


Oral Administration of GnRH and Domperidone via Gel Feed and Their Effect on Reproductive Performance of Devario devario (Bengal Danio), an Ornamental Fish


 , , , , ,
, , , , ,  , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
2.1. Reproductive Performance
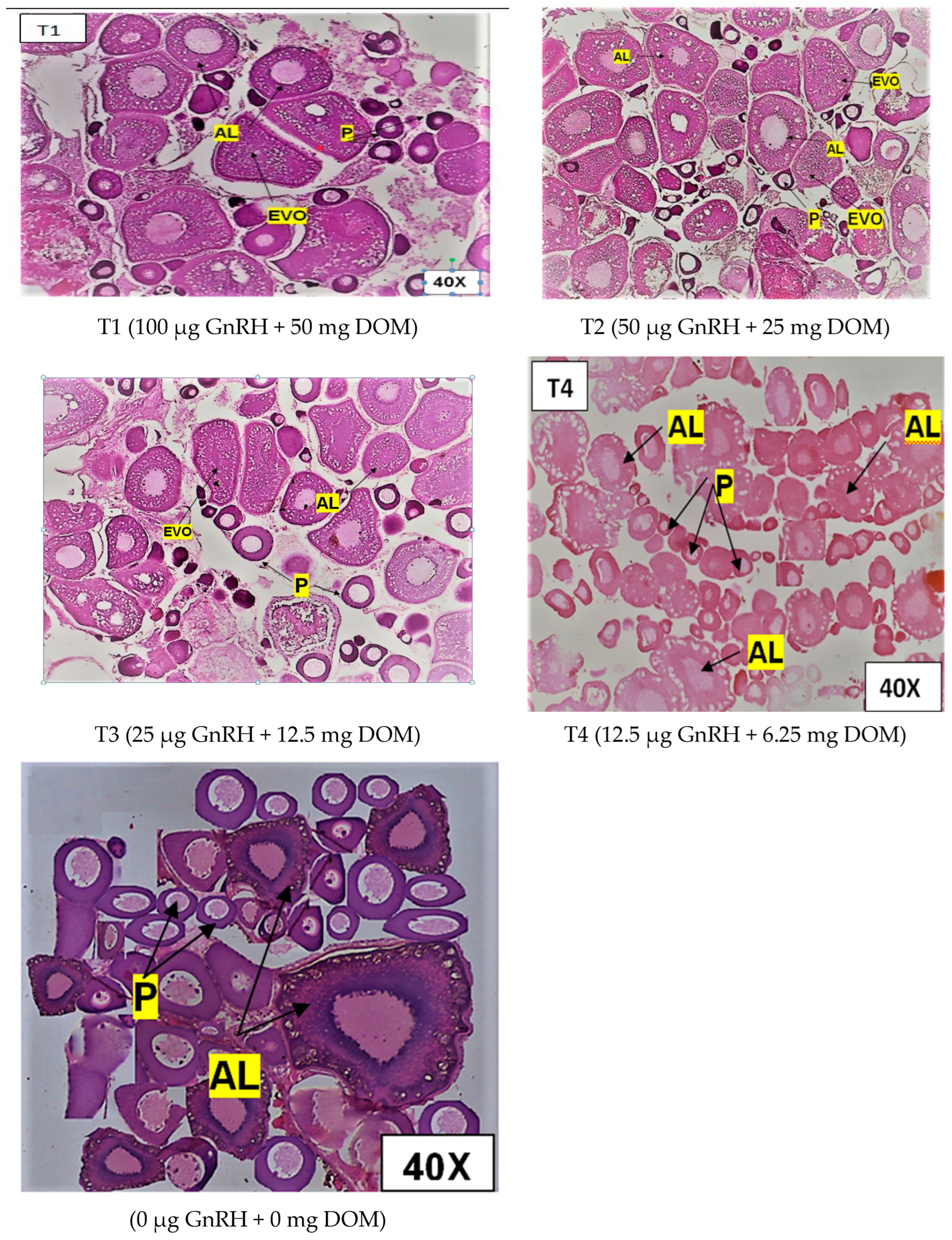
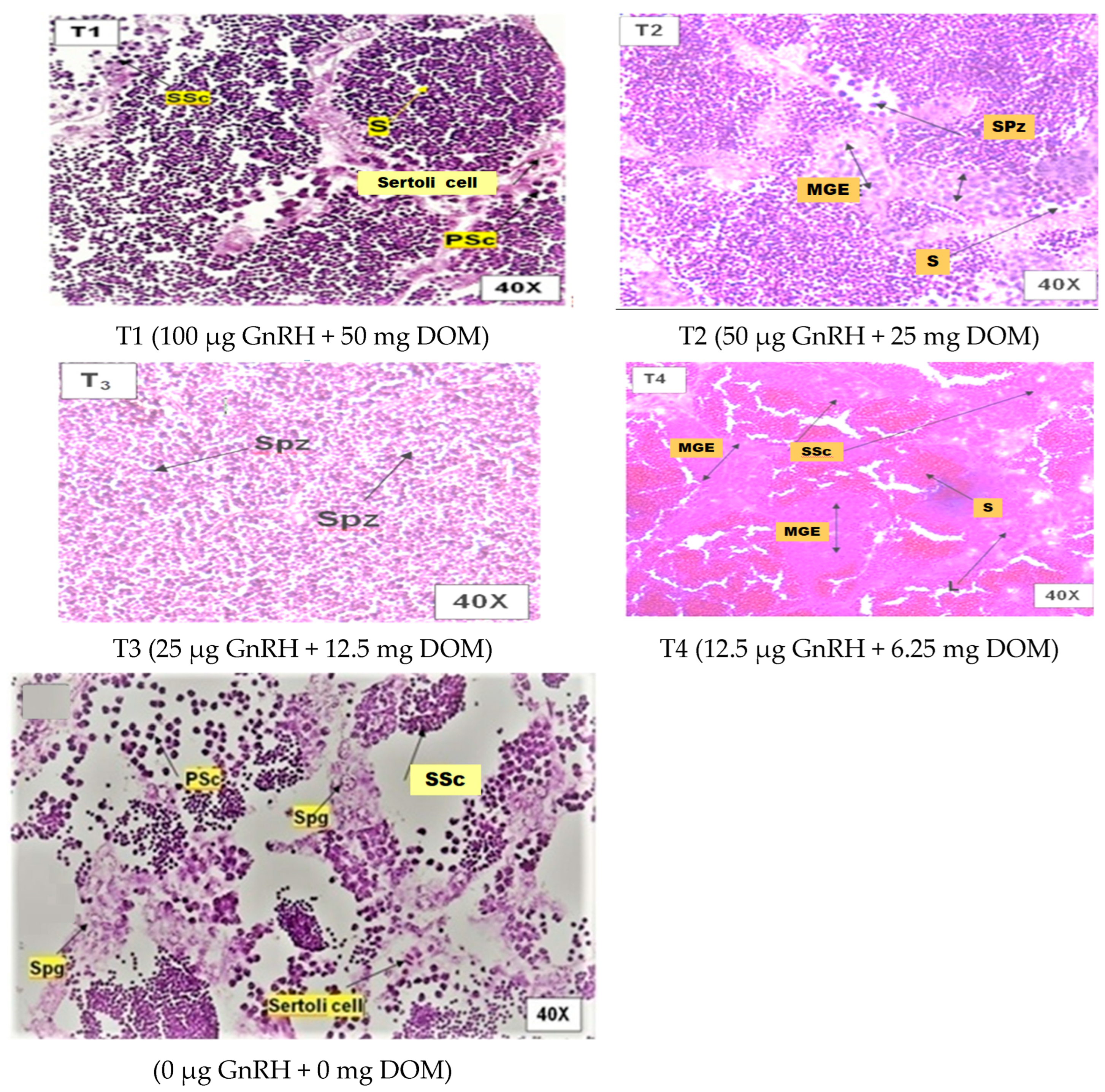
2.2. Histological Gonad Changes
2.3. Breeding Performance
2.4. Sperm Quality and Quantity
2.5. Stress Hormones
2.6. Antioxidant Enzymes Status
2.7. Levels of Sex Hormones
2.8. Discussion
2.8.1. Gonadosomatic Index (GSI)
2.8.2. Ovarian Maturation of Bengal Danio
2.8.3. Effect on Testicular Maturation
2.8.4. Effect on Reproductive Hormones
2.8.5. Effect on Sperm Parameters
2.8.6. Effect on Antioxidant Enzyme Activity
2.8.7. Effect on Breeding Performance of Fish
3. Conclusions
4. Materials and Methods
4.1. Animal Ethics
4.2. Experimental Fish and Husbandry
4.3. Formulation of Feed
4.4. Experimental Design and Set-Up
4.5. Water Quality Analysis
4.6. Reproductive Performance
4.7. Sperm Parameters
4.8. Breeding Performance
4.9. Reproductive Hormone
4.10. Antioxidant Enzyme Assay
4.11. Stress Parameters
4.11.1. Cortisol Level Analysis
4.11.2. Glucose Analysis
4.12. Histological Study of Gonads
4.13. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mylonas, C.C.; Zohar, Y. Endocrine regulation and artificial induction of oocyte maturation and spermiation in basses of the genus Morone. Aquaculture 2001, 202, 205–220. [Google Scholar] [CrossRef]
- Schreck, C.B.; Contreras-Sanchez, W.; Fitzpatrick, M.S. Effects of stress on fish reproduction, gamete quality, and progeny. In Reproductive Biotechnology in Finfish Aquaculture; Elsevier: Amsterdam, The Netherlands, 2001; pp. 3–24. [Google Scholar]
- Damle, D.; Kumar, R.; Ahilan, B.; Pillai, B.R.; Chidambaram, P.; Swain, P.P.; Debbarma, J.; Sundaray, J.K. The effect of habitat manipulation on early gonad maturation of Channa striata in captive condition. Indian. J. Anim. Res. 2023, 57, 1462–1468. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.; Shukry, M.; Saad, M.F.; Mohamed, N.A.; Nowosad, J.; Kucharczyk, D. Effects of GnRHa and hCG with or without dopamine receptor antagonists on the spawning efficiency of African catfish (Clarias gariepinus) reared in hatchery conditions. Anim. Reprod. Sci. 2021, 231, 106798. [Google Scholar] [CrossRef]
- Mohammadzadeh, S.; Moradian, F.; Yeganeh, S.; Falahatkar, B.; Milla, S. Design, production and purification of a novel recombinant gonadotropin-releasing hormone associated peptide as a spawning inducing agent for fish. Protein Expr. Purif. 2020, 166, 105510. [Google Scholar] [CrossRef]
- Mechaly, A.S.; Awruch, C.; Cabrita, E.; Costas, B.; Fernandes, J.M.; Gallego, V.; Fatsini, E. Cutting-Edge Methods in Teleost and Chondrichthyan Reproductive Biology. Rev. Fish. Sci. Aquac. 2024, 33, 77–112. [Google Scholar] [CrossRef]
- Weltzien, F.A.; Pasqualini, C.; Sébert, M.E.; Vidal, B.; Le Belle, N.; Kah, O.; Dufour, S. Androgen-dependent stimulation of brain dopaminergic systems in the female European eel (Anguilla anguilla). Endocrinology 2006, 147, 2964–2973. [Google Scholar] [CrossRef] [PubMed]
