
Exploiting 1,3-Propanediol Production by a Clostridium beijerinckii Strain: The Role of Glycerol and Ammonium Sulfate Concentrations
 , ,
, ,
Abstract
1. Introduction
2. Material and Methods
2.1. Microorganism and Inoculum
2.2. Culture Medium
2.3. Central Composite Rotational Design (CCRD)
2.4. Comparison of Culture Media in Kinetic Assays
2.5. Absolute Gene Expression Level
2.6. Analytical Determination
2.7. Statistical Validation
3. Results and Discussions
3.1. Optimization of Glycerol and (NH4)2SO4 Concentrations
3.2. Comparative Assay to Assess the Impact of Media Optimization on Product Formation, Substrate Consumption, and the Expression of Key Metabolic Enzymes
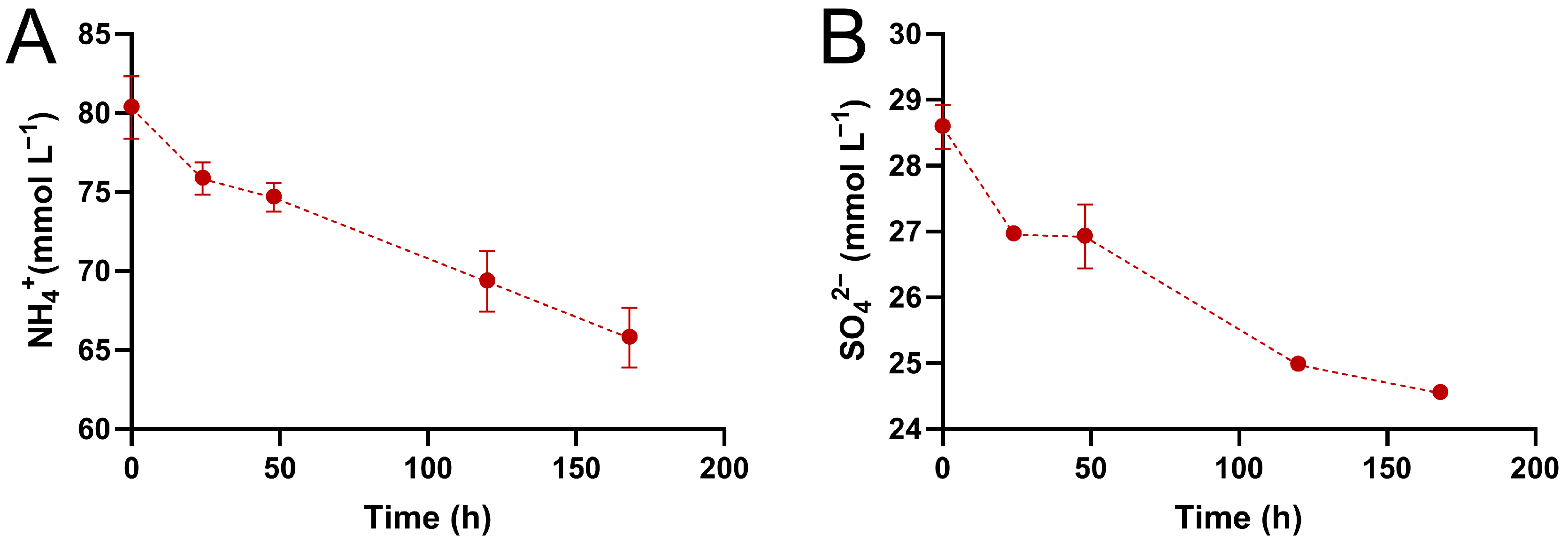
3.3. Sulfate and Ammonium Consumption
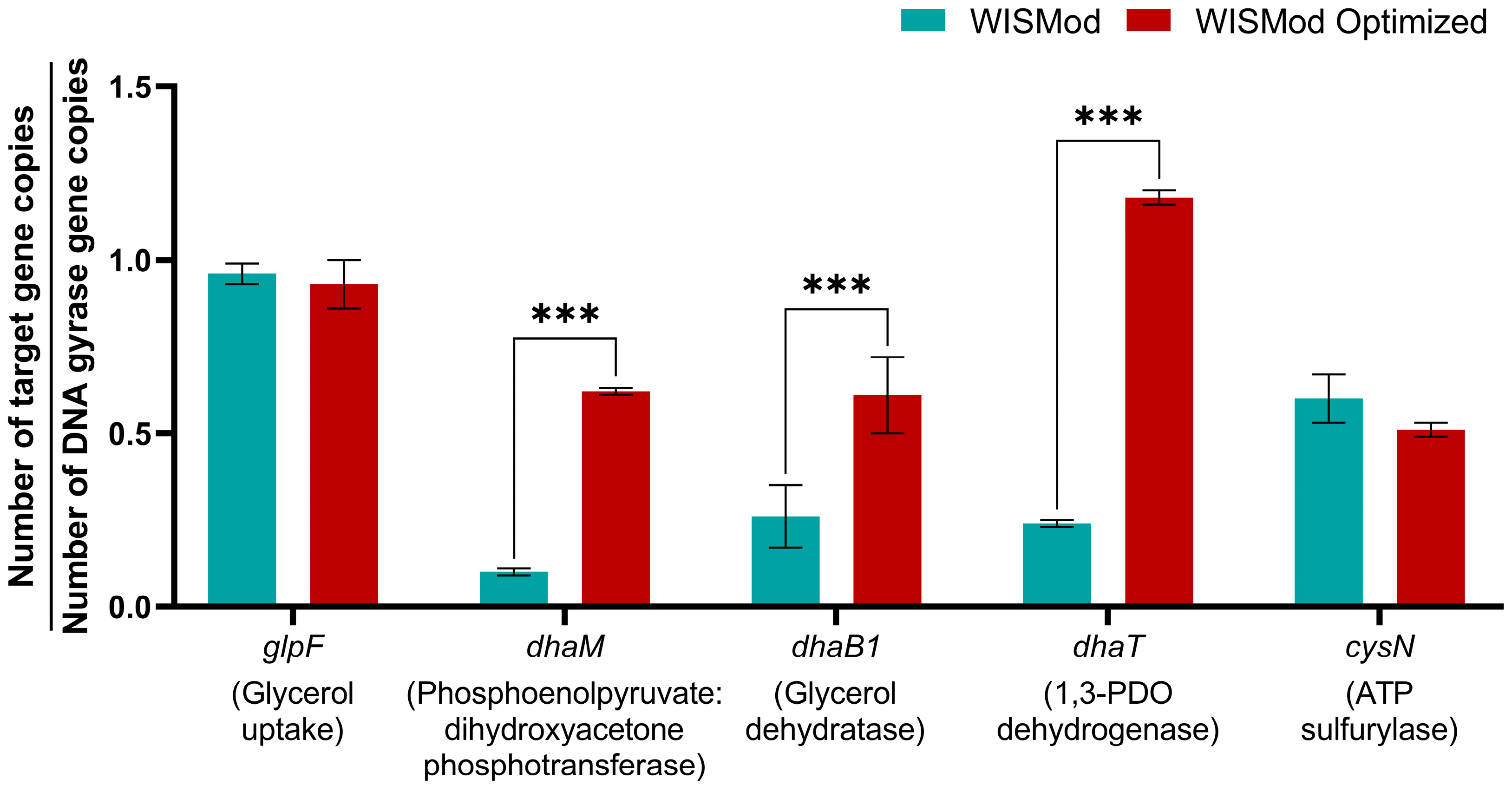
3.4. Gene Expression
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shahzad, S.J.H.; Bouri, E.; Kayani, G.M.; Nasir, R.M.; Kristoufek, L. Are clean energy stocks efficient? Asymmetric multifractal scaling behaviour. Phys. A Stat. Mech. Appl. 2020, 550, 124519. [Google Scholar] [CrossRef]
- Quah, R.V.; Tan, Y.H.; Mubarak, N.M.; Khalid, M.; Abdullah, E.C.; Nolasco-Hipolito, C. An overview of biodiesel production using recyclable biomass and non-biomass derived magnetic catalysts. J. Environ. Chem. Eng. 2019, 7, 103219. [Google Scholar]
- OECD-FAO Agricultural Outlook 2022–2031. Available online: https://www.oecd.org/en/publications/oecd-fao-agricultural-outlook-2022-2031_f1b0b29c-en.html (accessed on 27 February 2025).
- Analysis of Biofuels’ Current Outlook 2023. Available online: https://www.epe.gov.br/en/publications/publications/analysis-of-biofuels-current-outlook-2023 (accessed on 27 February 2025).
