

Broadening the Knowledge of Mexican Boletes: Addition of a New Genus, Seven New Species, and Three New Combinations

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Morphology Studies
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Alignments, Model Selection, and Phylogenetic Analyses
3. Results
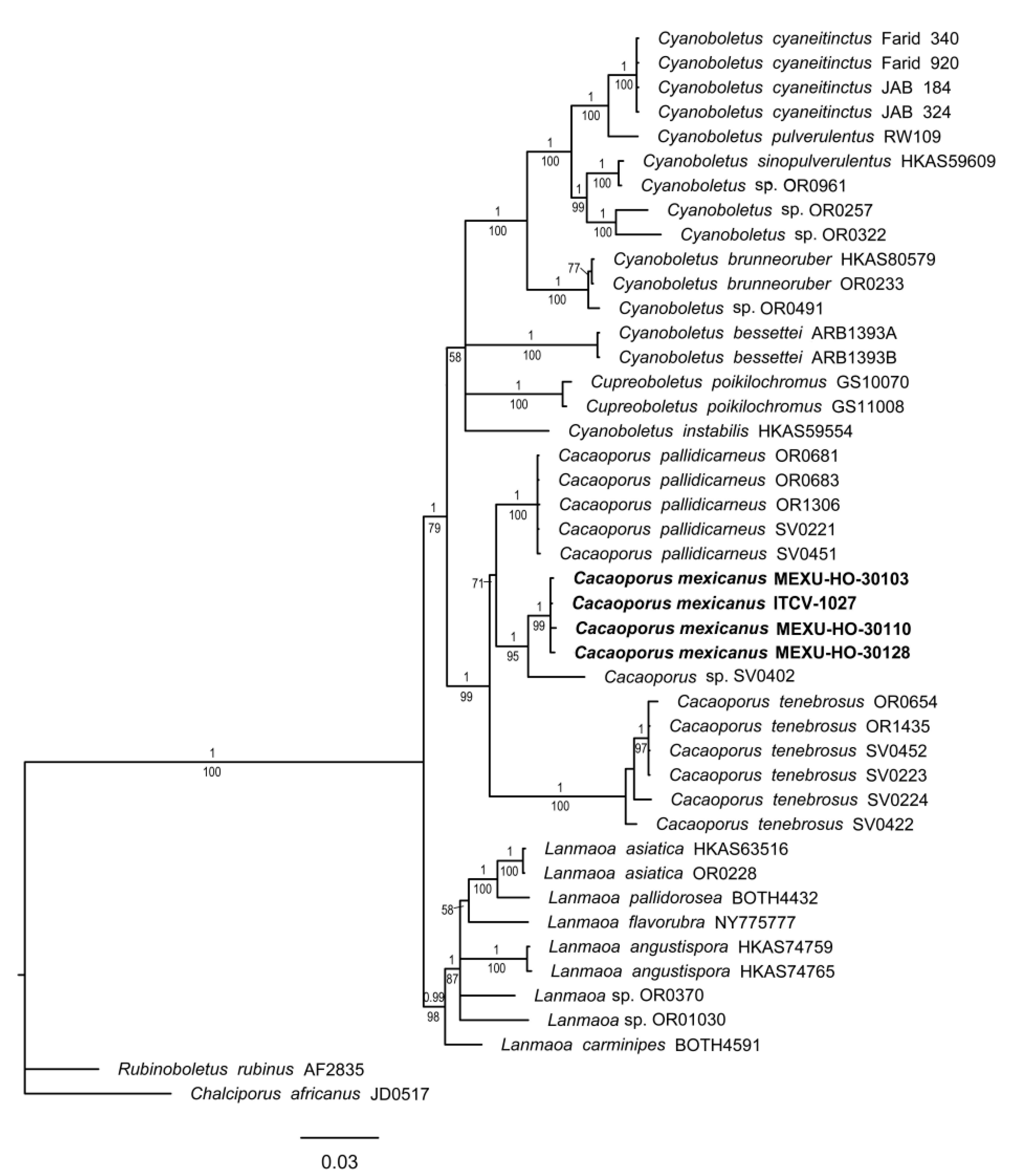
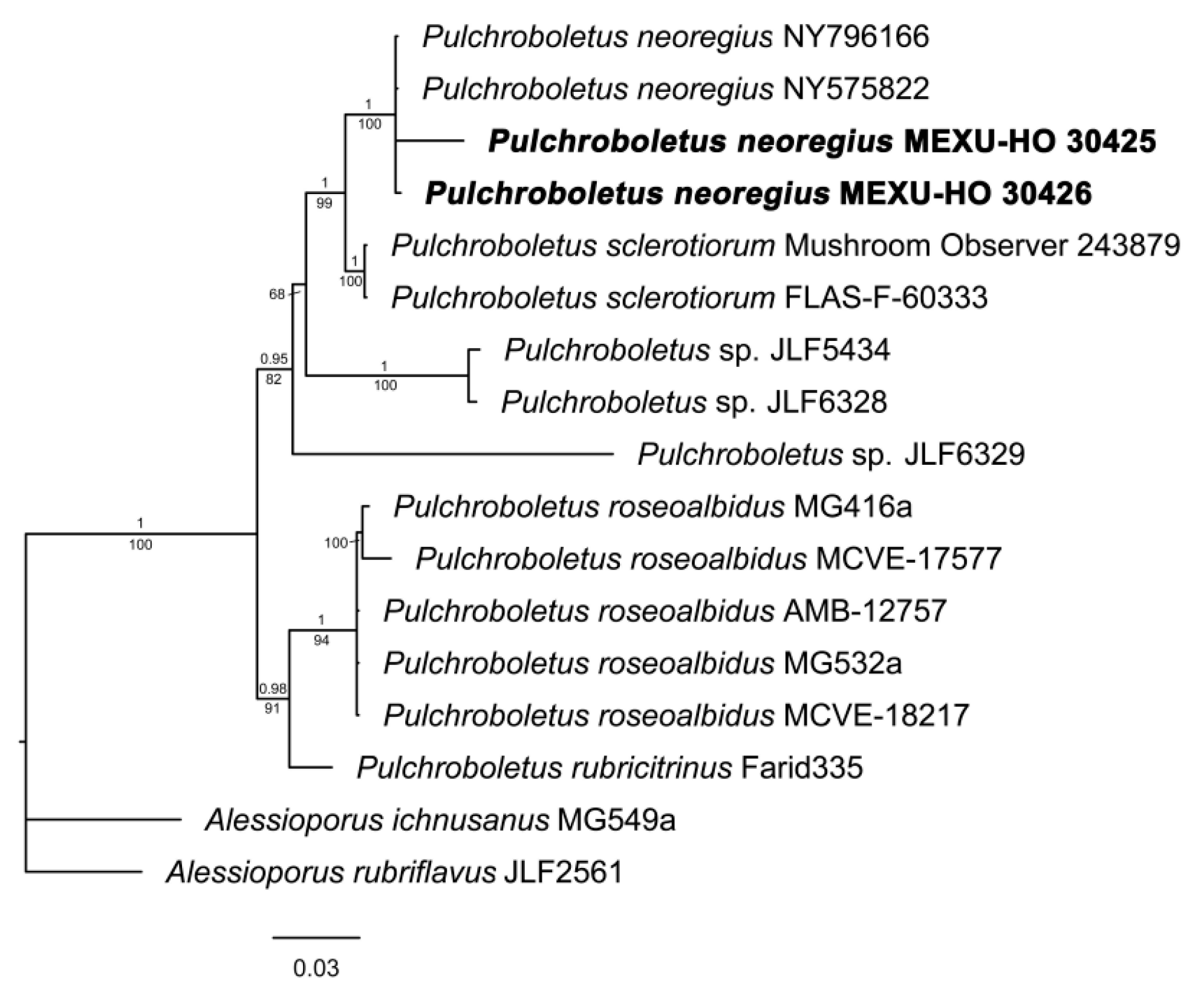
3.1. Phylogenetic Analyses
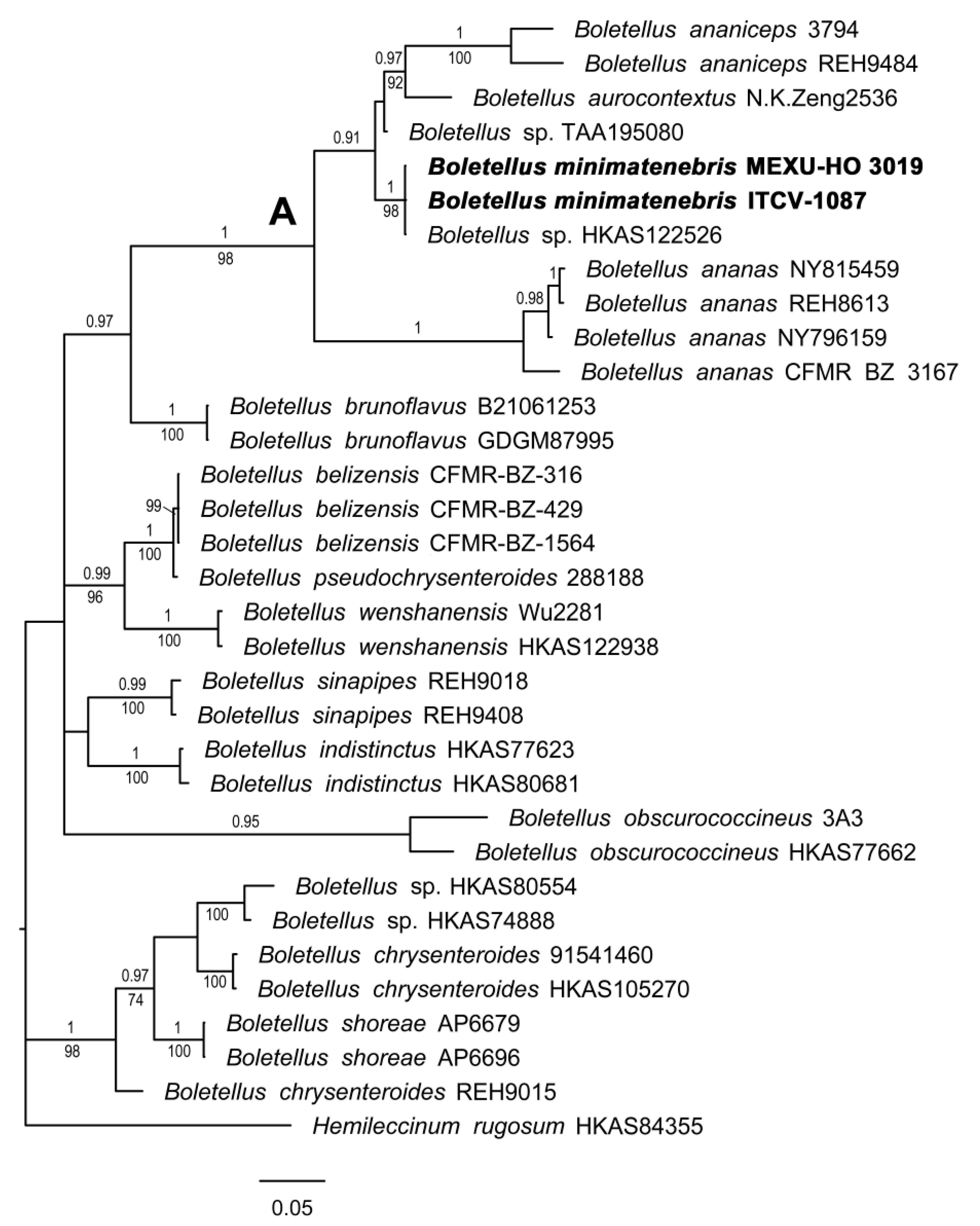
3.1.1. Boletellus Phylogeny
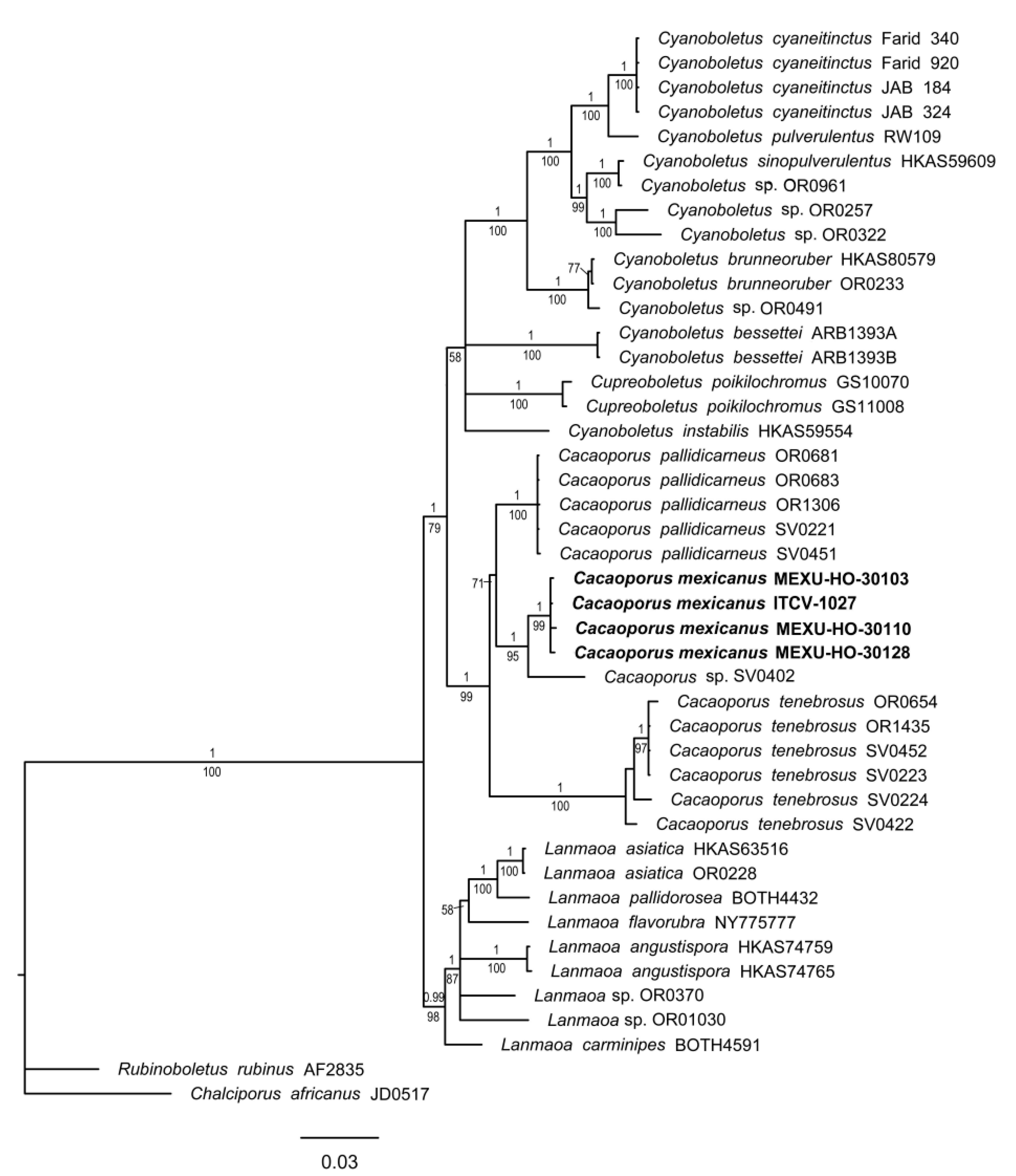
3.1.2. Cacaoporus Phylogeny
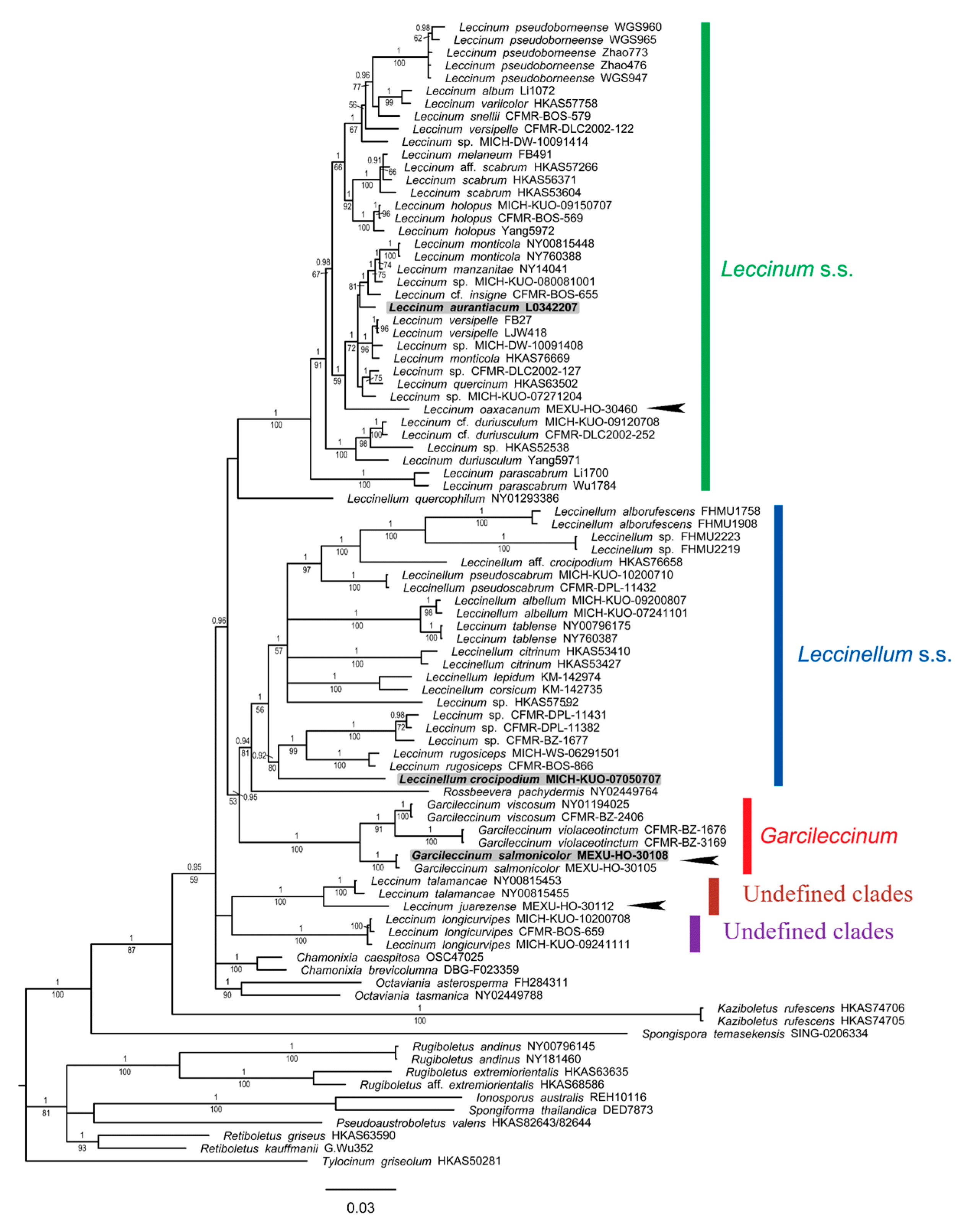
3.1.3. Leccinoideae Subfamily Phylogeny
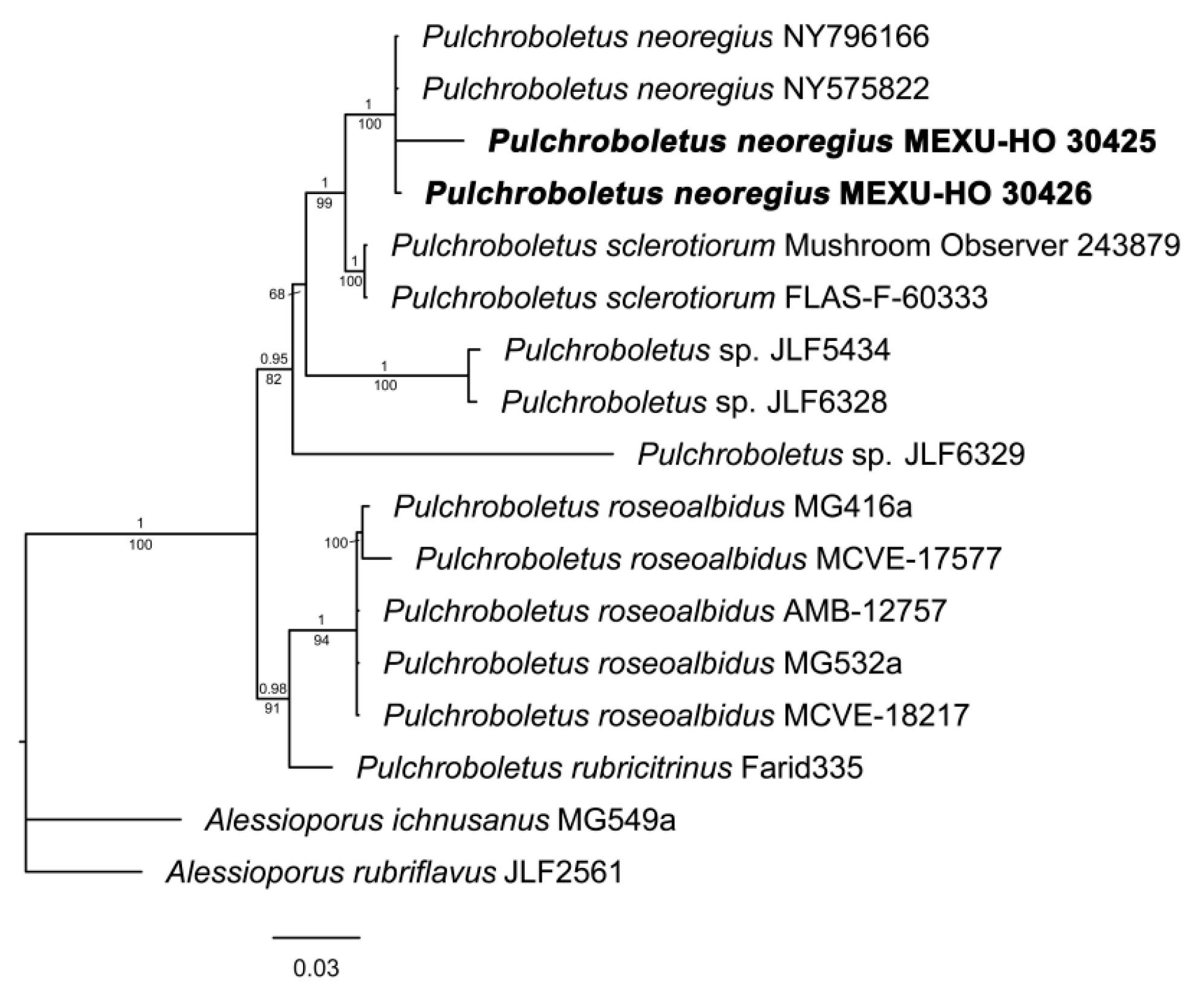
3.1.4. Pulchroboletus
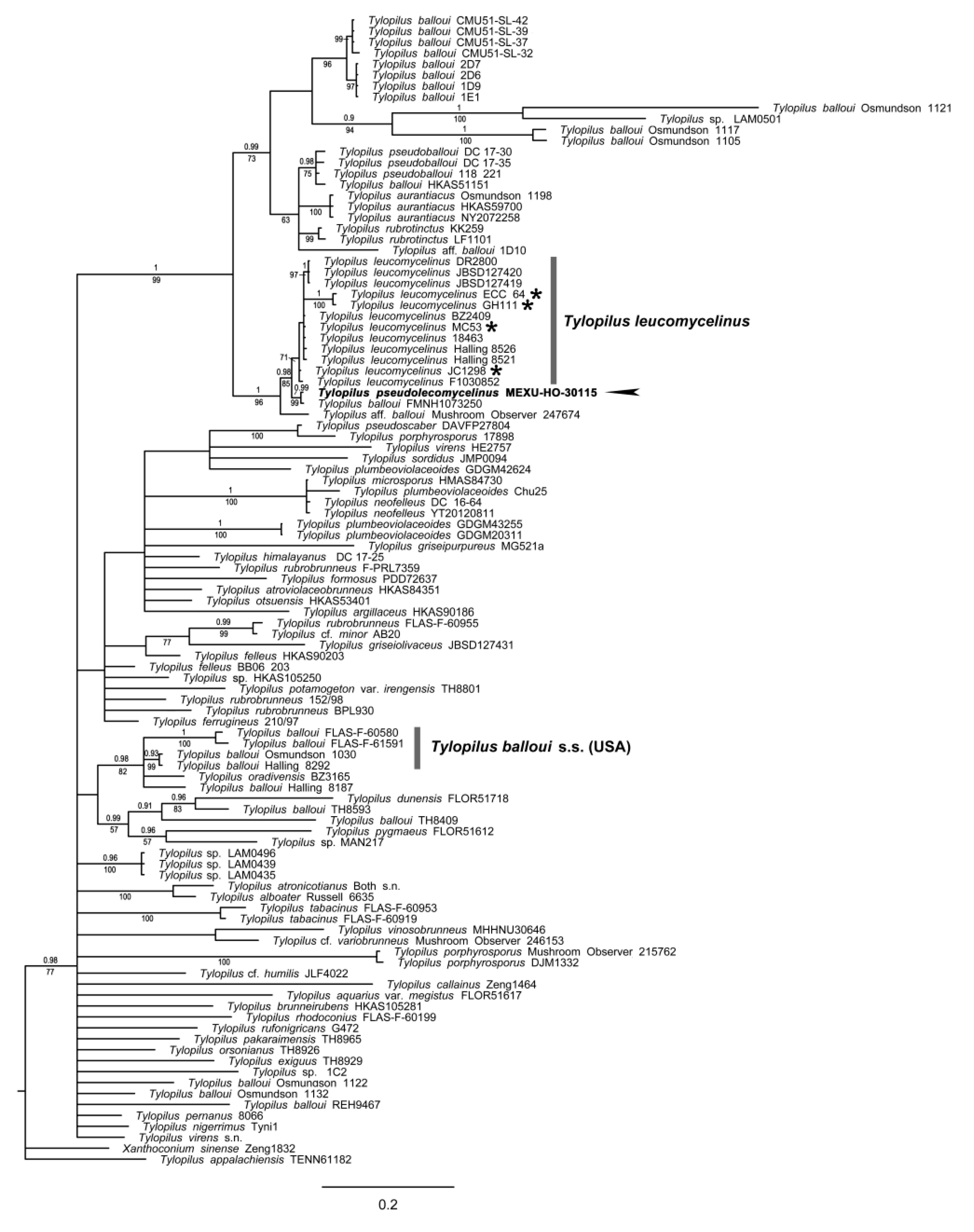
3.1.5. Tylopilus Phylogeny
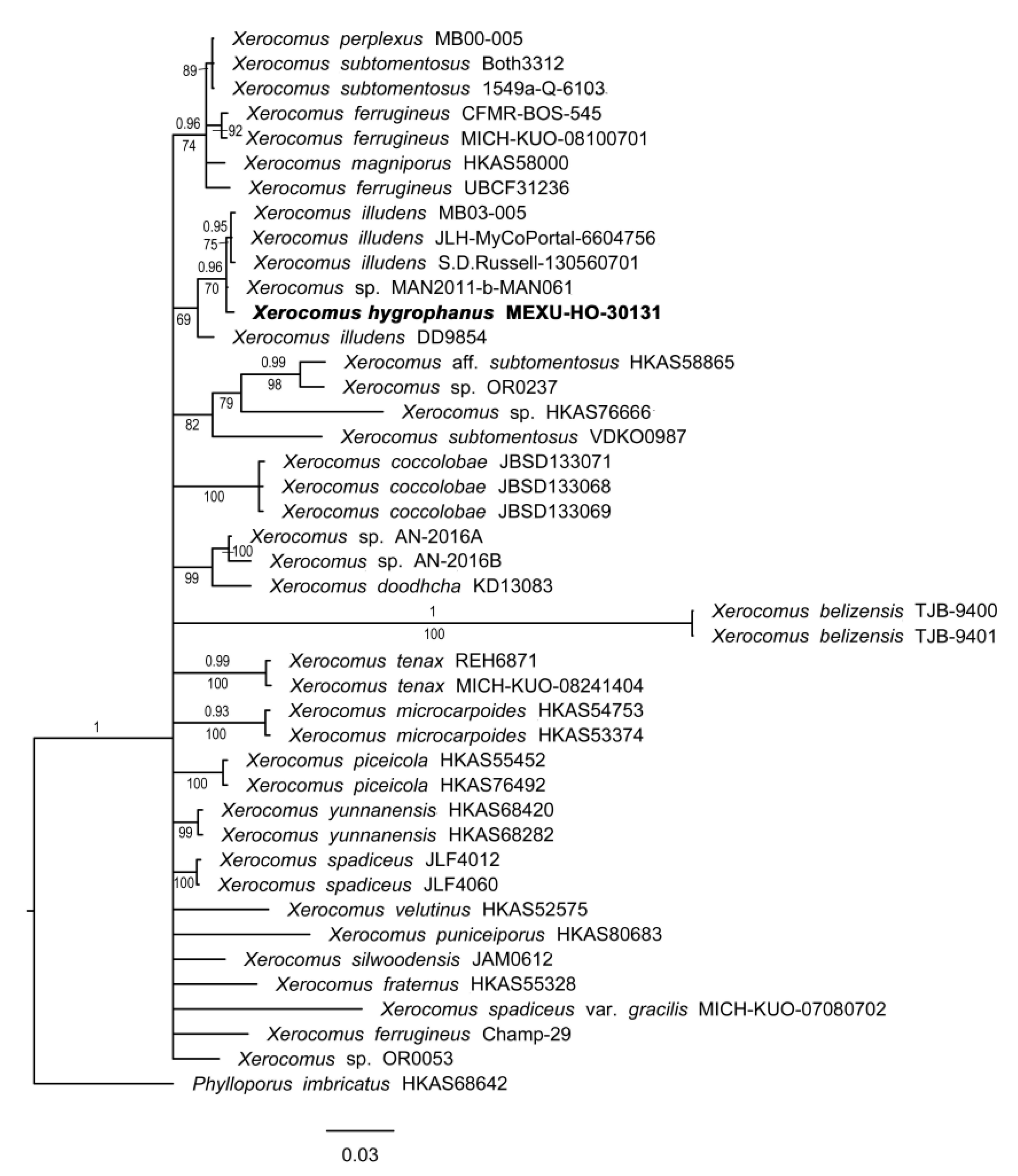
3.1.6. Xerocomus Phylogeny
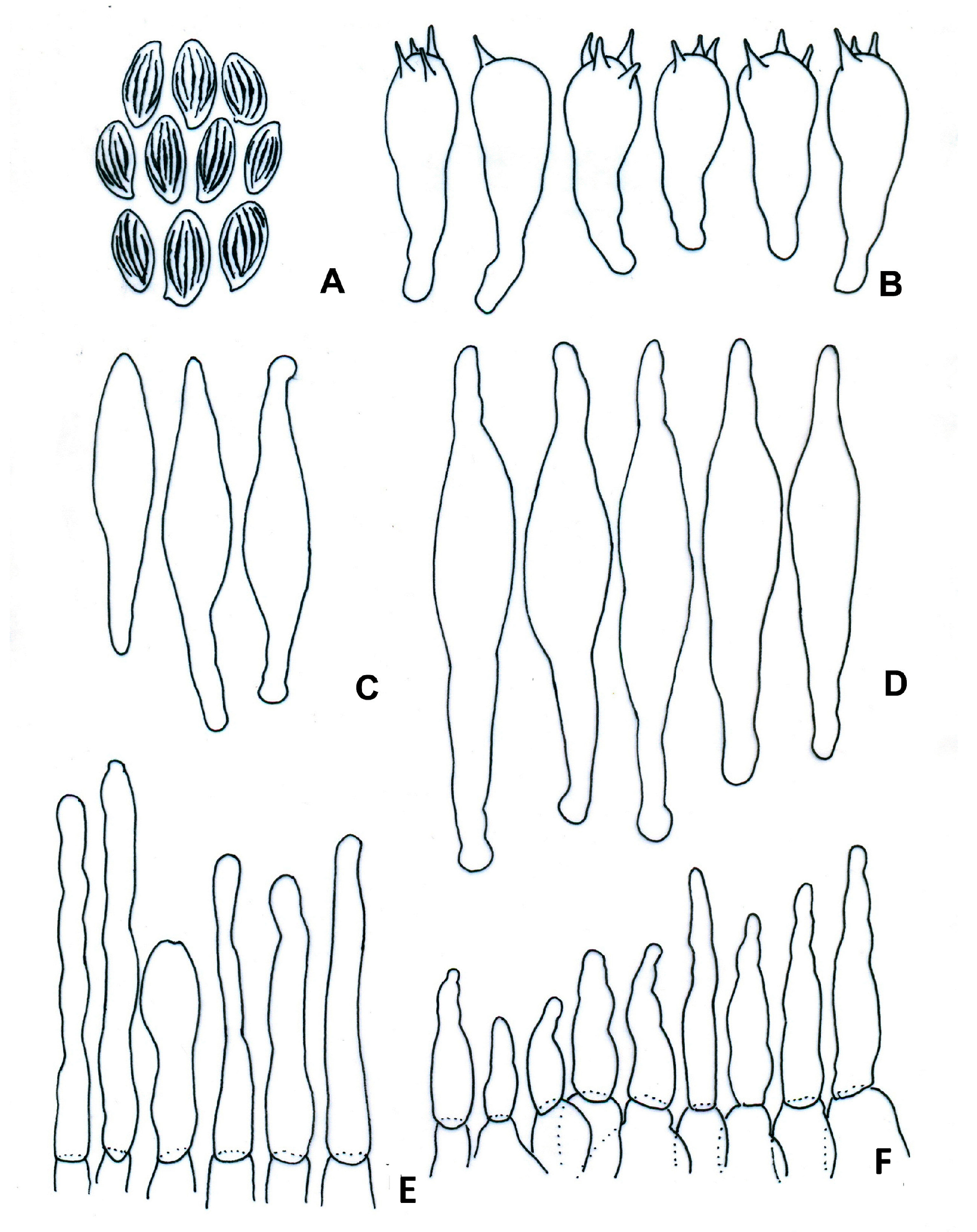
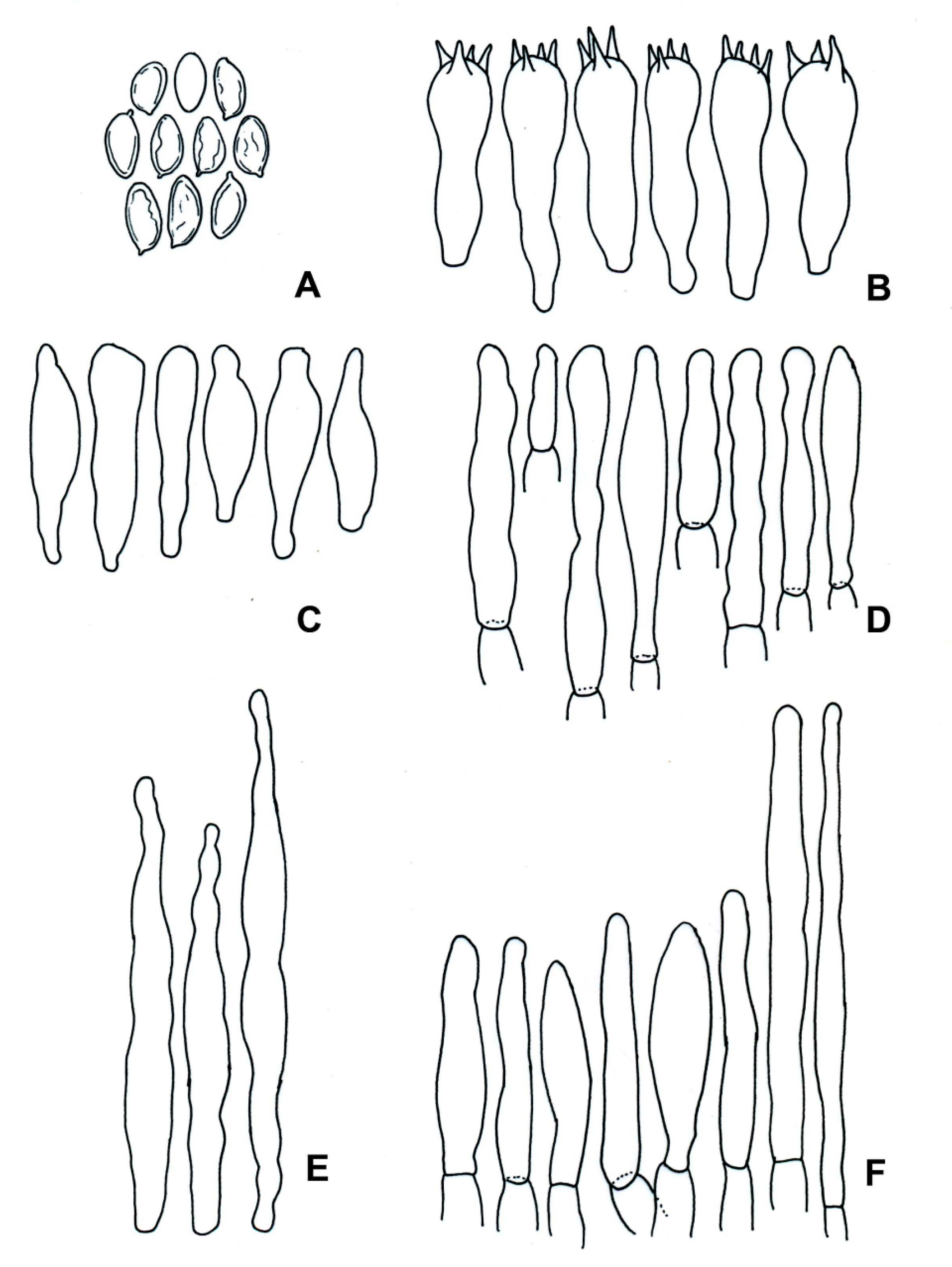
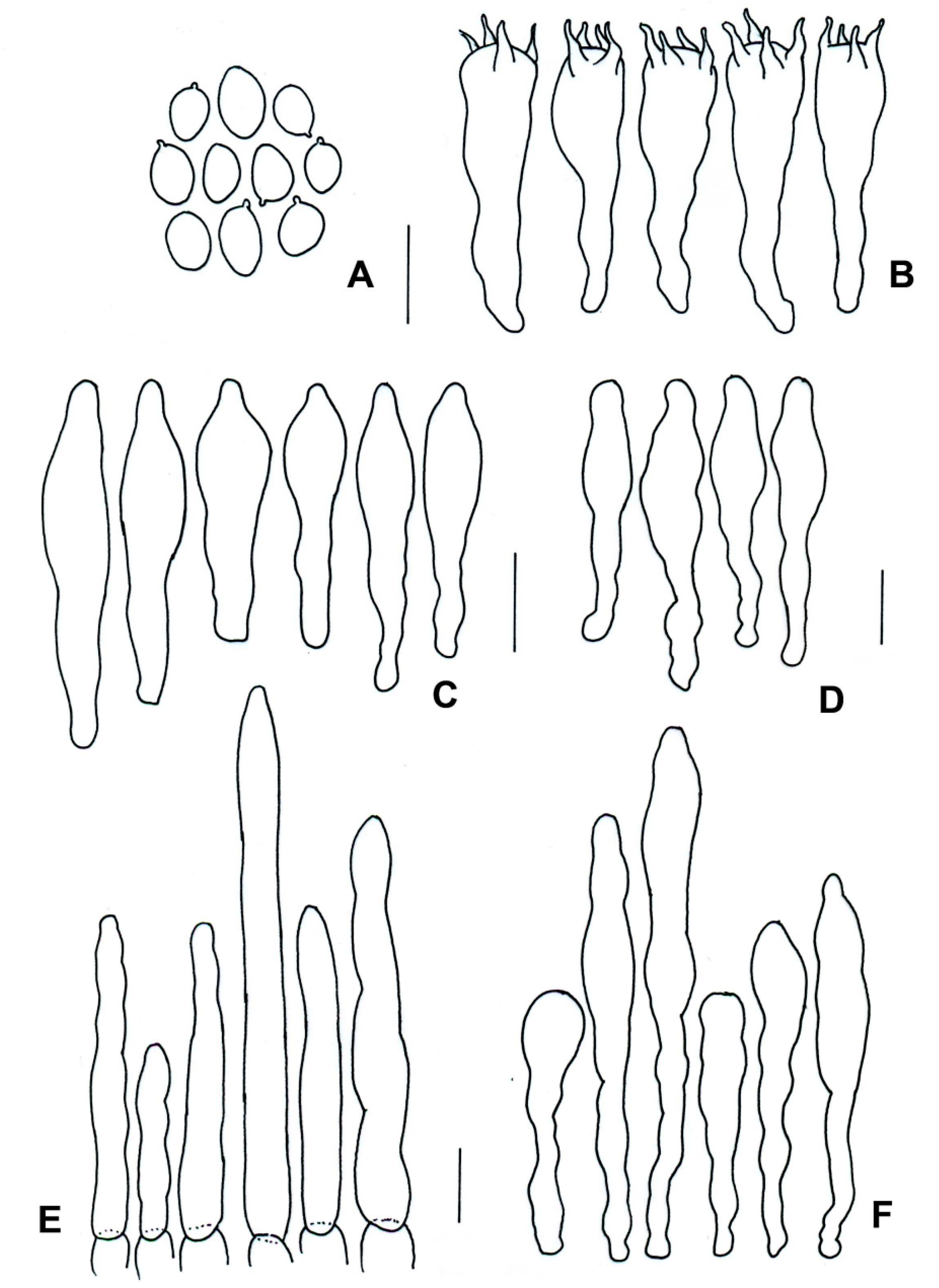
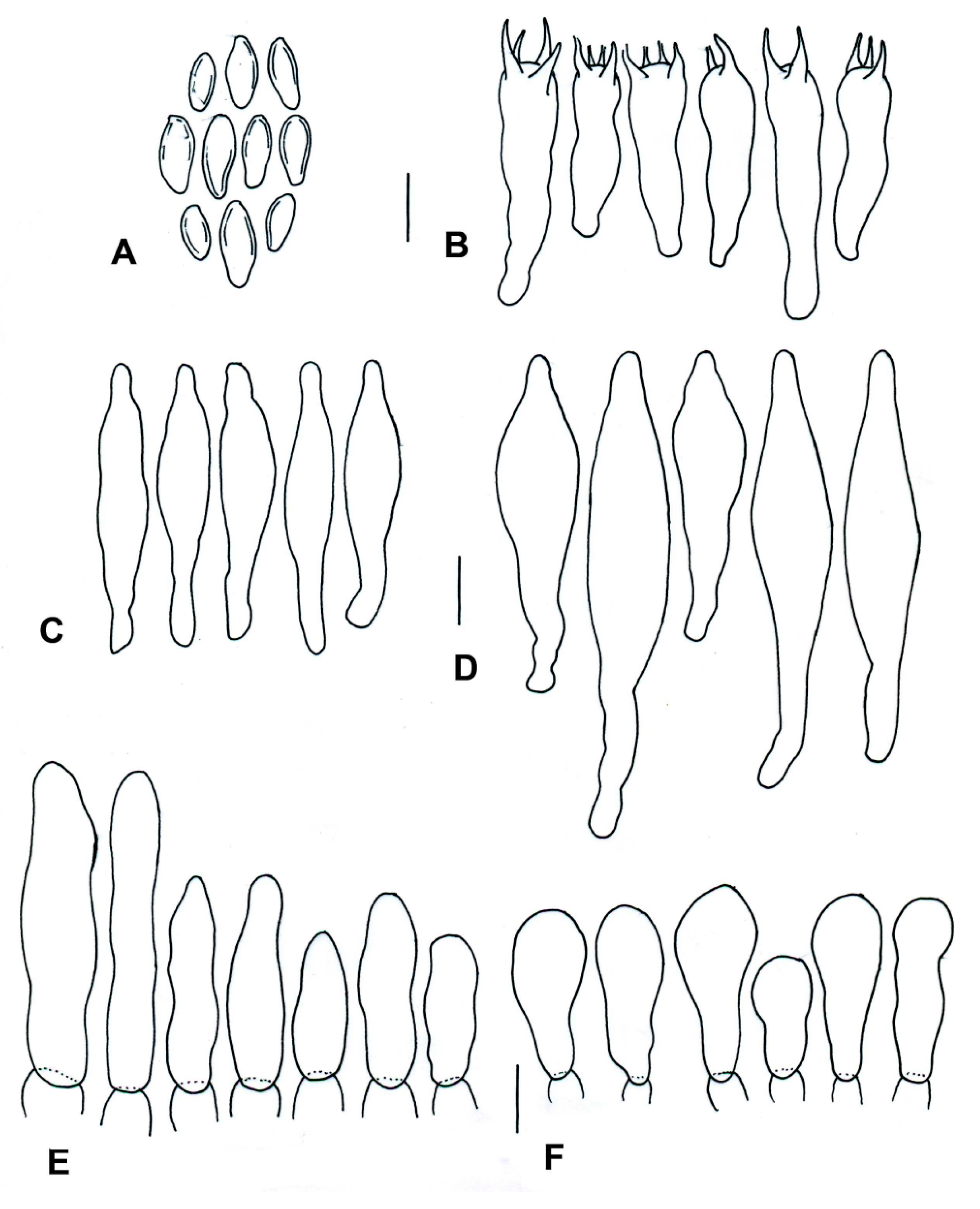
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morrone, J.J. Regionalización biogeográfica y evolución biótica de México: Encrucijada de la biodiversidad del Nuevo Mundo. Rev. Mex. Biodiv. México 2019, 90, e902980. [Google Scholar] [CrossRef]
- Ayala-Vasquez, O. Taxonomía, Biogeografía y Filogenia de Los Hongos del Orden Boletales en la Sierra Norte de Oaxaca, México. Ph.D. Thesis, Instituto Tecnológico de Ciudad Victoria, Tamaulipas, México, 2021. [Google Scholar]
- Guzmán, G. Análisis cualitativo y cuantitativo de la diversidad de los hongos en México. In La Diversidad Biológica de Iberoamérica II; Acta Zoologica Mexicana, Nueva Serie; Halffter, G., Ed.; CYTED, Instituto de Ecología: Veracruz, México, 1998; pp. 111–175. [Google Scholar]
- Guzmán, G. Los hongos de la península de Yucatán II. Nuevas exploraciones y adiciones micológicas. Biotica 1983, 8, 71–87. [Google Scholar]
- De la Fuente, J.I.; García-Jiménez, J.; López, C.Y.; Oros-Ortega, I.; Vela-Hernández, R.Y.; Guevara-Guerrero, G.; Garza-Ocañas, F.; Chay-Casanova, J.A.; Ibarra-Garibay, L.E.; Bandala-Muñoz, V.M. An annotated checklist of the macrofungi (Ascomycota, Basidiomycota, and Glomeromycota) from Quintana Roo, Mexico. Check List 2022, 16, 627–648. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Mueller, G.M. Fungal communities: Their diversity and distribution. In The Fungal Community: Its Organization and Role in the Ecosystem; Dighton, J., White, J.F., Oudemans, P., Eds.; CRC Taylor and Francis: New York, NY, USA, 2005; pp. 27–37. [Google Scholar]
- Pérez-Moreno, J.; Guerin-Laguette, A.; Rinaldi, A.C.; Yu, F.; Verbeken, A.; Hernández-Santiago, F.; Martínez-Reyes, M. Edible mycorrhizal fungi of the world: What is their role in forest sustainability, food security, biocultural conservation and climate change? Plants People Planet 2020, 3, 471–490. [Google Scholar] [CrossRef]