- Szukiewicz, D. Current Insights in Prolactin Signaling and Ovulatory Function in Health and Disease. Int. J. Mol. Sci. 2024, 25, 1976. [Google Scholar] [CrossRef] [PubMed]
- Kookaram, K.; Mojazi Amiri, B.; Dorkoosh, F.A.; Nematollahi, M.A.; Mortazavian, E.; Abed Elmdoust, A. Effect of oral administration of GnRHa+ nanoparticles of chitosan in oogenesis acceleration of goldfish (Carassius auratus). Fish. Physiol. Biochem. 2021, 47, 477–486. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Fostier, A.; Zanuy, S. Broodstock management and hormonal manipulations of fish reproduction. Gen. Comp. Endocrinol. 2010, 165, 516–534. [Google Scholar] [CrossRef]
- Klojdová, I.; Milota, T.; Smetanová, J.; Stathopoulos, C. Encapsulation: A strategy to deliver therapeutics and bioactive compounds? Pharmaceuticals 2023, 16, 362. [Google Scholar] [CrossRef]
- Lal, J.; Biswas, P.; Singh, S.K.; Debbarma, R.; Mehta, N.K.; Deb, S.; Patel, A.B. Moving towards Gel for Fish Feeding: Focus on Functional Properties and Its Acceptance. Gels 2023, 9, 305. [Google Scholar] [CrossRef] [PubMed]
- Shraborni, A.; Mandal, S.C.; Parhi, J. Freshwater Ornamental Fishes of India: Sustainable Management and Conservation. In Aquaculture and Conservation of Inland Coldwater Fishes; Springer Nature: Singapore, 2024; pp. 155–173. [Google Scholar]
- Felix, S. Developing ornamental aquaculture (Aquariculture) present scenario and scope in Tamil Nadu. Fish. Chimes 2009, 29, 8–9. [Google Scholar]
- Raja, K.; Aanand, P.; Padmavathy, S.; Sampathkumar, J.S. Present and future market trends of Indian ornamental fish sector. Int. J. Fish. Aquac. Sci. 2019, 7, 6–15. [Google Scholar]
- Mukherjee, R.; Maity, J. The Study of Freshwater Indigenous Ornamental Fishes Available in Selected Sites of Southern Bengal and Their Present Status in Natural Habitats. Asian J. Fish. Aqu. Res. 2023, 24, 1–9. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Saha, N.C. Consequences of sodium dodecyl sulfate exposure on the antioxidant status and steroidogenesis in fish gonad. Environ. Sci. Pollut. Res. 2021, 28, 19247–19259. [Google Scholar] [CrossRef]
- Wang, J.; Liu, W.; Li, P.; Tang, F.; Lu, W. Estimation of Coregonus ussuriensis age, growth, and maturation in China’s Amur River. PeerJ 2022, 10, e12817. [Google Scholar] [CrossRef]
- Sahadan, F.N.; Christianus, A.; Ina-Salwany, M.Y.; Ismail, F.S.; Othman, R.; Zulperi, Z. Gonadotropin-releasing hormone (GnRH)-its approaches to improve reproduction in fish. Sains Malays. 2022, 51, 3539–3549. [Google Scholar] [CrossRef]
- Mondal, P.; Hira, S.K.; Saha, N.C. The functional role of melatonin and kisspeptin in fish reproduction. Innov. J. Sci. 2019, 7, 1–7. [Google Scholar]
- Bhat, S.K.; Ganesh, C.B. Domperidone treatment attenuates stress-induced suppression of reproduction in viviparous mosquitofish Gambusia affinis. J. Fish. Biol. 2020, 96, 37–48. [Google Scholar] [CrossRef]
- Acharyya, A.; Das, J.; Hasan, K.N. Melatonin as a multipotent component of fish feed: Basic information for its potential application in aquaculture. Front. Mar. Sci. 2021, 8, 734066. [Google Scholar] [CrossRef]
