

The Effects of Different Doses of 3-NOP on Ruminal Fermentation Parameters, Methane Production, and the Microbiota of Lambs In Vitro


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diet, and Experimental Design
2.2. In Vitro Incubation
2.3. Sample Collection and Measurements
2.4. DNA Extraction, Real-Time PCR (qPCR) Analysis, Sequencing, and Diversity Analysis
2.5. Statistical Analysis
3. Results
3.1. The Effects of 3-NOP on Gas Production and the Degradability of the Substrate
3.2. The Effects of 3-NOP on In Vitro Fermentation Characteristics and Microbial Profile
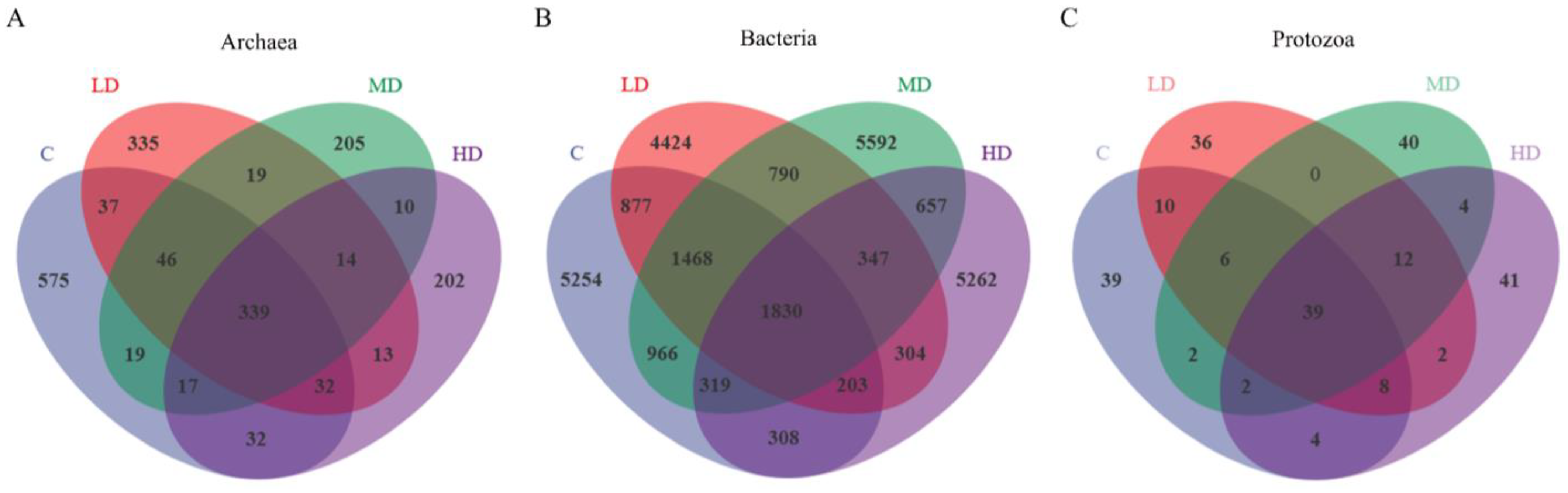
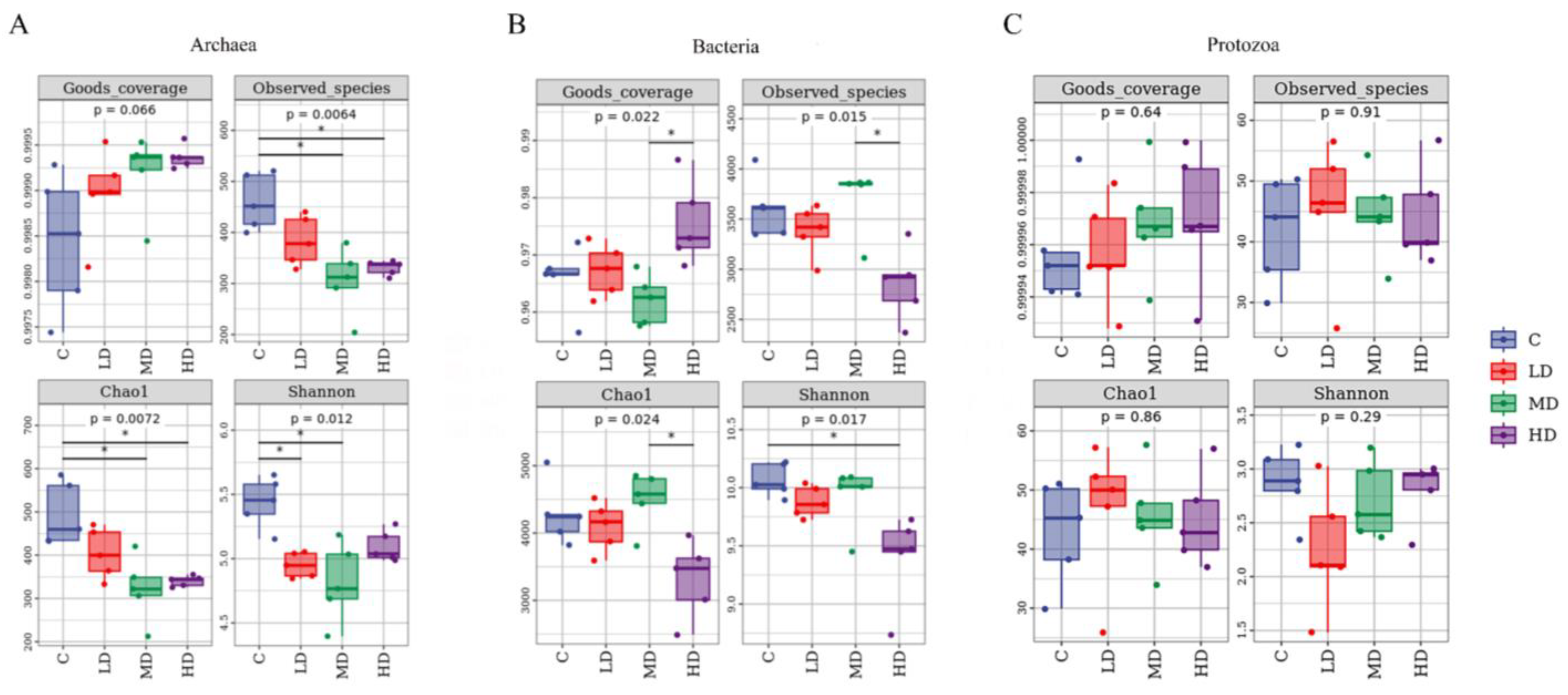
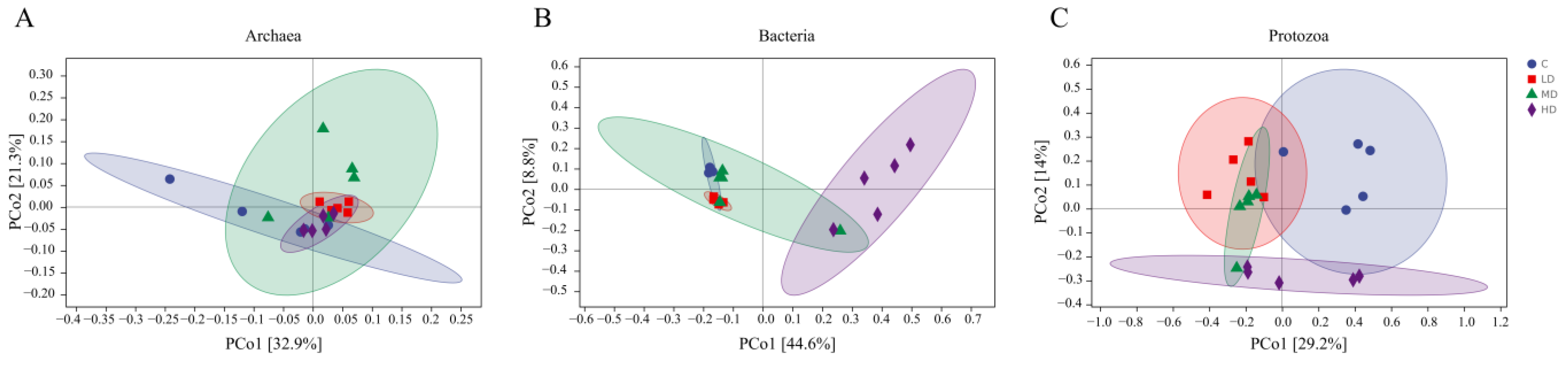
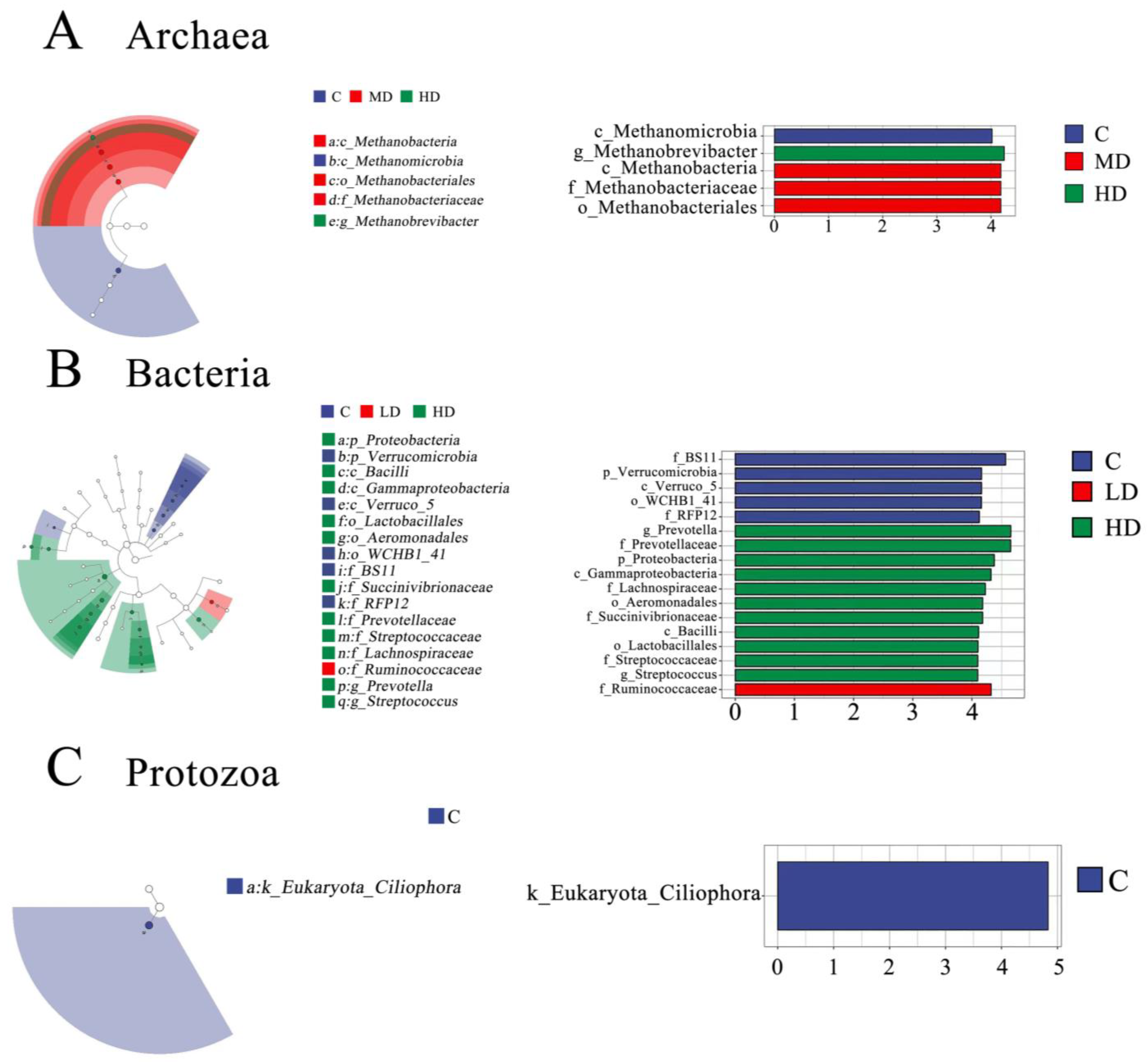
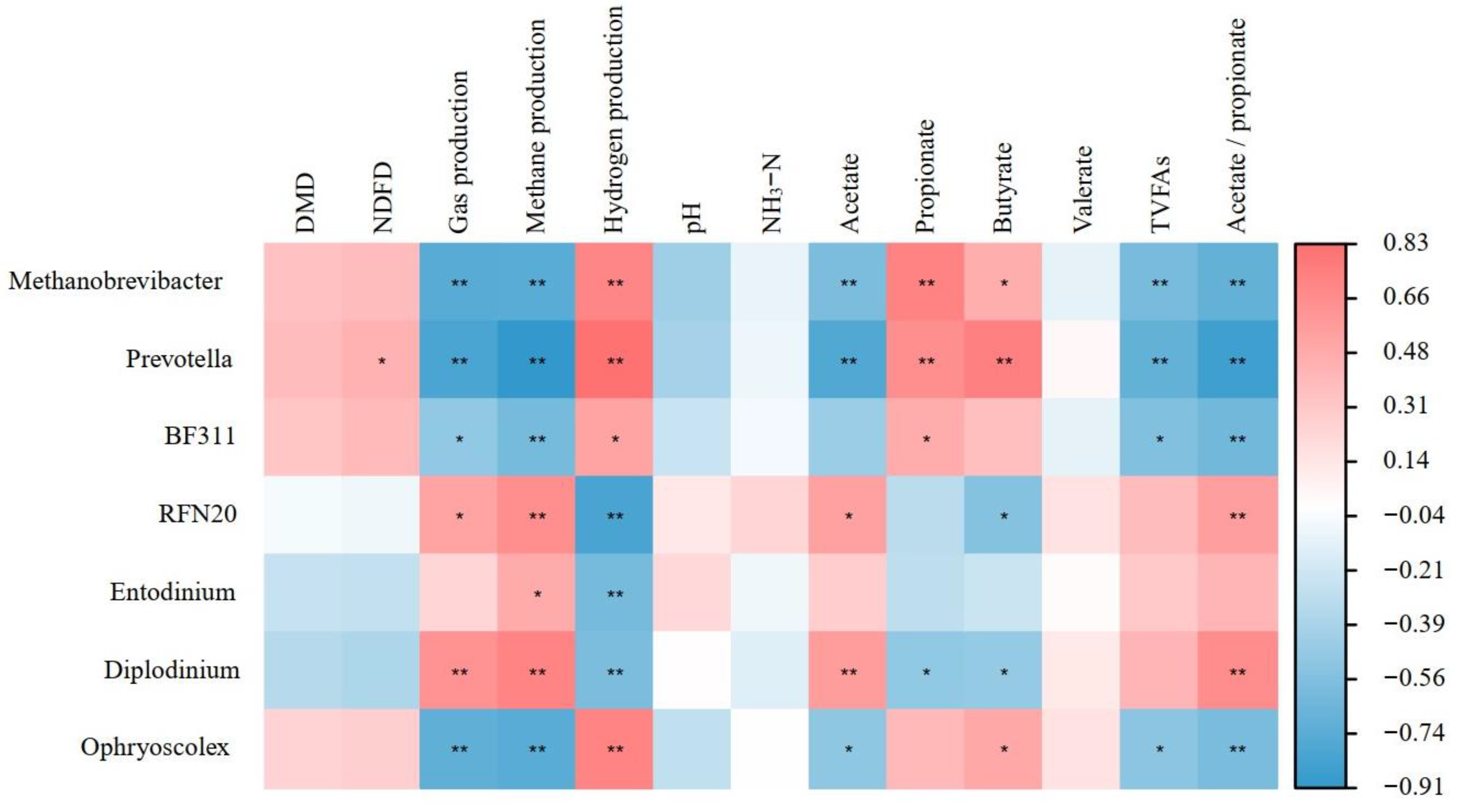
3.3. The Effects of 3-NOP on the Incubation Microbiota
4. Discussion
4.1. The Effects of 3-NOP on Ruminal Fermentation Parameters
4.2. The Effects of Different Doses of 3-NOP on Ruminal Microbiota
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Giallongo, F.; Frederick, T.W.; Harper, M.T.; Weeks, H.L.; Branco, A.F.; Moate, P.J.; Deighton, M.H.; Williams, S.R.O.; et al. An inhibitor persistently decreased enteric methane emission from dairy cows with no negative effect on milk production. Proc. Natl. Acad. Sci. USA 2015, 11, 10663–10668. [Google Scholar] [CrossRef]
- Duin, E.C.; Wagner, T.; Shima, S.; Prakash, D.; Cronin, B.; Yáñez-Ruiz, D.R.; Duval, S.; Rümbeli, R.; Stemmler, R.T.; Thauer, R.K. Mode of action uncovered for the specific reduction of methane emissions from ruminants by the small molecule 3-nitrooxypropanol. Proc. Natl. Acad. Sci. USA 2016, 113, 6172–6177. [Google Scholar] [CrossRef]
- Romero-Pérez, A.; Okine, E.K.; Guan, L.L.; Duval, S.M.; Kindermann, M.; Beauchemin, K.A. Effects of 3-nitrooxypropanol on methane production using the rumen simulation technique (Rusitec). Anim. Feed Sci. Technol. 2015, 209, 98–109. [Google Scholar] [CrossRef]
- Schilde, M.; von Soosten, D.; Hüther, L.; Kersten, S.; Meyer, U.; Zeyner, A.; Dänicke, S. Dose–response effects of 3-nitrooxypropanol combined with low-and high-concentrate feed proportions in the dairy cow ration on fermentation parameters in a rumen simulation technique. Animals 2021, 11, 1784. [Google Scholar] [CrossRef]
- Dijkstra, J.; Bannink, A.; France, J.; Kebreab, E.; van Gastelen, S. Short communication: Antimethanogenic effects of 3-nitrooxypropanol depend on supplementation dose, dietary fiber content, and cattle type. J. Dairy Sci. 2018, 101, 9041–9047. [Google Scholar] [CrossRef]
- Melgar, A.; Welter, K.C.; Nedelkov, K.; Martins, C.M.M.R.; Harper, M.T.; Oh, J.; Räisänen, S.E.; Chen, X.J.; Cueva, S.F.; Duval, S.; et al. Dose-response effect of 3-nitrooxypropanol on enteric methane emissions in dairy cows. J. Dairy Sci. 2020, 103, 6145–6156. [Google Scholar] [CrossRef]
- Vyas, D.; McGinn, S.M.; Duval, S.M.; Kindermann, M.K.; Beauchemin, K.A. Optimal dose of 3-nitrooxypropanol for decreasing enteric methane emissions from beef cattle fed high-forage and high-grain diets. Anim. Prod. Sci. 2016, 58, 1049–1055. [Google Scholar] [CrossRef]
- Yu, G.; Beauchemin, K.A.; Dong, R. A review of 3-nitrooxypropanol for enteric methane mitigation from ruminant livestock. Animals 2021, 11, 3540. [Google Scholar] [CrossRef]
- Haisan, J.; Sun, Y.; Guan, L.L.; Beauchemin, K.A.; Iwaasa, A.; Duval, S.; Barreda, D.R.; Oba, M. The effects of feeding 3-nitrooxypropanol on methane emissions and productivity of Holstein cows in mid lactation. J. Dairy Sci. 2014, 97, 3110–3119. [Google Scholar] [CrossRef]
