
Pathogenic and Harmful Bacteria in Dairy Technology: Genomic Characterization and Its Correlation with Physicochemical Parameters of Sheep’s Cheese Sold in Southern Brazil


 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Characterization and Collection
2.2. Characterization of Sheep Cheese
2.2.1. Physicochemical Analyses
2.2.2. Fatty Acid Profile Analysis by GC-FID
2.2.3. Volatile Compounds Profile Analysis by GC-MS
2.3. 16S Amplicon Sequencing and Analysis
2.4. Pangenomic Analysis
2.4.1. Comparative Analysis of Genomes
Phylogenetic Analysis
Complete Reference Genomes for In Silico Analysis
Genomic Similarity Analysis between 16S rRNA Genes and Reference Genes
2.4.2. Research of Virulence and Antimicrobial Resistance Genes
2.5. Statistical Analysis
3. Results and Discussion
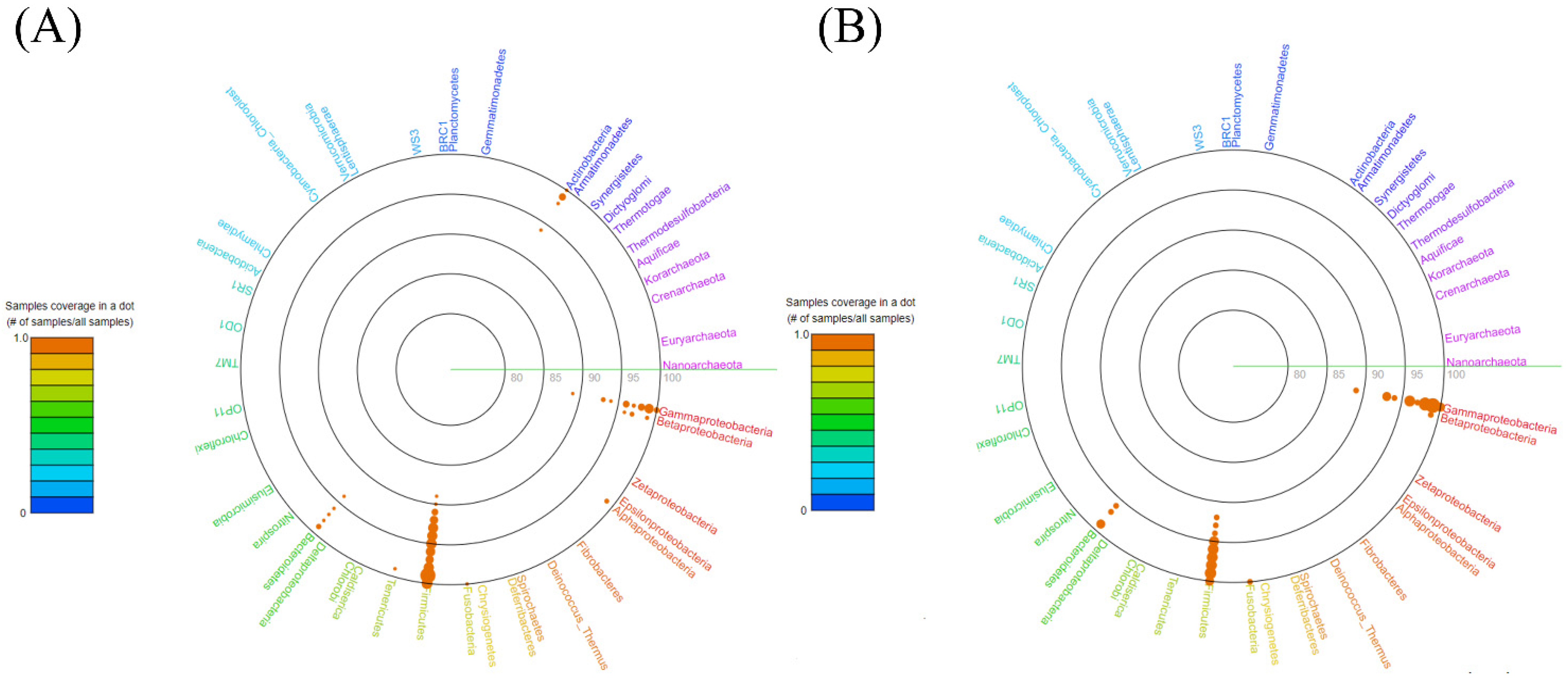
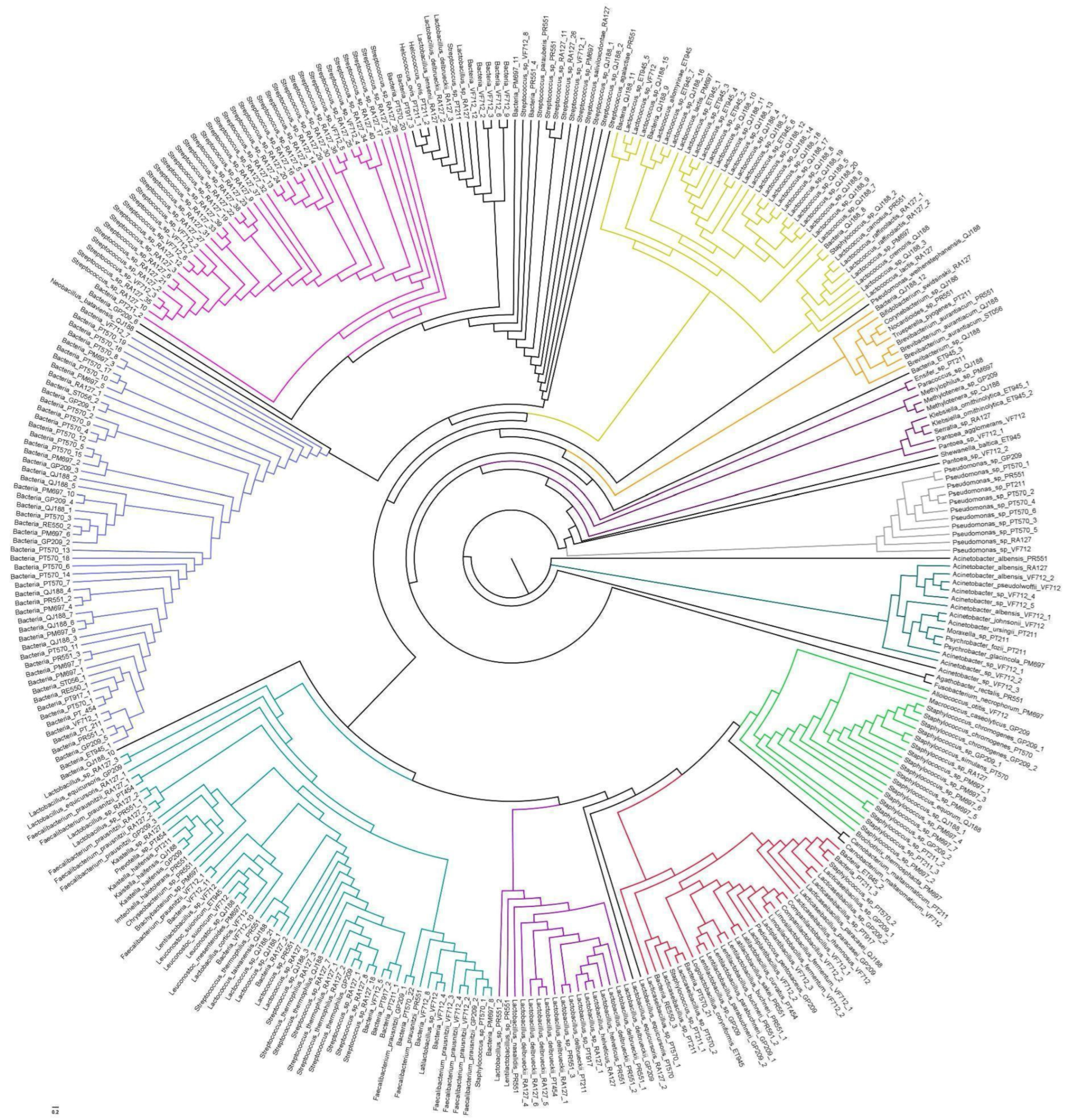
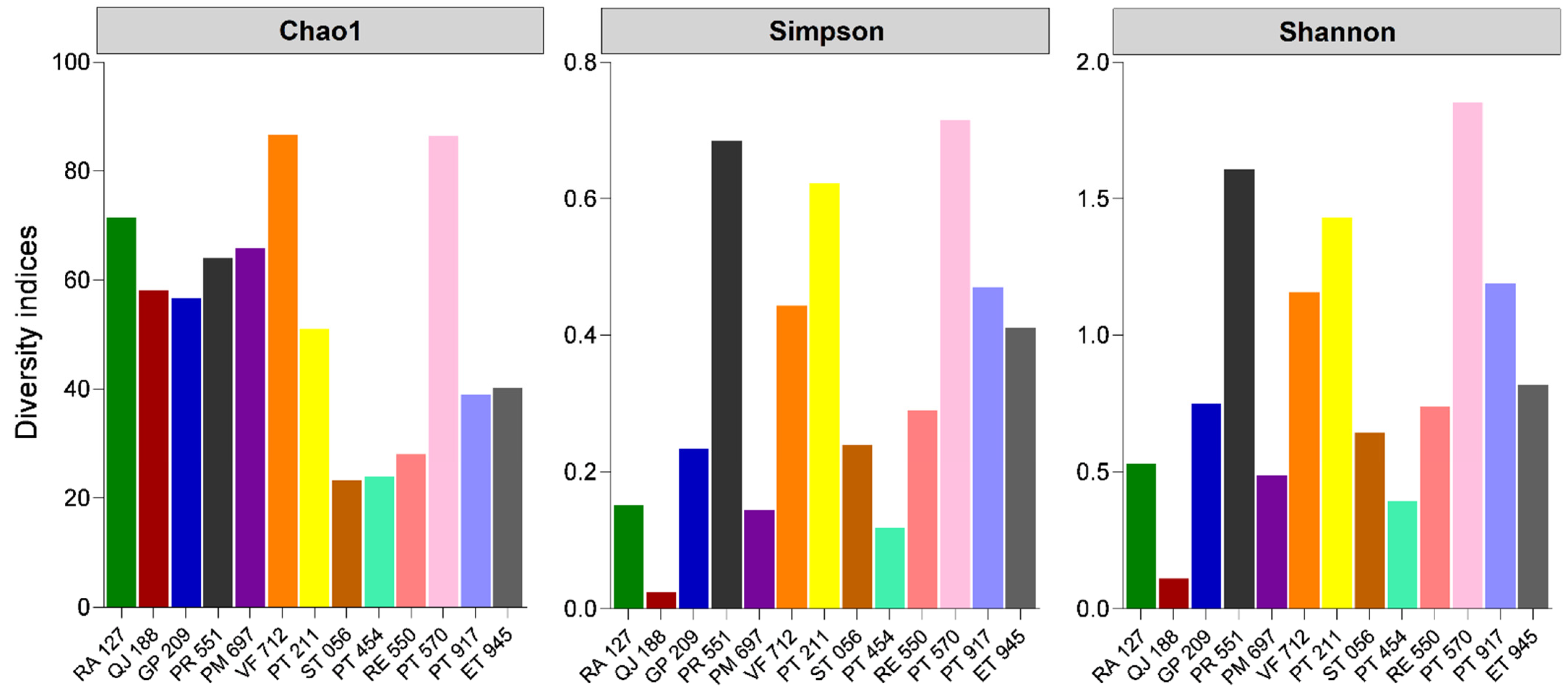
3.1. Bacterial Population Profile in Sheep’s Cheese
3.2. Profile of Bacteria with Pathogenic Potential and Harmful to Milk Technology
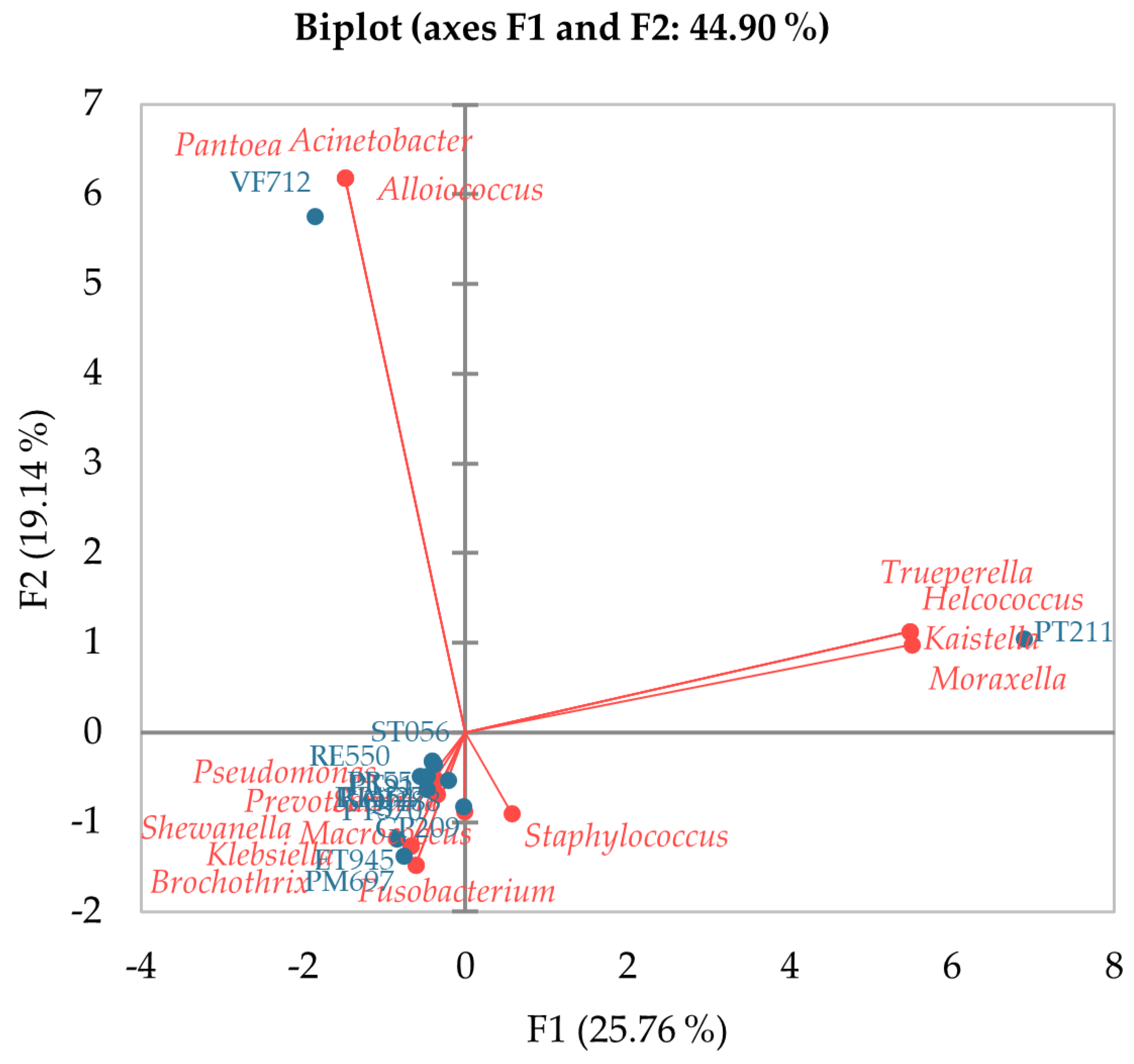
3.3. Correlation between Groups of Bacteria with Pathogenic and Harmful Potential for Milk Technology and Physicochemical Properties