- Pérez-Moreno, J.; Guerin-Laguette, A.; Flores, A.R.; Qiang, F. Mushrooms, Humans and Nature in a Changing World; Springer Nature Switzerland AG: Basel, Switzerland, 2020; p. 473. [Google Scholar] [CrossRef]
- Li, H.; Tian, Y.; Menolli, N., Jr.; Ye, L.; Karunarathna, S.C.; Pérez-Moreno, J.; Mohammad-Mahmudur-Rahman, M.M.; Rashid, M.D.H.; Phengsintham, P.; Rizal, L.; et al. Reviewing the world’s edible mushroom species: A new evidence-based classification system. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1982–2014. Available online: https://ift.onlinelibrary.wiley.com/doi/abs/10.1111/1541-4337.12708 (accessed on 6 July 2023). [CrossRef]
- Pérez-Moreno, J.; Mortimer, P.E.; Xu, J.; Karunarathna, S.C.; Li, H. Global perspectives on the ecological, cultural and socioeconomic relevance of wild edible fungi. Stud. Fungi 2021, 6, 408–424. [Google Scholar] [CrossRef]
- Gilbert, J.E. Les Boletes. In Les Livres du Mycologue; Librairie E. Le François: Paris, France, 1931; Volume 33, p. 254. [Google Scholar]
- Wu, G.; Yan-Chun, L.; Xue-Tai, Z.; Kuan, Z.; Li-Hong, H.; Yang-Yang, C.; Fang, L.; Jian-Ping, X.; Zhu, L.Y. One hundred noteworthy boletes from China. Fungal Divers. 2016, 81, 25–188. [Google Scholar] [CrossRef]
- Li, Y.C.; Yang, Z.L. The Boletes of China: Tylopilus S.L.; Springer: Singapore, 2021; pp. 259–360. [Google Scholar] [CrossRef]
- Orihara, T.; Healy, R.; Corrales, A.; Smith, M.E. Multilocus phylogenies reveal three new truffle-like taxa and the traces of interspecific hybridization in Octaviania (Boletaceae, Boletales). IMA Fungus 2021, 12, 12–14. [Google Scholar] [CrossRef]
- Hosen, M.d.I.; Yang, Z.L. Kaziboletus, a new boletoid genus of Boletaceae associated with Shorea robusta in Bangladesh. Mycol Prog 2021, 20, 1145–1156. [Google Scholar] [CrossRef]
- Kuo, M.; Ortiz-Santana, B. Revision of leccinoid fungi, with emphasis on North American taxa, based on molecular and morphological data. Mycologia 2020, 112, 197–211. [Google Scholar] [CrossRef]
- Ortiz-Santana, B.D.; Halling, E.R. A new species of Leccinum (Basidiomycota, Boletes) from Belize. Brittonia 2009, 61, 172–174. [Google Scholar] [CrossRef]
- Ortiz-Santana, B.D.; Lodge, D.J.; Baroni, T.J.; Both, E.E. Boletes from Belize and the Dominican Republic. Fungal Diver. 2007, 27, 247–416. [Google Scholar]
- Newman, E.I.; Reddell, P. The distribution of mycorrhizas among families of vascular plants. New Phytol. 1987, 106, 745–751. [Google Scholar] [CrossRef]
- Binder, M.; Hibbett, D.S. Molecular systematics and biological diversification of Boletales. Mycologia 2006, 98, 971–983. [Google Scholar] [CrossRef]