- Zupa, R.; Duncan, N.; Giménez, I.; Mylonas, C.C.; Pousis, C.; Passantino, L.; Corriero, A. Male germ cell proliferation and apoptosis in sexually immature meagre Argyrosomus regius (Asso, 1801) treated with recombinant follicle stimulating hormone. Sci. Rep. 2023, 13, 7013. [Google Scholar] [CrossRef]
- Feng, K.; Cui, X.; Song, Y.; Tao, B.; Chen, J.; Wang, J.; Hu, W. Gnrh3 regulates PGC proliferation and sex differentiation in developing zebrafish. Endocrinology 2020, 161, bqz024. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, T.; Bhandari, R.K.; Kobayashi, Y.; Nakamura, S.; Nakamura, M. GnRHa-accelerated spermatogenesis in the testes of underyearling golden rabbitfish (Siganus guttatus) (Bloch). Aquaculture 2006, 257, 558–565. [Google Scholar] [CrossRef]
- Guzmán, J.M.; Cal, R.; García-López, Á.; Chereguini, O.; Kight, K.; Olmedo, M.; Mañanós, E.L. Effects of in vivo treatment with the dopamine antagonist pimozide and gonadotropin-releasing hormone agonist (GnRHa) on the reproductive axis of Senegalese sole (Solea senegalensis). Comp. Biochem. Physiol. 2011, 158, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Q.; Zhang, Y.; Huang, W.; Liu, X.; Li, Y.; Zhu, P.; Lin, H. Gonadotropin-releasing hormone analogue multiple injection potentially accelerated testicular maturation of male yellow catfish (Pelteobagrusfluvidraco, Richardson) in captivity. Aquac. Res. 2012, 43, 467–480. [Google Scholar] [CrossRef]
- Kim, S.W.; Hong, W.H.; Han, S.J.; Kwon, J.; Ko, H.; Lee, S.B.; Park, S.C. Use of synthetic salmon GnRH and Domperidone (Ovaprim®) in sharks: Preparation for ex situ conservation. Front. Mar. Sci. 2020, 7, 571741. [Google Scholar] [CrossRef]
- Kumakura, N.; Okuzawa, K.; Gen, K.; Kagawa, H. Effects of gonadotropin-releasing hormone agonist and dopamine antagonist on hypothalamus–pituitary–gonadal axis of pre-pubertal female red seabream (Pagrus major). Gen. Comp. Endocrinol. 2003, 131, 264–273. [Google Scholar] [CrossRef]
- Iegorova, V.; Naraine, R.; Psenicka, M.; Zelazowska, M.; Sindelka, R. Comparison of RNA localization during oogenesis within Acipenser ruthenus and Xenopus laevis. Front. Cell Dev. Biol. 2022, 10, 982732. [Google Scholar] [CrossRef]
- Li, C.; Meng, X.; Liu, S.; Li, W.; Zhang, X.; Zhou, J.; Liu, H. Oocytes and hypoxanthine orchestrate the G2-M switch mechanism in ovarian granulosa cells. Development 2020, 147, dev184838. [Google Scholar] [CrossRef]
- Tokumoto, T.; Tokumoto, M.; Horiguchi, R.; Ishikawa, K.; Nagahama, Y. Diethylstilbestrol induces fish oocyte maturation. Proc. Natl. Acad. Sci. USA 2004, 101, 3686–3690. [Google Scholar] [CrossRef]
- Rizzo, E.; Bazzoli, N. Reproduction and embryogenesis. In Biology and Physiology of Freshwater Neotropical Fish; Academic Press: Cambridge, MA, USA, 2020; pp. 287–313. [Google Scholar]
- Ramasubramanian, V.; Shabana, M.S.; Ragunath, C. Effect of Vitellogenin in the Growth of Ornamental Fishes. In Vitellogenin in Fishes-Diversification, Biological Properties, and Future Perspectives; Springer Nature: Singapore, 2023; pp. 251–269. [Google Scholar]
- Babin, P.J.; Carnevali, O.; Lubzens, E.; Schneider, W.J. Molecular aspects of oocyte vitellogenesis in fish. In The Fish Oocyte; Springer: Berlin/Heidelberg, Germany, 2007; pp. 39–76. [Google Scholar]
