
Bacterial Cellulose for Scalable and Sustainable Bio-Gels in the Circular Economy

 ,
,  , ,
, , 
Abstract
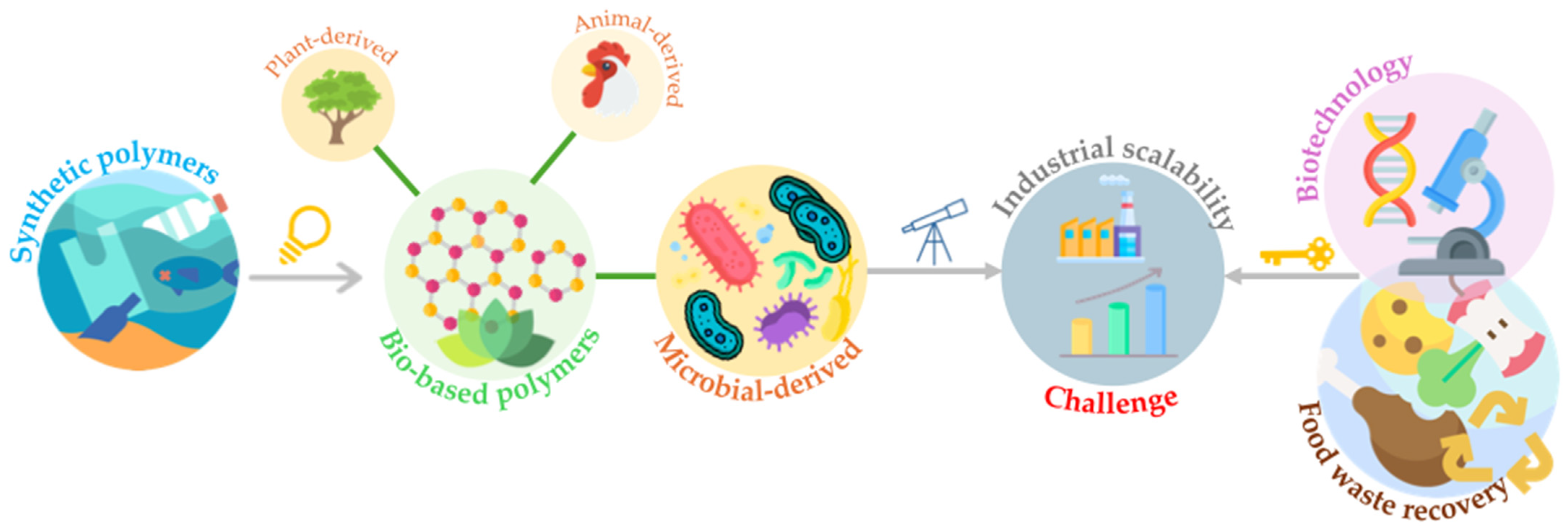
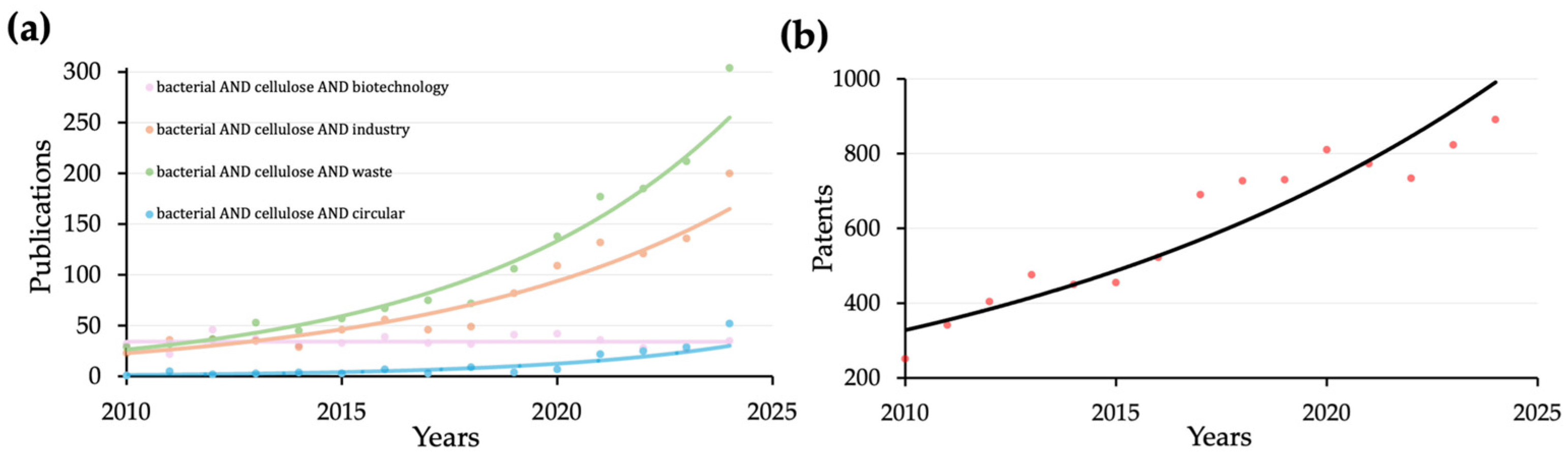
1. Introduction
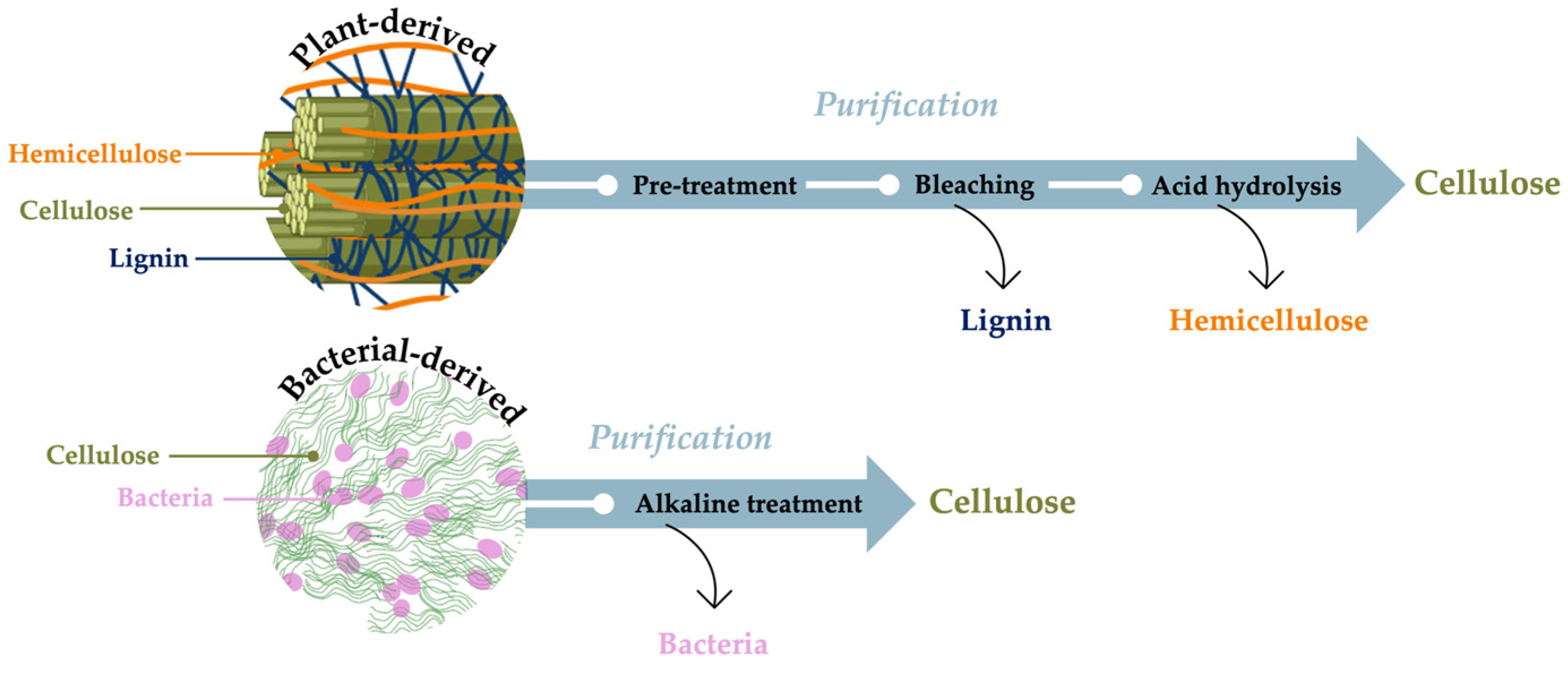
2. Bacterial-Derived Materials
3. Bacterial Cellulose: Features and Biosynthetic Pathway
4. Towards Industrial Production of Bacterial Cellulose
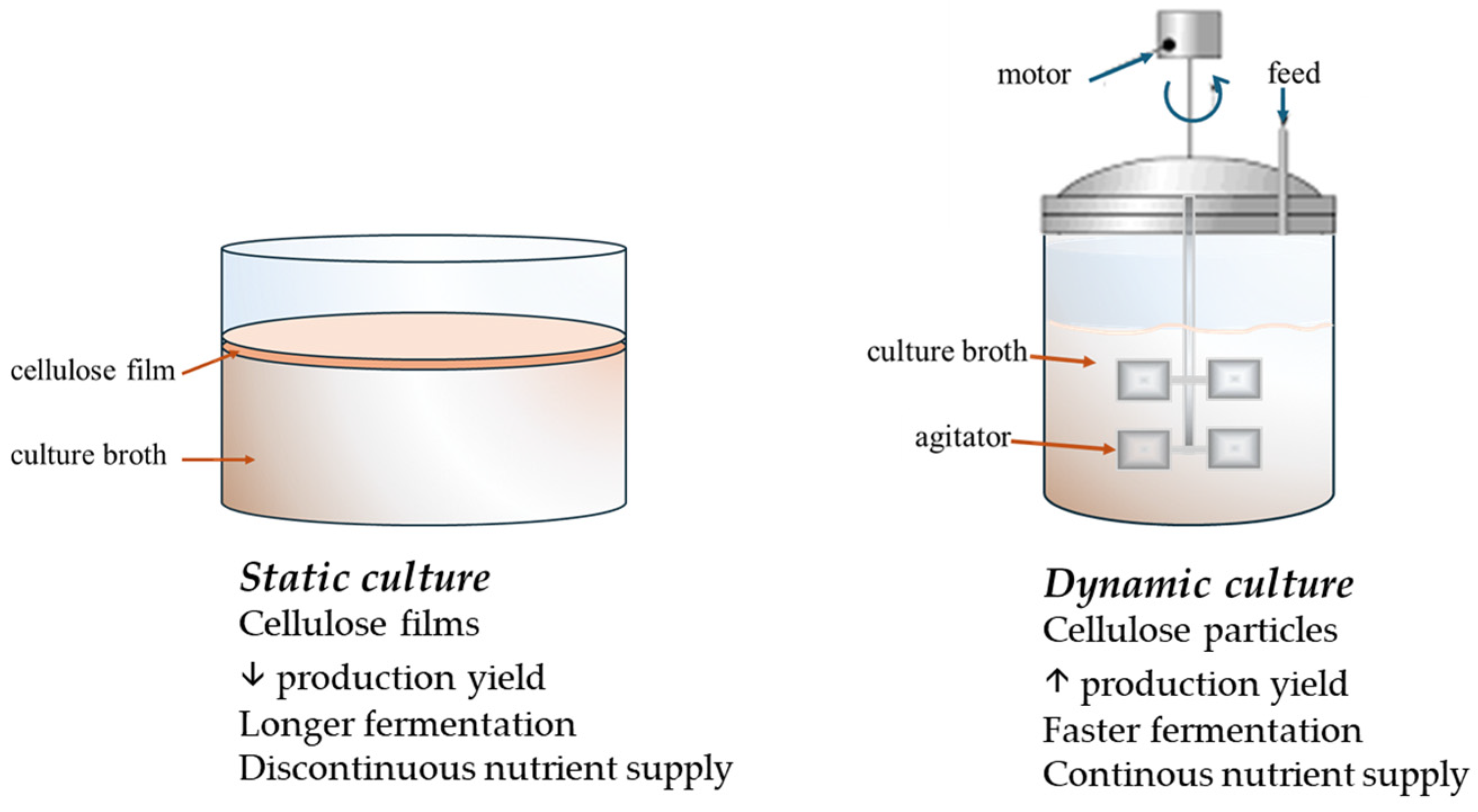
4.1. Biotechnological Process to Boost Bacterial Cellulose Production Yield
4.2. Biotechnological Approaches to Tailor Bacterial Cellulose Properties
5. Cost-Effective Production of Bacterial Cellulose
5.1. Nata De Coco
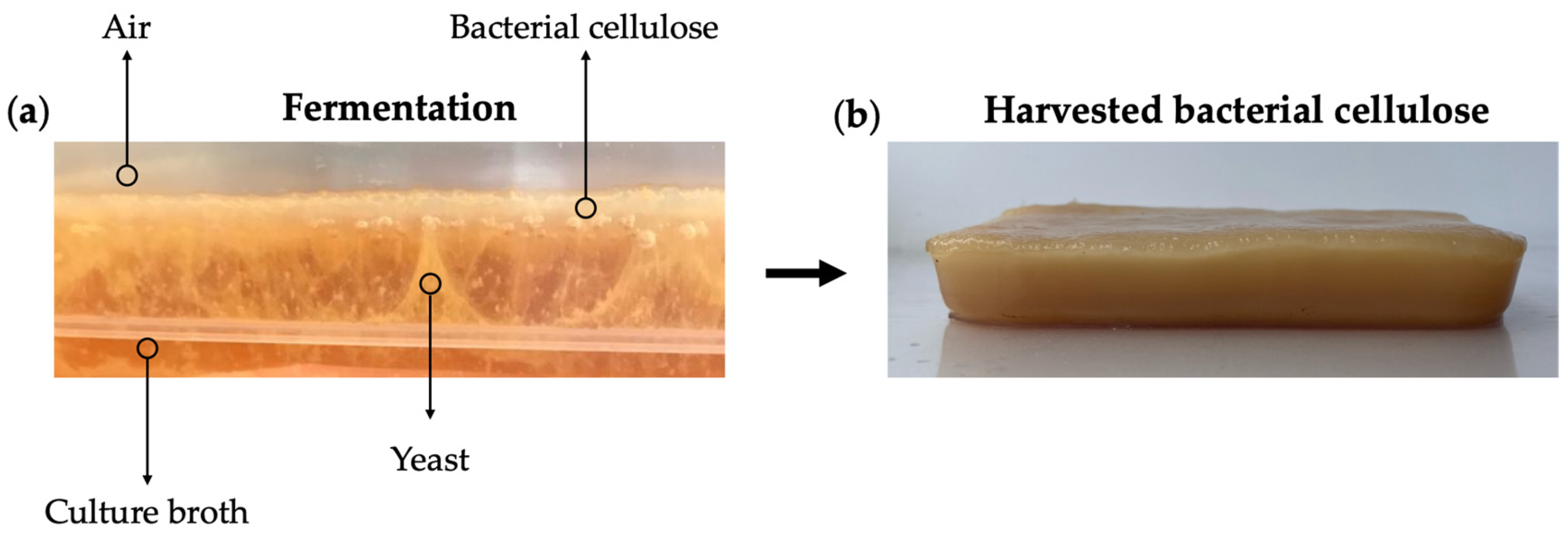
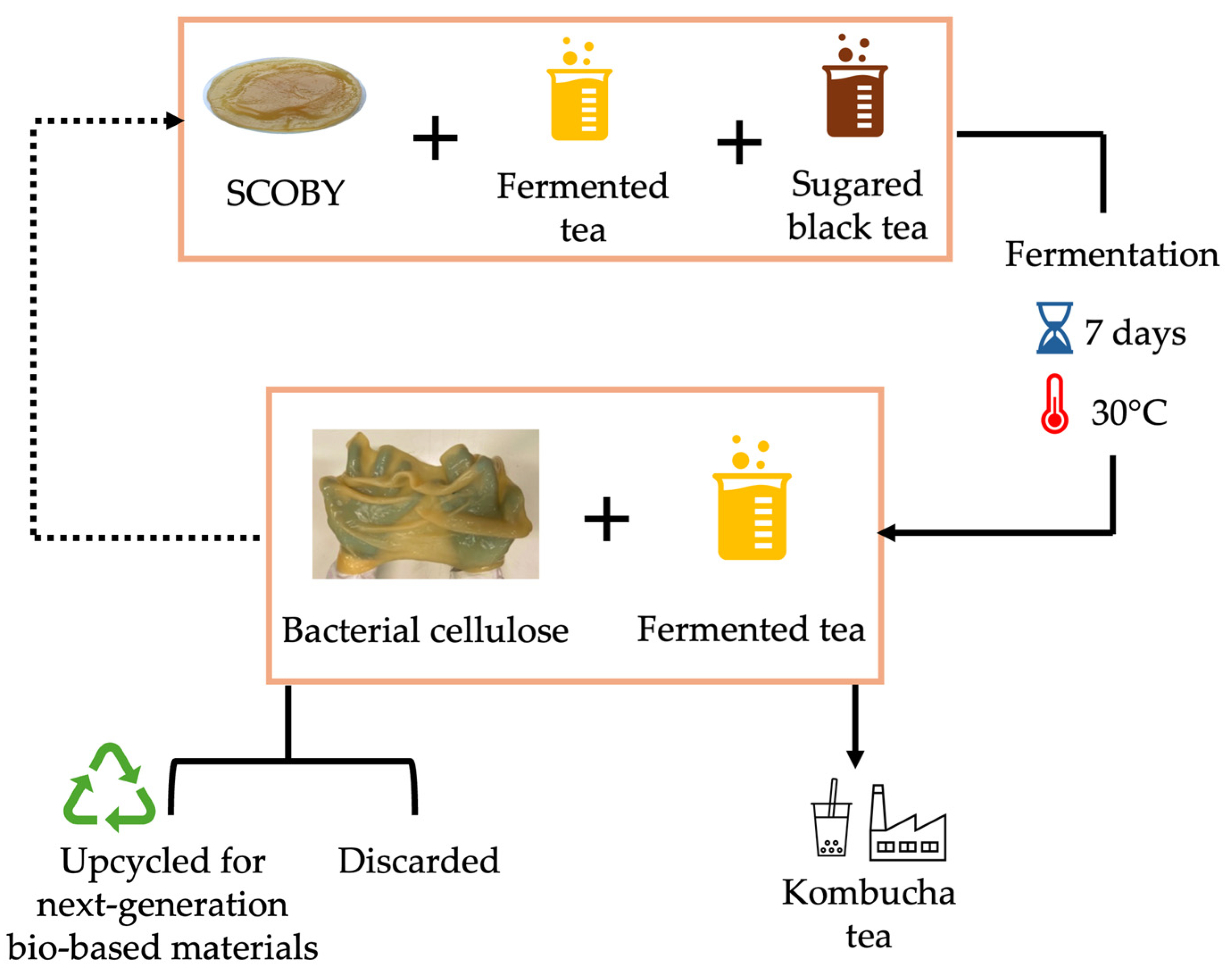
5.2. Kombucha Tea Fermentation
6. Circular Production of Bacterial Cellulose

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agri-Food Waste and Byproducts as Substrate | Application | Dry Production Yield | Yield Increase | Industrial Production | References |
|---|---|---|---|---|---|
| Wastewater from rice wine | - | 1.83 g/L | +26% | - | [150] |
| Potato peel | - | 4.7 g/L | +288% | Favorable results using potato peel waste for cost-effective industrial production | [151] |
| Banana leaves | Cardboard paper | 30 g/L (**) | - | Tunable properties for industrial needs | [135] |
| Coffee husk | - | 8.2 g/L | +446% | - | [144] |
| Wheat straw | - | - | - | [152] | |
| Pineapple | - | 3.24 g/L | +63% | - | [136] |
| Fruit juices | - | Up to 6 g/L | - | - | [137] |
| Rotten fruits and milk whey | Different envisioned applications | 60 mg/mL | +100% | - | [141] |
| Grape, cheese whey, and sulfite pulping liquor | - | Up to 2.5 g/L | - | Further characterization needed | [153] |
| Candied jujube | Further characterization needed | 2.25 g/L | - | Further characterization needed | [154] |
| Carob and Haricot bean | - | Up to 3.2 g/L | - | Optimized media for large-scale production | [142] |
| Maple syrup | Different possible applications | 1.51 g/L | - | - | [145] |
| Pineapple and sugarcane juices | - | 2.8 g/L | +30% | Low-cost substrate for large-scale industrial production. | [146] |
| Brewery waste | - | 5.05 g/L | - | - | [155] |
| Olive mill wastewater and cheese whey | - | - | - | Basis for industrial scale-up | [138] |
| Figs waste | - | 8.45 g/L | - | - | [15] |
| Bread waste | - | - | - | Low-cost substrate for large-scale industrial production. | [139] |
| Fruits peel | - | Up to 0.48 g/L | - | - | [140] |
| Pineapple peels and banana extracts | - | 2 g/L | 300% | Production suitable for future industrialization | [147] |
| Beet molasses, vinasse, and waste beer fermentation broth | - | Up to 5 g/L | - | - | [143] |
| Asparagus waste | Biomaterials | 2.57 g/L | - | - | [148] |
| Domestic food waste | Packaging and biomedical applications | 25 g/L | - | - | [115] |
| Waste apple pulp and stale bread | - | Up to 3.4 g/L | - | - | [149] |
7. Bacterial Cellulose for Industrial Applications
7.1. Biomedical Application
7.2. Sensors
7.3. Other Applications of Bacterial Cellulose
8. Bacterial Cellulose in the Fashion Sector
8.1. Environmental Stress Due to the Textile Sector
8.2. Environmental Stress Due to the Leather Industry
8.3. Bacterial Cellulose as a Textile and Leather Alternative
| BC Source | Alternative Material Features | Fashion Application | Scalability | Refs |
|---|---|---|---|---|
| - | Malleable, breathable, and water-impermeable BC-based nanocomposites impregnated with polydimethylsiloxane and perfluorocarbon | Textiles and shoes | Potential large-scale production | [193] |
| Kombucha tea fermentation | BC-derived hydrogel | Accessories and textile printings | Potential large-scale production | [190] |
| Kombucha tea fermentation | Dried BC sheets | Clothing material | - | [195] |
| Kombucha tea fermentation | BC-based composited with PVA, glycerol, PCL, and sunflower oil | Leather alternative | Scalable production | [187] |
| Kombucha tea fermentation | BC-polyurethane-polylactic acid composites | Textile, footwear, bags, and upholstery | - | [113] |
| Kombucha tea fermentation | BC-based materials enriched with soy and mushroom proteins | Leather alternative | - | [192] |
| Kombucha tea fermentation | Tailored-shaped BC | Textiles | - | [191] |
| Kombucha tea fermentation | BC-based composited with poly-vinyl-alcohol, glycerol, polycaprolactone, poly-lactic-acid | Textiles and shoes | - | [196] |
| Kombucha tea fermentation | BC-based composite with gold nanoparticles, silver nanoparticles, and graphene oxide | Leather alternative | - | [194] |
| Coconut water fermentation | Tanned BC sheets, oil-crosslinked and plasticized with glycerol | Leather alternative | To be improved to unlock industrial potential | [189] |
| Kombucha tea fermentation | Dried BC sheets dyed with coffee, ginger and sappan wood | - | Industrialization could be challenging | [127] |
| - | BC oil-plasticized | Leather alternative | - | [197] |
| - | Textile fibers produced with dry-jet BC | Textiles | Scaled process | [198] |
| Kombucha tea fermentation | Dried BC-sheets | Textiles | - | [199] |
| - | Melanated BC sheets | Textiles | Scalable BC | [200] |
9. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rocha, A.R.F.d.S.; Venturim, B.C.; Ellwanger, E.R.A.; Pagnan, C.S.; Silveira, W.B.d.; Martin, J.G.P. Bacterial Cellulose: Strategies for Its Production in the Context of Bioeconomy. J. Basic Microbiol. 2023, 63, 257–275. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.; Sicher, E.; Ayala-Garcia, C.; Sanchez-Fayos, I.M.; Conterno, L.; Ugur Yavuz, S. Innocell Bioreactor: An Open-Source Development to Produce Biomaterials for Food and Packaging Based on Fermentation Processes. Fermentation 2023, 9, 915. [Google Scholar] [CrossRef]
- Moreno-Díaz, C.; González-Arranz, S.; Martínez-Cerezo, C. Bacterial Cellulose Production within a Circular Economy Framework: Utilizing Organic Waste. Polymers 2024, 16, 2735. [Google Scholar] [CrossRef]
- Yermagambetova, A.; Tazhibayeva, S.; Takhistov, P.; Tyussyupova, B.; Tapia-Hernández, J.A.; Musabekov, K. Microbial Polysaccharides as Functional Components of Packaging and Drug Delivery Applications. Polymers 2024, 16, 2854. [Google Scholar] [CrossRef]
- Roh, E.K.; Oh, K.W.; Kim, S.H. Classification of Synthetic Polyurethane Leather by Mechanical Properties According to Consumers’ Preference for Fashion Items. Fibers Polym. 2013, 14, 1731–1738. [Google Scholar] [CrossRef]
- Wood, J.; Verran, J.; Redfern, J. Bacterial Cellulose Grown from Kombucha: Assessment of Textile Performance Properties Using Fashion Apparel Tests. Text. Res. J. 2023, 93, 3094–3108. [Google Scholar] [CrossRef]