- Romero-Pérez, A.; Okine, E.K.; Guan, L.L.; Duval, S.M.; Kindermann, M.; Beauchemin, K.A. Rapid Communication: Evaluation of methane inhibitor 3-nitrooxypropanol and monensin in a high-grain diet using the rumen simulation technique (Rusitec). J. Anim. Sci. 2017, 95, 4072–4077. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Fernández, G.; Duval, S.; Kindermann, M.; Schirra, H.J.; Denman, S.E.; McSweeney, C.S. 3-NOP vs. halogenated compound: Methane production, ruminal fermentation and microbial community response in forage fed cattle. Front. Microbiol. 2018, 9, 1582. [Google Scholar] [CrossRef]
- Gruninger, R.J.; Zhang, X.M.; Smith, M.L.; Kung, L.J.; Vyas, D.; McGinn, S.M.; Kindermann, M.; Wang, M.; Tan, Z.L.; Beauchemin, K.A. Application of 3-nitrooxypropanol and canola oil to mitigate enteric methane emissions of beef cattle results in distinctly different effects on the rumen microbial community. Anim. Microbiome 2022, 4, 35. [Google Scholar] [CrossRef]
- Pitta, D.W.; Melgar, A.; Hristov, A.N.; Indugu, N.; Narayan, K.S.; Pappalardo, C. Temporal changes in total and metabolically active ruminal methanogens in dairy cows supplemented with 3-nitrooxypropanol. J. Dairy. Sci. 2021, 104, 8721–8735. [Google Scholar] [CrossRef]
- Pitta, D.W.; Indugu, N.; Melgar, A.; Hristov, A.N.; Challa, K.; Vecchiarelli, B.; Hennessy, M.L.; Narayan, K.S.; Duval, S.; Kindermann, M.; et al. The effect of 3-nitrooxypropanol, a potent methane inhibitor, on ruminal microbial gene expression profiles in dairy cows. Microbiome 2022, 10, 1–21. [Google Scholar] [CrossRef]
- Martínez-Fernández, G.; Abecia, L.; Arco, A.; Cantalapiedra-Hijar, G.; Martín-García, A.I.; Molina-Alcaide, E.; Kindermann, M.; Duval, S.; Yáñez-Ruíz, D.R. Effects of ethyl-3-nitrooxy propionate and 3-nitrooxypropanol on ruminal fermentation, microbial abundance, and methane emissions in sheep. J. Dairy Sci. 2014, 97, 3790–3799. [Google Scholar] [CrossRef]
- Romero-Pérez, A.; Okine, E.K.; McGinn, S.M.; Guan, L.L.; Oba, M.; Duval, S.; Kindermann, M.; Beauchemin, K.A. The potential of 3-nitrooxypropanol to lower enteric methane emissions from beef cattle. J. Anim. Sci. 2014, 92, 4682–4693. [Google Scholar] [CrossRef]
- Haisan, J.; Sun, Y.; Guan, L.; Beauchemin, K.A.; Iwaasa, A.; Duval, S.; Kindermann, M.; Barreda, D.R.; Oba, M. The effects of feeding 3-nitrooxypropanol at two doses on milk production, rumen fermentation, plasma metabolites, nutrient digestibility, and methane emissions in lactating Holstein cows. Anim. Prod. Sci. 2016, 57, 282–289. [Google Scholar] [CrossRef]
- Liu, Z.H.; Wang, K.; Nan, X.M.; Yang, L.G.; Wang, Y.; Zhang, F.; Cai, M.; Zhao, Y.G.; Xiong, B.H. Effects of combined addition of 3-nitrooxypropanol and vitamin B12 on methane and propionate production in dairy cows by in vitro-simulated fermentation. J. Dairy. Sci. 2023, 106, 219–232. [Google Scholar] [CrossRef]
- Liu, Z.H.; Wang, K.; Nan, X.M.; Cai, M.; Yang, L.; Xiong, B.H.; Zhao, Y.G. Synergistic effects of 3-Nitrooxypropanol with fumarate in the regulation of propionate formation and methanogenesis in dairy cows in vitro. Appl. Environ. Microbiol. 2022, 88, e0190821. [Google Scholar] [CrossRef]
- van Gastelen, S.; Burgers, E.E.A.; Dijkstra, J.; de Mol, R.; Muizelaar, W.; Walker, N.; Bannink, A. Long-term effects of 3-nitrooxypropanol on methane emission and milk production characteristics in Holstein Friesian dairy cows. J. Dairy Sci. 2024, 107, 5556–5573. [Google Scholar] [CrossRef]
- Romero-Pérez, A.; Okine, E.K.; McGinn, S.M.; Guan, L.L.; Oba, M.; Duval, S.; Kindermann, M.; Beauchemin, K.A. Sustained reduction in methane production from long-term addition of 3-nitrooxypropanol to a beef cattle diet. J. Anim. Sci. 2015, 93, 1780–1791. [Google Scholar] [CrossRef]
