
Soil Fungal Community Characteristics at Timberlines of Sejila Mountain in Southeast Tibet, China

 ,
,
Abstract
1. Introduction
2. Materials and Methods
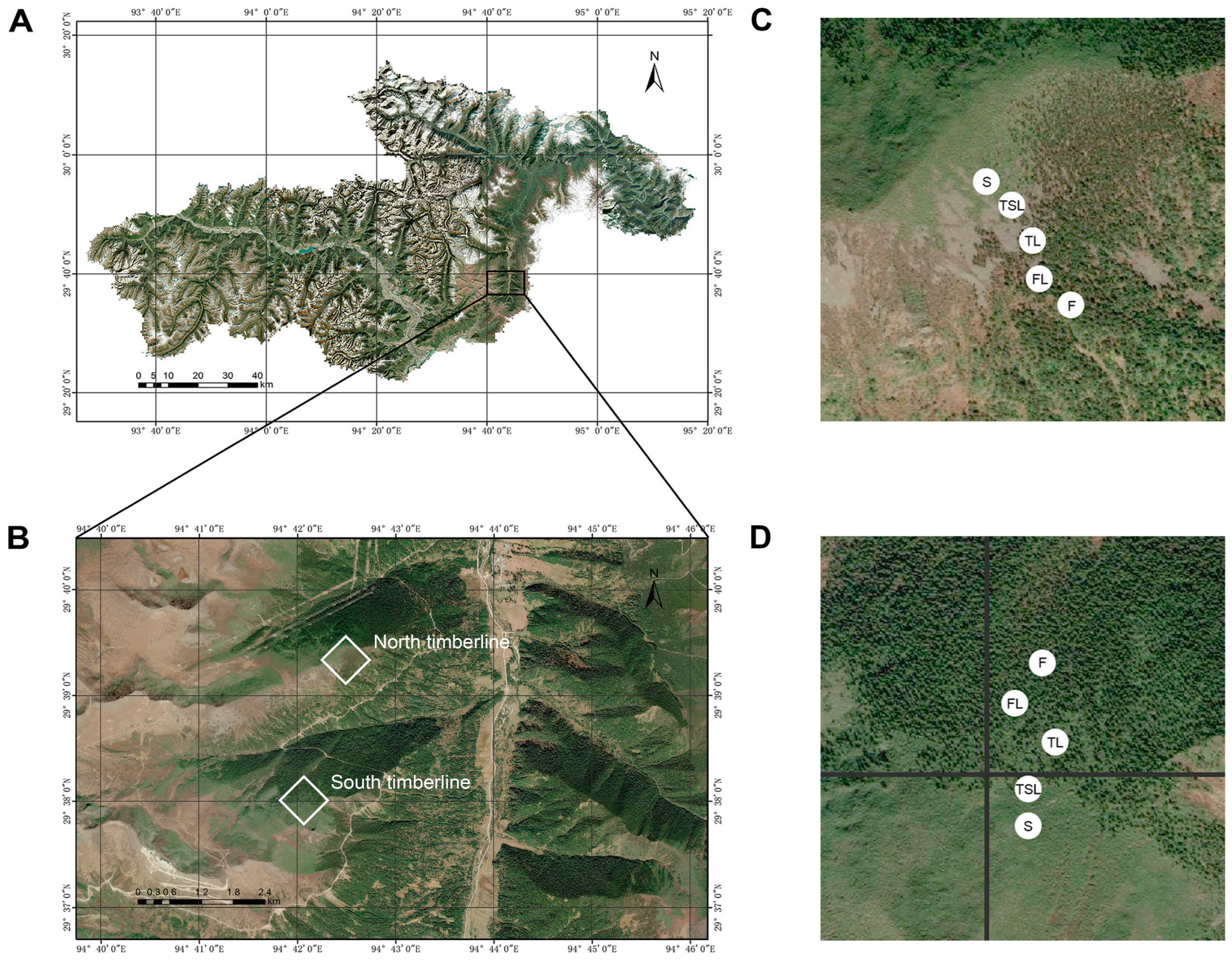
2.1. Study Site
2.2. Plot Setting and Vegetation Survey
2.3. Sample Collection
2.4. Determination of Soil Physical and Chemical Properties
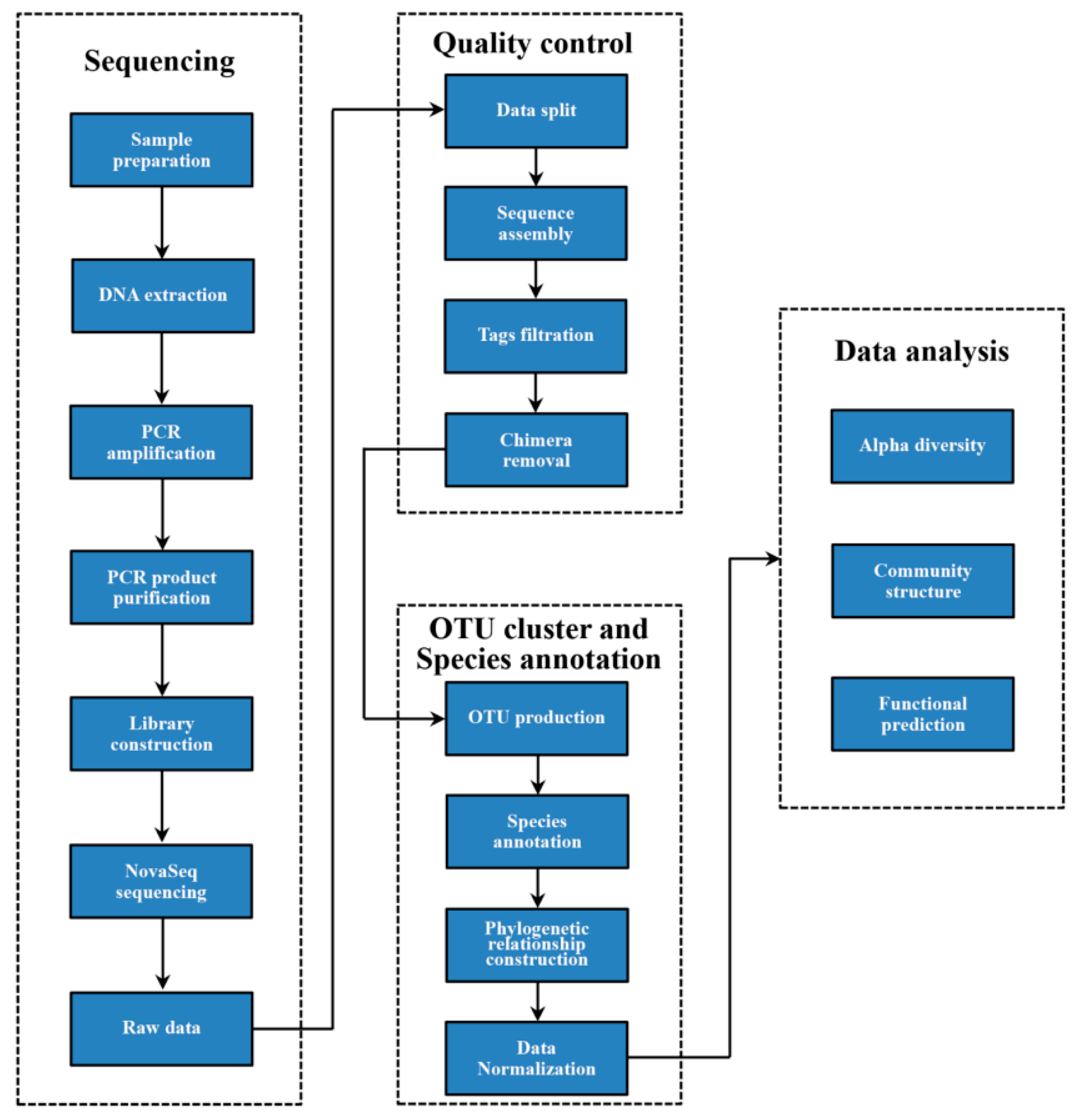
2.5. High-Throughput Sequencing of Soil Fungi
2.5.1. Soil Fungal Genomic DNA Extraction and PCR Amplification
2.5.2. Library Construction and Sequencing
2.5.3. OTU Clustering and Taxonomic Annotation
2.6. Data Analysis
3. Results
3.1. Vegetation and Soil Characteristics
3.1.1. Vegetation
3.1.2. Soil
3.2. Overview of High-Throughput Sequencing
3.3. Soil Fungal Diversity
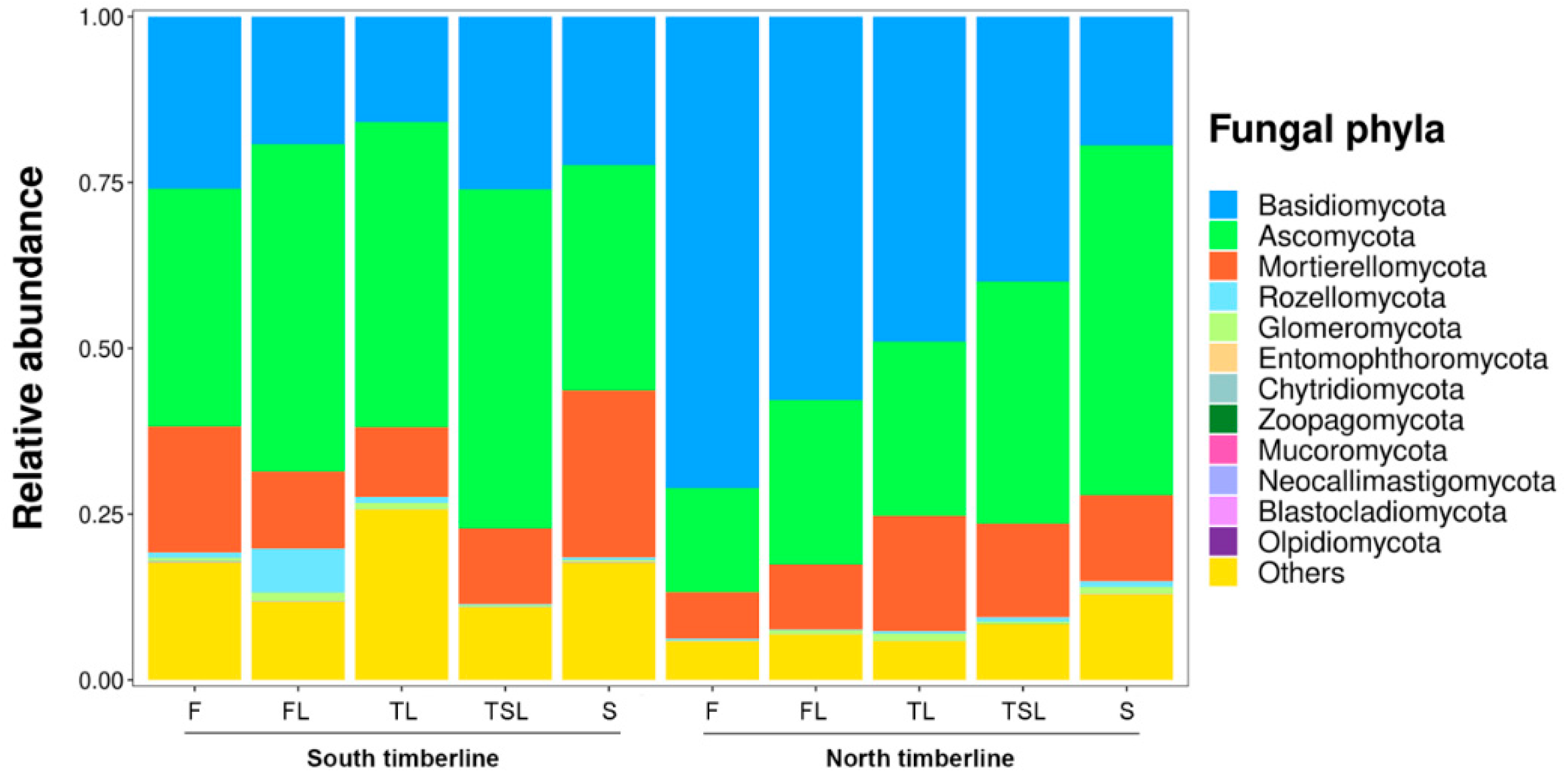
3.4. Soil Fungal Community Composition
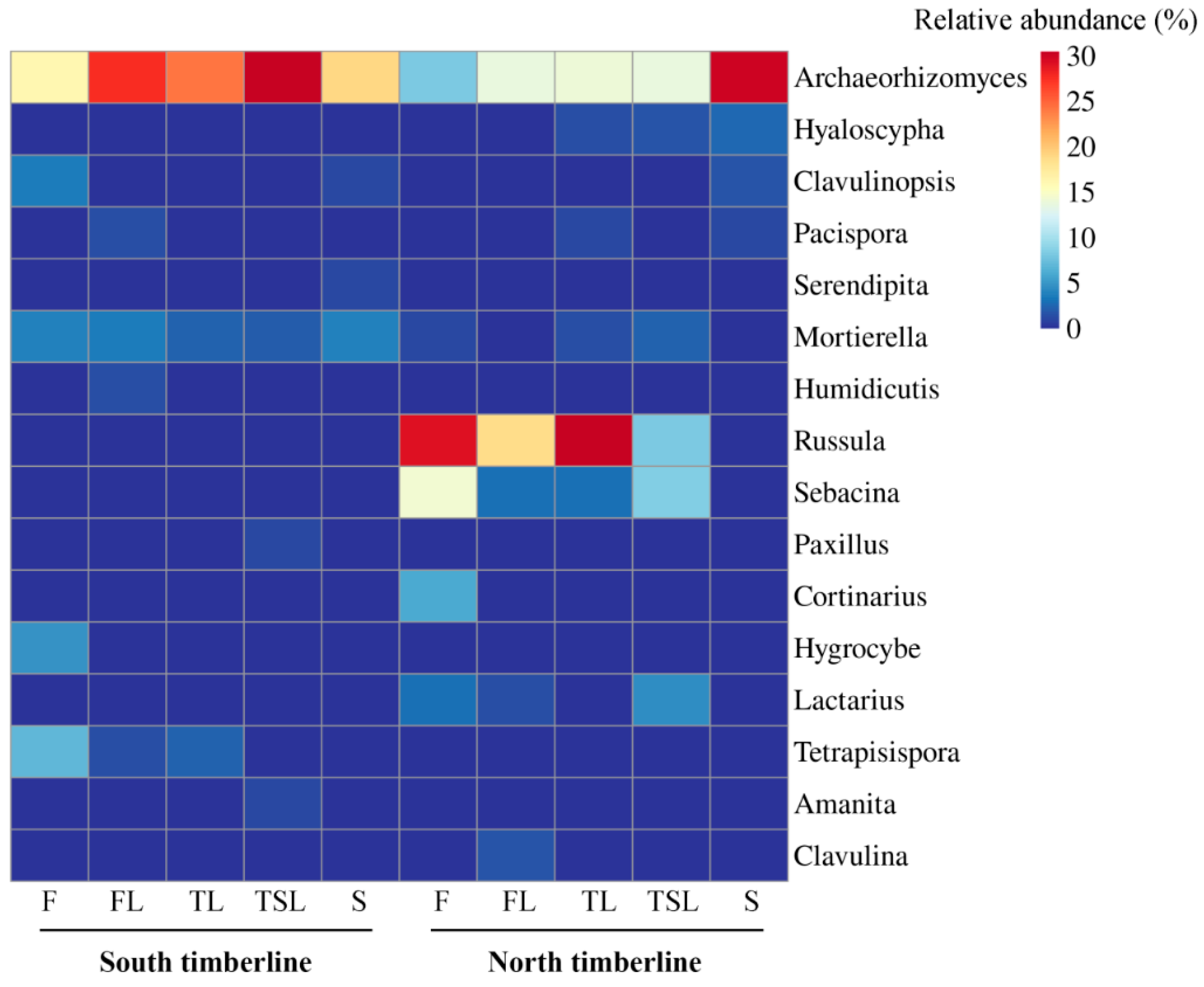
3.4.1. Relative Abundance of Soil Fungal Phyla and Genera
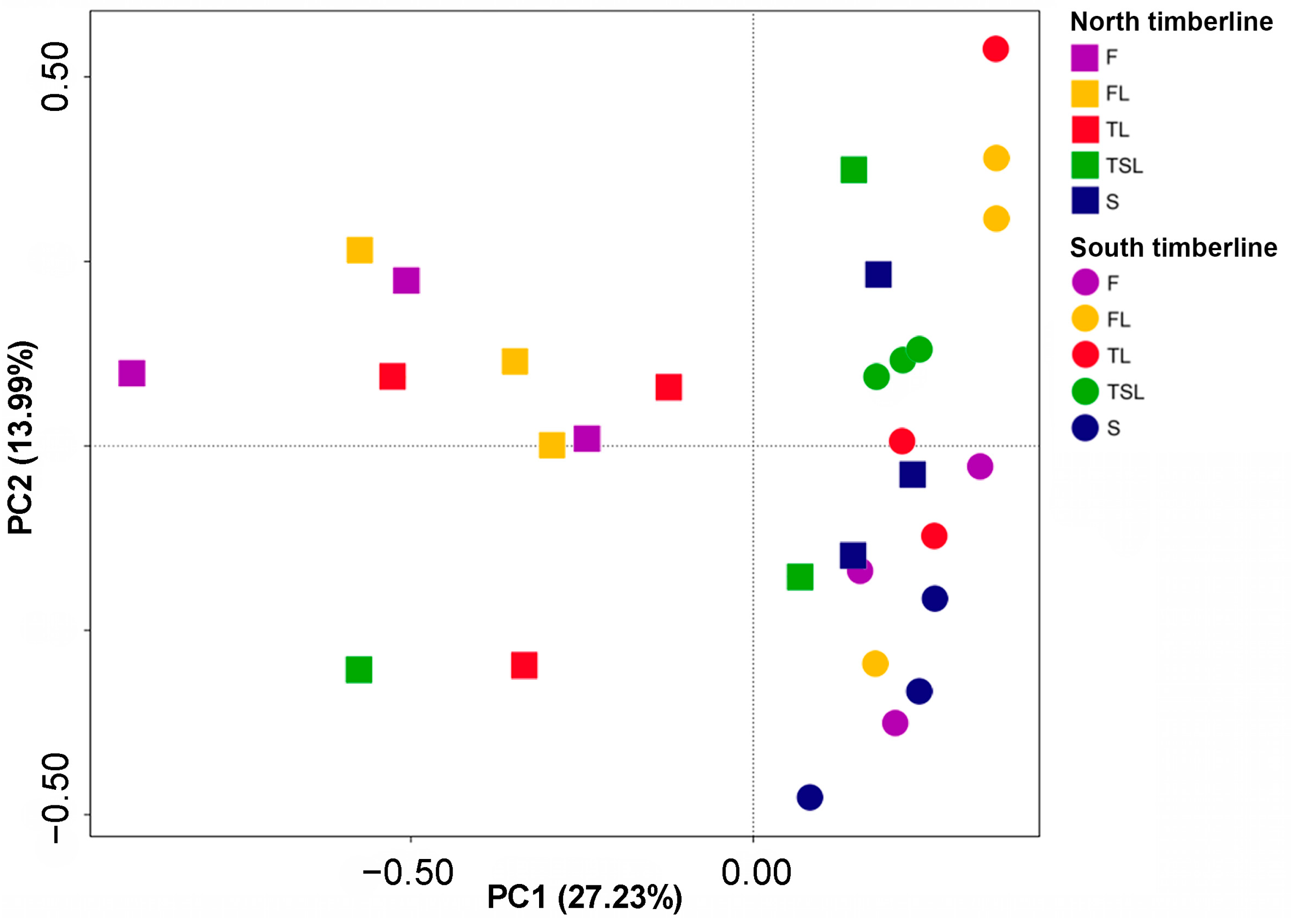
3.4.2. Soil Fungal Community Similarity
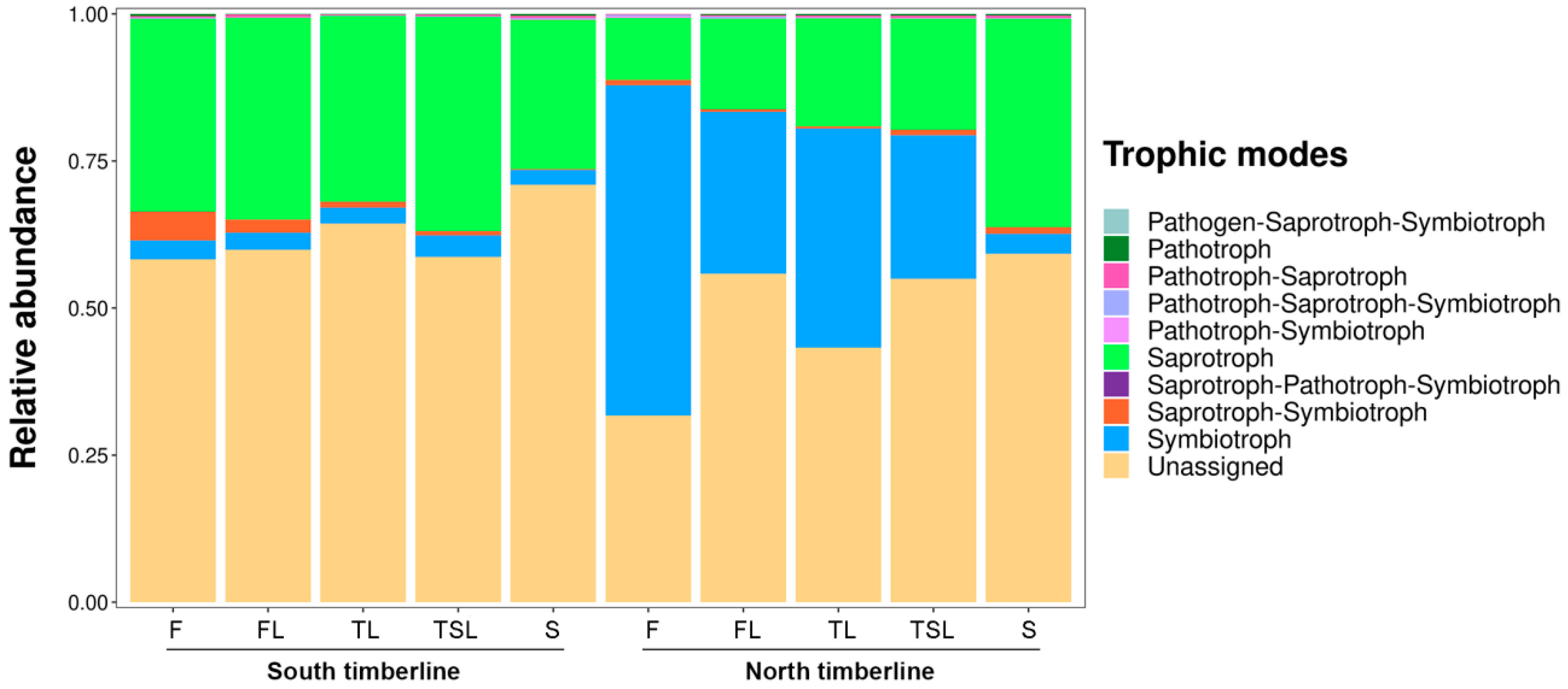
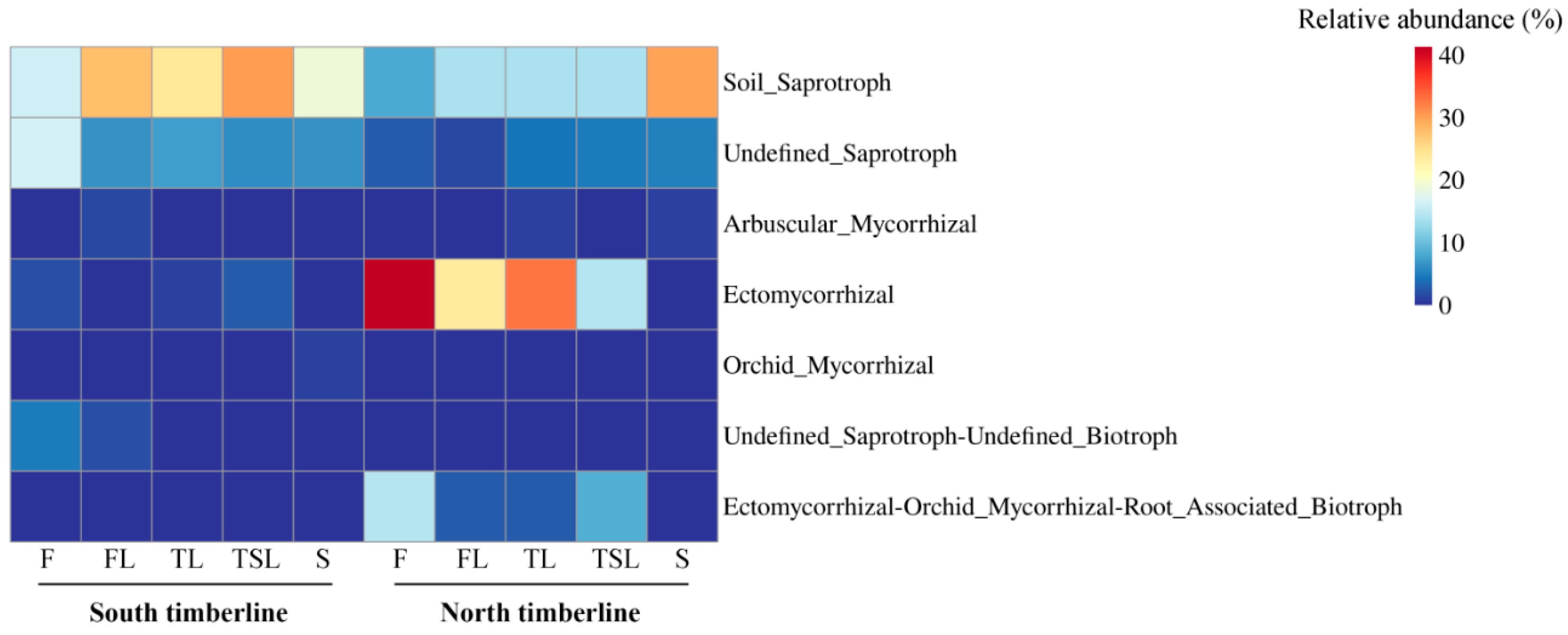
3.5. Ecological Function of Soil Fungi
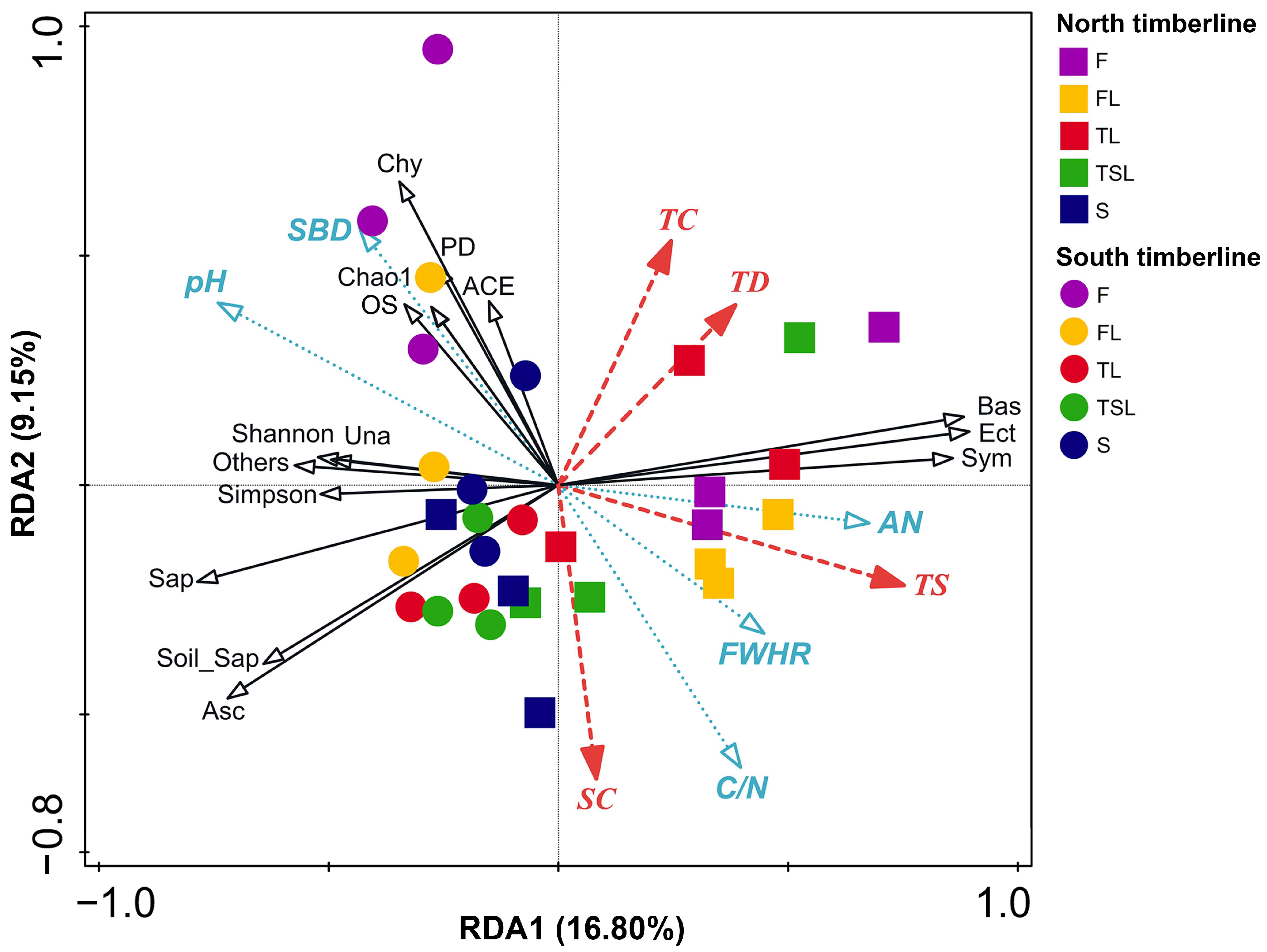
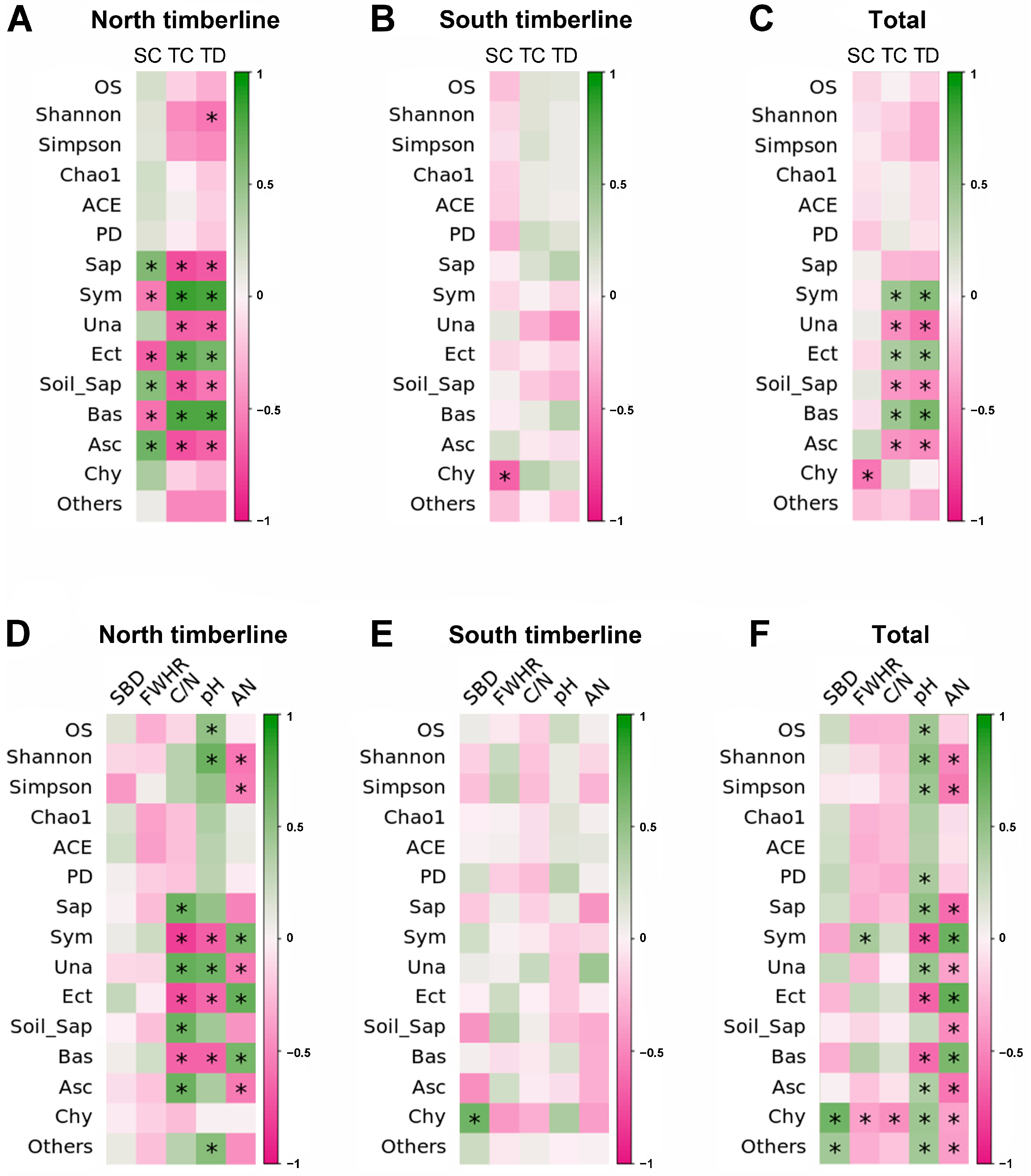
3.6. Relationship between Soil Fungal Community Characteristics and Vegetation and Soil Factors
4. Discussion
4.1. Distribution of Mycorrhizal Fungi and Its Relationship with Vegetation
4.2. Distribution of Saprophytic Fungi and Its Relationship with Vegetation
4.3. Relationship between Fungal Distribution and Soil Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, W.K.; Germino, M.J.; Hancock, T.E.; Johnson, D.M. Another perspective on altitudinal limits of alpine timberlines. Tree Physiol. 2003, 16, 1101–1112. [Google Scholar] [CrossRef]