- Rathinamoorthy, R.; Kiruba, T. Bacterial Cellulose—A Potential Material for Sustainable Eco-Friendly Fashion Products. J. Nat. Fibers 2022, 19, 3275–3287. [Google Scholar] [CrossRef]
- Grzybek, P.; Dudek, G.; van der Bruggen, B. Cellulose-Based Films and Membranes: A Comprehensive Review on Preparation and Applications. Chem. Eng. J. 2024, 495, 153500. [Google Scholar] [CrossRef]
- Taokaew, S. Bacterial Nanocellulose Produced by Cost-Effective and Sustainable Methods and Its Applications: A Review. Fermentation 2024, 10, 316. [Google Scholar] [CrossRef]
- Wang, B.; Qiu, S.; Chen, Z.; Hu, Y.; Shi, G.; Zhuo, H.; Zhang, H.; Zhong, L. Assembling Nanocelluloses into Fibrous Materials and Their Emerging Applications. Carbohydr. Polym. 2023, 299, 120008. [Google Scholar] [CrossRef]
- Walling, B.; Bharali, P.; Giridharan, B.; Gogoi, B.; Sorhie, V.; Alemtoshi; Mani, S.K. Bacterial Nanocellulose: A Novel Nanostructured Bio-Adsorbent for Green Remediation Technology. Acta Ecol. Sin. 2023, 43, 946–967. [Google Scholar]
- Wu, Z.; Chen, S.; Li, J.; Wang, B.; Jin, M.; Liang, Q.; Zhang, D.; Han, Z.; Deng, L.; Qu, X.; et al. Insights into Hierarchical Structure–Property–Application Relationships of Advanced Bacterial Cellulose Materials. Adv. Funct. Mater. 2023, 33, 2214327. [Google Scholar] [CrossRef]
- Cubas, A.L.V.; Provin, A.P.; Dutra, A.R.A.; Mouro, C.; Gouveia, I.C. Advances in the Production of Biomaterials through Kombucha Using Food Waste: Concepts, Challenges, and Potential. Polymers 2023, 15, 1701. [Google Scholar] [CrossRef]
- Pal, P.; Singh, A.K.; Srivastava, R.K.; Rathore, S.S.; Sahoo, U.K.; Subudhi, S.; Sarangi, P.K.; Prus, P. Circular Bioeconomy in Action: Transforming Food Wastes into Renewable Food Resources. Foods 2024, 13, 3007. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Goksungur, Y. Optimization of Bacterial Cellulose Production from Waste Figs by Komagataeibacter xylinus. Fermentation 2024, 10, 466. [Google Scholar] [CrossRef]
- Kumar Sarangi, P.; Subudhi, S.; Bhatia, L.; Saha, K.; Mudgil, D.; Prasad Shadangi, K.; Srivastava, R.K.; Pattnaik, B.; Arya, R.K. Utilization of Agricultural Waste Biomass and Recycling toward Circular Bioeconomy. Environ. Sci. Pollut. Res. 2023, 30, 8526–8539. [Google Scholar]
- Guleria, J.; Khan, M.A. Biomaterials and Biopolymers in Circular Economy: Latest Trends and Applications. In AIP Conference Proceedings; American Institute of Physics: College Park, MD, USA, 2024; Volume 2986. [Google Scholar]
- Patel, A.; Patel, P.; Shukla, A.; Wong, J.W.C.; Varjani, S.; Gosai, H. Sustainable Bioconversion of Industrial Wastes into Bacterial Cellulose for Diverse Applications: A Way Towards Pollution Control and Abatement. Curr. Pollut. Rep. 2023, 9, 226–242. [Google Scholar]
- El-Gendi, H.; Taha, T.H.; Ray, J.B.; Saleh, A.K. Recent Advances in Bacterial Cellulose: A Low-Cost Effective Production Media, Optimization Strategies and Applications. Cellulose 2022, 29, 7495–7533. [Google Scholar]
- Avcioglu, N.H. Bacterial Cellulose: Recent Progress in Production and Industrial Applications. World J. Microbiol. Biotechnol. 2022, 38, 86. [Google Scholar]
- Malcı, K.; Li, I.S.; Kisseroudis, N.; Ellis, T. Modulating Microbial Materials—Engineering Bacterial Cellulose with Synthetic Biology. ACS Synth. Biol. 2024, 13, 3857–3875. [Google Scholar] [CrossRef]
- Sumini, M.; Andrade, G.J.S.; Tischer, C.A.; Kobayashi, R.K.T.; Nakazato, G. Production of Bacterial Cellulose by Komagataeibacter xylinus: Biochemistry, Synthesis and Applications. Cellulose 2025, 32, 81–94. [Google Scholar] [CrossRef]
- Mcmeeking, A.; Dieckmann, E.; Cheeseman, C. Production Methods for Bacterial Biomaterials: A Review. Mater. Today Sustain. 2024, 25, 100623. [Google Scholar] [CrossRef]
- Wei, M.; Huang, Y.; Zhu, J.; Qiao, Y.; Xiao, N.; Jin, M.; Gao, H.; Huang, Y.; Hu, X.; Li, O. Advances in Hyaluronic Acid Production: Biosynthesis and Genetic Engineering Strategies Based on Streptococcus—A Review. Int. J. Biol. Macromol. 2024, 270, 132334. [Google Scholar]
- Ucm, R.; Aem, M.; Lhb, Z.; Kumar, V.; Taherzadeh, M.J.; Garlapati, V.K.; Chandel, A.K. Comprehensive Review on Biotechnological Production of Hyaluronic Acid: Status, Innovation, Market and Applications. Bioengineered 2022, 13, 9645–9661. [Google Scholar] [CrossRef] [PubMed]
- Ciriminna, R.; Scurria, A.; Pagliaro, M. Microbial Production of Hyaluronic Acid: The Case of an Emergent Technology in the Bioeconomy. Biofuels Bioprod. Biorefining 2021, 15, 1604–1610. [Google Scholar] [CrossRef]
- Serra, M.; Casas, A.; Toubarro, D.; Barros, A.N.; Teixeira, J.A. Microbial Hyaluronic Acid Production: A Review. Molecules 2023, 28, 2084. [Google Scholar] [CrossRef]
- Graciela, C.Q.; José Juan, E.C.; Gieraldin, C.L.; Xóchitl Alejandra, P.M.; Gabriel, A.Á. Hyaluronic Acid—Extraction Methods, Sources and Applications. Polymers 2023, 15, 3473. [Google Scholar] [CrossRef]
- Cerminati, S.; Leroux, M.; Anselmi, P.; Peirú, S.; Alonso, J.C.; Priem, B.; Menzella, H.G. Low Cost and Sustainable Hyaluronic Acid Production in a Manufacturing Platform Based on Bacillus subtilis 3NA Strain. Appl. Microbiol. Biotechnol. 2021, 105, 3075–3086. [Google Scholar] [CrossRef]
- Amado, I.R.; Vázquez, J.A.; Pastrana, L.; Teixeira, J.A. Microbial Production of Hyaluronic Acid from Agro-Industrial by-Products: Molasses and Corn Steep Liquor. Biochem. Eng. J. 2017, 117, 181–187. [Google Scholar] [CrossRef]
- Amado, I.R.; Vázquez, J.A.; Pastrana, L.; Teixeira, J.A. Cheese Whey: A Cost-Effective Alternative for Hyaluronic Acid Production by Streptococcus zooepidemicus. Food Chem. 2016, 198, 54–61. [Google Scholar] [CrossRef]
- Sun, H.; Ni, J.; Yang, G.; Liu, Z.; Wang, Z.; Zhu, S.; Li, Z.; Jiang, Y.; Zhan, X.; Wang, Y.; et al. A Novel Coupled Fermentation System for Low-Molecular-Weight Xanthan Gum with Diverse Biological Activities. Int. J. Biol. Macromol. 2024, 279, 135283. [Google Scholar] [CrossRef]
- Bhat, I.M.; Wani, S.M.; Mir, S.A.; Masoodi, F.A. Advances in Xanthan Gum Production, Modifications and Its Applications. Biocatal. Agric. Biotechnol. 2022, 42, 102328. [Google Scholar]
- Dos Santos, F.P.; Oliveira, A.M.; Nunesa, T.P.; De Farias Silva, C.E.; De Souza Abud, A.K. Bioconversion of Agro-Industrial Wastes into Xanthan Gum. Chem. Eng. Trans. 2016, 49, 145–150. [Google Scholar] [CrossRef]
- Bhat, R. Fermentation of Black Tea Broth (Kombucha): I. Effects of Sucrose Concentration and Fermentation Time on the Yield of Microbial Cellulose. Int. Food Res. J. 2012, 19, 109–117. [Google Scholar]
- Girard, V.D.; Chaussé, J.; Vermette, P. Bacterial Cellulose: A Comprehensive Review. J. Appl. Polym. Sci. 2024, 141, e55163. [Google Scholar]