- Kim, H.; Lee, H.G.; Baek, Y.C.; Lee, S.; Seo, J. The effects of dietary supplementation with 3-nitrooxypropanol on enteric methane emissions, rumen fermentation, and production performance in ruminants: A meta-analysis. J. Anim. Sci. Technol. 2020, 62, 31–42. [Google Scholar] [CrossRef]
- Kebreab, E.; Bannink, A.; Pressman, E.M.; Walker, N.; Karagiannis, A.; van Gastelen, S.; Dijkstra, J. A meta-analysis of effects of 3-nitrooxypropanol on methane production, yield, and intensity in dairy cattle. J. Dairy Sci. 2023, 106, 927–936. [Google Scholar] [CrossRef]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. Isme J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Kelly, W.J.; Janssen, P.H.; Attwood, G.T. Rumen microbial (meta) genomics and its application to ruminant production. Animal 2013, 7, 184–201. [Google Scholar] [CrossRef]
- Yáñez-Ruiz, D.R.; Abecia, L.; Newbold, C.J. Manipulating rumen microbiome and fermentation through interventions during early life: A review. Front. Microbiol. 2015, 6, 1133. [Google Scholar] [CrossRef]
- Abecia, L.; Waddams, K.E.; Martínez-Fernandez, G.; Martín-García, A.I.; Ramos-Morales, E.; Newbold, C.J.; Yáñez-Ruiz, D.R. An antimethanogenic nutritional intervention in early life of ruminants modifies ruminal colonization by Archaea. Archaea 2014, 2014, 841463. [Google Scholar] [CrossRef]
- Meale, S.J.; Popova, M.; Saro, C.; Martin, C.; Bernard, A.; Lagrée, M.; Yáñez-Ruíz, D.R.; Boudra, H.; Duval, S.; Morgavi, D.P. Early life dietary intervention in dairy calves results in a long-term reduction in methane emissions. Sci. Rep. 2011, 11, 3003. [Google Scholar] [CrossRef]
- NY/T 816-2021; Nutrient Requirements of Meat-Type Sheep and Goat. Ministry of Agriculture of the PRC. China Agriculture Press: Beijing, China, 2021. (In Chinese)
- Menke, K.H.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feeding stuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef]
- Kim, W.Y.; Hanigan, M.D.; Lee, S.J.; Lee, S.M.; Kim, D.H.; Hyun, J.H.; Yeo, J.M. Effects of cordyceps militaris on the growth of rumen microorganisms and in vitro rumen fermentation with respect to methane emissions. J. Dairy Sci. 2014, 97, 7065–7075. [Google Scholar] [CrossRef] [PubMed]
- Morsy, A.S.; Soltan, Y.A.; Sallam, S.M.A.; Kreuzer, M.; Alencar, S.M.; Abdalla, A.L. Comparison of the in vitro efficiency of supplementary bee propolis of different origin in enhancing ruminal nutrient degradation and mitigating methane formation. Anim. Feed. Sci. Technol. 2015, 199, 51–60. [Google Scholar] [CrossRef]
- Soltan, Y.A.; Morsy, A.S.; Lucas, R.C.; Abdalla, A.L. Potential of mimosine of Leucaena leucocephala for modulating ruminal nutrient degradability and methanogenesis. Anim. Feed. Sci. Technol. 2017, 223, 30–41. [Google Scholar] [CrossRef]
- Nolan, J.V.; Hegarty, R.S.; Hegarty, J.; Godwin, I.R.; Woodgate, R. Effects of dietary nitrate on fermentation, methane production and digesta kinetics in sheep. Anim. Prod. Sci. 2010, 50, 801–806. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Kim, H.; Jung, E.; Lee, H.G.; Kim, B.; Cho, S.; Lee, S.; Kwon, I.; Seo, J. Essential oil mixture on rumen fermentation and microbial community-an in vitro study. Asian Austral J. Anim. 2019, 32, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Shen, Y.B.; Ma, H.; Li, Y.; Lambo, M.T.; Dai, B.S.; Shen, W.Z.; Qu, Y.L.; Zhang, Y.G. Silibinin reduces in vitro methane production by regulating the rumen microbiome and metabolites. Front. Microbiol. 2023, 14, 1225643. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Analytical Chemists: Gaithersburg, MD, USA, 1995. [Google Scholar]
