
Production of Hydrogen with Ruminal Microbiota: Finding Culture Conditions for High Yields

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. General Procedures
2.2.1. Incubation Time
2.2.2. Selection of Culture Media and Preparation of Culture Medium
2.2.3. Headspace
2.2.4. Heat Treatment
2.2.5. Production and Productivity of Hydrogen
2.2.6. Cell Growth
2.3. Inoculum Preparation
2.4. Production and Quantification of Hydrogen
2.5. Massive Sequencing
2.6. Statistical Analysis
3. Results and Discussion
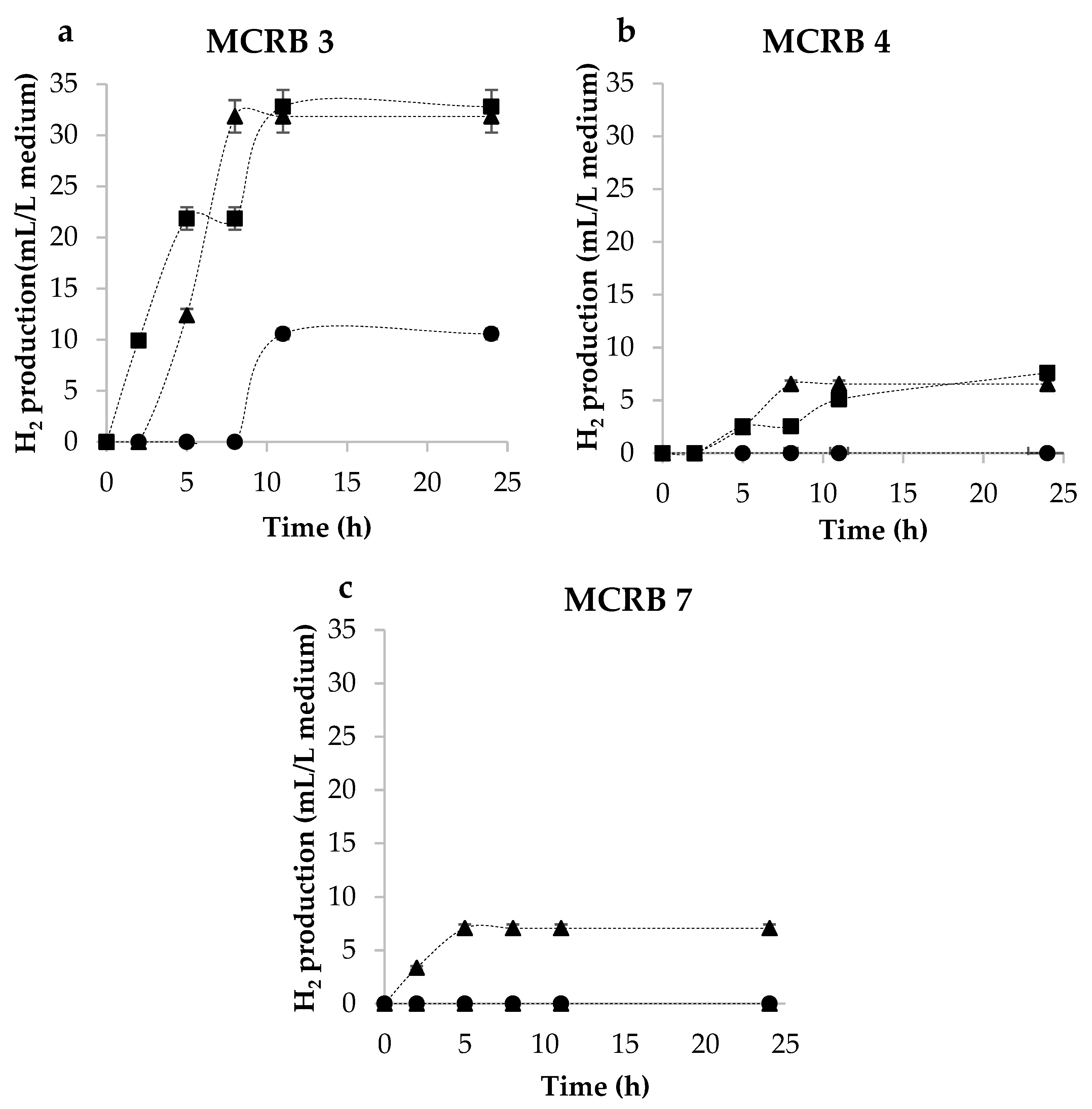
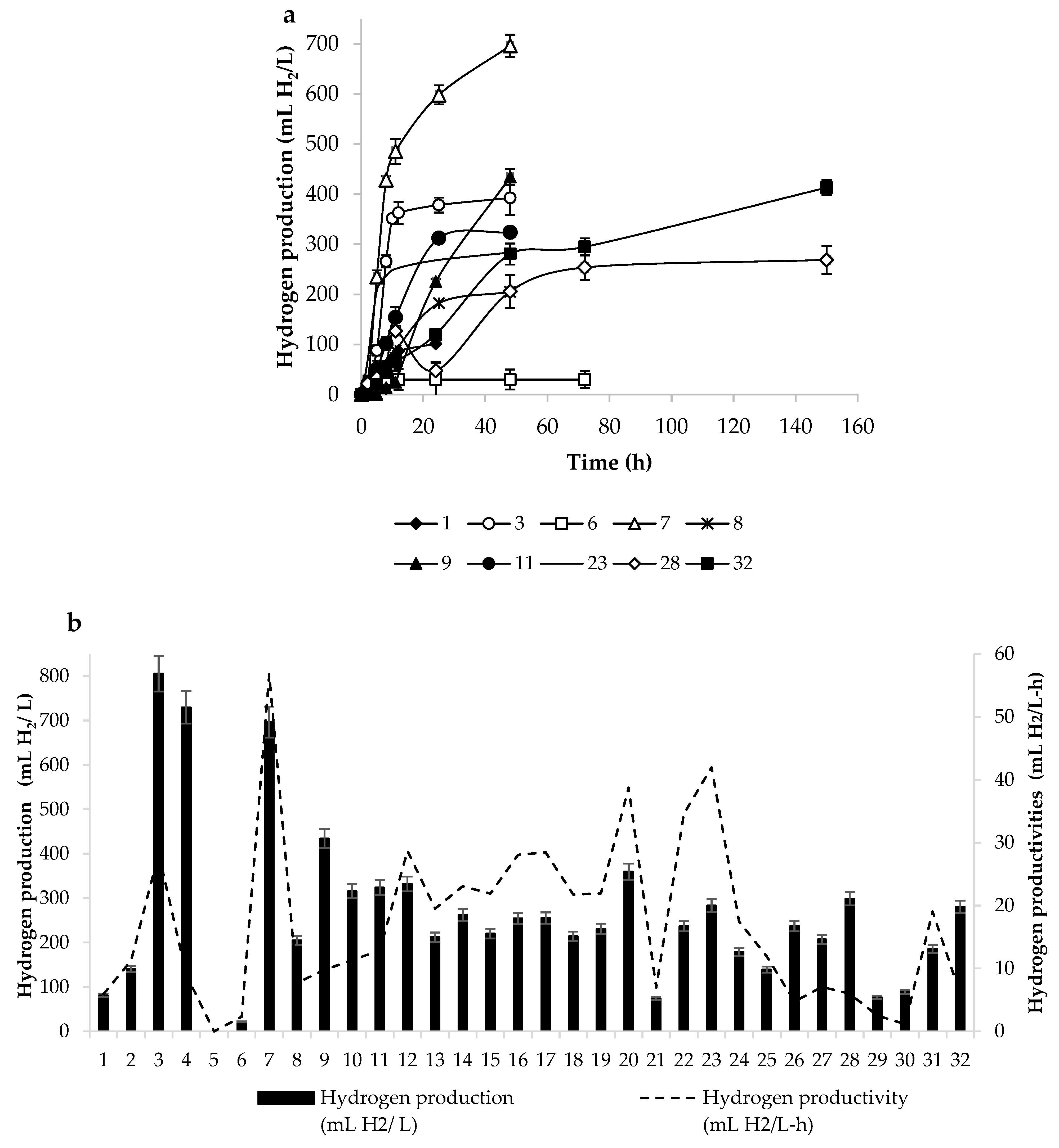
3.1. Incubation Time
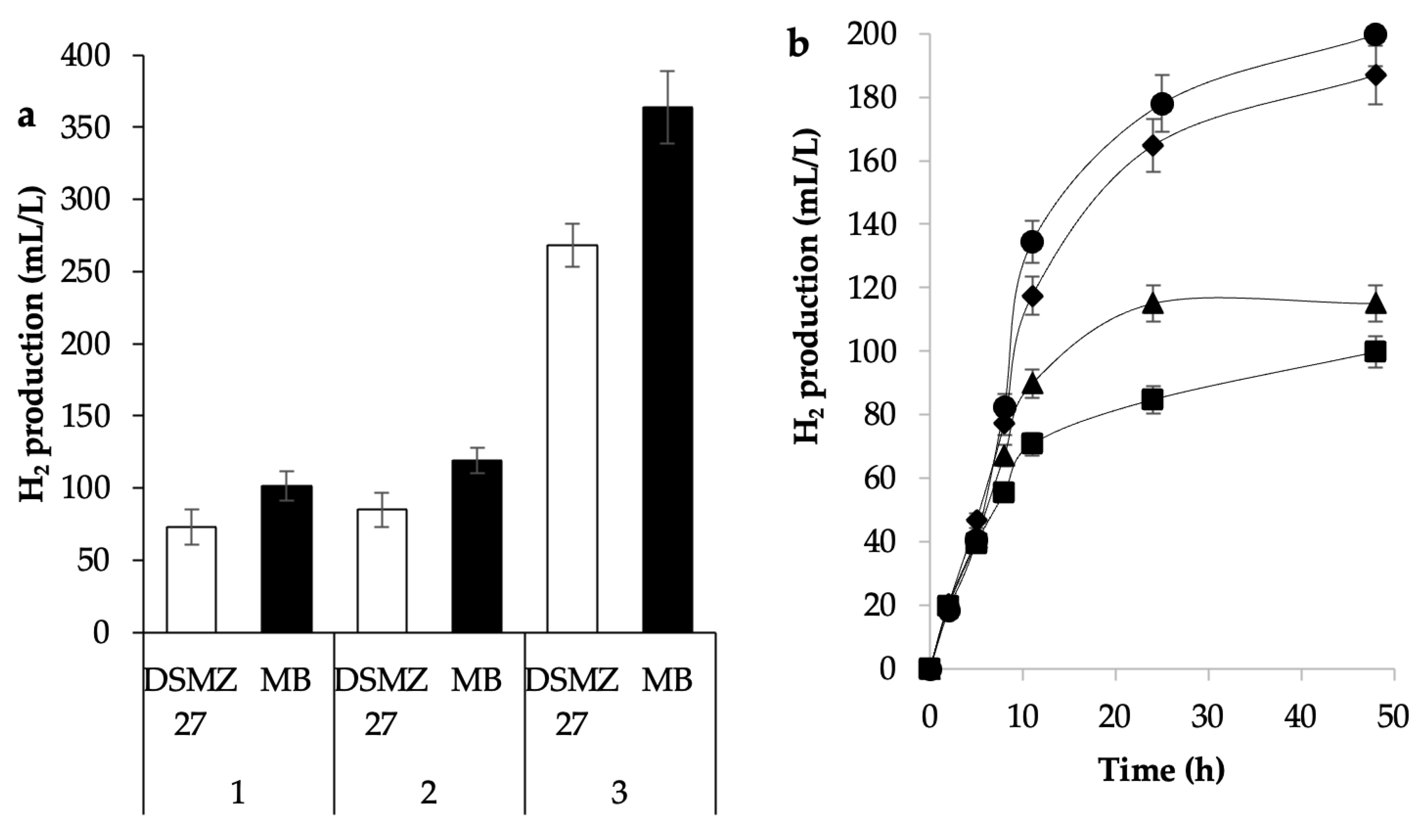
3.2. Culture Media to Produce Hydrogen
3.2.1. Composition of Culture Medium
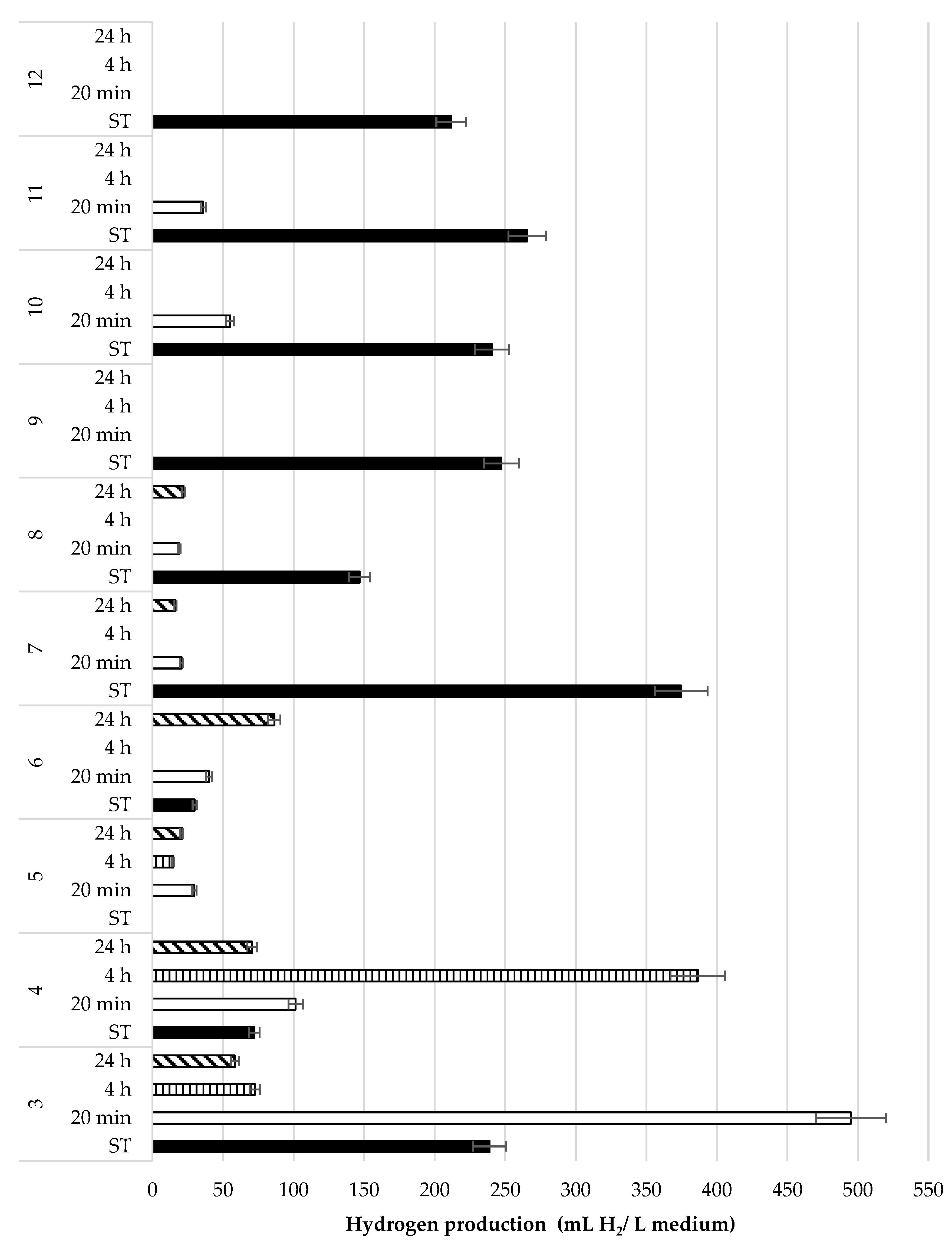
3.2.2. Culture Medium Preparation Conditions
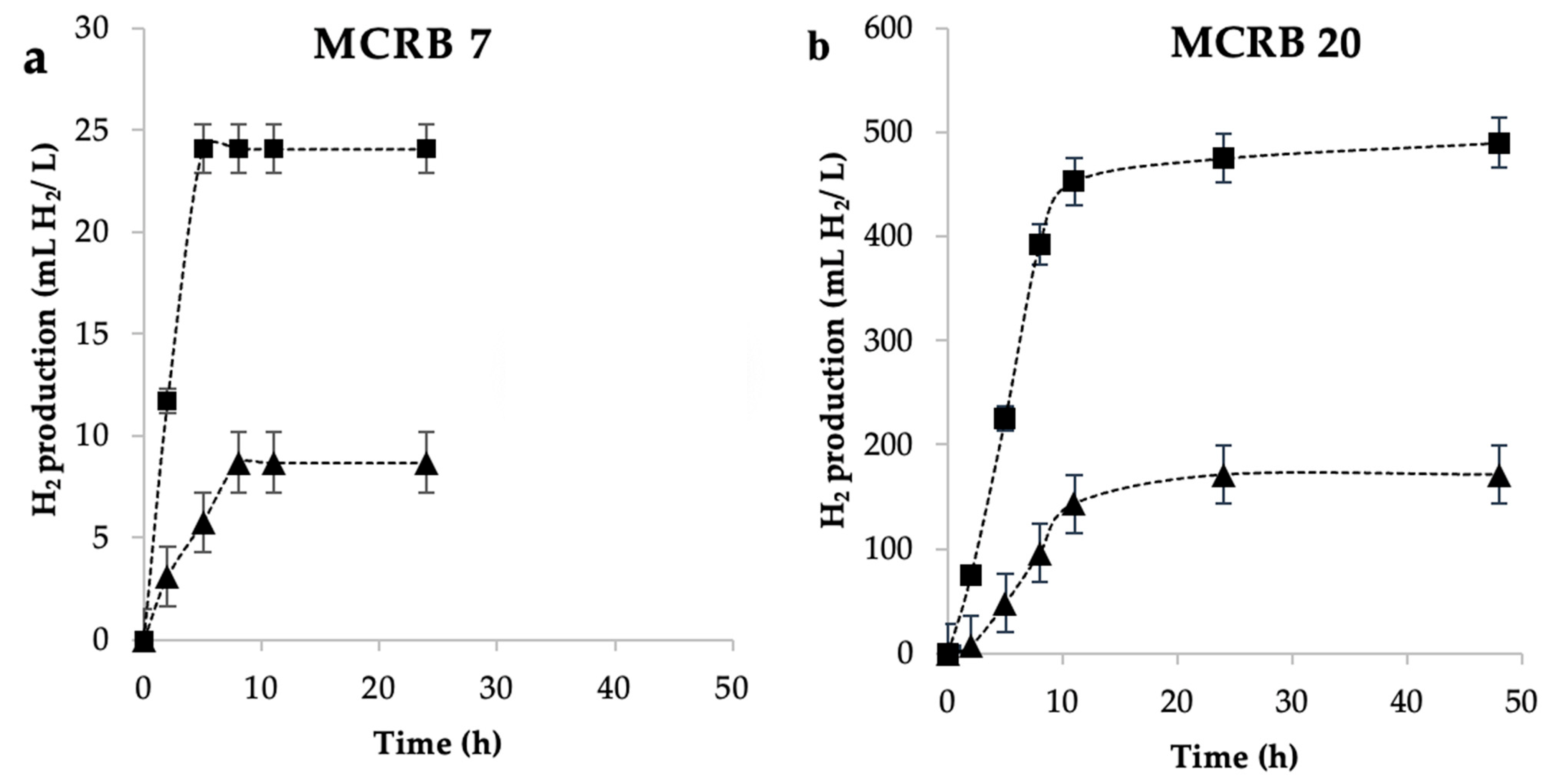