- Yuan, Z.; Yin, H.; Zheng, M.; Chen, X.; Peng, W.; Zhou, H.; Xing, J.; Wang, L.; Hu, S. Biodegradable, Robust, and Conductive Bacterial Cellulose @PPy-P Macrofibers as Resistive Strain Sensors for Smart Textiles. Carbohydr. Polym. 2025, 349, 122963. [Google Scholar] [CrossRef]
- Dáger-López, D.; Chenché, Ó.; Ricaurte-Párraga, R.; Núñez-Rodríguez, P.; Bajaña, J.M.; Fiallos-Cárdenas, M. Advances in the Production of Sustainable Bacterial Nanocellulose from Banana Leaves. Polymers 2024, 16, 1157. [Google Scholar] [CrossRef]
- Pandey, A.; Singh, M.K.; Singh, A. Bacterial Cellulose: A Smart Biomaterial for Biomedical Applications. J. Mater. Res. 2024, 39, 2–18. [Google Scholar]
- Núñez, D.; Oyarzún, P.; González, S.; Martínez, I. Toward Biomanufacturing of Next-Generation Bacterial Nanocellulose (BNC)-Based Materials with Tailored Properties: A Review on Genetic Engineering Approaches. Biotechnol. Adv. 2024, 74, 108390. [Google Scholar]
- Shen, L.; Worrell, E.; Patel, M.K. Environmental Impact Assessment of Man-Made Cellulose Fibres. Resour. Conserv. Recycl. 2010, 55, 260–274. [Google Scholar] [CrossRef]
- Moradali, M.F.; Rehm, B.H.A. Bacterial Biopolymers: From Pathogenesis to Advanced Materials. Nat. Rev. Microbiol. 2020, 18, 195–210. [Google Scholar] [CrossRef]
- Zhong, C. Industrial-Scale Production and Applications of Bacterial Cellulose. Front. Bioeng. Biotechnol. 2020, 8, 605374. [Google Scholar] [CrossRef]
- Gupte, Y.; Kulkarni, A.; Raut, B.; Sarkar, P.; Choudhury, R.; Chawande, A.; Kumar, G.R.K.; Bhadra, B.; Satapathy, A.; Das, G.; et al. Characterization of Nanocellulose Production by Strains of Komagataeibacter sp. Isolated from Organic Waste and Kombucha. Carbohydr. Polym. 2021, 266, 118176. [Google Scholar] [CrossRef] [PubMed]
- Manan, S.; Ullah, M.W.; Ul-Islam, M.; Shi, Z.; Gauthier, M.; Yang, G. Bacterial Cellulose: Molecular Regulation of Biosynthesis, Supramolecular Assembly, and Tailored Structural and Functional Properties. Prog. Mater. Sci. 2022, 129, 100972. [Google Scholar] [CrossRef]
- Noack, L.C.; Persson, S. Cellulose Synthesis across Kingdoms. Curr. Biol. 2023, 33, R251–R254. [Google Scholar] [CrossRef] [PubMed]
- McManus, J.B.; Yang, H.; Wilson, L.; Kubicki, J.D.; Tien, M. Initiation, Elongation, and Termination of Bacterial Cellulose Synthesis. ACS Omega 2018, 3, 2690–2698. [Google Scholar] [CrossRef]
- Omadjela, O.; Narahari, A.; Strumillo, J.; Mélida, H.; Mazur, O.; Bulone, V.; Zimmer, J. BcsA and BcsB Form the Catalytically Active Core of Bacterial Cellulose Synthase Sufficient for in Vitro Cellulose Synthesis. Proc. Natl. Acad. Sci. USA 2013, 110, 17856–17861. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.-Q.; Gao, Y.-G.; Tajima, K.; Sunagawa, N.; Zhou, Y.; Kawano, S.; Fujiwara, T.; Yoda, T.; Shimura, D.; Satoh, Y.; et al. Structure of Bacterial Cellulose Synthase Subunit D Octamer with Four Inner Passageways. Proc. Natl. Acad. Sci. USA 2010, 107, 17957–17961. [Google Scholar] [CrossRef]
- Kawano, S.; Tajima, K.; Kono, H.; Numata, Y.; Yamashita, H.; Satoh, Y.; Munekata, M. Regulation of Endoglucanase Gene (Cmcax) Expression in Acetobacter xylinum. J. Biosci. Bioeng. 2008, 106, 88–94. [Google Scholar] [CrossRef]
- Güzel, M. Characterization of Cellulose Produced by Bacteria Isolated from Different Vinegars. Int. J. Biol. Macromol. 2024, 277, 134436. [Google Scholar] [CrossRef]
- Gullo, M.; La China, S.; Falcone, P.M.; Giudici, P. Biotechnological Production of Cellulose by Acetic Acid Bacteria: Current State and Perspectives. Appl. Microbiol. Biotechnol. 2018, 102, 6885–6898. [Google Scholar] [CrossRef]
- Lu, Y.; Mehling, M.; Huan, S.; Bai, L.; Rojas, O.J. Biofabrication with Microbial Cellulose: From Bioadaptive Designs to Living Materials. Chem. Soc. Rev. 2024, 53, 7363–7391. [Google Scholar]
- Avcioglu, N.H.; Birben, M.; Seyis Bilkay, I. Optimization and Physicochemical Characterization of Enhanced Microbial Cellulose Production with a New Kombucha Consortium. Process Biochem. 2021, 108, 60–68. [Google Scholar] [CrossRef]
- Choi, S.M.; Rao, K.M.; Zo, S.M.; Shin, E.J.; Han, S.S. Bacterial Cellulose and Its Applications. Polymers 2022, 14, 1080. [Google Scholar] [CrossRef]
- Wang, J.; Li, C.; Tang, Y. Constructing Bacterial Cellulose and Its Composites: Regulating Treatments towards Applications. Cellulose 2024, 31, 7793–7817. [Google Scholar]
- Rosson, L.; Tan, B.; Best, W.; Byrne, N. Applications of Regenerated Bacterial Cellulose: A Review. Cellulose 2024, 31, 10165–10190. [Google Scholar]
- Campano, C.; Balea, A.; Blanco, A.; Negro, C. Enhancement of the Fermentation Process and Properties of Bacterial Cellulose: A Review. Cellulose 2016, 23, 57–91. [Google Scholar]
- Raut, M.P.; Asare, E.; Syed Mohamed, S.M.D.; Amadi, E.N.; Roy, I. Bacterial Cellulose-Based Blends and Composites: Versatile Biomaterials for Tissue Engineering Applications. Int. J. Mol. Sci. 2023, 24, 986. [Google Scholar] [CrossRef]
- Emenike, E.C.; Iwuozor, K.O.; Saliu, O.D.; Ramontja, J.; Adeniyi, A.G. Advances in the Extraction, Classification, Modification, Emerging and Advanced Applications of Crystalline Cellulose: A Review. Carbohydr. Polym. Technol. Appl. 2023, 6, 100337. [Google Scholar]
- Kunz, A.; Swine, E.; Mujtaba, M.; Michelin, M.; Farris, S. Extraction of High-Quality Grade Cellulose and Cellulose Nanocrystals from Different Lignocellulosic Agri-Food Wastes. Front. Sustain. Food Syst. 2023, 6, 1087867. [Google Scholar]
- Manimaran, M.; Norizan, M.N.; Kassim, M.H.M.; Adam, M.R.; Norrrahim, M.N.F.; Knight, V.F. Critical Assessment of the Thermal Stability and Degradation of Chemically Functionalized Nanocellulose-Based Polymer Nanocomposites. Nanotechnol. Rev. 2024, 13, 20240005. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Tseng, Y.-S.; Kumar, V.; Chen, C.-W.; Haldar, D.; Saini, J.K.; Dong, C.-D. Developments in Bioprocess for Bacterial Cellulose Production. Bioresour. Technol. 2022, 344, 126343. [Google Scholar]
- Infante-Neta, A.A.; D’Almeida, A.P.; de Albuquerque, T.L. Bacterial Cellulose in Food Packaging: A Bibliometric Analysis and Review of Sustainable Innovations and Prospects. Processes 2024, 12, 1975. [Google Scholar] [CrossRef]
- Navya, P.V.; Gayathri, V.; Samanta, D.; Sampath, S. Bacterial Cellulose: A Promising Biopolymer with Interesting Properties and Applications. Int. J. Biol. Macromol. 2022, 220, 435–461. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Mathur, G. Bacterial Cellulose: A Multipurpose Biomaterial for Manmade World. Curr. Appl. Sci. Technol. 2023, 23, 10–55003. [Google Scholar] [CrossRef]
- Cannazza, P.; Rissanen, A.J.; Guizelini, D.; Losoi, P.; Sarlin, E.; Romano, D.; Santala, V.; Mangayil, R. Characterization of Komagataeibacter Isolate Reveals New Prospects in Waste Stream Valorization for Bacterial Cellulose Production. Microorganisms 2021, 9, 2230. [Google Scholar] [CrossRef]