- Nomanbhay, S.; Hussein, R.; Ong, M.Y. Sustainability of biodiesel production in Malaysia by production of bio-oil from crude glycerol using microwave pyrolysis: A review. Green Chem. Lett. Rev. 2018, 11, 135–157. [Google Scholar]
- Wang, H.; Li, H.; Lee, C.K.; Mat Nanyan, N.S.; Tay, G.S. A systematic review on utilization of biodiesel-derived crude glycerol in sustainable polymers preparation. Int. J. Biol. Macromol. 2024, 216, 129536. [Google Scholar]
- Chilakamarry, C.R.; Sakinah, A.M.M.; Zularisam, A.W.; Pandey, A. Glycerol waste to value added products and its potential applications. Syst. Microbiol. Biomanuf. 2021, 1, 378–396. [Google Scholar] [CrossRef]
- Maru, B.T.; Bielen, A.A.M.; Constantí, M.; Medina, F.; Kengen, S.W.M. Glycerol fermentation to hydrogen by Thermotoga maritima: Proposed pathway and bioenergetic considerations. Int. J. Hydrogen Energy 2013, 38, 5563–5572. [Google Scholar]
- Seta, K.; Suzuki, T.; Kiyoshi, K.; Shigeno, T.; Nakajima-Kambe, T. Potential use of methane fermentation digested slurry as a low-cost, environmentally-friendly nutrient for bioethanol production from crude glycerol by Klebsiella variicola TB-83D. New Biotechnol. 2018, 44, 1–5. [Google Scholar]
- Sarchami, T.; Johnson, E.; Rehmann, L. Optimization of fermentation condition favoring butanol production from glycerol by Clostridium pasteurianum DSM 525. Bioresour. Technol. 2016, 208, 73–80. [Google Scholar] [PubMed]
- Mermejo, B.d.C.; Bortolucci, J.; de Andrade, A.R.; Reginatto, V. The Non-solventogenic Clostridium beijerinckii Br21 Produces 1,3-Propanediol From Glycerol With Butyrate as the Main By-Product. Front. Sustain. Food Syst. 2022, 6, 848022. [Google Scholar]
- Marr, A.C. 1,3-Propanediol, an Exemplary Bio-Renewable Organic Platform Chemical. Adv. Synth. Catal. 2024, 366, 4835–4845. [Google Scholar]
- Global 1,3 Propanediol Market Report, Trends, Size and Share Analysis-Growth Trends and Forecasts (2024–2031). Available online: https://www.coherentmarketinsights.com/industry-reports/global-1-3-propanediol-market (accessed on 27 February 2025).
- Johnson, E.E.; Rehmann, L. The role of 1,3-propanediol production in fermentation of glycerol by Clostridium pasteurianum. Bioresour. Technol. 2016, 209, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wischral, D.; Zhang, J.; Cheng, C.; Lin, M.; De Souza, L.M.G.; Pessoa, F.L.P.; Pereira, N.; Yang, S.T. Production of 1,3-propanediol by Clostridium beijerinckii DSM 791 from crude glycerol and corn steep liquor: Process optimization and metabolic engineering. Bioresour. Technol. 2016, 212, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Apiwatanapiwat, W.; Vaithanomsat, P.; Thanapase, W.; Ratanakhanokchai, K.; Kosugi, A. Xylan supplement improves 1,3-propanediol fermentation by Clostridium butyricum. J. Biosci. Bioeng. 2018, 125, 662–668. [Google Scholar] [CrossRef]
- Silva, G.P.D.; Contiero, J.; Neto, P.M.Á.; De Lima, C.J.B. 1,3-propanediol: Production, applications and biotechnological potential. Quím. Nova 2014, 37, 527–534. [Google Scholar] [CrossRef]
- Gallardo, R.; Alves, M.; Rodrigues, L.R. Modulation of crude glycerol fermentation by Clostridium pasteurianum DSM 525 towards the production of butanol. Biomass Bioenerg. 2014, 71, 134–143. [Google Scholar] [CrossRef]