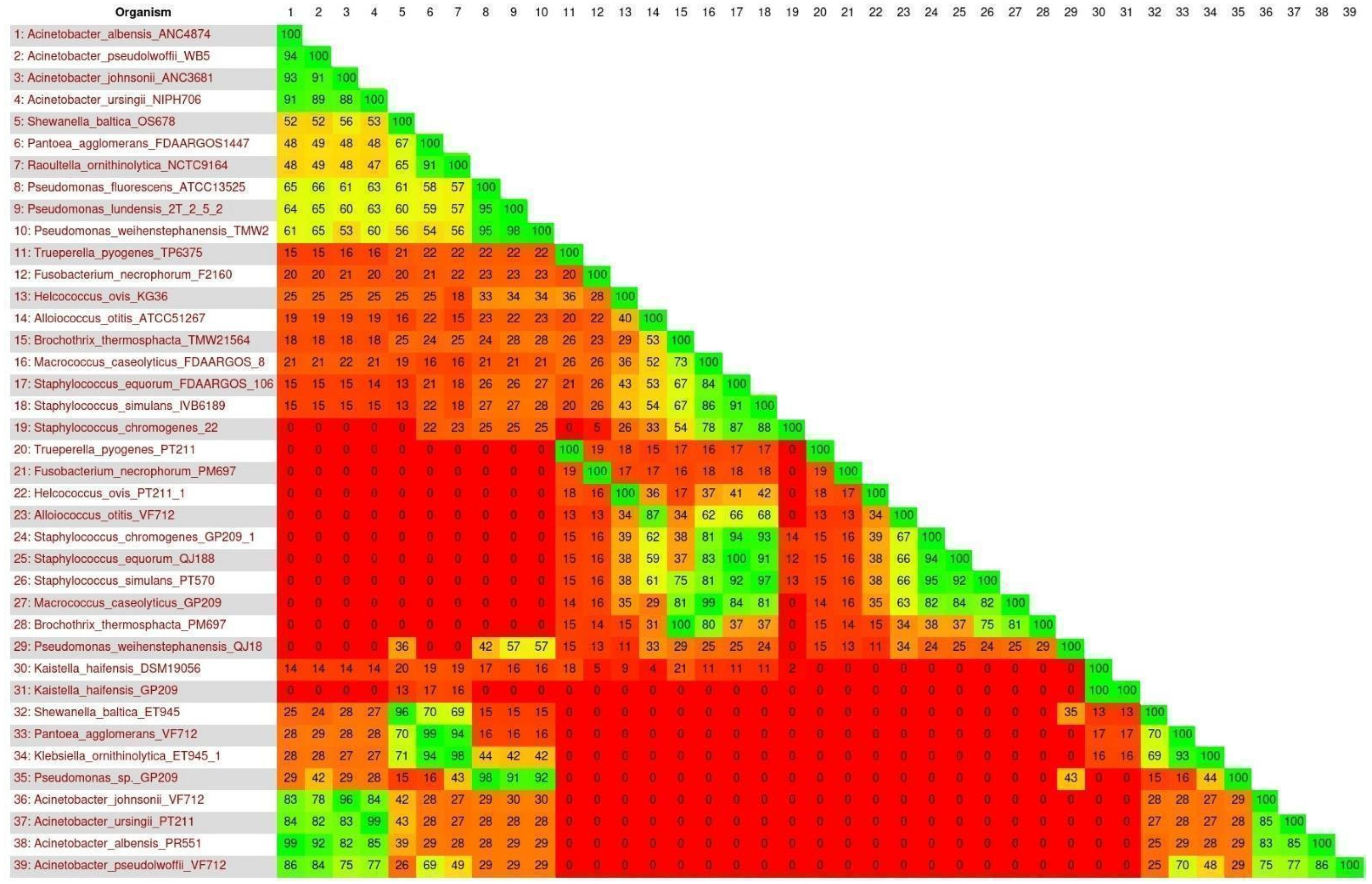
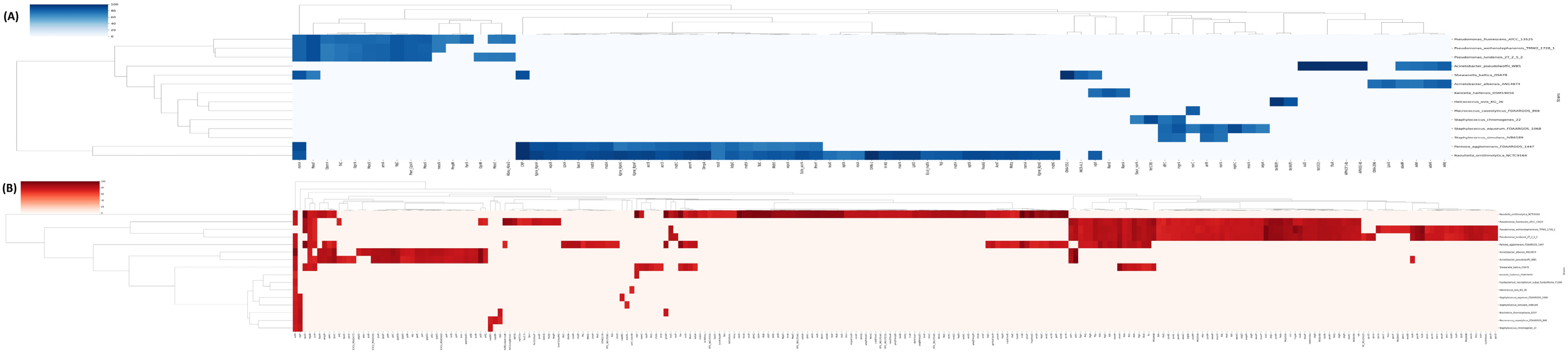
3.4. Search for Resistance and Virulence Genes in Reference Genomes
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 23 July 2024).
- Bianchi, A.E.; Monteiro, A.L.G.; Morais, O.R.d.; Beltrão, R.B.F.; Debortoli, E.C. Caracterização dos sistemas produtivos de ovinos de leite no Brasil. MilkPoint. 2016, Seção Radar Técnico, Ovinos e Caprinos, 1-5. Available online: http://www.infoteca.cnptia.embrapa.br/infoteca/handle/doc/1073171 (accessed on 23 July 2024).