- Osmundson, T.W.; Halling, R.E.; den Bakker, H.C. Morphological and molecular evidence supporting an arbutoid mycorrhizal relationship in the Costa Rican paramo. Mycorrhiza 2007, 17, 217–222. [Google Scholar] [CrossRef]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhizal 2010, 20, 217–263. [Google Scholar] [CrossRef]
- Halling, R.E.; Mueller, G.M. Agarics and boletes of Neotropical oakwoods. In Tropical Mycology, Macromycetes; Watling, R., Frankland, J.C., Ainsworth, A.M., Isaac, S., Robinson, C.H., Eds.; CABI Publishing: Wallingford, UK, 2001; Volume 1, pp. 1–10. [Google Scholar] [CrossRef]
- Halling, R.E.; Osmundson, T.W.; Neves, M.A. Pacific boletes: Implications for biogeographic relationships. Mycol. Res. 2008, 112, 437–447. [Google Scholar] [CrossRef]
- Halling, R.E.; Mueller, G.M. Common Mushrooms of the Talamanca Mountains, Costa Rica; New York Botanical Garden Press: New York, NY, USA, 2005; pp. 169–173. [Google Scholar]
- Gelardi, M.; Angelini, C.; Costanzo, F.; Dovana, F.; Ortiz-Santana, B.; Vizzini, A. Tylopilus griseiolivaceus sp. nov. and T. leucomycelinus (Boletaceae) revisited from the Dominican Republic within a comprehensive phylogeny of Tylopilus s. str. Mycol. Prog. 2019, 18, 1039–1056. [Google Scholar] [CrossRef]
- Gelardi, M.; Angelini, C.; Biketova, A.Y.; Suz, L.M.; Ercole, E.; Svetasheva, Y.T.; Miller, K.O.; de la Fuente, J.I.; García Jiménez, J.; Vizzini, A. Coccoloba-associated xerocomoid boletes (Boletaceae) from the Caribbean and Mexico: Tropicoboletus ruborculus gen. et comb. nov., revision of Xerocomus coccolobae, phylogenetic assessment of Singerocomus guadelupae comb. nov., and type studies of Xerocomus caeruleonigrescens, X. cuneipes, and X. pseudoboletinus var. pini-caribaeae. Mycol. Prog. 2023, 22, 22–29. [Google Scholar] [CrossRef]
- Halling, R.E. New Leccinum Species from Costa Rica. Kew Bull. 1999, 54, 747–753. [Google Scholar] [CrossRef]
- Halling, R.E.; Mueller, G.M. New Boletes from Costa Rica. Mycologia 1999, 91, 893–899. [Google Scholar] [CrossRef]
- Halling, R.E. Ectomycorrhizae: Co-evolution, significance and biogeography. Ann. Mo. Bot. Gard. 2001, 88, 5–13. Available online: http://biostor.org/reference/13128 (accessed on 25 April 2023). [CrossRef]
- Halling, R.E.; Mueller, G.M. Leccinum (Boletaceae) in Costa Rica. Mycologia 2003, 95, 488–499. [Google Scholar] [CrossRef]
- Halling, R.E.; Fechner, N.; Nuhn, M.; Osmundson, T.; Soytong, K.; Arora, D.; Binder, M.; Hibbett, D. Evolutionary relationships of Heimioporus and Boletellus (Boletales) with an emphasis on Australian taxa including new species and new combinations in Aureoboletus, Hemileccinum, and Xerocomus. Aust. Syst. Bot. 2015, 28, 1–22. [Google Scholar] [CrossRef]