- Brugnoli, M.; Robotti, F.; La China, S.; Anguluri, K.; Haghighi, H.; Bottan, S.; Ferrari, A.; Gullo, M. Assessing Effectiveness of Komagataeibacter Strains for Producing Surface-Microstructured Cellulose via Guided Assembly-Based Biolithography. Sci. Rep. 2021, 11, 19311. [Google Scholar] [CrossRef]
- Sathianathan, N.; Karri, R.R.; Gunavijayan, G.; Raj, A.; Mubarak, N.M.; Latha, V.; Mukundan, A.V.; Krishnankutty, R.E.; Chellappanpillai, S. Optimisation of Bacterial Cellulose Production by Novacetimonas hansenii Isolated from Pomegranate Fruit Wastes. J. Mol. Liq. 2025, 422, 126912. [Google Scholar] [CrossRef]
- Lasagni, F.; Cassanelli, S.; Gullo, M. How Carbon Sources Drive Cellulose Synthesis in Two Komagataeibacter xylinus Strains. Sci. Rep. 2024, 14, 20494. [Google Scholar] [CrossRef]
- Barshan, S.; Rezazadeh-Bari, M.; Almasi, H.; Amiri, S. Optimization and Characterization of Bacterial Cellulose Produced by Komagatacibacter xylinus PTCC 1734 Using Vinasse as a Cheap Cultivation Medium. Int. J. Biol. Macromol. 2019, 136, 1188–1195. [Google Scholar] [CrossRef]
- Li, Z.-Y.; Azi, F.; Ge, Z.-W.; Liu, Y.-F.; Yin, X.-T.; Dong, M.-S. Bio-Conversion of Kitchen Waste into Bacterial Cellulose Using a New Multiple Carbon Utilizing Komagataeibacter rhaeticus: Fermentation Profiles and Genome-Wide Analysis. Int. J. Biol. Macromol. 2021, 191, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Uğurel, C.; Öğüt, H. Optimization of Bacterial Cellulose Production by Komagataeibacter rhaeticus K23. Fibers 2024, 12, 29. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, C.; Yang, J.; Sun, B.; Lin, J.; Sun, D. Effect of Culture Conditions on Cellulose Production by a Komagataeibacter xylinus Strain. Macromol. Biosci. 2022, 22, 2100476. [Google Scholar] [CrossRef]
- Li, W.; Huang, X.; Liu, H.; Lian, H.; Xu, B.; Zhang, W.; Sun, X.; Wang, W.; Jia, S.; Zhong, C. Improvement in Bacterial Cellulose Production by Co-Culturing Bacillus Cereus and Komagataeibacter xylinus. Carbohydr. Polym. 2023, 313, 120892. [Google Scholar] [CrossRef]
- Akki, A.J.; Hiremath, L.D.; Badkillaya, R.R. Harnessing Symbiotic Association of Lactic Acid Bacteria and Cellulose-Synthesizing Bacteria for Enhanced Biological Activity. Iran. J. Sci. 2024, 48, 311–320. [Google Scholar] [CrossRef]
- Hu, H.; Catchmark, J.M.; Demirci, A. Effects of Pullulan Additive and Co-Culture of Aureobasidium pullulans on Bacterial Cellulose Produced by Komagataeibacter hansenii. Bioprocess Biosyst. Eng. 2022, 45, 573–587. [Google Scholar] [CrossRef]
- Teh, M.Y.; Ooi, K.H.; Danny Teo, S.X.; Bin Mansoor, M.E.; Shaun Lim, W.Z.; Tan, M.H. An Expanded Synthetic Biology Toolkit for Gene Expression Control in Acetobacteraceae. ACS Synth. Biol. 2019, 8, 708–723. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, X.; Chen, X.; Yuan, F.; Sun, B.; Xu, Y.; Yang, J.; Sun, D. Complete Genome Sequence of the Cellulose-Producing Strain Komagataeibacter nataicola RZS01. Sci. Rep. 2017, 7, 4431. [Google Scholar] [CrossRef]
- Florea, M.; Hagemann, H.; Santosa, G.; Abbott, J.; Micklem, C.N.; Spencer-Milnes, X.; de Arroyo Garcia, L.; Paschou, D.; Lazenbatt, C.; Kong, D.; et al. Engineering Control of Bacterial Cellulose Production Using a Genetic Toolkit and a New Cellulose-Producing Strain. Proc. Natl. Acad. Sci. USA 2016, 113, E3431–E3440. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Zhao, X.Q.; Li, D.M.; Wu, Y.M.; Wahid, F.; Xie, Y.Y.; Zhong, C. Review on the Strategies for Enhancing Mechanical Properties of Bacterial Cellulose. J. Mater. Sci. 2023, 58, 15265–15293. [Google Scholar] [CrossRef]
- Montenegro-Silva, P.; Ellis, T.; Dourado, F.; Gama, M.; Domingues, L. Enhanced Bacterial Cellulose Production in Komagataeibacter sucrofermentans: Impact of Different PQQ-Dependent Dehydrogenase Knockouts and Ethanol Supplementation. Biotechnol. Biofuels Bioprod. 2024, 17, 35. [Google Scholar] [CrossRef]
- Sajadi, E.; Fatemi, S.S.-A.; Babaeipour, V.; Deldar, A.A.; Yakhchali, B.; Anvar, M.S. Increased Cellulose Production by Heterologous Expression of BcsA and B Genes from Gluconacetobacterxylinus in E. Coli Nissle 1917. Bioprocess Biosyst. Eng. 2019, 42, 2023–2034. [Google Scholar] [CrossRef]
- Liu, D.; Cao, Y.; Qu, R.; Gao, G.; Chen, S.; Zhang, Y.; Wu, M.; Ma, T.; Li, G. Production of Bacterial Cellulose Hydrogels with Tailored Crystallinity from Enterobacter sp. FY-07 by the Controlled Expression of Colanic Acid Synthetic Genes. Carbohydr. Polym. 2019, 207, 563–570. [Google Scholar] [CrossRef]
- Andrade, F.K.; Costa, R.; Domingues, L.; Soares, R.; Gama, M. Improving Bacterial Cellulose for Blood Vessel Replacement: Functionalization with a Chimeric Protein Containing a Cellulose-Binding Module and an Adhesion Peptide. Acta Biomater. 2010, 6, 4034–4041. [Google Scholar] [CrossRef] [PubMed]
- Hur, D.H.; Choi, W.S.; Kim, T.Y.; Lee, S.Y.; Park, J.H.; Jeong, K.J. Enhanced Production of Bacterial Cellulose in Komagataeibacter xylinus Via Tuning of Biosynthesis Genes with Synthetic RBS. J. Microbiol. Biotechnol. 2020, 30, 1430–1435. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Lv, Z.; Liu, J.; Wang, Y.; Zhang, T.; Xie, Y.; Jia, S.; Xin, B.; Zhong, C. Engineering PTS-Based Glucose Metabolism for Efficient Biosynthesis of Bacterial Cellulose by Komagataeibacter xylinus. Carbohydr. Polym. 2024, 343, 122459. [Google Scholar] [CrossRef]
- Yang, L.; Zhu, X.; Chen, Y.; Jun, W. Enhanced Bacterial Cellulose Production in Gluconacetobacter xylinus by Overexpression of Two Genes (BscC and BcsD) and a Modified Static Culture. Int. J. Biol. Macromol. 2024, 260, 129552. [Google Scholar] [CrossRef]
- Huang, L.-H.; Li, X.-J.; Wang, Y.-T.; Jia, S.-R.; Xin, B.; Zhong, C. Enhancing Bacterial Cellulose Production with Hypoxia-Inducible Factors. Appl. Microbiol. Biotechnol. 2022, 106, 7099–7112. [Google Scholar] [CrossRef]
- Yang, F.; Cao, Z.; Li, C.; Chen, L.; Wu, G.; Zhou, X.; Hong, F.F. A Recombinant Strain of Komagataeibacter xylinus ATCC 23770 for Production of Bacterial Cellulose from Mannose-Rich Resources. New Biotechnol. 2023, 76, 72–81. [Google Scholar] [CrossRef]
- Jacek, P.; Szustak, M.; Kubiak, K.; Gendaszewska-Darmach, E.; Ludwicka, K.; Bielecki, S. Scaffolds for Chondrogenic Cells Cultivation Prepared from Bacterial Cellulose with Relaxed Fibers Structure Induced Genetically. Nanomaterials 2018, 8, 1066. [Google Scholar] [CrossRef]
- Jacek, P.; Kubiak, K.; Ryngajłło, M.; Rytczak, P.; Paluch, P.; Bielecki, S. Modification of Bacterial Nanocellulose Properties through Mutation of Motility Related Genes in Komagataeibacter hansenii ATCC 53582. New Biotechnol. 2019, 52, 60–68. [Google Scholar] [CrossRef]