- Barłowska, J.; Szwajkowska, M.; Litwińczuk, Z.; Król, J. Nutritional Value and Technological Suitability of Milk from Various Animal Species Used for Dairy Production. Compr. Rev. Food Sci. Food Saf. 2011, 10, 291–302. [Google Scholar] [CrossRef]
- Bittante, G.; Amalfitano, N.; Bergamaschi, M.; Patel, N.; Haddi, M.-L.; Benabid, H.; Pazzola, M.; Vacca, G.M.; Tagliapietra, F.; Schiavon, S. Composition and Aptitude for Cheese-Making of Milk from Cows, Buffaloes, Goats, Sheep, Dromedary Camels, and Donkeys. J. Dairy Sci. 2022, 105, 2132–2152. [Google Scholar] [CrossRef] [PubMed]
- Dalmina, E.M.; Malta, D.S.; Silva, F.C.; Tribst, A.A.L.; Rigo, E.; Cavalheiro, D. Sodium Reduction in “Requeijão Cremoso” Processed Cheese Made from Fresh and Refrigerated Sheep Milk. J. Food Process. Preserv. 2022, 46, e16418. [Google Scholar] [CrossRef]
- Hassan, F.A.; Abbas, H.M.; Abd El-Gawad, M.A.; Enab, A.K. Goats Dairy Products as a Potentially Functional Food. Life Sci. J. 2014, 11, 648–657. [Google Scholar]
- Ramos, T.M.; Gajo, A.A.; Pinto, S.M.; Abreu, L.R.; Pinheiro, A.C. Perfil de textura de Labneh (iogurte grego). Rev. Inst. Laticínios Cândido Tostes 2009, 64, 8–12. [Google Scholar]
- Robinson, R.K.; Tamime, A.Y. Feta & Related Cheeses; CRC Press: Boca Raton, FL, USA, 1996; ISBN 978-0-7476-0077-0. [Google Scholar]
- Furtado, M.M. Quesos Típicos de Latinoamérica; Danisco: São Paulo, Brazil, 2005. [Google Scholar]
- Addis, M.; Piredda, G.; Pes, M.; Di Salvo, R.; Scintu, M.F.; Pirisi, A. Effect of the Use of Three Different Lamb Paste Rennets on Lipolysis of the PDO Pecorino Romano Cheese. Int. Dairy J. 2005, 15, 563–569. [Google Scholar] [CrossRef]
- Nespolo, C.R.; Bianchi, A.E.; Queirós, A.A.E.; Farinatti, L.H.E. A Produção de Leite Ovino e Seus Derivados No Oeste Catarinense: Uma Alternativa Para o Produtor e Para o Consumidor. In Anais do Simpósio de Segurança Alimentar; FAURGS: Gramado, Brazil, 2012; Volume 4. [Google Scholar]
- Pedroso, M.A.P.; Nunes, L.S.; Uliana, G.C.; Richards, N.S.P.d.S. Estratégias e Inovações No Mercado Lácteo: Uma Análise Dos Produtos de Leite de Ovelha No Sul Do Brasil. Fazer Ciência 2023, 157, 55–60. [Google Scholar]
- Tilocca, B.; Costanzo, N.; Morittu, V.M.; Spina, A.A.; Soggiu, A.; Britti, D.; Roncada, P.; Piras, C. Milk Microbiota: Characterization Methods and Role in Cheese Production. J. Proteom. 2020, 210, 103534. [Google Scholar] [CrossRef] [PubMed]
- Paszczyk, B.; Łuczyńska, J. The Comparison of Fatty Acid Composition and Lipid Quality Indices in Hard Cow, Sheep, and Goat Cheeses. Foods 2020, 9, 1667. [Google Scholar] [CrossRef]
- Fusieger, A.; Martins, M.C.F.; de Freitas, R.; Nero, L.A.; de Carvalho, A.F. Technological Properties of Lactococcus Lactis Subsp. Lactis Bv. Diacetylactis Obtained from Dairy and Non-Dairy Niches. Braz. J. Microbiol. 2020, 51, 313–321. [Google Scholar] [CrossRef]
- Bovolenta, S.; Romanzin, A.; Corazzin, M.; Spanghero, M.; Aprea, E.; Gasperi, F.; Piasentier, E. Volatile Compounds and Sensory Properties of Montasio Cheese Made from the Milk of Simmental Cows Grazing on Alpine Pastures. J. Dairy Sci. 2014, 97, 7373–7385. [Google Scholar] [CrossRef] [PubMed]
- Pillonel, L.; Ampuero, S.; Tabacchi, R.; Bosset, J. Analytical Methods for the Determination of the Geographic Origin of Emmental Cheese: Volatile Compounds by GC/MS-FID and Electronic Nose. Eur. Food Res. Technol. 2003, 216, 179–183. [Google Scholar] [CrossRef]
- Possas, A.; Bonilla-Luque, O.M.; Valero, A. From Cheese-Making to Consumption: Exploring the Microbial Safety of Cheeses through Predictive Microbiology Models. Foods 2021, 10, 355. [Google Scholar] [CrossRef] [PubMed]
- Remor, A.; de Vasconcelos, T.C.B.; Belo, V.S.; Zanella, R.; dos Santos, E.D.; Dickel, E.L. Metagenomic investigation in colonial cheese produced with pasteurized milk and sold in the state of Rio Grande do Sul, Brazil. Veterinária Zootec. 2021, 28, 1–9. [Google Scholar] [CrossRef]
- Amato, E.; Filipello, V.; Gori, M.; Lomonaco, S.; Losio, M.N.; Parisi, A.; Huedo, P.; Knabel, S.J.; Pontello, M. Identification of a Major Listeria Monocytogenes Outbreak Clone Linked to Soft Cheese in Northern Italy—2009–2011. BMC Infect. Dis. 2017, 17, 342. [Google Scholar] [CrossRef] [PubMed]
- Castro, R.D.; Pedroso, S.H.S.P.; Sandes, S.H.C.; Silva, G.O.; Luiz, K.C.M.; Dias, R.S.; Filho, R.A.T.; Figueiredo, H.C.P.; Santos, S.G.; Nunes, A.C.; et al. Virulence Factors and Antimicrobial Resistance of Staphylococcus aureus Isolated from the Production Process of Minas Artisanal Cheese from the Region of Campo Das Vertentes, Brazil. J. Dairy Sci. 2020, 103, 2098–2110. [Google Scholar] [CrossRef] [PubMed]
- Alotaibi, K.; Khan, A.A. Prevalence and Molecular Characterization of Shiga Toxin-Producing Escherichia coli from Food and Clinical Samples. Pathogens 2023, 12, 1302. [Google Scholar] [CrossRef] [PubMed]
- Withenshaw, S.M.; Smith, R.P.; Davies, R.; Smith, A.E.O.; Gray, E.; Rodgers, J. A Systematized Review and Qualitative Synthesis of Potential Risk Factors Associated with the Occurrence of Non-O157 Shiga Toxin-Producing Escherichia coli (STEC) in the Primary Production of Cattle. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2363–2390. [Google Scholar] [CrossRef]
- Pinedo, L.C.; Mughini-Gras, L.; Franz, E.; Hald, T.; Pires, S.M. Sources and Trends of Human Salmonellosis in Europe, 2015–2019: An Analysis of Outbreak Data. Int. J. Food Microbiol. 2022, 379, 109850. [Google Scholar] [CrossRef]
- Primavilla, S.; Roila, R.; Rocchegiani, E.; Blasi, G.; Petruzzelli, A.; Gabucci, C.; Ottaviani, D.; Di Lullo, S.; Branciari, R.; Ranucci, D.; et al. Assessment of the Microbiological Safety and Hygiene of Raw and Thermally Treated Milk Cheeses Marketed in Central Italy between 2013 and 2020. Life 2023, 13, 2324. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. High-Throughput Sequencing for Detection of Subpopulations of Bacteria Not Previously Associated with Artisanal Cheeses. Appl. Environ. Microbiol. 2012, 78, 5717–5723. [Google Scholar] [CrossRef] [PubMed]
- Regecová, I.; Výrostková, J.; Zigo, F.; Gregová, G.; Kováčová, M. Detection of Antimicrobial Resistance of Bacteria Staphylococcus chromogenes Isolated from Sheep’s Milk and Cheese. Antibiotics 2021, 10, 570. [Google Scholar] [CrossRef] [PubMed]
- Danquah, C.A.; Minkah, P.A.B.; Duah-Junior, I.O.; Amankwah, K.B.; Somuah, S.O. Antimicrobial Compounds from Microorganisms. Antibiotics 2022, 11, 285. [Google Scholar] [CrossRef] [PubMed]
- Endres, C.M.; Moreira, E.; de Freitas, A.B.; Castel, A.P.D.; Graciano, F.; Mann, M.B.; Frazzon, A.P.G.; Mayer, F.Q.; Frazzon, J. Evaluation of Enterotoxins and Antimicrobial Resistance in Microorganisms Isolated from Raw Sheep Milk and Cheese: Ensuring the Microbiological Safety of These Products in Southern Brazil. Microorganisms 2023, 11, 1618. [Google Scholar] [CrossRef] [PubMed]