- Magnago, A.C.; Neves, M.A.; Borges da Silveira, R.M. Fistulinella ruschii, sp. nov., and a new record of Fistulinella campinaranae var. scrobiculata for the Atlantic Forest, Brazil. Mycologia 2017, 109, 1003–1013. [Google Scholar] [CrossRef]
- Magnago, A.C.; Alves-Silva, G.; Terry, T.W.; Borges da Silveira, R.M. New genera, species, and combinations of Boletaceae from Brazil and Guyana. Mycologia 2022, 114, 607–625. [Google Scholar] [CrossRef]
- Neves, M.A.; Binder, M.; Halling, R.; Hibbett, D.; Soytong, K. The phylogeny of selected Phylloporus species, inferred from NUC-LSU and ITS sequences, and descriptions of new species from the Old World. Fungal Divers. 2012, 55, 109–123. [Google Scholar] [CrossRef]
- Singer, R.; García-Jiménez, J.; Gómez, L.D. The Boletineae of México and Central America III. Nova Hedwig. Beih. 1991, 102, 1–99. [Google Scholar]
- Singer, R.; García-Jiménez, J.; Gómez, L.D. The Boletineae of México and Central America. América IV. Nova Hedwig. Beih. 1992, 105, 1–62. [Google Scholar]
- Heim, R.; Perreau, J. Les generes Porphyrellus et Strobilomyces au Mexique. Bull. Soc. Mycol. Fr. 1964, 80, 88–101. [Google Scholar]
- Ayala-Vázquez, O.; Valenzuela, R.; Aguirre-Acosta, E.; Raymundo, T.; García-Jiménez, J. Species of Boletaceae (Boletales, Basidiomycota) with ornamented spores from temperate forest at the state of Oaxaca, Mexico. Stud. Fungi 2018, 3, 271–292. [Google Scholar] [CrossRef]
- Ayala-Vásquez, O.; García-Jiménez, J.; Aguirre-Acosta, C.E.; Castro-Rivera, R.; Angeles-Argaiz, R.E.; Saldivar, A.E.; Garibay-Orijel, R. Hemiaustroboletus: A new genus in the subfamily Austroboletoideae (Boletaceae, Boletales). Mycokeys 2022, 88, 55–78. [Google Scholar] [CrossRef]
- García-Jiménez, J.; Castillo, J. Las especies de Boletáceos y Gomfidiaceos conocidos en el Estado de Nuevo León. Bol. Soc. Mic. 1981, 15, 121–197. [Google Scholar]
- García-Jiménez, J.; Garza-Ocañas, F. Hongos ectomicorrícicos en el estado de Nuevo León. Rev Mex Micol 1986, 2, 197–205. [Google Scholar]
- García-Jiménez, J.; Garza Ocañas, F. Conocimiento de los hongos de la familia Boletaceae de México. Ciencia UANL 2001, 4, 336–343. [Google Scholar]
- García-Jiménez, J.; Singer, R.; Estrada, E.; Garza-Ocañas, F.; Valenzuela, R. Dos especies nuevas del género Boletus (Boletales: Agaricomycetes) en México. Rev. Mex. Biodivers. 2013, 84, 152–162. [Google Scholar] [CrossRef]
- García-Jiménez, J.; Garza-Ocañas, F.; De la Fuente, J.I.; Saldivar, A.E.; Ayala-Vázquez, O. Three new records of Aureoboletus Pouzar (Boletaceae, Boletales) from Mexico. Check List 2019, 15, 759–765. [Google Scholar] [CrossRef]
- González-Velázquez, A.; Valenzuela, R. Los Boletáceos y Gonfidiáceos del Estado de México I. Rev. Mex. Micol. 1993, 9, 35–46. [Google Scholar]