- Beckage, B.; Osborne, B.; Gavin, G.D.; Pucko, C.; Siccama, T.; Perkins, T. A rapid upward shift of a forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proc. Natl. Acad. Sci. USA 2007, 105, 4197–4202. [Google Scholar] [CrossRef] [PubMed]
- Liang, E.Y.; Wang, Y.F.; Piao, S.L.; Lu, X.M.; Camarero, J.J.; Zhu, H.F.; Zhu, L.P.; Ellison, A.M.; Ciais, P.; Peñuelas, J. Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2016, 113, 4380–4385. [Google Scholar] [CrossRef] [PubMed]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Hoffmann, W.A.; Meinzer, F.C.; Franco, A.C.; Giambelluca, T.; Miralles-Wilhelm, F. Controls on stand transpiration and soil water utilization along a tree density gradient in a neotropical savanna. Agric. For. Meteorol. 2008, 148, 839–849. [Google Scholar] [CrossRef]
- Giambelluca, T.W.; Scholz, F.G.; Bucci, S.J.; Meinzer, F.C.; Hoffmann, G. Evapotranspiration and energy balance of Brazilian savannas with contrasting tree density. Agric. For. Meteorol. 2009, 149, 1365–1376. [Google Scholar] [CrossRef]
- Hamdan, K. Spatial and Temporal Variation of Forest Floor, Throughfall, and Stemflow Properties Associated with Bigleaf Maple in a Mixed Conifer Forest of Coastal British Columbia. Master’s Thesis, Simon Fraser University, Burnaby, BC, Canada, 2012. [Google Scholar]
- Garg, S.; Joshi, R.K.; Garkoti, S.C. Effect of tree canopy on species composition, diversity and biomass of herbaceous vegetation and soil characteristics in semi-arid forests of the Aravalli hills. Arid Land Res. Manag. 2022, 36, 224–242. [Google Scholar] [CrossRef]
- Vasutova, M.; Edwards-Jonasova, M.; Baldrian, P.; Cermak, M.; Cudlin, P. Distinct environmental variables drive the community composition of mycorrhizal and saprotrophic fungi at the alpine treeline ecotone. Fungal Ecol. 2017, 27, 116–124. [Google Scholar] [CrossRef]