- Huang, L.; Liu, Q.; Sun, X.; Li, X.; Liu, M.; Jia, S.; Xie, Y.; Zhong, C. Tailoring Bacterial Cellulose Structure through CRISPR Interference-mediated Downregulation of GalU in Komagataeibacter xylinus CGMCC 2955. Biotechnol. Bioeng. 2020, 117, 2165–2176. [Google Scholar] [CrossRef] [PubMed]
- Takahama, R.; Kato, H.; Tajima, K.; Tagawa, S.; Kondo, T. Biofabrication of a Hyaluronan/Bacterial Cellulose Composite Nanofibril by Secretion from Engineered Gluconacetobacter. Biomacromolecules 2021, 22, 4709–4719. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Catchmark, J.M. Bacterial Cellulose/Hyaluronic Acid Nanocomposites Production through Co-Culturing Gluconacetobacter hansenii and Lactococcus Lactis under Different Initial PH Values of Fermentation Media. Cellulose 2020, 27, 2529–2540. [Google Scholar] [CrossRef]
- Fang, J.; Kawano, S.; Tajima, K.; Kondo, T. In Vivo Curdlan/Cellulose Bionanocomposite Synthesis by Genetically Modified Gluconacetobacter xylinus. Biomacromolecules 2015, 16, 3154–3160. [Google Scholar] [CrossRef]
- Nie, W.; He, Z.; Gu, M.; Zhou, T.; Xu, J.; Zhong, J.; Yang, Y.; Zhong, W. Improved Bacterial Cellulose Production by Acetobacter Oryzoeni MGC-N8819 in Tobacco Waste Extract Coupled with Nicotine Removal by Pseudomonas sp. JY-Q/5∆. Int. J. Biol. Macromol. 2025, 293, 139336. [Google Scholar] [CrossRef]
- Gilbert, C.; Tang, T.-C.; Ott, W.; Dorr, B.A.; Shaw, W.M.; Sun, G.L.; Lu, T.K.; Ellis, T. Living Materials with Programmable Functionalities Grown from Engineered Microbial Co-Cultures. Nat. Mater. 2021, 20, 691–700. [Google Scholar] [CrossRef]
- Li, S.; Liu, G. Harnessing Cellulose-Binding Protein Domains for the Development of Functionalized Cellulose Materials. Bioresour. Bioprocess. 2024, 11, 74. [Google Scholar] [CrossRef]
- Lahiri, D.; Nag, M.; Dutta, B.; Dey, A.; Sarkar, T.; Pati, S.; Edinur, H.A.; Kari, Z.A.; Noor, N.H.M.; Ray, R.R. Bacterial Cellulose: Production, Characterization and Application as Antimicrobial Agent. Int. J. Mol. Sci. 2021, 22, 12984. [Google Scholar] [CrossRef]
- Costa, A.F.S.; Almeida, F.C.G.; Vinhas, G.M.; Sarubbo, L.A. Production of Bacterial Cellulose by Gluconacetobacter hansenii Using Corn Steep Liquor as Nutrient Sources. Front. Microbiol. 2017, 8, 2027. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, Q.; Wang, Y.; Liu, J.; Zhu, P. Preparation and Characterization of Bacterial Cellulose by Kombucha Using Corncob. Cellulose 2024, 31, 6625–6642. [Google Scholar] [CrossRef]
- Behera, B.; Laavanya, D.; Balasubramanian, P. Techno-Economic Feasibility Assessment of Bacterial Cellulose Biofilm Production during the Kombucha Fermentation Process. Bioresour. Technol. 2022, 346, 126659. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Kang, J.; Fei, S.; Qin, X.; Lin, X.; Wang, X.; Zhao, B.; He, C.; Liu, S.; Li, C. Lactiplantibacillus plantarum Causes the Abnormal Fermentation of Bacterial Cellulose by Komagataeibacter nataicola during nata de coco Production. Food Biosci. 2024, 61, 104603. [Google Scholar] [CrossRef]
- Qin, X.; Yuan, Y.; Fei, S.; Lin, X.; Shi, S.; Wang, X.; Pang, Q.; Kang, J.; Li, C.; Liu, S. Exploring the Biotic and Abiotic Drivers Influencing nata de coco Production by Komagataeibacter nataicola in Pre-Fermented Coconut Water. Int. J. Food Microbiol. 2024, 414, 110620. [Google Scholar] [CrossRef]
- Fei, S.; Yang, X.; Xu, W.; Zhang, J.; Li, J.; Chen, H.; Lin, X.; Liu, S.; Li, C. Insights into Proteomics Reveal Mechanisms of Ethanol-Enhanced Bacterial Cellulose Biosynthesis by Komagataeibacter nataicola. Fermentation 2023, 9, 575. [Google Scholar] [CrossRef]
- Fei, S.; Wang, X.; Qin, X.; Yuan, Y.; Zheng, Y.; Lin, X.; Kang, J.; Liu, S.; Li, C. Efficient nata de coco Production of Komagataeibacter nataicola Driven by Microbiota-Fermented Coconut Water: Biological and Structural Characteristics. Food Biosci. 2024, 62, 105184. [Google Scholar] [CrossRef]
- Rahmayanti, H.D.; Ginting, J.C.; Kartika, T.R.; Ardiani, S.; Akmalia, N.; Zulfi, A. A Study of Optical Properties of Edible Film Based Coconut Water for Sustainable and Environmentally Friendly Materials Packaging. IOP Conf. Ser. Earth Environ. Sci. 2024, 1359, 012109. [Google Scholar] [CrossRef]
- Nyhan, L.M.; Lynch, K.M.; Sahin, A.W.; Arendt, E.K. Advances in Kombucha Tea Fermentation: A Review. Appl. Microbiol. 2022, 2, 73–103. [Google Scholar] [CrossRef]
- Bergottini, V.M.; Bernhardt, D. Bacterial Cellulose Aerogel Enriched in Nanofibers Obtained from Kombucha SCOBY Byproduct. Mater. Today Commun. 2023, 35, 105975. [Google Scholar] [CrossRef]
- Laavanya, D.; Shirkole, S.; Balasubramanian, P. Current Challenges, Applications and Future Perspectives of SCOBY Cellulose of Kombucha Fermentation. J. Clean. Prod. 2021, 295, 126454. [Google Scholar] [CrossRef]
- Tapias, Y.A.R.; Di Monte, M.V.; Peltzer, M.A.; Salvay, A.G. Kombucha Fermentation in Yerba Mate: Cellulose Production, Films Formulation and Its Characterisation. Carbohydr. Polym. Technol. Appl. 2023, 5, 100310. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Saha, N.; Ngwabebhoh, F.A.; Zandraa, O.; Saha, T.; Saha, P. Silane-Modified Kombucha-Derived Cellulose/Polyurethane/Polylactic Acid Biocomposites for Prospective Application as Leather Alternative. Sustain. Mater. Technol. 2023, 36, e00611. [Google Scholar] [CrossRef]
- Charoenrak, S.; Charumanee, S.; Sirisa-ard, P.; Bovonsombut, S.; Kumdhitiahutsawakul, L.; Kiatkarun, S.; Pathom-Aree, W.; Chitov, T.; Bovonsombut, S. Nanobacterial Cellulose from Kombucha Fermentation as a Potential Protective Carrier of Lactobacillus plantarum under Simulated Gastrointestinal Tract Conditions. Polymers 2023, 15, 1356. [Google Scholar] [CrossRef]
- Karuppannan, S.; N, S. Influence of Domestic Food Waste Intrusion on Microbes Producing Cellulose. Biomass Convers. Biorefinery 2024, 14, 1–10. [Google Scholar] [CrossRef]
- Kalashnikova, O.; Pankova, E.; Sukhikh, S.; Babich, O.; Samusev, I.; Tcibulnikova, A.; Ivanova, S.; Kriger, O. Production of Bacterial Cellulose Using a Symbiotic Consortium of Bacteria and Yeast on Soybean Molasses Medium. LWT 2024, 205, 116480. [Google Scholar] [CrossRef]
- Wang, B.; Rutherfurd-Markwick, K.; Zhang, X.X.; Mutukumira, A.N. Kombucha: Production and Microbiological Research. Foods 2022, 11, 3456. [Google Scholar] [CrossRef]
- Aulitto, M.; Strazzulli, A.; Sansone, F.; Cozzolino, F.; Monti, M.; Moracci, M.; Fiorentino, G.; Limauro, D.; Bartolucci, S.; Contursi, P. Prebiotic Properties of Bacillus coagulans MA-13: Production of Galactoside Hydrolyzing Enzymes and Characterization of the Transglycosylation Properties of a GH42 β-Galactosidase. Microb. Cell Factories 2021, 20, 71. [Google Scholar] [CrossRef]
- Ryngajłło, M.; Jacek, P.; Cielecka, I.; Kalinowska, H.; Bielecki, S. Effect of Ethanol Supplementation on the Transcriptional Landscape of Bionanocellulose Producer Komagataeibacter xylinus E25. Appl. Microbiol. Biotechnol. 2019, 103, 6673–6688. [Google Scholar] [CrossRef]