- van Soest, P.V.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Huse, S.M.; Dethlefsen, L.; Huber, J.A.; Welch, D.M.; Relman, D.A.; Sogin, M.L. Exploring microbial diversity and taxonomy using SSU rRNA hypervariable tag sequencing. PLoS Genet. 2008, 4, e1000255. [Google Scholar] [CrossRef]
- Pires, A.C.C.; Cleary, D.F.R.; Almeida, A.; Cunha, Â.; Dealtry, S.; Mendonça-Hagler, L.C.; Smalla, K.; Gomes, N.C.M. Denaturing gradient gel electrophoresis and barcoded pyrosequencing reveal unprecedented archaeal diversity in mangrove sediment and rhizosphere samples. AEM 2012, 78, 5520–5528. [Google Scholar] [CrossRef]
- Sylvester, J.T.; Karnati, S.K.R.; Yu, Z.T.; Morrison, M.; Firkins, J.L. Development of an assay to quantify rumen ciliate protozoal biomass in cows using real-time PCR. J. Nutr. 2004, 134, 3378–3384. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic. Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.X.; Zhang, B.; Chen, Z.; Qin, W.T.; Wen, X.H. Sludge retention time affects the microbial community structure: A large-scale sampling of aeration tanks throughout China. Environ. Pollut. 2020, 261, 114140. [Google Scholar] [CrossRef] [PubMed]
- Guyader, J.; Ungerfeld, E.M.; Beauchemin, K.A. Redirection of metabolic hydrogen by inhibiting methanogenesis in the rumen simulation technique (RUSITEC). Front. Microbiol. 2017, 8, 393. [Google Scholar] [CrossRef]
- Romero-Pérez, A.; Okine, E.K.; Guan, L.L.; Duval, S.M.; Kindermann, M.; Beauchemin, K.A. Effects of 3-nitrooxypropanol and monensin on methane production using a forage-based diet in Rusitec fermenters. Anim. Feed. Sci. Technol. 2016, 220, 67–72. [Google Scholar] [CrossRef]
- Pereira, D.H.; Pereira, O.G.; da Silva, B.C.; Leão, M.I.; de Campos Valadares Filho, S.; Chizzotti, M.L.; García, R. Intake and total and partial digestibility of nutrients, ruminal pH and ammonia concentration and microbial efficiency in beef cattle fed with diets containing sorghum (Sorghum bicolor (L.) Moench) silage and concentrate in different ratios. Livest. Sci. 2007, 107, 53–61. [Google Scholar] [CrossRef]
- Ali, C.S.; Khaliq, T.; Sarwar, M.; Javaid, A.; Shahzad, M.A.; Nisa, M.; Zakir, S. Effect of various non protein nitrogen sources on in vitro dry matter digestibility, ammonia production, microbial growth and pH changes by rumen bacteria. Pak. Vet. J. 2009, 29, 25–30. Available online: https://www.researchgate.net/publication/26520131 (accessed on 16 May 2024).
- Bannink, A.; Kogut, J.; Dijkstra, J.; France, J.; Kebreab, E.; van Vuuren, A.M.; Tamminga, S. Estimation of the stoichiometry of volatile fatty acid production in the rumen of lactating cows. J. Theor. Biol. 2006, 238, 36–51. [Google Scholar] [CrossRef]
- Orzuna-Orzuna, J.F.; Godina-Rodríguez, J.E.; Garay-Martínez, J.R.; Granados-Rivera, L.D.; Maldonado-Jáquez, J.A.; Lara-Bueno, A. A Meta-Analysis of 3-Nitrooxypropanol Dietary Supplementation on Growth Performance, Ruminal Fermentation, and Enteric Methane Emissions of Beef Cattle. Fermentation 2024, 10, 273. [Google Scholar] [CrossRef]
- Melgar, A.; Harper, M.T.; Oh, J.; Giallongo, F.; Young, M.E.; Ott, T.L.; Duval, S.; Hristov, A.N. Effects of 3-nitrooxypropanol on rumen fermentation, lactational performance, and resumption of ovarian cyclicity in dairy cows. J. Dairy Sci. 2020, 103, 410–432. [Google Scholar] [CrossRef]