- Dacres, H.; Weihs, F.; Wang, J.; Anderson, A.; Trowell, S.C. Bioluminescence Resonance Energy Transfer Biosensor for Measuring Activity of a Protease Secreted by Pseudomonas fluorescens Growing in Milk. Anal. Chim. Acta 2023, 1270, 341401. [Google Scholar] [CrossRef]
- Otero-Asman, J.R.; Sánchez-Jiménez, A.; Bastiaansen, K.C.; Wettstadt, S.; Civantos, C.; García-Puente, A.; Bitter, W.; Llamas, M.A. The Prc and CtpA Proteases Modulate Cell-Surface Signaling Activity and Virulence in Pseudomonas aeruginosa. Science 2023, 26, 107216. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Ye, A.; Jiang, N. The Role of Bacteria in Gallstone Formation. Folia Microbiol. 2024, 69, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Parija, S.C. Pseudomonas, Burkholderia and Acinetobacter. In Textbook of Microbiology and Immunology; Parija, S.C., Ed.; Springer Nature: Singapore, 2023; pp. 553–561. ISBN 978-981-19331-5-8. [Google Scholar]
- Leonard, S.R.; Mammel, M.K.; Lacher, D.W.; Elkins, C.A. Application of Metagenomic Sequencing to Food Safety: Detection of Shiga Toxin-Producing Escherichia coli on Fresh Bagged Spinach. Appl. Environ. Microbiol. 2015, 81, 8183–8191. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Waldron, L.; Ballarini, A.; Narasimhan, V.; Jousson, O.; Huttenhower, C. Metagenomic Microbial Community Profiling Using Unique Clade-Specific Marker Genes. Nat. Methods 2012, 9, 811–814. [Google Scholar] [CrossRef]
- Erhardt, M.M.; Oliveira, W.d.C.; Fröder, H.; Marques, P.H.; Oliveira, M.B.P.P.; Richards, N.S.P.d.S. Lactic Bacteria in Artisanal Cheese: Characterization through Metagenomics. Fermentation 2023, 9, 41. [Google Scholar] [CrossRef]
- Keenum, I.; Wind, L.; Ray, P.; Guron, G.; Chen, C.; Knowlton, K.; Ponder, M.; Pruden, A. Metagenomic Tracking of Antibiotic Resistance Genes through a Pre-Harvest Vegetable Production System: An Integrated Lab-, Microcosm- and Greenhouse-Scale Analysis. Environ. Microbiol. 2022, 24, 3705–3721. [Google Scholar] [CrossRef]
- Moon, S.H.; Udaondo, Z.; Abram, K.Z.; Li, X.; Yang, X.; DiCaprio, E.L.; Jun, S.-R.; Huang, E. Isolation of AmpC- and Extended Spectrum β-Lactamase-Producing Enterobacterales from Fresh Vegetables in the United States. Food Control 2022, 132, 108559. [Google Scholar] [CrossRef]
- Onalenna, O.; Rahube, T.O. Assessing Bacterial Diversity and Antibiotic Resistance Dynamics in Wastewater Effluent-Irrigated Soil and Vegetables in a Microcosm Setting. Heliyon 2022, 8, e09089. [Google Scholar] [CrossRef]
- Valentino, V.; Sequino, G.; Cobo-Díaz, J.F.; Álvarez-Ordóñez, A.; De Filippis, F.; Ercolini, D. Evidence of Virulence and Antibiotic Resistance Genes from the Microbiome Mapping in Minimally Processed Vegetables Producing Facilities. Food Res. Int. 2022, 162, 112202. [Google Scholar] [CrossRef]
- Yasir, M.; Al-Zahrani, I.A.; Bibi, F.; Abd El Ghany, M.; Azhar, E.I. New Insights of Bacterial Communities in Fermented Vegetables from Shotgun Metagenomics and Identification of Antibiotic Resistance Genes and Probiotic Bacteria. Food Res. Int. 2022, 157, 111190. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Li, L.; Xu, Y.; Li, H.; Yu, Y.; Xu, Z. Metagenomics Reveals the Microbial Community Responsible for Producing Biogenic Amines During Mustard [Brassica Juncea (L.)] Fermentation. Front. Microbiol. 2022, 13, 824644. [Google Scholar] [CrossRef] [PubMed]
- Yurt, M.N.Z.; Ersoy Omeroglu, E.; Tasbasi, B.B.; Acar, E.E.; Altunbas, O.; Ozalp, V.C.; Sudagidan, M. Bacterial and Fungal Microbiota of Mould-Ripened Cheese Produced in Konya. Int. J. Dairy Technol. 2023, 76, 627–637. [Google Scholar] [CrossRef]
- Papadakis, P.; Konteles, S.; Batrinou, A.; Ouzounis, S.; Tsironi, T.; Halvatsiotis, P.; Tsakali, E.; Van Impe, J.F.M.; Vougiouklaki, D.; Strati, I.F.; et al. Characterization of Bacterial Microbiota of P.D.O. Feta Cheese by 16S Metagenomic Analysis. Microorganisms 2021, 9, 2377. [Google Scholar] [CrossRef]
- Masenya, K.; Manganyi, M.C.; Dikobe, T.B. Exploring Cereal Metagenomics: Unravelling Microbial Communities for Improved Food Security. Microorganisms 2024, 12, 510. [Google Scholar] [CrossRef]
- Pérez-Cobas, A.E.; Gomez-Valero, L.; Buchrieser, C. Metagenomic Approaches in Microbial Ecology: An Update on Whole-Genome and Marker Gene Sequencing Analyses. Microb. Genom. 2020, 6, e000409. [Google Scholar] [CrossRef]
- Taş, N.; de Jong, A.E.; Li, Y.; Trubl, G.; Xue, Y.; Dove, N.C. Metagenomic Tools in Microbial Ecology Research. Curr. Opin. Biotechnol. 2021, 67, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; Di Cagno, R.; Calasso, M.; Neviani, E.; Fox, P.F.; De Angelis, M. Drivers That Establish and Assembly the Lactic Acid Bacteria Biota in Cheeses. Trends Food Sci. Technol. 2018, 78, 244–254. [Google Scholar] [CrossRef]
- Lemes, M.R.; Rodrigues, T.C.V.; Jaiswal, A.K.; Tiwari, S.; Sales-Campos, H.; Andrade-Silva, L.E.; Oliveira, C.J.F.; Azevedo, V.; Rodrigues, V.; Soares, S.C.; et al. In Silico Designing of a Recombinant Multi-Epitope Antigen for Leprosy Diagnosis. J. Genet. Eng. Biotechnol. 2022, 20, 128. [Google Scholar] [CrossRef] [PubMed]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S.; et al. Genome Analysis of Multiple Pathogenic Isolates of Streptococcus agalactiae: Implications for the Microbial “Pan-Genome”. Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef]
- Barh, D.; Tiwari, S.; Jain, N.; Ali, A.; Santos, A.R.; Misra, A.N.; Azevedo, V.; Kumar, A. In Silico Subtractive Genomics for Target Identification in Human Bacterial Pathogens. Drug Dev. Res. 2011, 72, 162–177. [Google Scholar] [CrossRef]
- Lux, T.M.; Lee, R.; Love, J. Genome-Wide Phylogenetic Analysis of the Pathogenic Potential of Vibrio furnissii. Front. Microbiol. 2014, 5, 435. [Google Scholar] [CrossRef]
- IAL Instituto Adolfo Lutz. Normas Analíticas Do Instituto Adolfo Lutz, 3rd ed.; IMESP: São Paulo, Brazil, 1985; Volume 1. [Google Scholar]
- IAL Instituto Adolfo Lutz. Métodos Físico-Químicos Para Análise de Alimentos; ANVISA: São Paulo, Brazil, 2008. [Google Scholar]
- Tontisirin, K.; MacLean, W.C.; Warwick, P. Food Energy: Methods of Analysis and Conversion Factors: Report of a Technical Workshop, Rome, 3–6 December 2002; FAO: Rome, Italy, 2003; ISBN 92-5-105014-7. [Google Scholar]
- Bligh, E.; Dyer, W. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Hartman, L.; Lago, R.C. Rapid Preparation of Fatty Acid Methyl Esters from Lipids. Lab. Pract. 1973, 22, 475–476. [Google Scholar]
- Visentainer, J.V. Aspectos analíticos da resposta do detector de ionização em chama para ésteres de ácidos graxos em biodiesel e alimentos. Quím. Nova 2012, 35, 274–279. [Google Scholar] [CrossRef]
- Zianni, R.; Mentana, A.; Tomaiuolo, M.; Campaniello, M.; Iammarino, M.; Centonze, D.; Palermo, C. Volatolomic Approach by HS-SPME/GC–MS and Chemometric Evaluations for the Discrimination of X-Ray Irradiated Mozzarella Cheese. Food Chem. 2023, 423, 136239. [Google Scholar] [CrossRef]
- NIST NIST V2.2 (National Institute of Standards and Technology, USA) Library Database. Available online: https://webbook.nist.gov/chemistry/name-ser/ (accessed on 16 June 2024).