- Leng, L.; Nobu, M.K.; Narihiro, T.; Yang, P.; Amy Tan, G.Y.; Lee, P.H. Shaping microbial consortia in coupling glycerol fermentation and carboxylate chain elongation for Co-production of 1,3-propanediol and caproate: Pathways and mechanisms. Water Res. 2019, 148, 281–291. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, D. Toward glycerol biorefinery: Metabolic engineering for the production of biofuels and chemicals from glycerol. Biotechnol. Biofuels Bioprod. 2016, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Fokum, E.; Zabed, H.M.; Ravikumar, Y.; Elshobary, M.E.; Chandankere, R.; Zhang, Y.; Yun, J.; Qi, X. Co-fermentation of glycerol and sugars by Clostridium beijerinckii: Enhancing the biosynthesis of 1,3-propanediol. Food Biosci. 2021, 2021, 101028. [Google Scholar] [CrossRef]
- Altafini, R.d.M.; Martins, T.M.T.; Bruni, A.T.; Reginatto, V. Upgraded medium composition highlights the relevance of iron sulfate for 1,3-propanediol production by a Clostridium beijerinckii strain. Biocatal. Agric. Biotechnol. 2022, 43, 102388. [Google Scholar] [CrossRef]
- Lee, S.; Lee, J.H.; Mitchell, R.J. Analysis of Clostridium beijerinckii NCIMB 8052’s transcriptional response to ferulic acid and its application to enhance the strain tolerance. Biotechnol. Biofuels Bioprod. 2015, 8, 68. [Google Scholar] [CrossRef]
- Fonseca, B.C.; Guazzaroni, M.E.; Reginatto, V. Fermentative production of H2 from different concentrations of galactose by the new isolate Clostridium beijerinckii Br21. Int. J. Hydrogen Energy 2016, 41, 21109–21120. [Google Scholar] [CrossRef]
- Egoburo, D.E.; Diaz Peña, R.; Kolender, A.; Pettinari, M.J. Optimization and Validation of a GC–FID Method for Quantitative Determination of 1,3-Propanediol in Bacterial Culture Aqueous Supernatants Containing Glycerol. Chromatographia 2017, 80, 1121–1127. [Google Scholar]
- Akagi, J.M.; Campbell, L.L. Studies on Thermophilic Sulfate-Reducing Bacteria III: Adenosine Triphosphate-sulfurylase of Clostridium nigrificans and Desulfovibrio desulfuricans. J. Bacteriol. 1962, 84, 1194–1201. [Google Scholar] [PubMed]
- Kováč, J.; Vítězová, M.; Kushkevych, I. Metabolic activity of sulfate-reducing bacteria from rodents with colitis. Open Med. 2018, 13, 344. [Google Scholar] [CrossRef] [PubMed]
- Kushkevych, I.; Vítězová, M.; Vítěz, T.; Kováč, J.; Kaucká, P.; Jesionek, W.; Bartoš, M.; Barton, L. A new combination of substrates: Biogas production and diversity of the methanogenic microorganisms. Open Life Sci. 2018, 13, 119–128. [Google Scholar]
- Borgnia, M.J.; Agre, P. Reconstitution and functional comparison of purified GlpF and AqpZ, the glycerol and water channels from Escherichia coli. Proc. Natl. Acad. Sci. USA 2001, 98, 2888–2893. [Google Scholar]
- Stroud, R.M.; Nollert, P.; Miercke, L. The glycerol facilitator GlpF its aquaporin family of channels, and their selectivity. Adv. Protein Chem. 2003, 63, 291–316. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Primers (5′ 3′) | Reporter 1 (5′ 3′) |
|---|---|---|
| DNA gyrase (normalizer) | FW: AGATGTTGTGATAACATTAACACATTCAGGAT R: GCTTGAATTCCTTTTCCACCTCTTC | TCTGCCGAAATTCTT |
| Glycerol Uptake | FW: CCAGCATTAACAATTGCGTTAGCA R: TGCACCAATACATTGTCCAGCTAAA | CCCATGCCACTTTACC |
| Glycerol dehydratase | FW: TGGATTAAATTAAATGATATTAATAAAGTAAGAGATGAGATTTCAACT R: TCTGAATTTTGCCCTCCAACGATTA | ATAGCCGCCAAAATG |
| Phosphoenolpyruvate: Dihydroxyacetone phosphotransferase | FW: CAGAGTAAACATCTTCTATTGCAGCTGAT R: GGCTGGAGGAACGTCAGAT | CAGTGCCAACTCTTCC |