- Solly, E.F.; Lindahl, B.D.; Dawes, M.A.; Peter, M.; Souza, R.C.; Rixen, C.; Hagedorn, F. Experimental soil warming shifts the fungal community composition at the alpine treeline. New Phytol. 2017, 215, 766–778. [Google Scholar] [CrossRef]
- Alves, G.S.; Bertini, S.; Barbosa, B.B.; Pimentel, J.P.; Azevedo, L. Fungal endophytes inoculation improves soil nutrient availability, arbuscular mycorrhizal colonization and common bean growth. Rhizosphere 2021, 18, 100330. [Google Scholar] [CrossRef]
- Nejad, R.; Kafi, M.; Jari, S.K.; Mozafari, H.; Motesharezadeh, B. Arbuscular mycorrhizal fungi improve growth, physiological status and nutrients accumulation of Ailanthus altissima seedlings under cadmium pollution and salinity. Russ. J. Plant Physl. 2021, 68, 266–273. [Google Scholar] [CrossRef]
- Adeyemi, N.O.; Atayese, M.O.; Sakariyawo, O.S.; Azeez, J.O.; Adeoye, S. Alleviation of heavy metal stress by arbuscular mycorrhizal symbiosis in Glycine max (L.) grown in copper, lead and zinc contaminated soils. Rhizosphere 2021, 18, 100325. [Google Scholar] [CrossRef]
- Alizadeh, S.; Gharagoz, S.F.; Pourakbar, L.; Moghaddam, S.S.; Omidi, M.J. Arbuscular mycorrhizal fungi alleviate salinity stress and alter phenolic compounds of Moldavian balm. Rhizosphere 2021, 19, 100417. [Google Scholar] [CrossRef]
- Koizumi, T.; Hattori, M.; Nara, K. Ectomycorrhizal fungal communities in alpine relict forests of Pinus pumila on Mt. Norikura, Japan. Mycorrhiza 2018, 28, 129–145. [Google Scholar] [CrossRef]
- Reithmeier, L.; Kernaghan, G. Availability of ectomycorrhizal fungi to black spruce above the present treeline in Eastern Labrador. PLoS ONE 2013, 8, e77527. [Google Scholar] [CrossRef]
- Han, Q.S.; Huang, J.; Long, D.F.; Wang, X.B.; Liu, J.J. Diversity and community structure of ectomycorrhizal fungi associated with Larix chinensis across the alpine treeline ecotone of Taibai Mountain. Mycorrhiza 2017, 27, 487–497. [Google Scholar] [CrossRef]