- Su, J.; Tan, Q.; Tang, Q.; Tong, Z.; Yang, M. Research Progress on Alternative Kombucha Substrate Transformation and the Resulting Active Components. Front. Microbiol. 2023, 14, 1254014. [Google Scholar]
- Treviño-Garza, M.Z.; Guerrero-Medina, A.S.; González-Sánchez, R.A.; García-Gómez, C.; Guzmán-Velasco, A.; Báez-González, J.G.; Márquez-Reyes, J.M. Production of Microbial Cellulose Films from Green Tea (Camellia sinensis) Kombucha with Various Carbon Sources. Coatings 2020, 10, 1132. [Google Scholar] [CrossRef]
- Agüero, A.; Lascano, D.; Ivorra-Martinez, J.; Gómez-Caturla, J.; Arrieta, M.P.; Balart, R. Use of Bacterial Cellulose Obtained from Kombucha Fermentation in Spent Coffee Grounds for Active Composites Based on PLA and Maleinized Linseed Oil. Ind. Crops Prod. 2023, 202, 116971. [Google Scholar] [CrossRef]
- Jarrell, J.A.; Walia, N.; Nemergut, D.; Agadi, A.; Bennett, J.W. Inoculation, Growth and Bactericidal Effects of Three Kombucha Cultures. Microbiol. Res. 2022, 13, 128–136. [Google Scholar] [CrossRef]
- Priyadharshini, T.; Nageshwari, K.; Vimaladhasan, S.; Parag Prakash, S.; Balasubramanian, P. Machine Learning Prediction of SCOBY Cellulose Yield from Kombucha Tea Fermentation. Bioresour. Technol. Rep. 2022, 18, 101027. [Google Scholar] [CrossRef]
- Al-Kalifawi, E.J.; Hassan, I.A. Factors Influence on the Yield of Bacterial Cellulose of Kombucha (Khubdat Humza). Baghdad Sci. J. 2014, 11, 1420–1428. [Google Scholar] [CrossRef]
- Balasubramanian, P.; Praharaj, P.T. Principal Component Analysis Revealed the Key Influencing Factors of Kombucha Bacterial Cellulose Yield and Productivity. Bioresour. Technol. Rep. 2023, 23, 101539. [Google Scholar] [CrossRef]
- Cahyaningtyas, H.A.A.; Renaldi, G.; Fibriana, F.; Mulyani, W.E. Cost-Effective Production of Kombucha Bacterial Cellulose by Evaluating Nutrient Sources, Quality Assessment, and Dyeing Methods. Environ. Sci. Pollut. Res. 2025, 32, 2713–2725. [Google Scholar] [CrossRef]
- Selvaraj, S.; Gurumurthy, K. Optimizing Bacterial Cellulose Production From Kombucha Tea Utilizing Molasses and Tea Dust in Growth Medium. Int. J. Polym. Sci. 2024, 2024, 2240464. [Google Scholar] [CrossRef]
- Feng, X.; Ge, Z.; Wang, Y.; Xia, X.; Zhao, B.; Dong, M. Production and Characterization of Bacterial Cellulose from Kombucha-Fermented Soy Whey. Food Prod. Process. Nutr. 2024, 6, 20. [Google Scholar] [CrossRef]
- Guimarães, A.C.; Leonarski, E.; Cesca, K.; Poletto, P. Bacterial Cellulose from Kombucha: Assessing Inoculum Age and Concentration, and Its Conversion via Enzymatic Hydrolysis into Cellobiose and Glucose. Biocatal. Agric. Biotechnol. 2024, 59, 103244. [Google Scholar] [CrossRef]
- Sharma, C.; Bhardwaj, N.K. Bacterial Nanocellulose: Present Status, Biomedical Applications and Future Perspectives. Mater. Sci. Eng. C 2019, 104, 109963. [Google Scholar]
- Qi, Y.; Guo, Y.; Liza, A.A.; Yang, G.; Sipponen, M.H.; Guo, J.; Li, H. Nanocellulose: A Review on Preparation Routes and Applications in Functional Materials. Cellulose 2023, 30, 4115–4147. [Google Scholar]
- Mishra, S.; Singh, P.K.; Pattnaik, R.; Kumar, S.; Ojha, S.K.; Srichandan, H.; Parhi, P.K.; Jyothi, R.K.; Sarangi, P.K. Biochemistry, Synthesis, and Applications of Bacterial Cellulose: A Review. Front. Bioeng. Biotechnol. 2022, 10, 780409. [Google Scholar] [CrossRef] [PubMed]
- Urbina, L.; Corcuera, M.Á.; Gabilondo, N.; Eceiza, A.; Retegi, A. A Review of Bacterial Cellulose: Sustainable Production from Agricultural Waste and Applications in Various Fields. Cellulose 2021, 28, 8229–8253. [Google Scholar]
- Martinez, J.C.; Davila, L.; Zuluaga, R.; Ricardo, A.; Figueroa, J.; Oliveira, M.G.d.A.; Meriño-Cabrera, Y. Production of Bacterial Cellulose from Banana Waste (Musa paradisiaca): A Sustainable Material for Making Cardboard Paper. Biocatal. Agric. Biotechnol. 2025, 63, 103484. [Google Scholar] [CrossRef]
- Algar, I.; Fernandes, S.C.M.; Mondragon, G.; Castro, C.; Garcia-Astrain, C.; Gabilondo, N.; Retegi, A.; Eceiza, A. Pineapple Agroindustrial Residues for the Production of High Value Bacterial Cellulose with Different Morphologies. J. Appl. Polym. Sci. 2015, 132, 41237. [Google Scholar] [CrossRef]
- Kurosumi, A.; Sasaki, C.; Yamashita, Y.; Nakamura, Y. Utilization of Various Fruit Juices as Carbon Source for Production of Bacterial Cellulose by Acetobacter xylinum NBRC 13693. Carbohydr. Polym. 2009, 76, 333–335. [Google Scholar] [CrossRef]
- Brugnoli, M.; La China, S.; Lasagni, F.; Romeo, F.V.; Pulvirenti, A.; Gullo, M. Acetic Acid Bacteria in Agro-Wastes: From Cheese Whey and Olive Mill Wastewater to Cellulose. Appl. Microbiol. Biotechnol. 2023, 107, 3729–3744. [Google Scholar] [CrossRef]
- Güzel, M. Valorisation of Bread Wastes via the Bacterial Cellulose Production. Biomass Convers. Biorefinery 2025, 15, 4777–4790. [Google Scholar] [CrossRef]
- Hasanin, M.S.; Abdelraof, M.; Hashem, A.H.; El Saied, H. Sustainable Bacterial Cellulose Production by Achromobacter Using Mango Peel Waste. Microb. Cell Factories 2023, 22, 24. [Google Scholar] [CrossRef]
- Jozala, A.F.; Pértile, R.A.N.; dos Santos, C.A.; de Carvalho Santos-Ebinuma, V.; Seckler, M.M.; Gama, F.M.; Pessoa, A. Bacterial Cellulose Production by Gluconacetobacter xylinus by Employing Alternative Culture Media. Appl. Microbiol. Biotechnol. 2015, 99, 1181–1190. [Google Scholar] [CrossRef]
- Bilgi, E.; Bayir, E.; Sendemir-Urkmez, A.; Hames, E.E. Optimization of Bacterial Cellulose Production by Gluconacetobacter xylinus Using Carob and Haricot Bean. Int. J. Biol. Macromol. 2016, 90, 2–10. [Google Scholar] [CrossRef]
- Heydorn, R.L.; Lammers, D.; Gottschling, M.; Dohnt, K. Effect of Food Industry By-Products on Bacterial Cellulose Production and Its Structural Properties. Cellulose 2023, 30, 4159–4179. [Google Scholar] [CrossRef]
- Rani, M.U.; Appaiah, K.A.A. Production of Bacterial Cellulose by Gluconacetobacter hansenii UAC09 Using Coffee Cherry Husk. J. Food Sci. Technol. 2013, 50, 755–762. [Google Scholar] [CrossRef]
- Zeng, X.; Small, D.P.; Wan, W. Statistical Optimization of Culture Conditions for Bacterial Cellulose Production by Acetobacter xylinum BPR 2001 from Maple Syrup. Carbohydr. Polym. 2011, 85, 506–513. [Google Scholar] [CrossRef]
- Castro, C.; Zuluaga, R.; Putaux, J.L.; Caro, G.; Mondragon, I.; Gañán, P. Structural Characterization of Bacterial Cellulose Produced by Gluconacetobacter swingsii sp. from Colombian Agroindustrial Wastes. Carbohydr. Polym. 2011, 84, 96–102. [Google Scholar] [CrossRef]