- Asiah, N.; Aryani, N.; Nuraini, N.; Lukistyowaty, I.; Harjoyudanto, Y. Effects of GnRH and Anti-Dopamine on Gonad Maturation of Osteochilus melanopleurus (Bleeker, 1852). IOP Conf. Ser. Earth Environ. Sci. 2021, 934, 12–14. [Google Scholar] [CrossRef]
- Lee, C.S.; Tamaru, C.S. Advances and future prospects of controlled maturation and spawning of grey mullet (Mugil cephalus L.) in captivity. Aquaculture 1988, 74, 63–73. [Google Scholar] [CrossRef]
- Ramos-Judéz, S.; Chauvigné, F.; González-López, W.Á.; Rosenfeld, H.; Cerdà, J.; Giménez, I.; Duncan, N. Providing recombinant gonadotropin-based therapies that induce oogenesis from previtellogenic oocytes to produce viable larvae in a teleost, the flathead grey mullet (Mugil cephalus). Aquaculture 2021, 536, 736418. [Google Scholar] [CrossRef]
- Monbrison, D.D.; Tzchori, I.; Holland, M.C.; Zohar, Y.; Yaron, Z.; Elizur, A. Acceleration of gonadal development and spawning induction in the Mediterranean grey mullet, (Mugil cephalus): Preliminary studies. Isr. J. Aquac. 1997, 49, 214–221. [Google Scholar]
- Zohar, Y.; Zmora, N.; Trudeau, V.L.; Muñoz-Cueto, J.A.; Golan, M. A half century of fish gonadotropin-releasing hormones: Breaking paradigms. J. Neuroendocrinol. 2022, 34, e13069. [Google Scholar] [CrossRef]
- Zohar, Y.; Mylonas, C.C. Endocrine manipulations of spawning in cultured fish: From hormones to genes. In Reproductive Biotechnology in Finfish Aquaculture; Elsevier: Amsterdam, The Netherlands, 2001; pp. 99–136. [Google Scholar]
- Oduwole, O.O.; Peltoketo, H.; Huhtaniemi, I.T. Role of follicle-stimulating hormone in spermatogenesis. Front. Endocrinol. 2018, 9, 763. [Google Scholar] [CrossRef]
- Nasello, A.G.; Vanzeler, M.L.; Madureira, E.H.; Felicio, L.F. Effects of acute and long-term domperidone treatment on prolactin and gonadal hormone levels and sexual behavior of male and female rats. Pharmacol. Biochem. Behav. 1997, 58, 1089–1094. [Google Scholar] [CrossRef]
- de Araújo Silva, E.F.; da Silva Gomes, J.A.; de Oliveira, M.L.F.; de Carvalho, A.G.A.F.; Magalhães, C.P.; da Silva, J.V.; da Silva, L.H.; Tenorio, B.M.; Valença, M.M.; Tenorio, F.D.C.A.M. Protective effect of exogenous melatonin on testicular histopathology and histomorphometry of adult rats with domperidone-induced hyperprolactinemia. Reprod. Biol. 2023, 23, 100791. [Google Scholar] [CrossRef]
- Roelants, I.; Mikolajczyk, T.; Epler, P.; Ollevier, F.; Chyb, J.; Breton, B. Induction of spermiation in common carp after enhanced intestinal uptake of sGnRH-A and Pimozide. J. Fish. Biol. 2000, 56, 1398–1407. [Google Scholar]
- Zadmajid, V. Comparative effects of human chorionic gonadotropin (hCG) and Ovaprim™ (sGnRHa+ domperidone) on the reproductive characteristics of wild-caught male Longspine scraper, Capoeta trutta (Heckel, 1843). Aquaculture 2016, 463, 7–15. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Hasan, K.N.; Maitra, S.K. Melatonin actions on ovaprim (synthetic GnRH and domperidone)-induced oocyte maturation in carp. Reproduction 2016, 151, 285–296. [Google Scholar] [CrossRef]
- Specker, J.L. Vitellogenesis in fishes: Status and perspectives. In Perspectives in Comparative Endocrinology; National Research Council: Ottawa, ON, Canada, 1994; pp. 304–315. [Google Scholar]