- González-Velázquez, A.; Valenzuela, R. Boletáceos y Gonfidiáceos del Estado de México II. An. Esc. Nac. Cienc. Biol. 1995, 41, 119–196. [Google Scholar]
- Martínez-Reyes, M.; Carrera-Martínez, A.; de la Fuente, J.I.; Ríos-García, U.; Ortiz-López, I.; Ayala-Vásquez, O. Xerocomellus perezmorenoi (Boletaceae, Boletales), a new edible species from Mexico. Phytotaxa 2023, 584, 104–114. [Google Scholar] [CrossRef]
- Montoya, L.; Bandala, V.M. Studies on the genus Phylloporus in Mexico I. Discussion of the known species and description of a new species and a new record. Mycotaxon 1991, 41, 471–482. [Google Scholar]
- Montoya, L.; Bandala, V.M. A new Phylloporus from two relict Fagus grandifolia var. mexicana populations in montane cloud forest. Mycotaxon 2011, 117, 9–18. [Google Scholar]
- Montoya, L.; Garay-Serrano, E.; Bandala, V. Two new species of Phylloporus (Fungi, Boletales) from tropical Quercus forests in eastern Mexico. MycoKeys 2019, 51, 107–123. [Google Scholar] [CrossRef]
- Fuentes García, O.; Ayala-Vásquez; Martínez-Reyes, M.; Martínez-González, C.R.; Díaz-Aguilar, I.; Pérez-Moreno, J. Xerocomellus piedracanteadensis: A new edible species and first record of X. bolinii (Boletaceae, Boletales) from Mexico. Phytotaxa 2023. accepted. [Google Scholar]
- Rodríguez, E.C.; Martínez-González, C.; González, A.; Luna-Vega, I. Tylopilus hayatae, a new endemic bolete species in relict Mexican beech forest. Phytotaxa 2020, 441, 35–46. [Google Scholar] [CrossRef]
- Saldivar, Á.E.; García Jiménez, J.; Herrera-Fonseca, M.J.; Rodríguez-Alcántar, O. Updated checklist and new records of Boletaceae (Fungi, Basidiomycota, Boletales) from Jalisco, Mexico. Polibotanica 2021, 52, 25–49. [Google Scholar] [CrossRef]
- De la Fuente, J.I.; Ayala-Vásquez, O.; Garza-Ocañas, F.; López, C.Y.; García-Jiménez, J. Some interesting Boletales (Basidiomycota) from the state of Quintana Roo, México. Scientia Fungorum 2018, 48, 77–86. [Google Scholar] [CrossRef]
- Guzmán, G. Inventorying the fungi of Mexico. Biodivers. Conserv. 1998, 7, 369–384. [Google Scholar] [CrossRef]
- González-Chicas, E.; Capello, S.; Cifuentes, J.; Torres-De la Cruz, M. New records of Boletales (Basidiomycota) in a tropical oak forest from Mexican Southeast. Bot. Sci. 2019, 97, 423–432. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. 2022—Outline of Fungi and fungus-like taxa. Mycosphere 2021, 13, 53–453. [Google Scholar] [CrossRef]
- Valencia, S. Diversidad del género Quercus (Fagaceae) en México. Bol. Soc. Bot. Mex. 2004, 75, 33–53. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre: London, UK, 1978; p. 252. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenies. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 135–322. [Google Scholar]
- Wu, G.; Feng, B.; Xu, J.; Zhu, X.T.; Li, Y.C.; Zeng, N.K.; Hosen, M.I.; Yang, Z.L. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 2014, 69, 93–115. [Google Scholar] [CrossRef]