- Arraiano-Castilho, R.; Bidartondo, M.I.; Niskanen, T.; Clarkson, J.J.; Brunner, I.; Zimmermann, S.; Senn-Irlet, B.; Frey, B.; Peintner, U.; Mrak, T.; et al. Habitat specialisation controls ectomycorrhizal fungi above the treeline in the European Alps. New Phytol. 2020, 229, 2901–2916. [Google Scholar] [CrossRef]
- Hasselquist, N.; Germino, M.J.; McGonigle, T.; Smith, W.K. Variability of Cenococcum colonization and its ecophysiological significance for young conifers at alpine–treeline. New Phytol. 2005, 165, 867–873. [Google Scholar] [CrossRef]
- Mohatt, R.K.; Cripps, L.C.; Lavin, M. Ectomycorrhizal fungi of whitebark pine (a tree in peril) revealed by sporocarps and molecular analysis of mycorrhizae from treeline forests in the Greater Yellowstone Ecosystem. Botany 2008, 86, 14–25. [Google Scholar] [CrossRef]
- Sheng, Y.Y.; Cong, J.; Lu, H.; Yang, K.H.; Yang, L.S.; Wang, M.; Zhang, Y.G. Soil fungal diversity of the timberline ecotone in Shennongjia National Park. Acta Ecol. Sin. 2018, 38, 5322–5330. [Google Scholar]
- Sinsabaugh, R.L.; Gallo, M.E.; Lauber, C.; Waldrop, M.P.; Zak, D.R. Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition. Biogeochemistry 2005, 75, 201–215. [Google Scholar] [CrossRef]
- Tonjer, L.; Thoen, E.; Morgado, L.; Botnen, S.; Mundra, S.; Nybakken, L.; Bryn, A.; Kauserud, H. Fungal community dynamics across a forest-alpine ecotone. Mol. Ecol. 2021, 30, 4926–4938. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.A.; Wu, H.; Wang, Z.R.; Dai, G.H.; Yu, D.P.; Jiang, Y.; Li, H. Soil microbial community structure and ecological function in the tree island at the treeline ecotone of Changbai Mountain. Acta Ecol. Sin. 2021, 41, 9847–9856. [Google Scholar]