- Jittaut, P.; Hongsachart, P.; Audtarat, S.; Dasri, T. Production and Characterization of Bacterial Cellulose Produced by Gluconacetobacter xylinus BNKC 19 Using Agricultural Waste Products as Nutrient Source. Arab. J. Basic Appl. Sci. 2023, 30, 221–230. [Google Scholar] [CrossRef]
- Quiñones-Cerna, C.; Rodríguez-Soto, J.C.; Barraza-Jáuregui, G.; Huanes-Carranza, J.; Cruz-Monzón, J.A.; Ugarte-López, W.; Hurtado-Butrón, F.; Samanamud-Moreno, F.; Haro-Carranza, D.; Valdivieso-Moreno, S.; et al. Bioconversion of Agroindustrial Asparagus Waste into Bacterial Cellulose by Komagataeibacter rhaeticus. Sustainability 2024, 16, 736. [Google Scholar] [CrossRef]
- Esmail, A.; Morais, M.; Yilmazer, U.D.; Neves, L.A.; Freitas, F. Bacterial Cellulose Production through the Valorization of Waste Apple Pulp and Stale Bread. Biomass Convers. Biorefinery 2024, 14, 1–16. [Google Scholar] [CrossRef]
- Wu, J.M.; Liu, R.H. Thin Stillage Supplementation Greatly Enhances Bacterial Cellulose Production by Gluconacetobacter xylinus. Carbohydr. Polym. 2012, 90, 116–121. [Google Scholar] [CrossRef]
- Abdelraof, M.; Hasanin, M.S.; El-Saied, H. Ecofriendly Green Conversion of Potato Peel Wastes to High Productivity Bacterial Cellulose. Carbohydr. Polym. 2019, 211, 75–83. [Google Scholar] [CrossRef]
- Chen, L.; Hong, F.; Yang, X.-X.; Han, S.-F. Biotransformation of Wheat Straw to Bacterial Cellulose and Its Mechanism. Bioresour. Technol. 2013, 135, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Carreira, P.; Mendes, J.A.S.; Trovatti, E.; Serafim, L.S.; Freire, C.S.R.; Silvestre, A.J.D.; Neto, C.P. Utilization of Residues from Agro-Forest Industries in the Production of High Value Bacterial Cellulose. Bioresour. Technol. 2011, 102, 7354–7360. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, L.; Hua, J.; Jia, S.; Zhang, J.; Liu, H. Production of Nano Bacterial Cellulose from Waste Water of Candied Jujube-Processing Industry Using Acetobacter xylinum. Carbohydr. Polym. 2015, 120, 115–119. [Google Scholar] [CrossRef]
- Ha, J.H.; Shehzad, O.; Khan, S.; Lee, Y.; Won Park, J.; Khan, T.; Park, J.K. Production of Bacterial Cellulose by a Static Cultivation Using the Waste from Beer Culture Broth. Korean J. Chem. Eng. 2008, 25, 812–815. [Google Scholar] [CrossRef]
- Riva, L.; Bonetti, L.; Utoiu, E.; Sorin Manoiu, V.; Oprita, E.I.; Craciunescu, O. Bacterial Cellulose: A Sustainable Source for Hydrogels and 3D-Printed Scaffolds for Tissue Engineering. Gels 2024, 10, 387. [Google Scholar] [CrossRef]
- Zarepour, A.; Gok, B.; Budama-Kilinc, Y.; Khosravi, A.; Iravani, S.; Zarrabi, A. Bacterial Nanocelluloses as Sustainable Biomaterials for Advanced Wound Healing and Dressings. J. Mater. Chem. B 2024, 12, 12489–12507. [Google Scholar] [CrossRef]
- Ansari, M.M.; Heo, Y.; Do, K.; Ghosh, M.; Son, Y.O. Nanocellulose Derived from Agricultural Biowaste By-Products–Sustainable Synthesis, Biocompatibility, Biomedical Applications, and Future Perspectives: A Review. Carbohydr. Polym. Technol. Appl. 2024, 8, 100529. [Google Scholar] [CrossRef]
- Cruz, M.A.; Flor-Unda, O.; Avila, A.; Garcia, M.D.; Cerda-Mejía, L. Advances in Bacterial Cellulose Production: A Scoping Review. Coatings 2024, 14, 1401. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, M.; Li, J.; Hu, C.; Li, J.; Xiong, R.; Huang, C. Nanocellulose Based Hydrogel for Flexible Sensors: Current Progress and Future Perspective. Nano Energy 2024, 129, 109974. [Google Scholar] [CrossRef]
- Choi, J.; Sung, K.; Hyun, J.; Shin, S. Sheet-Laminated Additive Manufacturing of Bacterial Cellulose Nanofiber-Reinforced Hydrogels. Carbohydr. Polym. 2025, 349, 122972. [Google Scholar] [CrossRef]
- Pillai, M.M.; Tran, H.N.; Sathishkumar, G.; Manimekalai, K.; Yoon, J.H.; Lim, D.Y.; Noh, I.; Bhattacharyya, A. Symbiotic Culture of Nanocellulose Pellicle: A Potential Matrix for 3D Bioprinting. Mater. Sci. Eng. C 2021, 119, 111552. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tavakoli, J.; Tang, Y. Bacterial Cellulose Production, Properties and Applications with Different Culture Methods—A Review. Carbohydr. Polym. 2019, 219, 63–76. [Google Scholar] [CrossRef]
- Wang, Z.; Li, S.; Zhao, X.; Liu, Z.; Shi, R.; Hao, M. Applications of Bacterial Cellulose in the Food Industry and Its Health-Promoting Potential. Food Chem. 2025, 464, 141763. [Google Scholar] [CrossRef] [PubMed]
- Acharyya, P.P.; Sarma, M.; Kashyap, A. Recent Advances in Synthesis and Bioengineering of Bacterial Nanocellulose Composite Films for Green, Active and Intelligent Food Packaging. Cellulose 2024, 31, 7163–7187. [Google Scholar]
- Chen, Q.; Yang, F.; Hou, Y.; Li, Z.; Yuan, D.; Liu, C.; Hu, F.; Zhao, R.; Wang, H.; Liu, W.; et al. Bacterial Cellulose-Based Pickering Emulsions Reinforced with Silver and Silica Nanoparticles for Advanced Antibacterial and Hydrophobic Food Packaging Solutions. Carbohydr. Polym. 2025, 355, 123357. [Google Scholar] [CrossRef]
- Liu, Z.; Lin, D.; Shen, R.; Yang, X. Bacterial Cellulose Nanofibers Improved the Emulsifying Capacity of Soy Protein Isolate as a Stabilizer for Pickering High Internal-Phase Emulsions. Food Hydrocoll. 2021, 112, 106279. [Google Scholar] [CrossRef]
- Xavier, J.R.; Ramana, K.V. Development of Slow Melting Dietary Fiber-Enriched Ice Cream Formulation Using Bacterial Cellulose and Inulin. J. Food Process. Preserv. 2022, 46, e15394. [Google Scholar] [CrossRef]
- Xie, Y.; Lei, Y.; Rong, J.; Zhang, X.; Li, J.; Chen, Y.; Liang, H.; Li, Y.; Li, B.; Fang, Z.; et al. Physico-Chemical Properties of Reduced-Fat Biscuits Prepared Using O/W Cellulose-Based Pickering Emulsion. LWT 2021, 148, 111745. [Google Scholar] [CrossRef]
- Lima, N.F.; Maciel, G.M.; Lima, N.P.; Fernandes, I.d.A.A.; Haminiuk, C.W.I. Bacterial Cellulose in Cosmetic Innovation: A Review. Int. J. Biol. Macromol. 2024, 275, 133396. [Google Scholar] [CrossRef]
- Yan, C.; Chen, J.; Wan, X.; Nilghaz, A.; Cao, R.; Xie, Y.; Zhang, C.; Tian, J. Smart Bacteria Cellulose Facial Mask for Sensing and Recovering Skin PH. Int. J. Biol. Macromol. 2025, 306, 141286. [Google Scholar] [CrossRef]
- Nashed Kabalan, O.; Marceau, S.; Ciblac, T.; Le Roy, R. Influence of Bacterial Cellulose Stabilization on Strength Characteristics of Construction Earthen Materials. Mater. Struct. 2024, 57, 118. [Google Scholar] [CrossRef]
- Schiros, T.N.; Antrobus, R.; Farías, D.; Chiu, Y.T.; Joseph, C.T.; Esdaille, S.; Sanchirico, G.K.; Miquelon, G.; An, D.; Russell, S.T.; et al. Microbial Nanocellulose Biotextiles for a Circular Materials Economy. Environ. Sci. Adv. 2022, 1, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Felgueiras, C.; Azoia, N.G.; Gonçalves, C.; Gama, M.; Dourado, F. Trends on the Cellulose-Based Textiles: Raw Materials and Technologies. Front. Bioeng. Biotechnol. 2021, 9, 608826. [Google Scholar] [CrossRef]