- Matheny, P.B.; Wang, Z.; Binder, M. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phyl. Evol. 2007, 43, 430–451. [Google Scholar] [CrossRef]
- Giachini, A.J.; Hosaka, K.; Nouhra, E.; Spatafora, J.; Trappe, J.M. Phylogenetic relationships of the Gomphales based on nuc-25S-rDNA, mit-12S-rDNA, and mit-atp6-DNA combined sequences. Fungal Biol. 2010, 114, 224–234. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Vadthanarat, S.; Lumyong, S.; Raspé, O. Cacaoporus, a new Boletaceae genus, with two new species from Thailand. MycoKeys 2019, 54, 1–29. [Google Scholar] [CrossRef]
- Das, K.; Ghosh, A.; Chakraborty, D.; Datta, S.; Bera, I.; Layola, R., Mr.; Banu, F.; Vizzini, A.; Wisitrassameewong, K. Four Novel Species and Two New Records of Boletes from India. J. Fungi 2023, 9, 754. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Darriba, D.; Taboada, G.I.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 529–542. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 6 August 2023. [Google Scholar]
- Wang, R.; Herrera, M.; Xu, W.; Zhang, P.; Moreno, J.P.; Colinas, C.; Yu, F. Ethnomycological study on wild mushrooms in Pu’er prefecture, Southwest Yunnan, China. J. Ethnobiol. Ethnomed. 2022, 18, 55. [Google Scholar] [CrossRef]
- Thiers, H.D. California Boletes, I.V. The genus Leccinum. Mycologia 1971, 63, 261–276. [Google Scholar] [CrossRef]
- Kuo, M.; Methven, A.S.; Minnis, A.M.; Halling, R.E. Studies of North American macrofungi, 1. Validation of Lactarius rubidus comb. nov. and Leccinellum quercophilum sp. nov. Mycotaxon 2013, 124, 323–332. [Google Scholar] [CrossRef]
- Singer, R.I.; Araujo, I.; Ivory, M.H. Ectotrophically Mycorrhizal Fungi of the Neotropical Lowlands, Especially Central Amazonia. Nova Hedwig. 1983, 77, 1–352. [Google Scholar]
- Flores, R.; Simonini, G. Contributo alla conoscenza delle Boletales del Guatemala. Riv. Micol. 2000, 43, 121–145. [Google Scholar]
- Singer, R. The Boletoideae of Florida. The Boletineae of Florida with Notes on extralimital Species. III. Amer. Midi. Nat. 1947, 37, 1–135. [Google Scholar] [CrossRef]
- Thiers, H.D. The genus Xerocomus Quélet in northern California. Madroño 1964, 17, 237–249. [Google Scholar]
- Singer, R. The Boletineae of Florida with Notes on Extralimital Species. II. The Boletaceae (Gyroporoideae). Farlowia 1945, 2, 223–303. [Google Scholar]
- Haelewaters, D.; Dima, B.; Abdel-Hafiz, B.I.I.; Abdel-Wahab, M.A.; Abul-Ezz, S.R.; Acar, I.; Aguirre-Acosta, E.; Aime, M.C.; Al demir, S.; Ali, M.; et al. Fungal Systematics and Evolution 6. Sydowia 2020, 72, 271–296. [Google Scholar]
- Index Fungorum. Available online: http://www.indexfungorum.org (accessed on 8 August 2023).
