureus for Sustainable Valorization of Persimmon into High-Quality Wine. Fermentation. 2025; 11(5):278. https://doi.org/10.3390/fermentation11050278
Chicago/Turabian StyleDu, Yamin, Yangyang Sun, Rongqi Chen, Ruoxuan Xu, Qing Li, Xiuhe Liu, Lanxiao Li, and Tao Zhao. 2025. "Staged Fermentation with a Saccharomyces cerevisiae Consortium and Monascus purpureus for Sustainable Valorization of Persimmon into High-Quality Wine" Fermentation 11, no. 5: 278. https://doi.org/10.3390/fermentation11050278
APA StyleDu, Y., Sun, Y., Chen, R., Xu, R., Li, Q., Liu, X., Li, L., & Zhao, T. (2025). Staged Fermentation with a Saccharomyces cerevisiae Consortium and Monascus purpureus for Sustainable Valorization of Persimmon into High-Quality Wine. Fermentation, 11(5), 278. https://doi.org/10.3390/fermentation11050278