- DePaula, J.; Cunha, S.C.; Cruz, A.; Sales, A.L.; Revi, I.; Fernandes, J.; Ferreira, I.M.P.L.V.O.; Miguel, M.A.L.; Farah, A. Volatile Fingerprinting and Sensory Profiles of Coffee Cascara Teas Produced in Latin American Countries. Foods 2022, 11, 3144. [Google Scholar] [CrossRef]
- Galvan-Lima, Â.; Cunha, S.C.; Martins, Z.E.; Soares, A.G.; Ferreira, I.M.P.L.V.O.; Farah, A. Headspace Volatolome of Peel Flours from Citrus Fruits Grown in Brazil. Food Res. Int. 2021, 150, 110801. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Wickham, H. Reshaping Data with the Reshape Package. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package 2024. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 23 July 2024).
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Mori, H.; Maruyama, T.; Yano, M.; Yamada, T.; Kurokawa, K. VITCOMIC2: Visualization Tool for the Phylogenetic Composition of Microbial Communities Based on 16S rRNA Gene Amplicons and Metagenomic Shotgun Sequencing. BMC Syst. Biol. 2018, 12, 30. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference Sequence (RefSeq) Database at NCBI: Current Status, Taxonomic Expansion, and Functional Annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Agren, J.; Sundström, A.; Håfström, T.; Segerman, B. Gegenees: Fragmented Alignment of Multiple Genomes for Determining Phylogenomic Distances and Genetic Signatures Unique for Specified Target Groups. PLoS ONE 2012, 7, e39107. [Google Scholar] [CrossRef]
- Rodrigues, D.L.N.; Ariute, J.C.; Rodrigues da Costa, F.M.; Benko-Iseppon, A.M.; Barh, D.; Azevedo, V.; Aburjaile, F. PanViTa: Pan Virulence and resistance Analysis. Front. Bioinform. 2023, 3, 1070406. [Google Scholar] [CrossRef] [PubMed]
- Bennato, F.; Di Domenico, M.; Ianni, A.; Di Gialleonardo, L.; Cammà, C.; Martino, G. Grape Pomace in Ewes Diet Affects Metagenomic Profile, Volatile Compounds and Biogenic Amines Contents of Ripened Cheese. Fermentation 2022, 8, 598. [Google Scholar] [CrossRef]
- Kamilari, E.; Anagnostopoulos, D.A.; Papademas, P.; Efthymiou, M.; Tretiak, S.; Tsaltas, D. Snapshot of Cyprus Raw Goat Milk Bacterial Diversity via 16S rDNA High-Throughput Sequencing; Impact of Cold Storage Conditions. Fermentation 2020, 6, 100. [Google Scholar] [CrossRef]
- Kamilari, E.; Anagnostopoulos, D.A.; Papademas, P.; Kamilaris, A.; Tsaltas, D. Characterizing Halloumi Cheese’s Bacterial Communities through Metagenomic Analysis. LWT 2020, 126, 109298. [Google Scholar] [CrossRef]
- Kamimura, B.A.; Cabral, L.; Noronha, M.F.; Baptista, R.C.; Nascimento, H.M.; Sant’Ana, A.S. Amplicon Sequencing Reveals the Bacterial Diversity in Milk, Dairy Premises and Serra Da Canastra Artisanal Cheeses Produced by Three Different Farms. Food Microbiol. 2020, 89, 103453. [Google Scholar] [CrossRef]
- Endres, C.M.; Castro, Í.M.S.; Trevisol, L.D.; Severo, J.M.; Mann, M.B.; Varela, A.P.M.; Frazzon, A.P.G.; Mayer, F.Q.; Frazzon, J. Molecular Characterization of the Bacterial Communities Present in Sheep’s Milk and Cheese Produced in South Brazilian Region via 16S rRNA Gene Metabarcoding Sequencing. LWT 2021, 147, 111579. [Google Scholar] [CrossRef]
- Porcellato, D.; Aspholm, M.; Skeie, S.B.; Monshaugen, M.; Brendehaug, J.; Mellegård, H. Microbial Diversity of Consumption Milk during Processing and Storage. Int. J. Food Microbiol. 2018, 266, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Qiu, S.; Wang, B.; Zeng, H. Metagenomic and Flavoromic Profiling Reveals the Correlation between the Microorganisms and Volatile Flavor Compounds in Monascus-Fermented Cheese. Food Res. Int. 2024, 188, 114483. [Google Scholar] [CrossRef]
- Araújo-Rodrigues, H.; Tavaria, F.K.; dos Santos, M.T.P.G.; Alvarenga, N.; Pintado, M.M. A Review on Microbiological and Technological Aspects of Serpa PDO Cheese: An Ovine Raw Milk Cheese. Int. Dairy J. 2020, 100, 104561. [Google Scholar] [CrossRef]
- Bencini, R.; Pulina, G. The Quality of Sheep Milk: A Review. Aust. J. Exp. Agric. 1997, 37, 485–504. [Google Scholar] [CrossRef]
- Felder, D.; Burns, D.; Chang, D. Defining Microbial Terroir: The Use of Native Fungi for the Study of Traditional Fermentative Processes. Int. J. Gastron. Food Sci. 2012, 1, 64–69. [Google Scholar] [CrossRef]
- Xu, Z.S.; Ju, T.; Yang, X.; Gänzle, M. A Meta-Analysis of Bacterial Communities in Food Processing Facilities: Driving Forces for Assembly of Core and Accessory Microbiomes across Different Food Commodities. Microorganisms 2023, 11, 1575. [Google Scholar] [CrossRef]
- Yuan, H.; Han, S.; Zhang, S.; Xue, Y.; Zhang, Y.; Lu, H.; Wang, S. Microbial Properties of Raw Milk throughout the Year and Their Relationships to Quality Parameters. Foods 2022, 11, 3077. [Google Scholar] [CrossRef]
- Rzewuska, M.; Kwiecień, E.; Chrobak-Chmiel, D.; Kizerwetter-Świda, M.; Stefańska, I.; Gieryńska, M. Pathogenicity and Virulence of Trueperella pyogenes: A Review. Int. J. Mol. Sci. 2019, 20, 2737. [Google Scholar] [CrossRef]
- Seng, P.; Boushab, B.M.; Romain, F.; Gouriet, F.; Bruder, N.; Martin, C.; Paganelli, F.; Bernit, E.; Treut, Y.P.L.; Thomas, P.; et al. Emerging Role of Raoultella ornithinolytica in Human Infections: A Series of Cases and Review of the Literature. Int. J. Infect. Dis. 2016, 45, 65–71. [Google Scholar] [CrossRef]
- Rodríguez-Medina, N.; Barrios-Camacho, H.; Duran-Bedolla, J.; Garza-Ramos, U. Klebsiella variicola: An Emerging Pathogen in Humans. Emerg. Microbes Infect. 2019, 8, 973–988. [Google Scholar] [CrossRef]
- Khoramrooz, S.S.; Mirsalehian, A.; Emaneini, M.; Jabalameli, F.; Aligholi, M.; Saedi, B.; Bazargani, A.; Taherikalani, M.; Borghaei, P.; Razmpa, E. Frequency of Alloicoccus otitidis, Streptococcus pneumoniae, Moraxella catarrhalis and Haemophilus influenzae in Children with Otitis Media with Effusion (OME) in Iranian Patients. Auris Nasus Larynx 2012, 39, 369–373. [Google Scholar] [CrossRef]
- Lima, B.B.; Leal, M.C. Parâmetros Indicadores de Qualidade de Queijos Artesanais Comercializados Em Castro-PR; Universidade Federal do Paraná: Curitiba, Brazil, 2017. [Google Scholar]