- Gelardi, M.; Simonini, G.; Ercole, E.; Vizzini, A. Alessioporus and Pulchroboletus (Boletaceae, Boletineae), two novel genera for Xerocomus ichnusanus and X. roseoalbidusfrom the European Mediterranean basin: Molecular and morphological evidence. Mycology 2014, 106, 1168–1187. [Google Scholar] [CrossRef]
- Chakraborty, D.; Vizzini, A.; Das, K. Two new species and one new record of the genus Tylopilus (Boletaceae) from Indian Himalaya with morphological details and phylogenetic estimations. MycoKeys 2018, 33, 103–124. [Google Scholar] [CrossRef]
- Mueller, G.M.; Strack, B.A. Evidence for a mycorrhizal host shift during migration of Laccaria trichodermophora and other agarics into neotropical oak forests. Mycotaxon 1992, 45, 249–256. [Google Scholar]
- Willis, K.J.; McElwain, J.C. The Evolution of Plants; Oxford University Press: New York, NY, USA, 2002; 378p. [Google Scholar]
- McIver, E.E.; Basinger, J.F. Early Tertiary floral evolution in the Canadian High Arctic. Ann. Mo. Bot. Gard. 1999, 86, 523–545. [Google Scholar] [CrossRef]
- Morrone, J.J. Fundamental biogeographic patterns across the Mexican Transition Zone: An evolutionary approach. Ecography 2010, 33, 355–361. [Google Scholar] [CrossRef]
- Robles-García, D.; Suzán-Azpiri, H.; Montoya-Esquivel, A.; García-Jiménez, J.; Esquivel-Naranjo, E.U.; Yahia, E.; Landeros-Jaime, F. Ethnomycological knowledge in three communities in Amealco, Quéretaro, México. J. Ethnobiol. Ethnomedicine 2018, 14, 7. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Taxon | Specimen Voucher | ITS | nrLSU | RPB2 |
|---|---|---|---|---|
| Boletellus ananas | REH8613 | ------------------------ | KP327629 | ------------------- |
| B. ananas | CFMR BZ-3167 | ------------------------ | MK601717 | MK766279 |
| B. ananas | NY815459 | ------------------------ | JQ924336 | KF112760 |
| B. ananas | NY796159 | ------------------------ | JQ924335 | ------------------- |
| B. ananiceps | REH9484 | ------------------------ | KP327655 | ------------------- |
| B. ananiceps | 3794 | ------------------------ | KP327666 | ------------------- |
| B. aurocontextus | N.K.Zeng2536 | MT822935 | MT829120 | ------------------- |
| B. belizensis | CFMR:BZ-316 TJB-9128 | MN250194 | MN250169 | ------------------- |
| B. belizensis | CFMR BZ-316 TJB-9128 | NG_078671 | ------------------------ | ------------------- |
| B. belizensis | CFMR:BZ-429 JCB-2001-238 | MN250210 | MN250185 | ------------------- |
| B. belizensis | CFMR:BZ-1564 BOS-218 | MN250209 | MN250184 | ------------------- |
| B. badiovinosus | REH8923 | KP327640 | ------------------------ | ------------------- |
| B. brunoflavus | B21061253 | ON364057 | ON195936 | ON390839 |
| B. brunoflavus | GDGM87995 | ON364056 | ON195935 | ------------------- |
| B. chrysenteron | 54/97 | DQ534634 | ------------------------ | ------------------- |
| B. chrysenteron | REH9015 | ------------------------ | KP327645 | ------------------- |
| B. chrysenteron | iNaturalist # 91541460 | OM809388 | OM809388 | ------------------- |
| B. chrysenteron | HKAS105270 | MT520080 | MT520080 | ------------------- |
| B. indistinctus | HKAS 77623 | ------------------------ | NG_058689 | KT990371 |
| B. indistinctus | HKAS80681 | ------------------------ | KT990532 | KT990368 |
| B. obscurococcineus | HKAS77662 | ------------------------ | ------------------------ | KT990372 |
| B. obscurococcineus | isolate: 3A3 | AB973769 | AB973769 | ------------------- |
| B. minimatenebris | MEXU-HO-30119; ITCV1076 | OR713119 | OR713121 | OQ938895 |
| B. minimatenebris | ITCV-1087 | OR713120 | OR713122 | OQ938894 |
| B. pseudochrysenteroides | Mushroom Observer #288188 | ------------------------ | MH257564 | ------------------- |
| B. sinapipes | REH9018 | ------------------------ | KP327647 | ------------------- |
| B. sinapipes | REH9408 | ------------------------ | KP327653 | ------------------- |
| B. shoreae | AP6679 | MH608209 | MH608211 | ------------------- |
| B. shoreae | AP6696 | MH608208 | MH608210 | ------------------- |
| B. wenshanensis | HKAS 122938 | ------------------------ | NG_154028 | ------------------- |
| B. wenshanensis | 532624MF_103_Wu2281 | ------------------------ | ON006512 | ON007376 |
| Boletellus sp. | HKAS 74888 | KF112413 | ------------------------ | KF112747 |
| Boletellus sp. | HKAS 80554 | ------------------------ | KT990535 | KT990535 |
| Boletellus sp. | HKAS122526 | ON794351 | ------------------------ | ------------------- |
| Boletellus sp. | TAA195080 | AM412293 | ------------------------ | ------------------- |
| Fungal Taxon | Specimen Voucher | RPB2 | ATP6 |
|---|---|---|---|
| Cacaoporus mexicanus | ITCV-1023, MEXU-HO 30103 | OQ938899 | ------------------------ |
| C. mexicanus | ITCV-904, MEXU-HO-30110 | OQ938897 | ------------------------ |
| C. mexicanus | ITCV-978, MEXU-HO 30128 | OQ938898 | OR683446 |
| C. mexicanus | ITCV-1027 | OQ938896 | OR683445 |
| C. pallidicarneus | OR0681 | MK372283 | MK372259 |
| C. pallidicarneus | OR0683 | MK372284 | MK372260 |
| C. pallidicarneus | OR1306 | MK372285 | MK372261 |
| C. pallidicarneus | SV0221 | MK372286 | MK372262 |
| C. pallidicarneus | SV0451 | MK372287 | MK372263 |
| C. tenebrosus | OR0654 | MK372288 | MK372265 |
| C. tenebrosus | OR1435 | MK372289 | MK372265 |
| C. tenebrosus | SV0223 | MK372290 | MK372266 |
| C. tenebrosus | SV0224 | MK372291 | MK372267 |
| C. tenebrosus | SV0452 | MK372292 | MK372269 |
| Cacaoporus sp. | SV0402 | MK372293 | MK372270 |
| Cupreoboletus poikilochromus | GS10070 | KT157068 | ----------- |
| Cu. poikilochromus | GS11008 | KT157067 | ----------- |
| Cyanoboletus bessettei | ARB1393B | MW737458 | ----------- |
| Cy. bessettei | MW737457 | ----------- | |
| Cy. brunneoruber | HKAS80579_1 | KT990401 | ----------- |
| Cy. brunneoruber | OR0233 | MG212628 | MG212542 |
| Cy. cyaneitinctus | Farid_340 | MW737461 | ----------- |
| Cy. cyaneitinctus | Farid_920 | MW737465 | ----------- |
| Cy. cyaneitinctus | JAB_324 | MW737469 | ----------- |
| Cy. cyaneitinctus | JAB_184 | MW737467 | ----------- |
| Cy. instabilis | HKAS59554 | KF112698 | ----------- |
| Cy. pulverulentus | RW109 | KT824013 | KT823980 |
| Cy. sinopulverulentus | HKAS59609 | KF112700 | ----------- |
| Cyanoboletus sp. | OR0257 | MG212629 | MG212543 |
| Cyanoboletus sp. | OR0322 | MH614768 | MH614673 |
| Cyanoboletus sp. | OR0491 | MT823996 | MH614674 |
| Cyanoboletus sp. | OR0961 | MH614770 | MH614675 |
| Chalciporus africanus | JD517 | KT823996 | KT823963 |
| Lanmaoa angustispora | HM605178 | KM605178 | ----------- |
| La. angustispora | HKAS74765 | KF112680 | ----------- |
| La. asiatica | HKAS63516 | KT990419 | ----------- |
| La. asiatica | OR0228 | MH614777 | MH614682 |
| La. carminipes | BOTH4591 | MG897439 | MG897419 |
| La. flavorubra | NY775777 | KF112681 | ----------- |
| La. pallidorosea | BOTH4432 | MG897437 | MG897417 |
| Lanmaoa sp. | OR0130 | MH614778 | MH614683 |
| Lanmaoa sp. | OR0370 | MH614779 | MH614684 |
| Rubinoboletus rubinus | AF2835 | KT823995 | KT823962 |
| Fungal Taxon | Specimen Voucher | nrLSU | RPB2 | TEF1 |
|---|---|---|---|---|
| Garcileccinum salmonicolor | MEXU-HO-30108 | OQ909093 | OQ938917 | OR683442 |
| G. salmonicolor | MEXU-HO-30105 | OQ909094 | OQ938918 | OR683443 |
| G. violaceotinctum | CFMR-BZ-1676 | -------------------- | QGW57932 | QGW57818 |
| G. violaceotinctum | CFMR-BZ-3169 | -------------------- | QGW57933 | QGW57819 |
| G. viscosum | NY 01194025 | -------------------- | QGW57910 | QGW57795 |
| G. viscosum | CFMR-BZ-2406 | ------------------- | ----------------- | ----------------- |
| Hemileccinum rugosum | HKAS 84355 | KT990578 | ----------------- | ----------------- |
| Lanmaoa angustispora | HM605178 | -------------------- | KM605178 | ----------------- |
| La. angustispora | HKAS74765 | -------------------- | KF112680 | ----------------- |
| La. asiatica | HKAS63516 | -------------------- | KT990419 | ----------------- |
| La. asiatica | OR0228 | -------------------- | MH614777 | ----------------- |
| La. carminipes | BOTH4591 | -------------------- | MG897439 | ----------------- |
| La. flavorubra | NY775777 | -------------------- | KF112681 | ----------------- |
| La. pallidorosea | BOTH4432 | -------------------- | MG897437 | ----------------- |
| Lanmaoa sp. | OR0130 | -------------------- | MH614778 | ----------------- |
| Lanmaoa sp. | OR0370 | -------------------- | MH614779 | ----------------- |
| Leccinellum albellum | MICH-KUO-09200807 | MK601747 | MK766309 | MK721101 |
| Le. albellum | MICH-KUO-07241101 | MK601746 | MK766308 | MK721100 |
| Le. alborufescens | FHMU 1758 | -------------------- | MK816332 | MK816330 |
| Le. alborufescens | FHMU1908 | -------------------- | MK816333 | MK816330 |
| Le. citrinum | HKAS53427 | KF112488 | KF112727 | KF112253 |
| Le. citrinum | HKAS53410 | KT990585 | KT990421 | ----------------- |
| Le. crocipodium | MICH-KUO-07050707 | MK601749 | MK766311 | MK721103 |
| Le. pseudoscabrum | MICH-KUO-10200710 | MK601753 | MK766314 | MK721107 |
| Le. pseudoscabrum | CFMR-DPL-11432 | MK601752 | MK766313 | MK721106 |
| Le. quercophilum | NY01293386 | -------------------- | ----------------- | ----------------- |
| Leccinum aff. scabrum | HKAS57266 | KF112442 | KF112722 | KF112248 |
| L. album | Li1072 | MZ392872 | MW439260 | MW439267 |
| L. aurantiacum | L0342207 | MK601759 | MK766318 | MK721113 |
| L. duriusculum | Yang5971 | MZ675541 | MZ707779 | MZ707785 |
| L. holopus | MICH-KUO-09150707 | MK601763 | MK766322 | MK721117 |