3.3. Effect of Culture Gas Volume
3.4. Thermal Treatment
3.5. Kinetics of Hydrogen Production
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lukajtis, R.; Hołowacz, I.; Kucharska, K.; Glinka, M.; Rybarczyk, P.; Przyjazny, A.; Kamiński, M. Hydrogen production from biomass using dark fermentation. Renew. Sustain. Energy Rev. 2018, 91, 665–694. [Google Scholar] [CrossRef]
- Yadav, S.; Singh, V.; Mahata, C.; Das, D. Optimization for simultaneous enhancement of biobutanol and biohydrogen production. Int. J. Hydrogen Energy. 2021, 46, 3726–3741. [Google Scholar] [CrossRef]
- García-Sánchez, R.; Ramos-Ibarra, R.; Guatemala-Morales, G.; Arriola-Guevara, E.; Toriz-González, G.; Corona-González, R.I. Photofermentation of tequila vinasses by Rhodopseudomonas pseudopalustris to produce hydrogen. Int. J. Hydrogen Energy 2018, 43, 15857–15869. [Google Scholar] [CrossRef]
- Teke, G.M.; Anye Cho, B.; Bosman, C.E.; Mapholi, Z.; Zhang, D.; Pott, R.W.M. Towards industrial biological hydrogen production: A review. World J. Microbiol. Biotechnol. 2024, 40, 37. [Google Scholar]
- IEA 50. Hydrogen. Overview. Conference. Last Update on 10 July 2023. Available online: https://www.iea.org/energy-system/low-emission-fuels/hydrogen (accessed on 6 May 2024).