- Kliem, K.E.; Humphries, D.J.; Grandison, A.S.; Morgan, R.; Livingstone, K.M.; Givens, D.I.; Reynolds, C.K. Effect of a Whey Protein and Rapeseed Oil Gel Feed Supplement on Milk Fatty Acid Composition of Holstein Cows. J. Dairy Sci. 2019, 102, 288–300. [Google Scholar] [CrossRef]
- Margalho, L.P.; Kamimura, B.A.; Pimentel, T.C.; Balthazar, C.F.; Araujo, J.V.A.; Silva, R.; Conte-Junior, C.A.; Raices, R.S.L.; Cruz, A.G.; Sant’Ana, A.S. A Large Survey of the Fatty Acid Profile and Gross Composition of Brazilian Artisanal Cheeses. J. Food Compos. Anal. 2021, 101, 103955. [Google Scholar] [CrossRef]
- Mitani, T.; Asakuma, S.; Shinoda, Y.; Ueda, Y.; Aoki, Y.; Oshita, T. Effects of Ear Corn Silage Supplementation on Milk Production and Milk Fatty Acid Profiles in Grazing Dairy Farms. Anim. Sci. J. 2020, 91, e13454. [Google Scholar] [CrossRef]
- Nudda, A.; Cannas, A.; Correddu, F.; Atzori, A.S.; Lunesu, M.F.; Battacone, G.; Pulina, G. Sheep and Goats Respond Differently to Feeding Strategies Directed to Improve the Fatty Acid Profile of Milk Fat. Animals 2020, 10, 1290. [Google Scholar] [CrossRef]
- Silva, H.L.A.; Balthazar, C.F.; Esmerino, E.A.; Vieira, A.H.; Cappato, L.P.; Neto, R.P.C.; Verruck, S.; Cavalcanti, R.N.; Portela, J.B.; Andrade, M.M.; et al. Effect of Sodium Reduction and Flavor Enhancer Addition on Probiotic Prato Cheese Processing. Food Res. Int. 2017, 99, 247–255. [Google Scholar] [CrossRef]
- Silva, H.L.A.; Balthazar, C.F.; Esmerino, E.A.; Neto, R.P.C.; Rocha, R.S.; Moraes, J.; Cavalcanti, R.N.; Franco, R.M.; Tavares, M.I.B.; Santos, J.S.; et al. Partial Substitution of NaCl by KCl and Addition of Flavor Enhancers on Probiotic Prato Cheese: A Study Covering Manufacturing, Ripening and Storage Time. Food Chem. 2018, 248, 192–200. [Google Scholar] [CrossRef]
- Ahmad, M.T.; Shariff, M.; Md. Yusoff, F.; Goh, Y.M.; Banerjee, S. Applications of Microalga Chlorella vulgaris in Aquaculture. Rev. Aquac. 2020, 12, 328–346. [Google Scholar] [CrossRef]
- Mostafa, S.S.M. Microalgal Biotechnology: Prospects and Applications. In Plant Science; IntechOpen: London, UK, 2012; pp. 275–314. ISBN 978-953-51-0905-1. [Google Scholar]
- Nassar, M.; Jaffery, A.; Ibrahim, B.; Baraka, B.; Abosheaishaa, H. The Multidimensional Benefits of Eicosapentaenoic Acid: From Heart Health to Inflammatory Control. Egypt. J. Intern. Med. 2023, 35, 81. [Google Scholar] [CrossRef]
- Park, N.-H.; Choi, J.-S.; Hwang, S.-Y.; Kim, Y.-C.; Hong, Y.-K.; Cho, K.K.; Choi, I.S. Antimicrobial Activities of Stearidonic and Gamma-Linolenic Acids from the Green Seaweed Enteromorpha Linza against Several Oral Pathogenic Bacteria. Bot. Stud. 2013, 54, 39. [Google Scholar] [CrossRef]
- Ru, I.T.K.; Sung, Y.Y.; Jusoh, M.; Wahid, M.E.A.; Nagappan, T. Chlorella vulgaris: A Perspective on Its Potential for Combining High Biomass with High Value Bioproducts. Appl. Phycol. 2020, 1, 2–11. [Google Scholar] [CrossRef]
- Sukhikh, S.; Prosekov, A.; Ivanova, S.; Maslennikov, P.; Andreeva, A.; Budenkova, E.; Kashirskikh, E.; Tcibulnikova, A.; Zemliakova, E.; Samusev, I.; et al. Identification of Metabolites with Antibacterial Activities by Analyzing the FTIR Spectra of Microalgae. Life 2022, 12, 1395. [Google Scholar] [CrossRef]
- Perry, K.S.P. Queijos: Aspectos químicos, bioquímicos e microbiológicos. Química Nova 2004, 27, 293–300. [Google Scholar] [CrossRef]
- Curioni, P.M.G.; Bosset, J.O. Key Odorants in Various Cheese Types as Determined by Gas Chromatography-Olfactometry. Int. Dairy J. 2002, 12, 959–984. [Google Scholar] [CrossRef]
- Cardoso, M.G.B. Microbiota bacteriana e compostos voláteis de queijo Minas Artesanal da região da Serra da Canastra; Universidade Federal de Lavras: Lavras, Brazil, 2022. [Google Scholar]
- Furtado, M.M. O Estufamento Tardio Dos Queijos: Características e Prevenção, Uma Revisão. Rev. Inst. Laticínios Cândido Tostes 1985, 40, 3–39. [Google Scholar]
- Vinayamohan, P.G.; Viju, L.S.; Joseph, D.; Venkitanarayanan, K. Fermented Foods as a Potential Vehicle of Antimicrobial-Resistant Bacteria and Genes. Fermentation 2023, 9, 688. [Google Scholar] [CrossRef]
- Gaurav, A.; Bakht, P.; Saini, M.; Pandey, S.; Pathania, R. Role of Bacterial Efflux Pumps in Antibiotic Resistance, Virulence, and Strategies to Discover Novel Efflux Pump Inhibitors. Microbiology 2023, 169, 001333. [Google Scholar] [CrossRef] [PubMed]
- Naidu, V.; Bartczak, A.; Brzoska, A.J.; Lewis, P.; Eijkelkamp, B.A.; Paulsen, I.T.; Elbourne, L.D.H.; Hassan, K.A. Evolution of RND Efflux Pumps in the Development of a Successful Pathogen. Drug Resist. Updates 2023, 66, 100911. [Google Scholar] [CrossRef] [PubMed]
- Saini, N.; Aamir, M.; Singh, V.K.; Deepak, B.; Mona, S. Unveiling the Microbial Diversity and Functional Dynamics of Shiv Kund, Sohna Hot Spring, India through a Shotgun Metagenomics Approach. Arch. Microbiol. 2023, 205, 323. [Google Scholar] [CrossRef] [PubMed]
- Darby, E.M.; Trampari, E.; Siasat, P.; Gaya, M.S.; Alav, I.; Webber, M.A.; Blair, J.M.A. Molecular Mechanisms of Antibiotic Resistance Revisited. Nat. Rev. Microbiol. 2023, 21, 280–295. [Google Scholar] [CrossRef]
- Gervasoni, S.; Mehla, J.; Bergen, C.R.; Leus, I.V.; Margiotta, E.; Malloci, G.; Bosin, A.; Vargiu, A.V.; Lomovskaya, O.; Rybenkov, V.V.; et al. Molecular Determinants of Avoidance and Inhibition of Pseudomonas aeruginosa MexB Efflux Pump. mBio 2023, 14, e01403-23. [Google Scholar] [CrossRef]
- Zhou, J.-X.; Feng, D.-Y.; Li, X.; Zhu, J.-X.; Wu, W.-B.; Zhang, T. Advances in Research on Virulence Factors of Acinetobacter baumannii and Their Potential as Novel Therapeutic Targets. J. Appl. Microbiol. 2023, 134, lxac089. [Google Scholar] [CrossRef]
- Shadan, A.; Pathak, A.; Ma, Y.; Pathania, R.; Singh, R.P. Deciphering the Virulence Factors, Regulation, and Immune Response to Acinetobacter baumannii Infection. Front. Cell. Infect. Microbiol. 2023, 13, 1053968. [Google Scholar] [CrossRef]