- Cejko, B.I.; Żarski, D.; Judycka, S.; Kucharczyk, D.; Sarosiek, B.; Kowalski, R.K. Effect of two commercial preparations containing different GnRH analogues with dopamine antagonists on barbel Barbus barbus (L.) sperm quantity and quality. Aquac. Int. 2014, 22, 97–109. [Google Scholar] [CrossRef]
- Kowalski, R.K.; Hliwa, P.; Cejko, B.I.; Król, J.; Stabiński, R.; Ciereszko, A. Quality and quantity of smelt (Osmerus eperlanus L.) sperm in relation to time after hormonal stimulation. Reprod. Bio. 2012, 12, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Sorbera, L.A.; Mylonas, C.C.; Zanuy, S.; Carrillo, M.; Zohar, Y. Sustained administration of GnRHa increases milt volume without altering sperm counts in the sea bass. J. Exp. Zool. 1996, 276, 361–368. [Google Scholar] [CrossRef]
- Lim, H.K.; Pankhurst, N.W.; Fitzgibbon, Q.P. Effects of slow-release gonadotropin releasing hormone analog on milt characteristics and plasma levels of gonadal steroids in greenback flounder, Rhombosolea tapirina. Aquaculture 2004, 240, 505–516. [Google Scholar] [CrossRef]
- Clearwater, S.J.; Crim, L.W. Gonadotropin releasing hormone-analogue treatment increases sperm motility, seminal plasma pH and sperm production in yellowtail flounder (Pleuronectes ferrugineus). Fish. Physiol. Biochem. 1998, 19, 349–357. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Berger, B.; Weismann, T.; Patzner, R.A. Motility of spermatozoa of Alburnus alburnus (Cyprinidae) and its relationship to seminal plasma composition and sperm metabolism. Fish. Physiol. Biochem. 1996, 15, 167–179. [Google Scholar] [CrossRef]
- Zadmajid, V.; Bashiri, S.; Sharafi, N.; Butts, I.A.E. Effect of hCG and Ovaprim™ on reproductive characteristics of male Levantine scraper, Capoetadamascina (Valenciennes, 1842). Theriogenology 2018, 115, 45–56. [Google Scholar] [CrossRef]
- Podhorec, P.; Knowles, J.; Vysloužil, J.; Boryshpolets, S.; Sotnikov, A.; Holická, M.; Dzyuba, B. The effect of hormonal treatment on selected sperm quality parameters and sex steroids in tropical cyprinid bala shark Balantiocheilosmelanopterus. Fishes 2022, 7, 122. [Google Scholar] [CrossRef]
- Eslami, M.M.; Javadian, S.R.; Bahram, S. The effect of different doses of GnRH on stress responses in female koi carp (Cyprinus carpio). Iran. J. Aquat. Anim. Health 2021, 7, 61–68. [Google Scholar] [CrossRef]
- Pankhurst, N.W. The endocrinology of stress in fish: An environmental perspective. Gen. Comp. Endocrinol. 2011, 170, 265–275. [Google Scholar] [CrossRef]
- Davies, K.J. An overview of oxidative stress. IUBMB Life 2000, 50, 241–244. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Carmo de Carvalho e Martins, M.D.; Martins da Silva Santos Oliveira, A.S.; da Silva, L.A.A.; Primo, M.G.S.; de Carvalho Lira, V.B. Biological indicators of oxidative stress [malondialdehyde, catalase, glutathione peroxidase, and superoxide dismutase] and their application in nutrition. In Biomarkers in Nutrition; Springer International Publishing: Cham, Switzerland, 2022; pp. 1–25. [Google Scholar]