- Katebah, M.; Al-Rawashdeh, M.; Linke, P. Analysis of hydrogen production costs in Steam-Methane Reforming considering integration with electrolysis and CO2 capture. Clean Eng. Technol. 2022, 10, 100552. [Google Scholar] [CrossRef]
- Kayfeci, M.; Keçebaş, A.; Bayat, M. Hydrogen production. In Solar Hydrogen Production: Processes, Systems and Technologies; Elsevier: Amsterdam, The Netherlands, 2019; Chapter 3; pp. 45–83. [Google Scholar] [CrossRef]
- Soares, J.F.; Confortin, T.C.; Todero, I.; Mayer, F.D.; Mazutti, M.A. Dark fermentative biohydrogen production from lignocellulosic biomass: Technological challenges and future prospects. Renew. Sustain. Energy Rev. 2020, 117, 109484. [Google Scholar] [CrossRef]
- Nikolaidis, P.; Poullikkas, A. A comparative overview of hydrogen production processes. Renew. Sustain. Energy Rev. 2017, 67, 597–611. [Google Scholar] [CrossRef]
- Ubando, A.T.; Chen, W.H.; Hurt, D.A.; Conversion, A.; Rajendran, S.; Lin, S.L. Biohydrogen in a circular bioeconomy: A critical review. Bioresource Technol. 2022, 366, 128168. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wan, W. Factors influencing fermentative hydrogen production: A review. Int. J. Hydrogen Energy 2009, 34, 799–811. [Google Scholar] [CrossRef]
- Nissila, M.E.; Tähti, H.P.; Rintala, J.A.; Puhakka, J.A. Thermophilic hydrogen production from cellulose with rumen fluid enrichment cultures: Effects of different heat treatments. Hydrogen Energy 2010, 36, 1482–1490. [Google Scholar] [CrossRef]
- DePeters, E.J.; George, L.W. Rumen transfaunation. Immunol. Lett. 2014, 162, 69–76. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, R.; Chang, J.; Chen, L.; Nabi, M.; Zhang, H.; Zhang, G.; Zhang, P. Rumen microbes, enzymes, metabolisms, and application in lignocellulosic waste conversion-A comprehensive review. Biotechnol. Adv. 2024, 71, 108308. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Nabi, M.; Zhang, P.; Zhang, G.; Cai, Y.; Wang, O.; Zhou, Z.; Ding, Y. Promising biological conversion of lignocellulosic biomass to renewable energy with rumen microorganisms: A comprehensive review. Renew. Sustain. Energy Rev. 2020, 134, 110335. [Google Scholar] [CrossRef]
- Liang, J.; Fang, W.; Wang, Q.; Zubair, M.; Zhang, G.; Ma, W.; Cai, Y.; Zhang, P. Metagenomic analysis of community, enzymes and metabolic pathways during corn straw fermentation with rumen microorganisms for volatile fatty acid production. Bioresour. Technol. 2021, 342, 126004. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef]
- Ratti, R.P.; Botta, L.S.; Sakamoto, I.K.; Silva, E.L.; Amancio, M.B. Production of H2 from cellulose by rumen microorganisms: Effects of inocula pre-treatment and enzymatic hydrolysis. Biotecnol. Lett. 2014, 36, 537–546. [Google Scholar] [CrossRef]
- Botta, L.S.; Ratti, R.P.; Sakamoto, I.K.; Rodrigues, L.; Silva, E.L.; Amâncio, M.B. Bioconversion of waste office paper to hydrogen using pretreated rumen fluid inoculum. Bioprocess Biosyst. Eng. 2016, 39, 1887–1897. [Google Scholar] [CrossRef] [PubMed]
- Fraga, M.; Perelmuter, K.; Valencia, M.J.; Cajarville, C.; Zunino, P. Caracterización de la microbiota bacteriana ruminal de un bovino a pastoreo mediante técnicas clásicas e independientes del cultivo. Veterinaria (Montev.) 2013, 49, 40–55. Available online: https://www.revistasmvu.com.uy/index.php/smvu/article/view/218 (accessed on 15 March 2024).