- Fursova, N.K.; Fursov, M.V.; Astashkin, E.I.; Fursova, A.D.; Novikova, T.S.; Kislichkina, A.A.; Sizova, A.A.; Fedyukina, G.N.; Savin, I.A.; Ershova, O.N. Multidrug-Resistant and Extensively Drug-Resistant Acinetobacter baumannii Causing Nosocomial Meningitis in the Neurological Intensive Care Unit. Microorganisms 2023, 11, 2020. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhao, D.; Huang, N.; Liu, S.; Zheng, J.; Cao, J.; Zeng, W.; Zheng, X.; Wang, L.; Zhou, T.; et al. Clinical Impact of the Type VI Secretion System on Clinical Characteristics, Virulence and Prognosis of Acinetobacter baumannii during Bloodstream Infection. Microb. Pathog. 2023, 182, 106252. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.A.; Saha, P. Chemotactic Response of P-Nitrophenol Degrading Pseudomonas asiatica Strain PNPG3 through Phenotypic and Genome Sequence-Based in Silico Studies. 3 Biotech 2023, 13, 408. [Google Scholar] [CrossRef]
- He, L.; Zhao, L.; Li, Q.; Huang, L.; Qin, Y.; Zhuang, Z.; Wang, X.; Huang, H.; Zhang, J.; Zhang, J.; et al. Flagellar Gene fliP Contributes to the Virulence of Pseudomonas plecoglossicida by Regulating Its Motility. Aquaculture 2023, 576, 739874. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Cheese | Type of Milk | Maturation (Days) | Origin of Collection | Origin of Production | Inspection Service |
|---|---|---|---|---|---|---|
| PT570 | Type Pecorino Toscano | P | 45 | RS | RS | SIF |
| PT211 | Type Pecorino Toscano | P | 90 | RS | RS | SIF |
| PT454 | Type Pecorino Toscano | P | 180 | RS | RS | SIF |
| PT917 | Type Pecorino Toscano | P | 270 | RS | RS | SIF |
| RE550 | Type Pecorino Toscano | P | 500 | RS | RS | SIF |
| ST056 | Colonial | P | UN | RS | RS | SISBI |
| ET945 | Colonial | P | UN | RS | RS | SISBI |
| QJ188 | Colonial | P | 60 | SC | SC | SISBI |
| PM697 | Matured | P | 90 | SC | SC | SISBI |
| GP209 | Matured | R | 90 | SC | SC | SISBI |
| VF712 | Matured | P | 180 | SC | SC | SISBI |
| PR551 | Pecorino Romano | R | UN | PR | IT | SIF |
| RA127 | Pecorino Romano | UN | UN | PR | IT | SIF |
| Phyla | Class | Order | Family | Genus | Species | Sample | TA 1 | BPP 2 | BHMT 3 |
|---|---|---|---|---|---|---|---|---|---|
| Actinobacteriota | Actinomycetia | Actinomycetales | Actinomycetaceae | Trueperella | T. pyogenes | PT211 | 0.112 | 0.132 | UC |
| Bacteroidota | Bacteroidia | Bacteroidales | Bacteroidaceae | Prevotella | Prevotella sp. | PT454 | 0.006 | 0.007 | UC |
| Flavobacteriales | Weeksellaceae | Kaistella | Kaistella sp. | RA127 | 0.006 | 0.007 | UC | ||
| K. haifensis | QJ188, GP209, PT211 | 0.216 | 0.255 | UC | |||||
| Firmicutes | Bacilli | Lactobacillales | Carnobacteriaceae | Alloiococcus | A. otitis | VF712 | 0.006 | 0.007 | UC |
| Listeriaceae | Brochothrix | B. thermosphacta | PM697 | 0.069 | 0.082 | UC | |||
| Staphylococcales | Staphylococcacea | Macrococcus | M. caseolyticus | GP209 | 0.020 | UC | 0.024 | ||
| Staphylococcus | Staphylococcus sp. | RA127, QJ188, GP209, PM697, VF712, PT211, PT570, PT917 | 0.803 | 0.944 | 0.955 | ||||
| S. chromogenes | GP209, PT570, PT917 | 0.120 | 0.142 | 0.143 | |||||
| S. equorum | QJ188, ST056, ET945 | 0.218 | 0.257 | 0.026 | |||||
| S. simulans | PT570 | 0.036 | 0.043 | 0.044 | |||||
| Clostridia | Tissierellales | Peptoniphilaceae | Helcococcus | H. ovis | PT211 | 0.185 | 0.192 | UC | |
| Fusobacteriota | Fusobacteriia | Fusobacteriales | Fusobacteriaceae | Fusobacterium | F. necrophorum | PM697 | 0.008 | 0.007 | UC |
| Klebsiella | K. ornithinolytica | ET945 | 0.190 | 0.224 | UC | ||||
| Pantoea | Pantoea sp. | VF712 | 0.185 | 0.139 | UC | ||||
| P. agglomerans | VF712 | 0.038 | 0.046 | UC | |||||
| Serratia | Serratia sp. | RA127 | 0.016 | 0.019 | 0.019 | ||||
| Shewanellaceae | Shewanella | S. baltica | ET945 | 0.012 | 0.014 | 0.015 | |||
| Pseudomonadales | Moraxellaceae | Acinetobacter | Acinetobacter sp. | VF712 | 3.799 | 4.465 | 4.515 | ||
| A. albensis | RA127, PR551, PM697, VF712, PT917, ET945 | 49.535 | 58.222 | 58.857 | |||||
| A. johnsonii | VF712 | 0.020 | 0.024 | 0.024 | |||||
| A. pseudolwoffii | VF712 | 0.020 | 0.024 | 0.024 | |||||
| A. ursingii | PT211, RE550 | 0.022 | 0.026 | 0.027 | |||||
| Moraxella | Moraxella sp. | PT211 | 0.004 | 0.005 | UC | ||||
| Pseudomonadaceae | Pseudomonas | Pseudomonas sp. | RA127, QJ188, GP209, PR551, PM697, VF712, PT211, ST056, RE550, PT570, PT917, ET945 | 29.389 | 34.660 | 35.038 | |||
| P. lundensis | ET945 | 0.028 | 0.034 | 0.034 | |||||
| P. weihenstephanensis | QJ188 | 0.018 | 0.022 | 0.022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pedroso, M.A.P.; Oliveira, W.d.C.; Felice, A.G.; Cunha, S.C.S.; Oliveira, M.B.P.P.; Richards, N.S.P.d.S. Pathogenic and Harmful Bacteria in Dairy Technology: Genomic Characterization and Its Correlation with Physicochemical Parameters of Sheep’s Cheese Sold in Southern Brazil. Fermentation 2024, 10, 397. https://doi.org/10.3390/fermentation10080397
Pedroso MAP, Oliveira WdC, Felice AG, Cunha SCS, Oliveira MBPP, Richards NSPdS. Pathogenic and Harmful Bacteria in Dairy Technology: Genomic Characterization and Its Correlation with Physicochemical Parameters of Sheep’s Cheese Sold in Southern Brazil. Fermentation. 2024; 10(8):397. https://doi.org/10.3390/fermentation10080397
Chicago/Turabian StylePedroso, Matheus Abraão Piovesan, Wemerson de Castro Oliveira, Andrei Giacchetto Felice, Sara Cristina Silva Cunha, Maria Beatriz Prior Pinto Oliveira, and Neila Silvia Pereira dos Santos Richards. 2024. "Pathogenic and Harmful Bacteria in Dairy Technology: Genomic Characterization and Its Correlation with Physicochemical Parameters of Sheep’s Cheese Sold in Southern Brazil" Fermentation 10, no. 8: 397. https://doi.org/10.3390/fermentation10080397
APA StylePedroso, M. A. P., Oliveira, W. d. C., Felice, A. G., Cunha, S. C. S., Oliveira, M. B. P. P., & Richards, N. S. P. d. S. (2024). Pathogenic and Harmful Bacteria in Dairy Technology: Genomic Characterization and Its Correlation with Physicochemical Parameters of Sheep’s Cheese Sold in Southern Brazil. Fermentation, 10(8), 397. https://doi.org/10.3390/fermentation10080397