- Reynolds, C.K.; Humphries, D.J.; Kirton, P.; Kindermann, M.; Duval, S.; Steinberg, W. Effects of 3-nitrooxypropanol on methane emission, digestion, and energy and nitrogen balance of lactating dairy cows. J. Dairy Sci. 2014, 97, 3777–3789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Smith, M.C.; Gruninger, R.J.; Kung, L.; Vyas, D.; McGinn, S.M.; Kindermann, M.; Wang, M.; Tan, Z.L.; Beauchemin, K.A. Combined effects of 3-nitrooxypropanol and canola oil supplementation on methane emissions, rumen fermentation and biohydrogenation, and total tract digestibility in beef cattle. J. Anim. Sci. 2021, 99, skab081. [Google Scholar] [CrossRef] [PubMed]
- Hungate, R.E. Hydrogen as an intermediate in the rumen fermentation. Arch. Mikrobiol. 1967, 59, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Trei, J.E.; Scott, G.C.; Parish, R.C. Influence of methane inhibition on energetic efficiency of lambs. J. Anim. Sci. 1972, 34, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Sun, X.Z.; Janssen, P.H.; Tang, S.X.; Tan, Z.L. Responses of methane production and fermentation pathways to the increased dissolved hydrogen concentration generated by eight substrates in in vitro ruminal cultures. Anim. Feed. Sci. Technol. 2014, 194, 1–11. [Google Scholar] [CrossRef]
- Wang, K.; Nan, X.M.; Zhao, Y.G.; Tong, J.J.; Jiang, L.S.; Xiong, B.H. Effects of propylene glycol on in vitro ruminal fermentation, methanogenesis, and microbial community structure. J. Dairy Sci. 2021, 104, 2924–2934. [Google Scholar] [CrossRef]
- Wang, K.; Xiong, B.H.; Zhao, X. Could propionate formation be used to reduce enteric methane emission in ruminants? Sci. Total. Environ. 2023, 855, 158867. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, N.B.; Hvelplund, T.; Weisbjerg, M.R.; Nørgaard, P. Mikrobiel omsætning i formaverne. In Kvægets Ernæring og Fysiologi-Bind 1; Hvelplund, T., Nørgaard, P., Eds.; Danmarks Jordbrugsforskning, Ministeriet for Fødevarer, Landbrug og Fiskeri: Tjele, Denmark, 2003; pp. 211–235. [Google Scholar]
- Zhang, X.M.; Gruninger, R.J.; Alemu, A.W.; Wang, M.; Tan, Z.L.; Kindermann, M.; Beauchemin, K.A. 3-Nitrooxypropanol supplementation had little effect on fiber degradation and microbial colonization of forage particles when evaluated using the in situ ruminal incubation technique. J. Dairy. Sci. 2020, 103, 8986–8997. [Google Scholar] [CrossRef] [PubMed]
- Ungerfeld, E.M.; Rust, S.R.; Boone, D.R.; Liu, Y. Effects of several inhibitors on pure cultures of ruminal methanogens. J. Appl. Microbiol. 2004, 97, 520–526. [Google Scholar] [CrossRef]
- Leahy, S.C.; Kelly, W.J.; Li, D.; Li, Y.; Altermann, E.; Lambie, S.C.; Cox, F.; Attwood, G.T. The complete genome sequence of Methanobrevibacter sp. AbM4. Stand. Genomic. Sci. 2013, 8, 215–227. [Google Scholar] [CrossRef]
- Pitta, D.; Indugu, N.; Narayan, K.; Hennessy, M. Symposium review: Understanding the role of the rumen microbiome in enteric methane mitigation and productivity in dairy cows. J. Dairy Sci. 2022, 105, 8569–8585. [Google Scholar] [CrossRef] [PubMed]
- Söllinger, A.; Tveit, A.T.; Poulsen, M.; Noel, S.J.; Bengtsson, M.M.; Bernhardt, J.; Hellwing, A.L.F.; Lund, P.; Riedal, K.; Schleper, C. Holistic assessment of rumen microbiome dynamics through quantitative metatranscriptomics reveals multifunctional redundancy during key steps of anaerobic feed degradation. Msystems 2018, 3, e00038-18. [Google Scholar] [CrossRef] [PubMed]
- Söllinger, A.; Urich, T. Methylotrophic methanogens everywhere- physiology and ecology of novel players in global methane cycling. Biochem. Soc. Trans. 2019, 47, 1895–1907. [Google Scholar] [CrossRef]
- Shi, W.B.; Moon, C.D.; Leahy, S.C.; Kang, D.W.; Froula, J.; Kittelmann, S.; Fan, C.; Deutach, S.; Gagić, D.; Seedorf, H.; et al. Methane yield phenotypes linked to differential gene expression in the sheep rumen microbiome. Genome. Res. 2014, 24, 1517–1525. [Google Scholar] [CrossRef]