| 1,3-Propanediol dehydrogenase | FW: GGACGCTTTAACTCATGCAATTGAA R: CTTGTATTGCTGAAGCATCTGTCA | CCGGATTTGCATCTTT |
| ATP sulfurylase (Adenosine Triphosphate sulfurylase) | FW: GACCAAATCATACTTTCAGAGGATTCCA R: GGAAGTGTCGTAATTTCATCTCCTACA | TTCCAGCTTCAATTTC |
| Conditions | Results | ||||||
|---|---|---|---|---|---|---|---|
| Conditions | Glycerol (mmol L−1) | (NH4)2SO4 (mmol L−1) | Final pH | OD600 | Consumed Glycerol (mmol L−1) | Butyric Acid (mmol L−1) | 1,3-PDO (mmol L−1) |
| 1 | 200 (−1) | 7.6 (−1) | 5.06 ± 0.10 | 2.09 ± 0.18 | 109.5 ± 6.38 | 18.29 ± 1.13 | 37.41 ± 3.73 |
| 2 | 200 (−1) | 22.7 (+1) | 5.46 ± 0.05 | 2.01 ± 0.30 | 152.80 ± 8.97 | 10.22 ± 0.01 | 21.30 ± 1.12 |
| 3 | 400 (+1) | 7.6 (−1) | 5.29 ± 0.06 | 2.06 ± 0.16 | 181.51 ± 14.58 | 19.57 ± 1.84 | 76.27 ± 7.34 |
| 4 | 400 (+1) | 22.7 (+1) | 5.16 ± 0.04 | 2.36 ± 0.11 | 185.62 ± 5.62 | 18.96 ± 1.26 | 53.88 ± 5.85 |
| 5 | 158 (−1.41) | 15.1 (0) | 6.03 ± 0.01 | 1.74 ± 0.00 | 51.36 ± 1.87 | 6.03 ± 0.01 | 3.44 ± 2.31 |
| 6 | 441 (+1.41) | 15.1 (0) | 5.49 ± 0.08 | 2.11 ± 0.08 | 157.42 ± 12.62 | 15.51 ± 0.66 | 41.89 ± 1.28 |
| 7 | 300 (0) | 4.4 (−1.41) | 5.15 ± 0.02 | 2.20 ± 0.23 | 164.54 ± 1.83 | 17.27 ± 0.29 | 50.98 ± 0.67 |
| 8 | 300 (0) | 25.8 (+1.41) | 5.16 ± 0.13 | 2.53 ± 0.11 | 183.32 ± 10.26 | 20.34 ± 2.56 | 69.18 ± 2.91 |
| 9 | 300 (0) | 15.1 (0) | 5.26 ± 0.02 | 2.04 ± 0.04 | 135.51 ± 13.76 | 15.84 ± 0.89 | 38.33 ± 0.66 |
| 10 | 300 (0) | 15.1 (0) | 5.39 ± 0.30 | 2.10 ± 0.25 | 123.72 ± 19.97 | 12.68 ± 0.66 | 35.70 ± 6.79 |
| Factors | Significance | Sum of Squares | Degrees of Freedom | Mean Square | F | p |
|---|---|---|---|---|---|---|
| Linear glycerol | Significant | 1909.76 | 1 | 1909.76 | 1699.83 | 0.015 |
| Quadratic glycerol | Significant | 235.04 | 1 | 235.04 | 209.20 | 0.044 |
| Quadratic (NH4)2SO4 | Significant | 561.29 | 1 | 561.29 | 499.59 | 0.028 |
| Lack of fit | Not significant | 372.37 | 5 | 74.47 | 66.29 | 0.093 |
| Pure error | 1.12 | 1 | 1.12 | |||
| Total | 3634.43 | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Castro, P.F.D.L.; Altafini, R.d.M.; Bortolucci, J.; Florêncio, J.; Campos, M.L.A.M.; Reginatto, V. Exploiting 1,3-Propanediol Production by a Clostridium beijerinckii Strain: The Role of Glycerol and Ammonium Sulfate Concentrations. Fermentation 2025, 11, 187. https://doi.org/10.3390/fermentation11040187
de Castro PFDL, Altafini RdM, Bortolucci J, Florêncio J, Campos MLAM, Reginatto V. Exploiting 1,3-Propanediol Production by a Clostridium beijerinckii Strain: The Role of Glycerol and Ammonium Sulfate Concentrations. Fermentation. 2025; 11(4):187. https://doi.org/10.3390/fermentation11040187
Chicago/Turabian Stylede Castro, Pedro Felipe Dassie Leonel, Rafael de Moraes Altafini, Jonatã Bortolucci, Jaques Florêncio, Maria Lucia Arruda Moura Campos, and Valeria Reginatto. 2025. "Exploiting 1,3-Propanediol Production by a Clostridium beijerinckii Strain: The Role of Glycerol and Ammonium Sulfate Concentrations" Fermentation 11, no. 4: 187. https://doi.org/10.3390/fermentation11040187
APA Stylede Castro, P. F. D. L., Altafini, R. d. M., Bortolucci, J., Florêncio, J., Campos, M. L. A. M., & Reginatto, V. (2025). Exploiting 1,3-Propanediol Production by a Clostridium beijerinckii Strain: The Role of Glycerol and Ammonium Sulfate Concentrations. Fermentation, 11(4), 187. https://doi.org/10.3390/fermentation11040187