- Tamura, H.; Takasaki, A.; Miwa, I.; Taniguchi, K.; Maekawa, R.; Asada, H.; Sugino, N. Oxidative stress impairs oocyte quality and melatonin protects oocytes from free radical damage and improves fertilization rate. J. Pineal Res. 2008, 44, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Yuniarti, T.; Basuki, F.; Hastuti, S.; Nugroho, R.A.; Marantika, S. Reproductive Performance of Java Barb (Punctius javanicus) Injected sGNRH and Domperidone of Different Dosage. IOP Conf. Ser. Earth Environ. Sci. 2021, 750, 12–25. [Google Scholar] [CrossRef]
- Campbell, B.; Dickey, J.; Beckman, B.; Young, G.; Pierce, A.; Fukada, H.; Swanson, P. Previtellogenic oocyte growth in salmon: Relationships among body growth, plasma insulin-like growth factor-1, estradiol-17beta, follicle-stimulating hormone and expression of ovarian genes for insulin-like growth factors, steroidogenic-acute regulatory protein and receptors for gonadotropins, growth hormone, and somatolactin. Biol. Reprod. 2006, 75, 34–44. [Google Scholar] [PubMed]
- Nuraini; Alawi, H.; Aryani, N.d.N. Pengaruhs GnRH + Domperidon dan Waktu Laten Berbedaterhadap Ovulasi dan Penetasan Telur Ikan Selais. Berk. Perikan. Terubuk 2013, 41, 1–8. [Google Scholar]
- Chowdhury, S.; Saikia, S.K. Oxidative Stress in Fish: A Review. J. Sci. Res. 2020, 12, 145–160. [Google Scholar] [CrossRef]
- Oyen, F.G.F.; Camps, L.E.C.M.M.; Bonga, S.W. Effect of acid stress on the embryonic development of the common carp (Cyprinus carpio). Aquat. Toxicol. 1991, 19, 1–12. [Google Scholar] [CrossRef]
- Syaifudin, S.M.; Sulmartiwi, D.L.; Andriyono, S. Penambahan Mikroalga Merah Porphyridiumcruentum pada Pakan terhadapKecerahan Warna Ikan Cupang (Betta splendens). J. Aquac. Fish. Health 2016, 6, 41. [Google Scholar]
- APHA. Standard Methods for the Examination of Water Wastewater, 21st ed.; APHA: Washington, DC, USA, 2005.
- Takahara, S.; Hamilton, H.B.; Neel, J.V.; Kobara, T.Y.; Ogura, Y.; Nishimura, E.T. Hypocatalasemia: A new genetic carrier state. J. Clin. Investig. 1960, 39, 610–619. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Trinder, P. Determination of blood glucose using 4-amino phenazone as oxygen acceptor. J. Clin. Pathol. 1969, 22, 246. [Google Scholar] [CrossRef]
- Roberts, B. Embryology of the Eye and its Adenxa; Kanger Publishers: Basel, Switzerland, 2001; p. 132. [Google Scholar]

{kind=link}
{kind=link}
| Parameters | Experimental Groups | ||||
|---|---|---|---|---|---|
| T1 (100 µg GnRH + 50 mg DOM) | T2 (50 µg GnRH + 25 mg DOM) | T3 (25 µg GnRH + 12.5 mg DOM) | T4 (12.5 µg GnRH + 6.25 mg DOM) | Control (0 µg GnRH + 0 mg DOM) | |
| GSI % (initial) | 0.4 ± 0.05 | 0.4 ± 0.05 | 0.4 ± 0.05 | 0.4 ± 0.05 | 0.4 ± 0.05 |
| GSI % (20th) | 7.72 ± 0.264 c | 6.78 ± 0.546 bc | 6.05 ± 0.226 b | 4.61 ± 0.205 a | 4.33 ± 0.0195 a |
| GSI % (40th) | 2.8 ± 0.741 a | 15.2 ± 0.638 c | 2.2 ± 0.864 a | 9.77 ± 0.159 b | 9.49 ± 0.059 b |
| Parameters | Experimental Groups | ||||
|---|---|---|---|---|---|
| T1 (100 µg GnRH + 50 mg DOM) | T2 (50 µg GnRH + 25 mg DOM) | T3 (25 µg GnRH + 12.5 mg DOM) | T4 (12.5 µg GnRH + 6.25 mg DOM) | Control (0 µg GnRH + 0 mg DOM) | |
| GSI % (initial) | 0.2 ± 0.05 | 0.2 ± 0.05 | 0.2 ± 0.05 | 0.2 ± 0.05 | 0.2 ± 0.05 |
| GSI % (20th) | 0.779 ± 0.00458 c | 0.837 ± 0.0405 c | 0.643 ± 0.0186 b | 0.643 ± 0.024 b | 0.623 ± 0.0231 b |
| GSI % (40th) | 2.62 ± 0.292 c | 2.7 ± 0.156 c | 3.16 ± 0.191 c | 1.19 ± 0.166 b | 1.25 ± 0.168 b |