- DSMZ. DSMZ-German Collection of Microorganisms and Cell Cultures GmbH. [WWW Document]. List Recomm Media Microorg. 2022. Available online: https://www.dsmz.de/collection/catalogue/microorganisms/culture-technology/list-of-media-for-microorganisms (accessed on 26 October 2023).
- Schären, M.; Kiri, K.; Riede, S.; Gardener, M.; Meyer, U.; Hummel, J.; Urich, T.; Breves, G.; Dänicke, S. Alterations in the rumen liquid- particle- and epithelium-associated microbiota of dairy cows during the transition from a silage- and concentrate-based ration to pasture in spring. Front. Microbiol. 2017, 8, 744. [Google Scholar] [CrossRef]
- Ji, S.; Zhang, H.; Yan, H.; Azarfar, A.; Shi, H.; Alugongo, G.; Li, S.; Cao, Z.; Wang, Y. Comparison of rumen bacteria distribution in original rumen digesta, rumen liquid and solid fractions in lactating Holstein cows. J. Anim. Sci. Biotechnol. 2017, 8, 16. [Google Scholar] [CrossRef]
- Petri, R.M.; Forster, R.J.; Yang, W.; McKinnon, J.J.; McAllister, T.A. Characterization of rumen bacterial diversity and fermentation parameters in concentrate fed cattle with and without forage. J. Appl. Microbiol. 2012, 112, 1152–1162. [Google Scholar] [CrossRef] [PubMed]
- Franco-Leon, J.J.; Arriola-Guevara, E.; Suarez-Hernandez, L.A.; Toriz, G.; Guatemala-Morales, G.; Corona-Gonzalez, R.I. Influence of supplemented nutrients in tequila vinasses for hydrogen and polyhydroxybutyrate production by photofermentation with Rhodopseudomonas pseudopalustris. Bioresour Technol. 2021, 329, 124865. [Google Scholar] [CrossRef]
- Guneser, O.; Karagul Yuceer, Y. Effect of ultraviolet light on water- and fat-soluble vitamins in cow and goat milk. J. Dairy Sci. 2012, 95, 6230–6241. [Google Scholar] [CrossRef] [PubMed]
- Saleem, F.; Bouatra, S.; Guo, A.C.; Psychogios, N.; Mandal, R.; Dunn, S.M.; Ameta, B.N.; Wishart, D.S. The Bovine Ruminal Fluid Metabolome. Metabolomics 2013, 9, 360–378. [Google Scholar] [CrossRef]
- Castelló, E.; Nunes Ferraz, A.D.; Andreani, C.; Anzola-Rojas, M.P.; Borzacconi, L.; Buitrón, G.; Carrillo-Reyes, J.; Gomes, S.D.; Maintinguer, S.I.; Moreno-Andrade, I.; et al. Stability problems in the hydrogen production by dark fermentation: Possible causes and solutions. Renew. Sustain. Energy Rev. 2020, 119, 109602. [Google Scholar] [CrossRef]
- Palomo-Briones, R.; Celis, L.B.; Méndez-Acosta, H.O.; Bernet, N.; Trably, E.; Razo-Flores, E. Enhancement of mass transfer conditions to increase the productivity and efficiency of dark fermentation in continuous reactors. Fuel 2019, 254, 115648. [Google Scholar] [CrossRef]
- Brentner, L.B.; Jordan, P.A.; Zimmerman, J.B. Challenges in developing biohydrogen as a sustainable energy source: Implications for a research agenda. Environ. Sci. Technol. 2010, 44, 2243–2254. [Google Scholar] [CrossRef] [PubMed]
- Cheong, D.Y.; Hansen, C.L. Bacterial stress enrichment enhances anaerobic hydrogen production in cattle manure sludge. Appl. Microbiol. Biotechnol. 2006, 72, 35–643. [Google Scholar] [CrossRef] [PubMed]
- Arreola-Vargas, J.; Alatriste-Mondragón, F.; Celis, L.B.; Razo-Flores, E.; López-López, A.; Méndez-Acosta, H.O. Continuous hydrogen production in a trickling bed reactor by using triticale silage as inoculum: Effect of simple and complex substrates. J. Chem. Technol. Biotechnol. 2014, 90, 1062–1069. [Google Scholar] [CrossRef]