- An, H.M.; Xu, A.S. Resource of fungal family Russulaceae in Shergyla subalpine in Tibet. Southwest China J. Agric. Sci. 2016, 5, 1202–1208. [Google Scholar]
- An, H.M.; Xu, A.S. Diversity and distribution of the ectomycorrhizal fungi in the Shergyla Mountain in Xizang. Guizhou Agric. Sci. 2016, 44, 162–165. [Google Scholar]
- Yao, H.F.; Lu, J.; Wang, C.; Zang, M. Prediction model of single tree canopy of Abies georgei var. Smithii forest based on mixed effect method. J. Plateau Agric. 2021, 5, 572–580. [Google Scholar]
- Ren, Y.H.; Zhou, Y.Z.; Hou, L.; Fang, J.P.; Luo, D.Q. Spatial distribution patterns of standing trees at different ages in Abies georgei var. Smithii forests in Sejila Mountain. Acta Ecol. Sin. 2021, 41, 5417–5424. [Google Scholar]
- Holtmeier, F.K. Ecological Aspects of Climatically-caused Timberline Fluctuations. In Mountain Environment in Changing Climates; Beniston, M., Ed.; Routledge: London, UK, 1994; pp. 220–233. [Google Scholar]
- Scuderi, L.A. Late-Holocene upper timberline variation in the southern Sierra Nevada. Nature 1987, 325, 242–244. [Google Scholar] [CrossRef]
- Jacob, M.; Annys, S.; Frankl, A.; De Ridder, M.; Beeckman, H.; Guyassa, E.; Nyssen, J. Tree line dynamics in the tropical African highlands—Identifying drivers and dynamics. J. Veg. Sci. 2015, 26, 9–20. [Google Scholar] [CrossRef]
- Ren, Q.S.; Yang, X.L.; Cui, G.F.; Wang, J.S.; Huang, Y.; Wei, X.H.; Li, Q.L. Smith fir poplllation structure and dynamics in the timberline ecotone of the Sejila Mountain, Tibet, China. Acta Ecol. Sin. 2007, 27, 2669–2677. [Google Scholar] [CrossRef]
- Lu, R.K. Soil Agrochemical Analysis Method; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- State Forestry Administration. Chemical Analysis Methods of Forest Soil Water: LY/T 1275-1999; Standards Press of China: Beijing, China, 1999.