| Parameters | Experimental Groups | ||||
|---|---|---|---|---|---|
| T1 (100 µg GnRH + 50 mg DOM) | T2 (50 µg GnRH + 25 mg DOM) | T3 (25 µg GnRH + 12.5 mg DOM) | T4 (12.5 µg GnRH + 6.25 mg DOM) | Control (0 µg GnRH + 0 mg DOM) | |
| Absolute fecundity | 1100 ± 36.3 b | 1300 ± 23 c | 1040 ± 27.5 b | 934 ± 39.1 a | 936 ± 25 a |
| Fertilization rate (%) | 75.7 ± 0.882 a | 76.3 ± 1.45 a | 74.3 ± 0.882 a | 85.3 ± 0.882 b | 86 ± 0.577 b |
| Hatching percentage (%) | 68.7 ± 2.4 b | 73.7 ± 1.45 c | 68.3 ± 1.76 b | 63.7 ± 0.882 ab | 62.7 ± 0.882 a |
| Survival (%) | 18.6 ± 0.176 a | 18.7 ± 0.0882 a | 18.6 ± 0.0577 a | 18.6 ± 0.115 a | 18.5 ± 0.115 a |
| Parameters | Experimental Groups | ||||
|---|---|---|---|---|---|
| T1 (100 µg GnRH + 50 mg DOM) | T2 (50 µg GnRH + 25 mg DOM) | T3 (25 µg GnRH + 12.5 mg DOM) | T4 (12.5 µg GnRH + 6.25 mg DOM) | Control (0 µg GnRH + 0 mg DOM) | |
| Sperm volume (µm3) | 15.2 ± 0.0577 b | 15.4 ± 0.0577 b | 15.6 ± 0.153 b | 7.37 ± 0.0882 a | 7.1 ± 0.115 a |
| Sperm concentration (107 cell/mL) | 3.76 ± 0.0636 bc | 4.08 ± 0.0917 cd | 4.34 ± 0.159 d | 3.47 ± 0.12 b | 2.45 ± 0.00882 a |
| Sperm motility (%) | 85.7 ± 0.882 a | 86 ± 0.577 a | 86 ± 0.577 a | 85.7 ± 0.882 a | 86 ± 0.577 a |
| Parameters | Experimental Groups | ||||
|---|---|---|---|---|---|
| T1 (100 µg GnRH + 50 mg DOM) | T2 (50 µg GnRH + 25 mg DOM) | T3 (25 µg GnRH + 12.5 mg DOM) | T4 (12.5 µg GnRH + 6.25 mg DOM) | Control (0 µg GnRH + 0 mg DOM) | |
| Liver cortisol (20th) | 1.55 ± 0.0186 d | 1.35 ± 0.0184 c | 1.32 ± 0.00318 c | 1.22 ± 0.0104 b | 1.15 ± 0.00231 a |
| Liver cortisol (40th) | 0.913 ± 0.0145 c | 0.913 ± 0.00882 c | 0.88 ± 0.00577 bc | 0.857 ± 0.0176 ab | 0.837 ± 0.00333 a |
| Muscle cortisol (20th) | 2.46 ± 0.414 c | 2.07 ± 0.0124 bc | 1.62 ± 0.0569 ab | 1.35 ± 0.0427 a | 1.28 ± 0.0388 a |
| Muscle cortisol (40th) | 0.973 ± 0.0145 d | 0.947 ± 0.012 cd | 0.927 ± 0.00882 bc | 0.893 ± 0.00882 ab | 0.867 ± 0.00882 a |
| Parameters | Experimental Groups | ||||
|---|---|---|---|---|---|
| T1 (100 µg GnRH + 50 mg DOM) | T2 (50 µg GnRH + 25 mg DOM) | T3 (25 µg GnRH + 12.5 mg DOM) | T4 (12.5 µg GnRH + 6.25 mg DOM) | Control (0 µg GnRH + 0 mg DOM) | |
| Liver glucose (20th day) | 522 ± 1.62 d | 489 ± 0.254 c | 434 ± 3.55 b | 429 ± 3.33 b | 357 ± 3.97 a |
| Liver glucose (40th day) | 365 ± 1.55 c | 362 ± 1.86 c | 357 ± 1.21 b | 355 ± 1.19 b | 345 ± 1.24 a |
| Muscle glucose (20th day) | 539 ± 10.2 c | 518 ± 10.8 c | 467 ± 2.79 b | 387 ± 4.1 a | 379 ± 7.86 a |
| Muscle glucose (40th day) | 393 ± 0.109 b | 391 ± 1.6 b | 389 ± 3.34 b | 382 ± 0.468 a | 377 ± 1.24 a |
| Parameters | Experimental Groups | |||||
|---|---|---|---|---|---|---|
| T1 (100 µg GnRH + 50 mg DOM) | T2 (50 µg GnRH + 25 mg DOM) | T3 (25 µg GnRH + 12.5 mg DOM) | T4 (12.5 µg GnRH + 6.25 mg DOM) | Control (0 µg GnRH + 0 mg DOM) | ||
| SOD (unit mg protein−1) | Brain | 35.2 ± 2.44 | 37.5 ± 2.57 | 37.9 ± 2.64 | 46.3 ± 2.15 | 35.7 ± 2.5 |
| Gill | 73.7 ± 1.83 b | 94.2 ± 2.45 c | 54.8 ± 2.38 a | 57.5 ± 2.51 a | 52.3 ± 2.06 a | |
| Liver | 24.6 ± 2.35 b | 34.4 ± 2.04 c | 12.5 ± 1.98 a | 17.6 ± 2.44 a | 12.4 ± 2.28 a | |
| Muscle | 41.6 ± 2.05 b | 44.6 ± 2.47 b | 40.6 ± 2.25 b | 30.6 ± 2.28 a | 29.7 ± 2.34 a | |