- Demeyer, D.I.; Van Nevel, C.J. Methanogenesis, an integrated part of carbohydrate fermentation and its control. Dig. Metab. Rumin. 1975, 366–382. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/19761433771 (accessed on 1 August 2024).
- Denman, S.E.; Fernandez, G.M.; Shinkai, T.; Mitsumori, M.; McSweeney, C.S. Metagenomic analysis of the rumen microbial community following inhibition of methane formation by a halogenated methane analog. Front. Microbiol. 2015, 6, 1087. [Google Scholar] [CrossRef]
- Aguilar-Marin, S.B.; Betancur-Murillo, C.L.; Isaza, G.A.; Mesa, H.; Jovel, J. Lower Methane Emissions Were Associated with Higher Abundance of Ruminal Prevotella in a Cohort of Colombian Buffalos. BMC Microbiol. 2020, 20, 364. [Google Scholar] [CrossRef] [PubMed]
- Betancur-Murillo, C.L.; Aguilar-Marín, S.B.; Jovel, J. Prevotella: A Key Player in Ruminal Metabolism. Microorganisms 2023, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.W.; Zhang, Z.; Zhang, G.H.; He, B.; Qin, Y.L.; Yang, B.; Yu, Z.T.; Wang, J.K. Maternal Rumen Bacteriota Shapes the Offspring Rumen Bacteriota, Affecting the Development of Young Ruminants. Microbiol. Spectr. 2023, 11, e03590-22. [Google Scholar] [CrossRef] [PubMed]
- Janssen, P.H.; Kirs, M. Structure of the archaeal community of the rumen. Appl. Environ. Microbiol. 2008, 74, 3619–3625. [Google Scholar] [CrossRef]
- Kang, S.H.; Evans, P.; Morrison, M.; McSweeney, C. Identification of metabolically active proteobacterial and archaeal communities in the rumen by DNA- and RNA-derived 16S rRNA gene. J. Appl. Microbiol. 2013, 115, 644–653. [Google Scholar] [CrossRef]
- Newbold, C.J.; De La Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The role of ciliate protozoa in the rumen. Front. Microbiol. 2015, 6, 1313. [Google Scholar] [CrossRef]
- Emerson, E.L.; Weimer, P.J. Fermentation of model hemicelluloses by Prevotella strains and Butyrivibrio fibrisolvens in pure culture and in ruminal enrichment cultures. Appl. Microbiol. Biotechnol. 2017, 101, 4269–4278. [Google Scholar] [CrossRef]
- Bi, Y.L.; Zeng, S.Q.; Zhang, R.; Diao, Q.Y.; Tu, Y. Effects of dietary energy levels on rumen bacterial community composition in Holstein heifers under the same forage to concentrate ratio condition. BMC Microbiol. 2018, 18, 69. [Google Scholar] [CrossRef]
- Li, N.; Xue, Y.G.; Chen, S.S.; Takahashi, J.; Dai, L.L.; Dai, X.H. Methanogenic population dynamics regulated by bacterial community responses to protein-rich organic wastes in a high solid anaerobic digester. Chem. Eng. J. 2017, 317, 444–453. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, J.Z.; Zhou, Y.L.; Almeida, A.; Bateman, A.; Danchin, A.; He, L.S. Phylogenomics of expanding uncultured environmental Tenericutes provides insights into their pathogenicity and evolutionary relationship with Bacilli. BMC Genom. 2020, 21, 408. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Content % |
|---|---|
| Ingredient | |
| Corn | 26.26 |
| Wheat bran | 2.10 |
| Soybean meal | 2.60 |
| Corn germ meal | 3.20 |
| CaHCO3 | 0.08 |
| CaCO3 | 0.26 |
| Sodium bicarbonate | 0.70 |
| Salt | 0.80 |
| Premix 1 | 4.00 |
| Leymus chinensis | 35 |
| Alfalfa hay | 25 |
| Total | 100 |
| Nutritional Level 2 | |
| DM | 91.32 |
| EE | 2.05 |
| CHO | 74.26 |
| NFC | 39.18 |
| CP | 12.13 |
| NDF | 34.85 |
| ADF | 22.04 |
| Ca | 0.43 |
| P | 0.35 |
| DE, MJ/kg | 9.35 |
| ME, MJ/kg | 7.67 |
| CP/ME | 1.58 |
| Items | DM | CP | EE | NDF | ADF | CA 1 |
|---|---|---|---|---|---|---|
| Corn | 86.45 | 9.11 | 3.61 | 9.01 | 3.01 | 1.23 |
| Wheat bran | 87.12 | 15.70 | 4.64 | 41.34 | 11.93 | 4.85 |