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for Basidiomycetes: Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-coverage ITS primers for the DNA-based identification of ascomycetes and basidiomycetes in environmental samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols a Guide to Methods and Applications; Innis, M.A., Garfield, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, G.J. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Caporaso, J.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F. QIIME allows analysis of high throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S RNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, R.H.; Kõljalg, U. Full UNITE + INSD Dataset for Fungi; Version 10.05.2021; UNITE Community: Tartu, Estonia, 2021. [Google Scholar]
- Edgar, R.C. High-accuracy alignment ensembles enable unbiased assessments of sequence homology and phylogeny. bioRxiv 2022. [CrossRef]
- Nguyen, H.N.; Song, Z.W.; Bates, T.S.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. Funguild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org (accessed on 6 March 2022).
- Chen, H. VennDiagram: Generate High-Resolution Venn and Euler Plots. R Package Version 1.7.1. Available online: https://CRAN.R-project.org/package=VennDiagram (accessed on 6 March 2022).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; SAGE Publications: Thousand Oaks, CA, USA, 2018. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kolde, R. pheatmap: Pretty Heatmaps. R Package Version 1.0.12. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 6 March 2022).
- Oksanen, J.; Blanchet, G.F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package. R Package Version 2.5–7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 6 March 2022).
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using Canoco 5; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Zhu, T.H.; Zhang, J.; Hu, T.X.; Li, X.W.; Liu, Y.G. Mycorrhizae type associated with Eucalyptus in Sichuan and effect of forest density on mycorrhizal fungi. J. Sichuan Agicultural Univ. 2001, 19, 222–224. [Google Scholar]
- Liu, Z.L.; Fang, J.Y.; Piao, S.L. Geographical Distribution of Species in Genera Abies, Picea and Larix in China. Acta Geogr. Sin. 2002, 57, 577–586. [Google Scholar]
- Liu, X.L.; Bau, T.; Wang, X.H. Species diversity of Russula from the Greater and Lesser Hinggan Mountains in Northeast China. Mycosystema 2017, 36, 1355–1368. [Google Scholar]
- Kernaghan, G.; Currah, R.S.; Bayer, R.J. Russulaceous ectomycorrhizae of Abies lasiocarpa and Picea engelmannii. Can. J. Bot. 1997, 75, 1843–1850. [Google Scholar] [CrossRef]
- Gray, L.; Kernaghan, G. Fungal succession during the decomposition of ectomycorrhizal fine roots. Microb. Ecol. 2019, 79, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Eo, J.K.; Eom, A.H. Diversity of ectomycorrhizal fungi of Abies koreana at Mt. Halla. Korean J. Mycol. 2009, 37, 134–138. [Google Scholar]
- Matsuda, Y.; Hijii, N. Ectomycorrhizal fungal communities in an Abies firma forest, with special reference to ectomycorrhizal associations between seedlings and mature trees. Can. J. Bot. 2004, 82, 822–829. [Google Scholar] [CrossRef]
- Suz, L.M.; Barsoum, N.; Benham, S.; Dietrich, H.P.; Fetzer, K.D.; Fischer, R.; García, P.; Gehrman, J.; Kristöfel, F.; Manninger, M.; et al. Environmental drivers of ectomycorrhizal communities in Europe’s temperate oak forests. Mol. Ecol. 2014, 23, 5628–5644. [Google Scholar] [CrossRef]
- Christian, K. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Menkis, A.; Urbina, H.; James, T.Y.; Rosling, A. Archaeorhizomyces borealis sp. nov. and a sequence based classification of related soil fungal species. Fungal Biol. 2014, 118, 943–955. [Google Scholar] [CrossRef]
- Rosling, A.; Cox, F.; Cruz-Martinez, K.; Ihrmark, K.; Grelet, G.A.; Lindahl, B.D.; Menkis, A.; James, T.Y. Archaeorhizomycetes: Unearthing an ancient class of ubiquitous soil fungi. Science 2011, 333, 876–879. [Google Scholar] [CrossRef]
- Sterkenburg, E.; Bahr, A.; Durling, M.B.; Clemmensen, K.E.; Lindahl, B.D. Changes in fungal communities along a boreal forest soil fertility gradient. New Phytol. 2015, 207, 1145–1158. [Google Scholar] [CrossRef]
- Pinto-Figueroa, E.A.; Seddon, E.; Yashiro, E.; Buri, A.; Niculita-Hirzel, H.; van der Meer, J.R.; Guisan, A. Archaeorhizomycetes spatial distribution in soils along wide elevational and environmental gradients reveal co-abundance patterns with other fungal saprobes and potential weathering capacities. Front. Microbiol. 2019, 10, 656. [Google Scholar] [CrossRef]
- Schadt, C.W.; Martin, A.P.; Lipson, D.A.; Schmidt, S.K. Seasonal dynamics of previously unknown fungal lineages in tundra soils. Science 2003, 301, 1359–1361. [Google Scholar] [CrossRef]
- Levy-Booth, D.J.; Giesbrecht, I.J.W.; Kellogg, C.T.E.; Heger, T.J.; D’Amore, D.V.; Keeling, P.J.; Hallam, S.J.; Mohn, W.W. Seasonal and ecohydrological regulation of active microbial populations involved in DOC, CO2, and CH4 fluxes in temperate rainforest soil. ISME J. 2018, 13, 950–963. [Google Scholar] [CrossRef]
- Fernández-Martínez, M.A.; Pérez-Ortega, S.; Pointing, S.B.; Allan Green, T.G.; Pintado, A.; Rozzi, R.; Sancho, L.G.; Ríos, A.D.L. Microbial succession dynamics along glacier forefield chronosequences in Tierra del Fuego (Chile). Polar Biol. 2017, 40, 1939–1957. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Finlay, R.D.; Dahlberg, A.; Stenlid, J.; Wardle, D.A.; Lindahl, B.D. Carbon sequestration is related to mycorrhizal fungal community shifts during long-term succession in boreal forests. New Phytol. 2015, 205, 1525–1536. [Google Scholar] [CrossRef]
- Borg Dahl, M.; Priemé, A.; Brejnrod, A.; Brusvang, P.; Lund, M.; Nymand, J.; Kramshøj, M.; Ro-Poulsen, H.; Haugwitz, M.S. Warming, shading and a moth outbreak reduce tundra carbon sink strength dramatically by changing plant cover and soil microbial activity. Sci. Rep. 2017, 7, 16035. [Google Scholar] [CrossRef]