- Contreras-Dávila, C.A.; Méndez-Acosta, H.O.; Arellano-García, L.; Alatriste-Mondragón, F.; Razo-Flores, E. Continuous hydrogen production from enzymatic hydrolysate of Agave tequilana bagasse: Effect of the organic loading rate and reactor configuration. Chem. Eng. J. 2017, 313, 671–679. [Google Scholar] [CrossRef]
- Arreola-Vargas, J.; Flores-Larios, A.; González-Álvarez, V.; Corona-González, R.I.; Mendez-Acosta, H.O. Single and two-stage anaerobic digestion for hydrogen and methane production from acid and enzymatic hydrolysates of Agave tequilana bagasse. Int. J. Hydrogen Energy 2015, 41, 897–904. [Google Scholar] [CrossRef]
- Hobson, P.N. The Rumen Microbial Ecosystem; Blacki Academic & Professional: Aberdeen, UK, 1997. [Google Scholar] [CrossRef]
- Kamra, D.N. Rumen microbial ecosystem. Curr. Sci. 2005, 89, 124–135. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Culture medium | |
|---|---|---|
| MB | Rhodospirillaceae | |
| Concentration | ||
| (g/L) | ||
| Glucose | 1.0 | |
| Starch | 1.0 | |
| Xylose | 1.0 | |
| Yeast extract | 0.5 | 0.3 |
| NaCl | 0.90 | 0.4 |
| KH2PO4 | 0.45 | 0.5 |
| NH4 | 0.90 ((NH4)2SO4) | 0.4 (NH4Cl) |
| K2HPO4 | 0.45 | |
| Ferric (III) citrate | 0.5 | |
| (μM) | g/L | |
| NH4-acetate | 0.5 | |
| Na2-succinate | 1 | |
| Acetic acid | 3.60 | |
| Ac. propionic | 0.99 | |
| Ac. butyric | 0.53 | |
| Ac. n-valeric | 0.11 | |
| Ac. isovaleric | 0.11 | |
| (mg/L) | ||
| CaCl2·2H2O | 30 | 50 |
| MgCl2·6H2O | 20 | 400 |
| MnCl2·4H2O | 10 | 0.03 |
| CoCl2·6H2O | 10 | 0.2 |
| ZnSO4·7H2O | 0.1 | |
| CuCl2·2H2O | 0.01 | |
| NiCl2·6H2O | 0.02 | |
| Na2MoO4·2H2O | 0.03 | |
| H3BO3 | 0.3 | |
| Pyridoxine hydrochloride | 2.0 | |
| Riboflavin | 2.0 | |
| Thiamine hydrochloride | 2.0 | |
| Ac. pantothenic | 2.0 | |
| Ac. P-aminobenzoic | 0.1 | |
| B-12 vitamin | 25 | 0.4 |
| Ac. Folic | 500 | |
| Biotin | 500 | |
| Clarified rumen fluid | 200 mL/L | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gándara-Arteaga, V.M.; Guatemala-Morales, G.M.; Martínez-Gómez, Á.d.J.; Toriz, G.; Pelayo-Ortiz, C.; Corona-González, R.I. Production of Hydrogen with Ruminal Microbiota: Finding Culture Conditions for High Yields. Fermentation 2024, 10, 274. https://doi.org/10.3390/fermentation10060274
Gándara-Arteaga VM, Guatemala-Morales GM, Martínez-Gómez ÁdJ, Toriz G, Pelayo-Ortiz C, Corona-González RI. Production of Hydrogen with Ruminal Microbiota: Finding Culture Conditions for High Yields. Fermentation. 2024; 10(6):274. https://doi.org/10.3390/fermentation10060274
Chicago/Turabian StyleGándara-Arteaga, Vianca Maribel, Guadalupe María Guatemala-Morales, Álvaro de Jesús Martínez-Gómez, Guillermo Toriz, Carlos Pelayo-Ortiz, and Rosa Isela Corona-González. 2024. "Production of Hydrogen with Ruminal Microbiota: Finding Culture Conditions for High Yields" Fermentation 10, no. 6: 274. https://doi.org/10.3390/fermentation10060274
APA StyleGándara-Arteaga, V. M., Guatemala-Morales, G. M., Martínez-Gómez, Á. d. J., Toriz, G., Pelayo-Ortiz, C., & Corona-González, R. I. (2024). Production of Hydrogen with Ruminal Microbiota: Finding Culture Conditions for High Yields. Fermentation, 10(6), 274. https://doi.org/10.3390/fermentation10060274