| Soybean meal | 89.25 | 44.20 | 1.9 | 13.06 | 9.06 | 6.11 |
| Corn germ meal | 90.11 | 20.83 | 2.32 | 28.3 | 12.533 | 5.93 |
| Leymus chinensis | 91.01 | 8.02 | 2.01 | 57.62 | 39.25 | 8.05 |
| Alfalfa hay | 88.10 | 18.78 | 1.34 | 41.75 | 33.01 | 8.24 |
| Item | Treatment 1 | SEM 2 | p-Value 3 | |||||
|---|---|---|---|---|---|---|---|---|
| C | LD | MD | HD | Trt | L | Q | ||
| Total gas production, mL | 144 a | 120.3 b | 116 c | 114.3 c | 1.11 | <0.01 | <0.01 | <0.01 |
| Methane production, mL | 20.76 a | 3.84 b | 0.51 c | 0.46 c | 0.10 | <0.01 | <0.01 | <0.01 |
| Hydrogen production, mL | 0.02 c | 5.88 b | 9.25 a | 9.68 a | 0.41 | <0.01 | <0.01 | <0.01 |
| Degradability of dry matter, % | 55.63 | 55.15 | 54.16 | 54.82 | 0.84 | 0.67 | 0.35 | 0.52 |
| Degradability of neutral detergent fiber, % | 48.85 | 48.68 | 49.69 | 48.66 | 1.41 | 0.95 | 0.94 | 0.95 |
| Items | Treatment | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| C | LD | MD | HD | Trt | L | Q | ||
| pH | 6.63 | 6.61 | 6.61 | 6.62 | 0.01 | 0.08 | 0.34 | 0.03 |
| NH3-N, mg/dL | 22.70 | 23.01 | 22.66 | 22.96 | 0.40 | 0.90 | 0.80 | 0.97 |
| TVFAs 1, mmol/L | 110.37 a | 106.45 a | 99.16 b | 101.53 b | 1.40 | <0.01 | <0.01 | <0.01 |
| Acetate, mol/100 mol | 50.35 a | 45.05 b | 42.37 b | 43.76 b | 0.89 | <0.01 | <0.01 | <0.01 |
| Propionate, mol/100 mol | 30.66 b | 32.46 a | 33.78 a | 33.64 a | 0.33 | <0.01 | <0.01 | <0.01 |
| Butyrate, mol/100 mol | 15.99 b | 19.53 a | 20.89 a | 19.70 a | 0.87 | 0.02 | 0.03 | <0.01 |
| Valerate, mol/100 mol | 3 | 2.96 | 2.96 | 2.91 | 0.12 | 0.96 | 0.57 | 0.86 |
| Acetate/propionate | 1.64 a | 1.39 b | 1.25 c | 1.30 c | 0.04 | <0.01 | <0.01 | <0.01 |
| Total bacteria counts 2, ×109 | 2.31 | 2.19 | 2.80 | 2.38 | 0.24 | 0.35 | 0.48 | 0.66 |
| Total methanogen counts 3, ×106 | 7.24 | 6.98 | 6.59 | 6.30 | 0.39 | 0.36 | 0.07 | 0.19 |
| Total protozoa counts 4, ×104 | 5.64 | 5.58 | 4.91 | 4.83 | 0.40 | 0.36 | 0.09 | 0.24 |
| Items | Treatment | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| C | LD | MD | HD | Trt | L | Q | ||
| Archaea at the Phylum Level | ||||||||
| Euryarchaeota | 99.45 b | 99.67 ab | 99.82 a | 99.76 a | 0.09 | 0.03 | 0.01 | 0.01 |
| Crenarchaeota | 0.02 | 0.01 | 0.004 | 0.002 | 0.01 | 0.22 | 0.04 | 0.10 |
| Others | 0.54 a | 0.32 ab | 0.18 b | 0.23 b | 0.08 | 0.04 | 0.01 | 0.01 |
| Bacteria at the Phylum Level | ||||||||
| Bacteroidetes | 53.48 | 50.50 | 54.50 | 51.15 | 1.13 | 0.07 | 0.61 | 0.87 |
| Firmicutes | 30.93 | 32.42 | 30.01 | 33.02 | 1.14 | 0.26 | 0.48 | 0.65 |
| Spirochaetes | 7.15 ab | 8.66 a | 7.83 a | 5.43 b | 0.66 | 0.02 | 0.10 | 0.01 |
| Verrucomicrobia | 4.23 a | 3.89 a | 3.11 a | 1.17 b | 0.54 | <0.01 | <0.01 | <0.01 |
| Proteobacteria | 0.75 b | 0.77 b | 1.23 b | 5.51 a | 0.47 | <0.01 | <0.01 | <0.01 |
| Lentisphaerae | 0.86 a | 0.92 a | 0.67 a | 0.19 b | 0.12 | <0.01 | <0.01 | <0.01 |
| Tenericutes | 0.70 | 0.74 | 0.61 | 0.55 | 0.05 | 0.10 | 0.03 | 0.07 |
| Fibrobacteres | 0.32 b | 0.46 b | 0.50 b | 1.07 a | 0.11 | <0.01 | <0.01 | <0.01 |
| TM7 | 0.36 a | 0.30 ab | 0.24 b | 0.16 c | 0.03 | <0.01 | <0.01 | <0.01 |
| Actinobacteria | 0.24 | 0.21 | 0.21 | 0.23 | 0.02 | 0.65 | 0.78 | 0.43 |
| Others | 0.99 b | 1.13 b | 1.10 b | 1.54 a | 0.11 | 0.01 | 0.04 | <0.01 |
| Protozoa at the Order Level | ||||||||
| Entodiniomorphida | 94.61 | 95.56 | 94.18 | 95.21 | 2.06 | 0.97 | 0.96 | 1 |
| Vestibuliferida | 0.51 | 2.42 | 1.92 | 2.08 | 1.20 | 0.69 | 0.42 | 0.56 |
| Others | 4.88 | 2.02 | 3.90 | 2.71 | 1.67 | 0.64 | 0.53 | 0.73 |
| Archaea at the Genus Level | ||||||||
| Methanobrevibacter | 94.27 b | 96.89 a | 97.37 a | 97.39 a | 0.60 | <0.01 | <0.01 | <0.01 |