- Zhang, Q.P.; Fang, R.Y.; Deng, C.Y.; Zhao, H.J.; Shen, M.H.; Wang, Q. Slope aspect effects on plant community characteristics and soil properties of alpine meadows on Eastern Qinghai-Tibetan plateau. Ecol. Indic. 2022, 143, 109400. [Google Scholar] [CrossRef]
- Akhtaruzzaman, M.; Roy, S.; Mahmud, M.S.; Shormin, T. Soil properties under different vegetation types in Chittagong university campus, Bangladesh. J. For. Environ. Sci. 2020, 36, 133–142. [Google Scholar]
- Cheng, F.; Wei, X.; Hou, L.; Shang, Z.C.; Peng, X.B.; Zhao, P.; Fei, Z.X.; Zhang, S.X. Soil fungal communities of montane natural secondary forest types in China. J. Microbiol. 2015, 53, 379–389. [Google Scholar] [CrossRef]
- Zhang, L.P.; Zhang, X.C.; Liu, Z.W.; Sun, Q.; Wang, S.B.; Sun, H.Y.; Yu, W.F.; Liu, Y.J. Effect of plantation litter decomposition on soil properties. J. Northwest A F Univ. 2008, 36, 87–92. [Google Scholar]
- Ek, H.A.; Arnebrant, K.; SÖderstrÖm, B. Growth and assimilation of NH4+ and NO3− by Paxillus involutus in association with Betula pendula and Picea abies as affected by substrate pH. New Phytol. 1994, 128, 629–637. [Google Scholar] [CrossRef]
- Rossi, S.; Bordeleau, A.; Houle, D.; Morin, H. Effect of chronic ammonium nitrate addition on the ectomycorrhizal community in a black spruce stand. Can. J. For. Res. 2012, 42, 1204–1212. [Google Scholar] [CrossRef]
- Termorshuizen, A.J.; Ket, P.C. Effects of ammonium and nitrate on mycorrhizal seedlings of Pinus sylvestris. For. Pathol. 1991, 21, 404–413. [Google Scholar] [CrossRef]
- Marty, C.; Houle, D.; Gagnon, C.; Courchesne, F. The relationships of soil total nitrogen concentrations, pools and C:N ratios with climate, vegetation types and nitrate deposition in temperate and boreal forests of eastern Canada. Catena 2017, 152, 163–172. [Google Scholar] [CrossRef]
- He, P.; Lu, J.; Ren, Y.H.; Li, J.R.; Hou, L.; Deng, X.S.; Gao, T.; Cheng, F. Altitude and slope aspects as the key factors affecting the change of C:N:P stoichiometry in the leaf-litter-soil system of alpine timberline ecotones of the Sygera Mountains in Southeast Tibet, China. Geoderma Reg. 2023, 32, e00602. [Google Scholar] [CrossRef]
- Shen, L.; Ye, S.M.; Liu, H.Y.; Deng, X.S.; He, P.; Cheng, F. Linkage between leaf-litter-soil, microbial resource limitation, and carbon-use efficiency in successive Chinese fir (Cunninghamia lanceolata) plantations. Forests 2023, 14, 357. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Timberline Sites a | Vegetation Zones | Altitude (m) | Slope Degree (°) | Dominant Vegetation |
|---|---|---|---|---|
| South | F | 4280–4287 | 18~28 | S. saltuaria, R. aganniphum, R. nyingchiens |
| FL | 4335–4360 | 11~17 | S. saltuaria, R. aganniphum, R. nyingchiens | |
| TL | 4354–4380 | 10~25 | R. aganniphum, R. nyingchiense, S. saltuaria | |
| TSL | 4384–4390 | 6~18 | R. aganniphum, R. nyingchiense | |
| S | 4390–4416 | 1~7 | R. aganniphum | |
| North | F | 4223–4250 | 18–23 | A. georgei, R. aganniphum, R. nyingchiense |
| FL | 4321–4339 | 8~18 | A. georgei, R. aganniphum, R. nyingchiense | |
| TL | 4353–4370 | 11~15 | R. aganniphum, R. nyingchiense, A. georgei | |
| TSL | 4387–4392 | 13~18 | R. aganniphum, R. nyingchiense | |
| S | 4403–4412 | 12~21 | R. aganniphum |
| Timberline Sites | Vegetation Zones | Vegetation Factors a | ||
|---|---|---|---|---|
| SC (%) | TC (%) | TD (Trees/hm2) | ||
| South | F | 56.67 ± 16.07 b | 65.00 ± 5.00 a | 341.67 ± 160.73 a |
| FL | 81.67 ± 2.89 a | 46.67 ± 7.64 b | 175.00 ± 66.14 b | |
| TL | 85.00 ± 5.00 a | 30.00 ± 5.00 c | 58.33 ± 14.43 bc | |
| TSL | 91.67 ± 5.77 a | 13.33 ± 2.89 d | 41.67 ± 14.43 bc | |
| S | 91.67 ± 5.77 a | 0.00 ± 0.00 e | 0.00 ± 0.00 c | |
| Mean | 81.33 ± 15.17 | 31.00 ± 24.29 | 123.33 ± 144.07 | |
| North | F | 78.33 ± 7.64 b | 65.00 ± 13.23 a | 400.00 ± 125.00 a |
| FL | 80.00 ± 5.00 b | 43.33 ± 5.77 b | 308.33 ± 62.92 a | |
| TL | 86.67 ± 2.89 ab | 36.67 ± 2.89 b | 166.67 ± 62.92 b | |
| TSL | 86.67 ± 7.64 ab | 18.33 ± 2.89 c | 50.00 ± 25.00 bc | |
| S | 95.00 ± 0.00 a | 0.00 ± 0.00 d | 0.00 ± 0.00 c | |
| Mean | 85.33 ± 7.67 | 32.67 ± 23.59 | 185.00 ± 167.12 | |
| FTimberline | 2.36 | 0.61 | 5.21 | |
| PTimberline | 0.14 | 0.44 | 0.03 | |
| FZone | 11.72 | 111.73 | 25.14 | |
| PZone | 0.00 | 0.00 | 0.00 | |
| FTimberline×Zone | 3.18 | 0.73 | 0.96 | |
| PTimberline×Zone | 0.04 | 0.58 | 0.45 | |
| Timberline Sites | Vegetation Zones | Soil Factors a | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SP | SBD | FWHC | pH | SOC | TP | TN | AN | AP | C/N | C/P | N/P | ||
| (%) | (g/cm3) | (%) | (g/kg) | (g/kg) | (g/kg) | (mg/kg) | (mg/kg) | ||||||
| South | F | 60.00 ± 2.91 a | 0.97 ± 0.21 a | 55.44 ± 11.05 b | 5.58 ± 0.02 a | 72.98 ± 17.77 b | 0.75 ± 0.03 ab | 3.40 ± 0.65 a | 47.50 ± 9.66 a | 2.14 ± 0.04 a | 21.27 ± 1.41 c | 97.11 ± 21.52 b | 4.54 ± 0.79 b |
| FL | 98.15 ± 54.11 a | 0.49 ± 0.28 b | 102.55 ± 30.52 ab | 5.36 ± 0.17 a | 104.58 ± 17.03 ab | 0.75 ± 0.10 ab | 5.00 ± 1.40 a | 60.08 ± 1.91 a | 1.79 ± 0.14 a | 21.38 ± 2.55 c | 139.39 ± 6.40 b | 6.61 ± 1.16 ab | |
| TL | 73.16 ± 2.94 a | 0.52 ± 0.24 b | 120.83 ± 62.71 ab | 5.02 ± 0.18 b | 131.09 ± 29.51 ab | 0.81 ± 0.06 a | 4.99 ± 1.17 a | 77.50 ± 20.91 a | 2.74 ± 1.08 a | 26.31 ± 0.28 b | 162.51 ± 40.89 b | 6.19 ± 1.62 b | |
| TSL | 74.62 ± 2.95 a | 0.36 ± 0.15 b | 138.39 ± 31.62 a | 4.90 ± 0.17 b | 138.54 ± 30.23 ab | 0.66 ± 0.05 bc | 5.02 ± 0.99 a | 68.25 ± 12.73 a | 1.97 ± 0.48 a | 27.55 ± 0.68 b | 211.13 ± 40.73 ab | 7.64 ± 1.29 ab | |
| S | 74.33 ± 5.32 a | 0.54 ± 0.06 b | 109.52 ± 4.82 ab | 4.86 ± 0.05 b | 182.22 ± 79.28 a | 0.57 ± 0.06 c | 5.85 ± 2.45 a | 76.75 ± 43.37 a | 1.54 ± 1.57 a | 30.99 ± 0.62 a | 319.26 ± 125.41 a | 10.26 ± 3.86 a | |
| Mean | 76.05 ± 24.26 | 0.58 ± 0.27 | 105.35 ± 41.02 | 4.86 ± 0.31 | 125.88 ± 51.54 | 0.71 ± 0.10 | 4.85 ± 1.49 | 66.02 ± 22.43 | 2.03 ± 0.85 | 25.50 ± 4.04 | 185.88 ± 94.93 | 7.05 ± 2.62 | |
| North | F | 65.43 ± 4.36 a | 0.23 ± 0.10 b | 165.69 ± 87.40 a | 4.26 ± 0.15 b | 139.47 ± 14.82 a | 0.62 ± 0.06 a | 5.20 ± 0.54 a | 127.08 ± 14.13 a | 2.22 ± 0.35 a | 26.83 ± 0.50 d | 223.37 ± 12.49 a | 8.33 ± 0.59 a |