- Peters, G.; Li, M.; Lenzen, M. The Need to Decelerate Fast Fashion in a Hot Climate—A Global Sustainability Perspective on the Garment Industry. J. Clean. Prod. 2021, 295, 126390. [Google Scholar] [CrossRef]
- Bailey, K.; Basu, A.; Sharma, S. The Environmental Impacts of Fast Fashion on Water Quality: A Systematic Review. Water 2022, 14, 1073. [Google Scholar] [CrossRef]
- Nayak, R.; Cleveland, D.; Tran, G.; Joseph, F. Potential of Bacterial Cellulose for Sustainable Fashion and Textile Applications: A Review. J. Mater. Sci. 2024, 59, 6685–6710. [Google Scholar] [CrossRef]
- Locker, C.R.; Theregowda, R. Life-Cycle Assessment of Bioleather1. Clean. Circ. Bioecon. 2022, 1, 100003. [Google Scholar] [CrossRef]
- Tewari, S.; Reshamwala, S.M.S.; Bhatt, L.; Kale, R.D. Vegan Leather: A Sustainable Reality or a Marketing Gimmick? Environ. Sci. Pollut. Res. 2023, 31, 3361–3375. [Google Scholar] [CrossRef]
- Kefale, G.Y.; Kebede, Z.T.; Birlie, A.A. A Systematic Review on Potential Bio Leather Substitute for Natural Leather. J. Eng. 2023, 2023, 1629174. [Google Scholar] [CrossRef]
- Williams, E.; Cenian, K.; Golsteijn, L.; Morris, B.; Scullin, M.L. Life Cycle Assessment of MycoWorks’ ReishiTM: The First Low-Carbon and Biodegradable Alternative Leather. Environ. Sci. Eur. 2022, 34, 120. [Google Scholar] [CrossRef]
- Rodrigues, I.; Mata, T.M.; Martins, A.A. Environmental Analysis of a Bio-Based Coating Material for Automobile Interiors. J. Clean. Prod. 2022, 367, 133011. [Google Scholar] [CrossRef]
- Mogas-Soldevila, L.; Matzeu, G.; Presti, M.L.; Omenetto, F.G. Additively Manufactured Leather-like Silk Protein Materials. Mater. Des. 2021, 203, 109631. [Google Scholar] [CrossRef]
- Raksaksri, L.; Phunpeng, V. Leather-like Composite Materials Prepared from Natural Rubber and Two Leather Wastes: Wet Blue Leather and Finished Leather. J. Elastomers Plast. 2022, 54, 1254–1276. [Google Scholar] [CrossRef]
- Wijayarathna, E.R.K.B.; Mohammadkhani, G.; Soufiani, A.M.; Adolfsson, K.H.; Ferreira, J.A.; Hakkarainen, M.; Berglund, L.; Heinmaa, I.; Root, A.; Zamani, A. Fungal Textile Alternatives from Bread Waste with Leather-like Properties. Resour. Conserv. Recycl. 2022, 179, 106041. [Google Scholar] [CrossRef]
- Bustillos, J.; Loganathan, A.; Agrawal, R.; Gonzalez, B.A.; Perez, M.G.; Ramaswamy, S.; Boesl, B.; Agarwal, A. Uncovering the Mechanical, Thermal, and Chemical Characteristics of Biodegradable Mushroom Leather with Intrinsic Antifungal and Antibacterial Properties. ACS Appl. Bio. Mater. 2020, 3, 3145–3156. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Saha, N.; Ngwabebhoh, F.A.; Zandraa, O.; Saha, T.; Saha, P. Kombucha-Derived Bacterial Cellulose from Diverse Wastes: A Prudent Leather Alternative. Cellulose 2021, 28, 9335–9353. [Google Scholar] [CrossRef]
- Silva, F.A.G.S.; Branco, S.; Dourado, F.; Neto, B.; Gama, M. Life Cycle Assessment of Bacterial Cellulose and Comparison to Other Cellulosic Sources. J. Clean. Prod. 2025, 493, 144876. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, Z.; Yuan, J.; Xu, J.; Ji, Q.; Bai, Y. Eco-Friendly Production of Leather-like Material from Bacterial Cellulose and Waste Resources. J. Clean. Prod. 2024, 476, 143700. [Google Scholar] [CrossRef]
- Kamiński, K.; Jarosz, M.; Grudzień, J.; Pawlik, J.; Zastawnik, F.; Pandyra, P.; Kołodziejczyk, A.M. Hydrogel Bacterial Cellulose: A Path to Improved Materials for New Eco-Friendly Textiles. Cellulose 2020, 27, 5353–5365. [Google Scholar] [CrossRef]
- Chan, C.K.; Shin, J.; Jiang, S.X.K. Development of Tailor-Shaped Bacterial Cellulose Textile Cultivation Techniques for Zero-Waste Design. Cloth. Text. Res. J. 2018, 36, 33–44. [Google Scholar] [CrossRef]
- Kim, H.; Song, J.E.; Kim, H.R. Comparative Study on the Physical Entrapment of Soy and Mushroom Proteins on the Durability of Bacterial Cellulose Bio-leather. Cellulose 2021, 28, 3183–3200. [Google Scholar] [CrossRef]
- Fernandes, M.; Gama, M.; Dourado, F.; Souto, A.P. Development of Novel Bacterial Cellulose Composites for the Textile and Shoe Industry. Microb. Biotechnol. 2019, 12, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Ayyappan, V.G.; Vhatkar, S.S.; Bose, S.; Sampath, S.; Das, S.K.; Samanta, D.; Mandal, A.B. Incorporations of Gold, Silver and Carbon Nanomaterials to Kombucha-Derived Bacterial Cellulose: Development of Antibacterial Leather-like Materials. J. Indian Chem. Soc. 2022, 99, 100278. [Google Scholar] [CrossRef]
- Domskiene, J.; Sederaviciute, F.; Simonaityte, J. Kombucha Bacterial Cellulose for Sustainable Fashion. Int. J. Cloth. Sci. Technol. 2019, 31, 644–652. [Google Scholar] [CrossRef]
- Saha, N.; Ngwabebhoh, F.A.; Nguyen, H.T.; Saha, P. Environmentally Friendly and Animal Free Leather: Fabrication and Characterization. In AIP Conference Proceedings; American Institute of Physics: College Park, MD, USA, 2020; Volume 2289. [Google Scholar]
- Lisowski, D.; Bielecki, S.; Cichosz, S.; Masek, A. Ecologically Modified Leather of Bacterial Origin. Materials 2024, 17, 2783. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.A.G.S.; Schlapp-Hackl, I.; Nygren, N.; Heimala, S.; Leinonen, A.; Dourado, F.; Gama, M.; Hummel, M. Upcycling of Cellulosic Textile Waste with Bacterial Cellulose via Ioncell® Technology. Int J Biol Macromol 2024, 271, 132194. [Google Scholar] [CrossRef]
- Abouzaid, H.A.K. Production and Investigation of Bio-Textile Films Produced from Bacterial Cellulose Biosynthesis from Black Tea and Ginger, and Cultivation on Sugar Cane Media. J. Ind. Text. 2024, 54, 15280837231224076. [Google Scholar] [CrossRef]
- Walker, K.T.; Li, I.S.; Keane, J.; Goosens, V.J.; Song, W.; Lee, K.Y.; Ellis, T. Self-Pigmenting Textiles Grown from Cellulose-Producing Bacteria with Engineered Tyrosinase Expression. Nat. Biotechnol. 2024, 43, 345–354. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venturelli, G.; Villa, F.; Petraretti, M.; Guagliano, G.; Levi, M.; Petrini, P. Bacterial Cellulose for Scalable and Sustainable Bio-Gels in the Circular Economy. Gels 2025, 11, 262. https://doi.org/10.3390/gels11040262
Venturelli G, Villa F, Petraretti M, Guagliano G, Levi M, Petrini P. Bacterial Cellulose for Scalable and Sustainable Bio-Gels in the Circular Economy. Gels. 2025; 11(4):262. https://doi.org/10.3390/gels11040262
Chicago/Turabian StyleVenturelli, Giovanni, Federica Villa, Mariagioia Petraretti, Giuseppe Guagliano, Marinella Levi, and Paola Petrini. 2025. "Bacterial Cellulose for Scalable and Sustainable Bio-Gels in the Circular Economy" Gels 11, no. 4: 262. https://doi.org/10.3390/gels11040262
APA StyleVenturelli, G., Villa, F., Petraretti, M., Guagliano, G., Levi, M., & Petrini, P. (2025). Bacterial Cellulose for Scalable and Sustainable Bio-Gels in the Circular Economy. Gels, 11(4), 262. https://doi.org/10.3390/gels11040262