| Methanosphaera | 0.94 | 0.87 | 0.83 | 0.59 | 0.11 | 0.15 | 0.09 | 0.07 |
| Methanosarcina | 1.92 a | 0.28 c | 0.20 c | 0.69 b | 0.41 | 0.03 | 0.03 | 0.01 |
| Methanimicrococcus | 0.46 | 0.31 | 0.36 | 0.22 | 0.13 | 0.64 | 0.25 | 0.52 |
| vadinCA11 | 0.05 | 0.03 | 0.10 | 0.03 | 0.02 | 0.17 | 0.95 | 0.56 |
| Methanoplanus | 0.01 | 0.04 | 0.02 | 0.02 | 0.01 | 0.31 | 0.85 | 0.53 |
| Methanobacterium | 0.07 | 0.03 | 0.01 | 0.003 | 0.03 | 0.18 | 0.10 | 0.12 |
| Methanosaeta | 0.03 a | 0.02 b | 0.02 b | 0.01 b | 0.01 | 0.03 | 0.15 | 0.06 |
| Candidatus_Nitrososphaera | 0.004 | 0.003 | 0.001 | 0.001 | 0.002 | 0.77 | 0.29 | 0.57 |
| Thermococcus | 0.001 | 0.006 | 0.0001 | 0.001 | 0.002 | 0.18 | 0.47 | 0.52 |
| Others | 2.24 a | 1.51 b | 1.10 b | 1.05 b | 0.19 | <0.01 | <0.01 | <0.01 |
| Bacteria at the Genus Level | ||||||||
| Prevotella | 12.32 c | 13.74 bc | 15.30 b | 21.74 a | 0.55 | <0.01 | <0.01 | <0.01 |
| Sphaerochaeta | 6.66 ab | 8.02 a | 7.37 a | 5 b | 0.64 | 0.02 | 0.11 | <0.01 |
| BF311 | 3.46 b | 3.43 b | 3.81 b | 5.31 a | 0.21 | <0.01 | <0.01 | <0.01 |
| Clostridiaceae_Clostridium | 2.51 | 3.03 | 2.73 | 2.53 | 0.22 | 0.34 | 0.82 | 0.28 |
| Succiniclasticum | 1.01 ab | 0.34 b | 0.87 ab | 1.67 a | 0.25 | 0.02 | 0.07 | <0.01 |
| RFN20 | 1 ab | 1.11 a | 0.81 b | 0.44 c | 0.07 | <0.01 | <0.01 | <0.01 |
| CF231 | 0.83 | 0.58 | 0.78 | 0.99 | 0.12 | 0.15 | 0.13 | 0.01 |
| Streptococcus | 0.01 | 0.01 | 0.23 | 2.69 | 0.81 | 0.09 | 0.04 | 0.04 |
| YRC22 | 0.65 | 0.55 | 0.65 | 0.74 | 0.06 | 0.21 | 0.22 | 0.13 |
| Fibrobacter | 0.32 b | 0.46 b | 0.50 b | 1.07 a | 0.11 | <0.01 | <0.01 | <0.01 |
| Others | 71.25 a | 68.73 a | 66.97 a | 57.83 b | 1.51 | <0.01 | <0.01 | <0.01 |
| Protozoa at the Genus Level | ||||||||
| Entodinium | 49.21 a | 48.33 a | 48.03 a | 37.08 b | 2.97 | 0.03 | 0.02 | 0.02 |
| Diplodinium | 30.08 a | 29.98 a | 23.11 b | 13.96 c | 1.84 | <0.01 | <0.01 | <0.01 |
| Ophryoscolex | 2.13 c | 4.96 bc | 14.14 b | 32.41 a | 3.13 | <0.01 | <0.01 | <0.01 |
| Ostracodinium | 3.01 | 2.86 | 1.99 | 4.26 | 1.45 | 0.74 | 0.65 | 0.65 |
| Isotricha | 1.51 | 1.42 | 1.92 | 2.08 | 1.02 | 0.96 | 0.61 | 0.88 |
| Eudiplodinium | 0 | 0.001 | 0 | 1.51 | 0.75 | 0.42 | 0.19 | 0.26 |
| Epidinium | 0 | 0 | 0 | 0.0005 | 0.0002 | 0.42 | 0.19 | 0.26 |
| Cycloposthium | 0 | 0 | 0 | 0.0005 | 0.0002 | 0.42 | 0.67 | 0.57 |
| Others | 14.06 | 12.45 | 10.81 | 8.70 | 3.31 | 0.70 | 0.22 | 0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xuan, T.; Zheng, T.; Li, T.; Wu, B.; Li, T.; Bao, W.; Qin, W. The Effects of Different Doses of 3-NOP on Ruminal Fermentation Parameters, Methane Production, and the Microbiota of Lambs In Vitro. Fermentation 2024, 10, 440. https://doi.org/10.3390/fermentation10090440
Xuan T, Zheng T, Li T, Wu B, Li T, Bao W, Qin W. The Effects of Different Doses of 3-NOP on Ruminal Fermentation Parameters, Methane Production, and the Microbiota of Lambs In Vitro. Fermentation. 2024; 10(9):440. https://doi.org/10.3390/fermentation10090440
Chicago/Turabian StyleXuan, Tianbao, Tingfang Zheng, Tiyu Li, Baiyila Wu, Tailin Li, Wenjun Bao, and Weize Qin. 2024. "The Effects of Different Doses of 3-NOP on Ruminal Fermentation Parameters, Methane Production, and the Microbiota of Lambs In Vitro" Fermentation 10, no. 9: 440. https://doi.org/10.3390/fermentation10090440
APA StyleXuan, T., Zheng, T., Li, T., Wu, B., Li, T., Bao, W., & Qin, W. (2024). The Effects of Different Doses of 3-NOP on Ruminal Fermentation Parameters, Methane Production, and the Microbiota of Lambs In Vitro. Fermentation, 10(9), 440. https://doi.org/10.3390/fermentation10090440