| FL | 67.53 ± 3.71 a | 0.26 ± 0.16 b | 168.39 ± 58.68 a | 4.71 ± 0.19 a | 161.81 ± 96.66 a | 0.60 ± 0.06 a | 5.32 ± 3.12 a | 106.25 ± 46.90 ab | 0.35 ± 0.15 b | 30.26 ± 0.43 ab | 262.84 ± 131.61 a | 8.65 ± 4.23 a | |
| TL | 67.12 ± 7.82 a | 0.53 ± 0.22 a | 96.90 ± 18.80 a | 4.93 ± 0.25 a | 154.84 ± 35.05 a | 0.70 ± 0.06 a | 5.45 ± 1.15 a | 119.75 ± 15.08 a | 0.52 ± 0.23 b | 28.36 ± 0.43 c | 221.52 ± 49.41 a | 7.80 ± 1.61 a | |
| TSL | 73.79 ± 8.50 a | 0.20 ± 0.05 b | 184.94 ± 70.43 a | 4.82 ± 0.14 a | 126.46 ± 8.77 a | 0.65 ± 0.06 a | 4.31 ± 0.23 a | 63.25 ± 6.95 b | 0.50 ± 0.08 b | 29.33 ± 1.04 bc | 194.46 ± 24.18 a | 6.64 ± 0.87 a | |
| S | 63.69 ± 7.35 a | 0.29 ± 0.10 ab | 130.75 ± 34.92 a | 4.89 ± 0.26 a | 151.23 ± 46.05 a | 0.71 ± 0.10 a | 4.92 ± 1.51 a | 70.25 ± 7.65 b | 0.40 ± 0.15 b | 30.72 ± 0.20 a | 209.58 ± 39.71 a | 6.82 ± 1.31 a | |
| Mean | 67.51 ± 6.63 | 0.30 ± 0.17 | 149.33 ± 59.89 | 4.72 ± 0.30 | 151.23 ± 46.05 | 0.66 ± 0.08 | 5.04 ± 1.46 | 97.32 ± 33.33 | 0.80 ± 0.76 | 29.10 ± 1.53 | 222.35 ± 60.89 | 7.65 ± 2.00 | |
| FTimberline | 1.71 | 19.20 | 6.14 | 45.21 | 1.51 | 3.83 | 0.11 | 13.84 | 27.8 | 87.27 | 2.4 | 0.6 | |
| PTimberline | 0.21 | 2.88 | 0.02 | 0.00 | 0.23 | 0.06 | 0.74 | 0.00 | 0.00 | 0.00 | 0.14 | 0.45 | |
| FZone | 1.03 | 3.16 | 1.22 | 1.05 | 1.31 | 2.71 | 0.50 | 1.81 | 3.56 | 36.02 | 2.06 | 0.85 | |
| PZone | 0.41 | 0.04 | 0.33 | 0.40 | 0.30 | 0.06 | 0.74 | 0.17 | 0.02 | 0.00 | 0.12 | 0.51 | |
| FTimberline×Zone | 0.88 | 4.00 | 1.59 | 16.3 | 1.24 | 5.04 | 0.73 | 3.83 | 2.57 | 17.68 | 3.63 | 2.71 | |
| PTimberline×Zone | 0.49 | 0.02 | 0.22 | 0.00 | 0.32 | 0.01 | 0.58 | 0.02 | 0.07 | 0.00 | 0.02 | 0.06 | |
| Timberline Sites | Vegetation Zones | OS | ACE | Chao1 | Shannon | Simpson | Goods_Coverage | PD |
|---|---|---|---|---|---|---|---|---|
| South | F | 744 ± 109 a | 870.534 ± 122.140 a | 831.879 ± 125.155 a | 5.665 ± 0.570 a | 0.943 ± 0.023 a | 0.997 ± 0.001 a | 209.136 ± 33.325 a |
| FL | 665 ± 121 a | 800.677 ± 123.711 a | 764.611 ± 121.938 a | 5.413 ± 0.472 a | 0.943 ± 0.015 a | 0.997 ± 0.001 a | 181.342 ± 30.852 a | |
| TL | 676 ± 204 a | 807.454 ± 230.070 a | 766.311 ± 231.254 a | 5.425 ± 1.257 a | 0.929 ± 0.066 a | 0.997 ± 0.001 a | 186.111 ± 66.182 a | |
| TSL | 657 ± 52 a | 790.033 ± 65.099 a | 759.933 ± 70.602 a | 5.676 ± 0.326 a | 0.953 ± 0.011 a | 0.997 ± 0.001 a | 179.771 ± 14.494 a | |
| S | 714 ± 32 a | 867.160 ± 47.436 a | 823.505 ± 33.026 a | 5.307 ± 0.510 a | 0.922 ± 0.044 a | 0.996 ± 0.001 a | 188.148 ± 18.947 a | |
| Mean | 691 ± 107 | 827.172 ± 118.663 | 789.248 ± 118.057 | 5.497 ± 0.616 | 0.938 ± 0.034 | 0.997 ± 0.001 | 188.902 ± 33.486 | |
| North | F | 603 ± 46 a | 751.066 ± 124.208 a | 718.568 ± 105.106 a | 4.542 ± 0.130 a | 0.869 ± 0.019 a | 0.997 ± 0.001 a | 167.133 ± 27.693 a |
| FL | 586 ± 70 a | 693.782 ± 95.361 a | 660.492 ± 76.691 a | 5.000 ± 0.799 a | 0.916 ± 0.062 a | 0.997 ± 0.001 a | 156.946 ± 31.501 a | |
| TL | 712 ± 153 a | 871.145 ± 173.517 a | 823.665 ± 169.359 a | 4.983 ± 0.976 a | 0.877 ± 0.108 a | 0.997 ± 0.001 a | 190.444 ± 47.168 a | |
| TSL | 609 ± 22 a | 722.291 ± 32.316 a | 694.143 ± 18.849 a | 5.173 ± 0.545 a | 0.922 ± 0.034 a | 0.997 ± 0.000 a | 165.631 ± 7.974 a | |
| S | 655 ± 111 a | 767.896 ± 121.008 a | 739.167 ± 124.540 a | 5.456 ± 0.416 a | 0.941 ± 0.019 a | 0.997 ± 0.000 a | 172.730 ± 35.279 a | |
| Mean | 633 ± 92 | 761.236 ± 118.176 | 727.207 ± 109.594 | 5.031 ± 0.626 | 0.905 ± 0.057 | 0.997 ± 0.001 | 170.577 ± 29.819 | |
| FTimberline | 2.229 | 2.038 | 1.905 | 3.568 | 3.304 | 1.333 | 2.047 | |
| PTimberline | 0.151 | 0.169 | 0.183 | 0.073 | 0.084 | 0.262 | 0.168 | |
| FZone | 0.499 | 0.610 | 0.518 | 0.237 | 0.619 | 0.292 | 0.373 | |
| PZone | 0.736 | 0.660 | 0.723 | 0.914 | 0.654 | 0.880 | 0.825 | |
| FTimberline × Zone | 0.532 | 0.527 | 0.474 | 0.668 | 0.725 | 1.125 | 0.347 | |
| PTimberline × Zone | 0.714 | 0.717 | 0.754 | 0.622 | 0.585 | 0.373 | 0.843 | |
| Factors a | RDA1 | RDA2 | r2 | p |
|---|---|---|---|---|
| SC | 0.143641 | −0.98963 | 0.496922 | 0.001 |
| TC | 0.351797 | 0.936076 | 0.436142 | 0.001 |
| TD | 0.632203 | 0.774803 | 0.357184 | 0.002 |
| TS | 0.966325 | −0.25733 | 0.619121 | 0.001 |
| SBD | −0.61957 | 0.784943 | 0.520066 | 0.001 |
| FWHC | 0.796237 | −0.60498 | 0.319073 | 0.006 |
| pH | −0.88802 | 0.459812 | 0.716007 | 0.001 |
| C/N | 0.531154 | −0.84728 | 0.603805 | 0.001 |
| AN | 0.998002 | −0.06318 | 0.470154 | 0.001 |
| OS | −0.69521 | 0.718802 | 0.236768 | 0.025 |
| Shannon | −0.99978 | 0.020968 | 0.26829 | 0.015 |
| Simpson | −0.99411 | −0.10838 | 0.265634 | 0.016 |
| ACE | −0.62356 | 0.781778 | 0.199583 | 0.045 |
| Chao1 | −0.62749 | 0.778628 | 0.199686 | 0.043 |
| PD | −0.52077 | 0.853696 | 0.27333 | 0.016 |
| Asc | −0.83238 | −0.55421 | 0.727328 | 0.001 |
| Bas | 0.98076 | 0.195217 | 0.808387 | 0.001 |
| Chy | −0.49419 | 0.869353 | 0.54539 | 0.002 |
| Others | −0.99535 | 0.096355 | 0.33698 | 0.005 |
| Sap | −0.96414 | −0.26541 | 0.659629 | 0.001 |
| Sym | 0.99291 | 0.118865 | 0.754456 | 0.001 |
| Soil_Sap | −0.8569 | −0.51549 | 0.548047 | 0.001 |
| Ect | 0.982745 | 0.184964 | 0.835384 | 0.001 |
| Una | −0.99994 | −0.0111 | 0.251452 | 0.022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, F.; Li, M.; Ren, Y.; Hou, L.; Gao, T.; He, P.; Deng, X.; Lu, J. Soil Fungal Community Characteristics at Timberlines of Sejila Mountain in Southeast Tibet, China. J. Fungi 2023, 9, 596. https://doi.org/10.3390/jof9050596
Cheng F, Li M, Ren Y, Hou L, Gao T, He P, Deng X, Lu J. Soil Fungal Community Characteristics at Timberlines of Sejila Mountain in Southeast Tibet, China. Journal of Fungi. 2023; 9(5):596. https://doi.org/10.3390/jof9050596
Chicago/Turabian StyleCheng, Fei, Mingman Li, Yihua Ren, Lei Hou, Tan Gao, Peng He, Xiangsheng Deng, and Jie Lu. 2023. "Soil Fungal Community Characteristics at Timberlines of Sejila Mountain in Southeast Tibet, China" Journal of Fungi 9, no. 5: 596. https://doi.org/10.3390/jof9050596
APA StyleCheng, F., Li, M., Ren, Y., Hou, L., Gao, T., He, P., Deng, X., & Lu, J. (2023). Soil Fungal Community Characteristics at Timberlines of Sejila Mountain in Southeast Tibet, China. Journal of Fungi, 9(5), 596. https://doi.org/10.3390/jof9050596