| Catalase (unit mg protein−1) | Brain | 184 ± 9.36 c | 281 ± 24.1 d | 145 ± 9.36 c | 92.5 ± 3.7 b | 40.1 ± 10 a |
| Gill | 134 ± 17.7 c | 281 ± 19.8 d | 67.2 ± 7.0 bc | 55.5 ± 12 bc | 50.1 ± 5.78 b | |
| Liver | 191 ± 6.13 c | 265 ± 3.96 d | 142 ± 3.54 b | 133 ± 6.41 b | 60.1 ± 10 a | |
| Muscle | 173 ± 3.54 c | 265 ± 14.3 d | 149 ± 6.13 c | 152 ± 3.7 c | 73.4 ± 8.83 b | |
| Parameters | Experimental Group | ||||
|---|---|---|---|---|---|
| T1 (100 µg GnRH + 50 mg DOM) | T2 (50 µg GnRH + 25 mg DOM) | T3 (25 µg GnRH + 12.5 mg DOM) | T4 (12.5 µg GnRH + 6.25 mg DOM) | Control (0 µg GnRH + 0 mg DOM) | |
| Testosterone (nmoL−1) | 16.1 ± 0.0897 c | 16.4 ± 0.255 cd | 16.6 ± 0.0176 d | 11.1 ± 0.103 b | 4.53 ± 0.0841 a |
| Vitellogenin (nmoL−1) | 183 ± 6.49 c | 189 ± 1.56 c | 144 ± 0.583 b | 138 ± 1.2 ab | 128 ± 14.81 a |
| Ingredients | Quantity (g) |
|---|---|
| Fish muscle | 75 |
| Corn flour | 30 |
| Salt | 2 |
| Lactogen | 5 |
| Gelling agent | 5 |
| Yeast | 2 |
| Parameters | Percentage | |
|---|---|---|
| Wet wt. Basis | Dry wt. Basis | |
| Moisture | 59.79% | --- |
| Protein | 22.63% | 56.27% |
| Lipid | 5% | 12.43% |
| Ash | 8.35% | 20.76% |
| NFE | 4.23% | 10.54% |
| Parameters | T1 | T2 | T3 | T4 | Control |
|---|---|---|---|---|---|
| pH | 6.9–7.16 | 6.8–7.15 | 6.8–7.12 | 6.5–7.09 | 6.5–7.15 |
| Temperature (°C) | 26–27.2 | 25.4–26.3 | 26.5–27.5 | 26.4–27.3 | 26.1–26.9 |
| Dissolved Oxygen (mg L−1) | 5.1–7.31 | 5.04–7.2 | 5.21–7.09 | 5.3–7.4 | 5.0–7.2 |
| Total Hardness (mg L−1) | 51.2–64.4 | 52.8–63.2 | 51.7–63.1 | 53.9–64.1 | 50.1–63.2 |
| Total Alkalinity (mg L−1) | 61–79.3 | 63.3–78.7 | 61.8–78.1 | 62.4–77.6 | 60.2–75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deb, S.; Biswas, P.; Singh, S.K.; Waikhom, G.; Debbarma, R.; Kashyap, S.; Lal, J.; Samal, K.; Malla, S.; Yadav, N.K.; et al. Oral Administration of GnRH and Domperidone via Gel Feed and Their Effect on Reproductive Performance of Devario devario (Bengal Danio), an Ornamental Fish. Gels 2025, 11, 554. https://doi.org/10.3390/gels11070554
Deb S, Biswas P, Singh SK, Waikhom G, Debbarma R, Kashyap S, Lal J, Samal K, Malla S, Yadav NK, et al. Oral Administration of GnRH and Domperidone via Gel Feed and Their Effect on Reproductive Performance of Devario devario (Bengal Danio), an Ornamental Fish. Gels. 2025; 11(7):554. https://doi.org/10.3390/gels11070554
Chicago/Turabian StyleDeb, Suparna, Pradyut Biswas, Soibam Khogen Singh, Gusheinzed Waikhom, Reshmi Debbarma, Shubham Kashyap, Jham Lal, Khusbu Samal, Supratim Malla, Nitesh Kumar Yadav, and et al. 2025. "Oral Administration of GnRH and Domperidone via Gel Feed and Their Effect on Reproductive Performance of Devario devario (Bengal Danio), an Ornamental Fish" Gels 11, no. 7: 554. https://doi.org/10.3390/gels11070554
APA StyleDeb, S., Biswas, P., Singh, S. K., Waikhom, G., Debbarma, R., Kashyap, S., Lal, J., Samal, K., Malla, S., Yadav, N. K., Devi, N. C., Das, P., Singh, N. S., Reddy, G. D., & Irungbam, S. (2025). Oral Administration of GnRH and Domperidone via Gel Feed and Their Effect on Reproductive Performance of Devario devario (Bengal Danio), an Ornamental Fish. Gels, 11(7), 554. https://doi.org/10.3390/gels11070554