| L. holopus | CFMR-BOS-569 | MK601762 | MK766321 | MK721116 |
| L. holopus | Yang 5972 | -------------------- | MW439258 | -------------------- |
| L. juarenzense | MEXU HO 30,112 | OQ909095 | OQ938916 | -------------------- |
| L. manzanitae | NY14041 | MK601765 | MK766324 | MK721119 |
| L. melaneum | FB491 | MZ675542 | MZ707780 | MZ707786 |
| L. monticola | NY00815448 | HQ161869 | HQ161838 | ----------------- |
| L. monticola | NY760388 | MK601766 | ----------------- | ----------------- |
| L. monticola | HKAS76669 | KF112443 | KF112723 | KF112249 |
| L. oaxacanum | MEXU-HO-30460 | OQ909096 | OQ938915 | OR683444 |
| L. parascabrum | Li1700 | MW413912 | MW439265 | MW439272 |
| L. parascabrum | Wu1784 | -------------------- | MW439264 | MW439271 |
| L. pseudoborneense | WGS960 | MW413909 | MW439262 | MW439270 |
| L. pseudoborneense | WGS965 | MW413910 | MW439263 | MW439269 |
| L. pseudoborneense | Zhao773 | -------------------- | MZ543309 | MZ543307 |
| L. pseudoborneense | Zhao476 | -------------------- | MZ543308 | MZ543306 |
| L. pseudoborneense | WGS947 | MW413908 | MW439261 | MW439268 |
| L. quercinum | HKAS63502 | KF112444 | KF112724 | KF112250 |
| L. scabrum | HKAS53604 | KT990586 | KT990422 | --------------------- |
| L. scabrum | HKAS56371 | KT990587 | KT990423 | KT990782 |
| L. snellii | CFMR-BOS-579 | MK601773 | MK766331 | MK721127 |
| L. tablense | NY00796175 | MK601775 | MK766333 | MK721129 |
| L. tablense | NY760387 | MK601774 | MK766332 | MK721128 |
| L. variicolor | HKAS57758 | KF112445 | KF112725 | KF112251 |
| L. versipelle | CFMR-DLC2002-122 | MK601778 | MK766336 | MK721132 |
| L. versipelle | FB27 | MZ675546 | MZ707782 | MZ707790 |
| L. versipelle | LJW418 | MZ675545 | MZ707781 | MZ707789 |
| Fungal Taxon | Specimen Voucher | ITS | nrLSU |
|---|---|---|---|
| Alessioporus ichnusanus | MG 549a | KJ729493 | KJ729506 |
| A. rubriflavus | JLF2561 | KC812305 | KC812306 |
| Pulchroboletus neoregius | MEXU-HO-30423 | ------------------------------- | OQ940034 |
| P. neoregius | MEXU-HO-30424 | OQ940040 | ------------------------ |
| P. neoregius | MEXU-HO-30425 | OQ940039 | OQ940035 |
| P. neoregius | MEXU-HO-30426 | OQ940040 | OQ940033 |
| P. neoregius | NY796166 | JQ924304 | JQ924344 |
| P. neoregius | NY575822 | JQ924303 | JQ924343 |
| P. roseoalbidus | AMB 12757 | NR_154305 | NG_060126 |
| P. roseoalbidus | MCVE 17577 | KJ729490 | KJ729503 |
| P. roseoalbidus | MCVE 18217 | KJ729488 | KJ729501 |
| P. roseoalbidus | MG416a | KJ729489 | KJ729502 |
| P. roseoalbidus | MG532a | KJ729487 | KJ729500 |
| P. rubricitrinus | Farid335 | MF193884 | MG026638 |
| P. sclerotiorum | Flas-F-60333 | MF098659 | MF614166 |
| P. sclerotiorum | Mushroom Observer #243879 | MH257550 | MH257545 |
| Pulchroboletus sp. | JLF5434 | MH213050 | MH201325 |
| Pulchroboletus sp. | JLF6328 | MH213051 | MH201326 |
| Pulchroboletus sp. | JLF6329 | MH213053 | MH201328 |
| Fungal Taxon | Specimen Voucher | ITS | nrLSU | RPB2 |
|---|---|---|---|---|
| Tylopilus aff. balloui | 1D10 | AB973734 | ------------- | ------------- |
| T. aff. balloui | Mushroom 247674 | KY859806 | ------------- | ------------- |
| T. argillaceus | HKAS90186 | ------------- | KT990589 | KT990424 |
| T. atroviolaceobrunneus | HKAS84351 | ------------- | KT990625 | MT110421 |
| T. aurantiacus | Osmundson 1198 | ------------- | EU430740 | --------------- |
| T. aurantiacus | HKAS59700 | ------------- | NG081276 | KF112740 |
| T. aurantiacus | NY2072258 | ------------- | EU430740 | ------------- |
| T. balloui | CMU51-SL-42 | KX017307 | KX017298 | ------------- |
| T. balloui | CMU51-SL-39 | KX017306 | KX017297 | ------------- |
| T. balloui | CMU51-SL-37 | KX017305 | KX017296 | ------------- |
| T. balloui | CMU51-SL-32 | KX017304 | KX017295 | ------------- |
| T. balloui | 2D7 | AB973758 | ------------- | ---------------------- |
| T. balloui | 2D6 | AB973757 | ------------- | ---------------------- |
| T. balloui | 1D9 | AB973733 | ------------- | ---------------------- |
| T. balloui | 1E1 | AB973735 | ------------- | ---------------------- |
| T. balloui | FMNH 1073250 | ------------- | EU430733 | ---------------------- |
| T. balloui | Osmundson 1121 | ------------- | EU430743 | EU434333 |
| T. balloui | Osmundson 1117 | ------------- | EU430741 | ---------------------- |
| T. balloui | Osmundson 1105 | ------------- | ------------- | ---------------------- |
| T. balloui | HKAS51151 | ------------- | KT990590 | KT990425 |
| T. formosus | PDD72637 | HM060320 | HM060319 | ---------------------- |
| T. griseipurpureus | MG521a | KM975484 | KM975493 | ---------------------- |
| T. himalayanus | DC 17-25 | MG799322 | MG799328 | ---------------------- |
| T. leucomycelinus | DR2800 | MN115814 | MN115804 | ---------------------- |
| T. leucomycelinus | JBSD127420 | MN115813 | MN115803 | MN095210 |
| T. leucomycelinus | JBSD127419 | MN115812 | MN115802 | MN095209 |
| T. leucomycelinus | ECC 64 | OQ984890 | ------------- | ---------------------- |
| T. leucomycelinus | GH111 | OQ984889 | ------------- | ---------------------- |
| T. leucomycelinus | BZ2409 | MN115815 | MN115805 | ---------------------- |
| T. leucomycelinus | MC53 | OQ984888 | ------------- | ---------------------- |
| T. leucomycelinus | 18463 | JF908789 | ------------- | ---------------------- |
| T. leucomycelinus | Halling 8526 | ------------- | EU430736 | EU434336 |
| T. leucomycelinus | Halling 8521 | ------------- | EU430735 | ---------------------- |
| T. leucomycelinus | JC1298 | OQ984887 | ------------- | ---------------------- |
| T. leucomycelinus | F1030852 | MN115811 | ------------- | ---------------------- |
| T. microspores | HMAS84730 | NR137924 | NG059538 | ---------------------- |
| T. neofelleus | DC 16-64 | MG777524 | MG777529 | ---------------------- |
| T. neofelleus | YT20120811 | KM975487 | KM975495 | ---------------------- |
| T. otsuensis | HKAS53401 | KF112449 | KF112797 | |
| T. plumbeoviolaceoides | GDGM:42624 | ------------- | KM975498 | ---------------------- |
| T. plumbeoviolaceoides | Chu25 | DQ407261 | ------------- | ---------------------- |
| T. plumbeoviolaceoides | GDGM43255 | KM975490 | ------------- | ---------------------- |
| T. plumbeoviolaceoides | GDGM20311 | NR137601 | NG059490 | ---------------------- |
| T. porphyrosporus | 17898 | JF908788 | ---------------------- | |
| T. pseudoballoui | DC 17-30 | MG799329 | MG799327 | ---------------------- |
| T. pseudoballoui | DC 17-35 | MG799324 | MG799325 | ---------------------- |
| T. pseudoleucomycelinus | MEXU-HO-30115 | OQ940043 | OQ940037 | OQ938903 |
| T. pseudoscaber | DAVFP27804 | JF899578 | ------------- | ---------------------- |
| T. rubrobrunneus | F-PRL7359 | GQ166869 | ------------- | ---------------------- |
| T. rubrotinctus | KK259 | --------------- | MT154733 | MW165283 |
| T. rubrotinctus | LF1101 | -------------- | MT154732 | MW165291 |
| T. sordidus | JMP0094 | EU819450 | -------------------- | ---------------------- |
| T. virens | HE2757 | KC505585 | -------------------- | ---------------------- |
| Species | Voucher | ITS | LSU | RPB2 |
|---|---|---|---|---|
| Xerocomus coccolobae | JBSD133071 (ANGE1405) | OQ108302 | OQ102365 | OQ117434 |
| X. coccolobae | JBSD133068 (ANGE915) | OQ108300 | OQ102363 | OQ117435 |
| X. coccolobae | JBSD133069 (ANGE965) | OQ108301 | OQ102364 | OQ117436 |
| X. hygrophorus | MEXU-HO-30131 | OQ984883 | OQ975751 | OQ938901 |
| X. illudens | JLH MyCoPortal 6604756 | MK578706 | MK578706 | ------------- |
| X. illudens | DD9854 | ------------- | AY612840 | ------------- |
| X. illudens | MB03-005 | JQ003658 | JQ003658 | ------------- |
| X. illudens | iNaturalist # 130560701 | OP643377 | OP643377 | ------------- |
| X. ferrugineus | CFMR BOS 545 | ------------- | MK601819 | MK766375 |
| X. ferrugineus | KUO-08100701 | ------------- | MK601820 | MK766376 |
| X. ferrugineus | UBC:F31236 | MZ817040 | MZ817040 | ------------- |
| X. ferrugineus | Champ-29 | KX449436 | KX449436 | ------------- |
| X. fulvipes | HKAS 76666 | -------------- | KF112390 | KF112789 |
| X. magniporus | HKAS 58000 | -------------- | KF112392 | KF112781 |
| X. puniceiporus | HKAS 80683 | -------------- | KU974141 | KU974146 |
| X. perplexus | MB00-005 | -------------- | JQ003702 | ------------- |
| X. rugosellus | HKAS 58865 | -------------- | KF112389 | KF112784 |
| Xerocomus sp. | MAN061 | JQ003656 | JQ003707 | ------------- |
| Xerocomus sp. | MAN061 | JQ003656 | ------------ | ------------- |
| Xerocomus sp. | OR0053 | ------------- | ------------ | MH580834 |
| Xerocomus sp. | OR0237 | MH580835 | ||
| X. spadiceus | MICH KUO 7080702 | ------------- | MK601822 | MK766378 |
| X. spadiceus | JLF4012 | KX534078 | -------------- | -------------- |
| X. spadiceus | JLF4060 | KX534079 | -------------- | -------------- |
| X. silwoodensis | JAM0612 | ------------- | KF030323 | -------------- |
| X. subtomentosus | VDKO0987 | ------------- | -------------- | MG212657 |
| X. subtomentosus | Xs1 | AF139716 | -------------- | -------------- |
| X. subtomentosus | Both3312 | DQ066413 | -------------- | -------------- |
| X. subtomentosus | 1549a-Q-6103 | KM248935 | -------------- | -------------- |
| X. tenax | REH6871 | ------------- | KF030320.1 | -------------- |
| X. tenax | MICH KUO 8241404 | ------------- | MK601823 | MK766379 |
| X. tenax | REH6871 | -------------- | KF030320 | -------------- |
| Phylloporus imbricatus | HKAS84355 | NG_059621 | KT990578 | KT990413 |
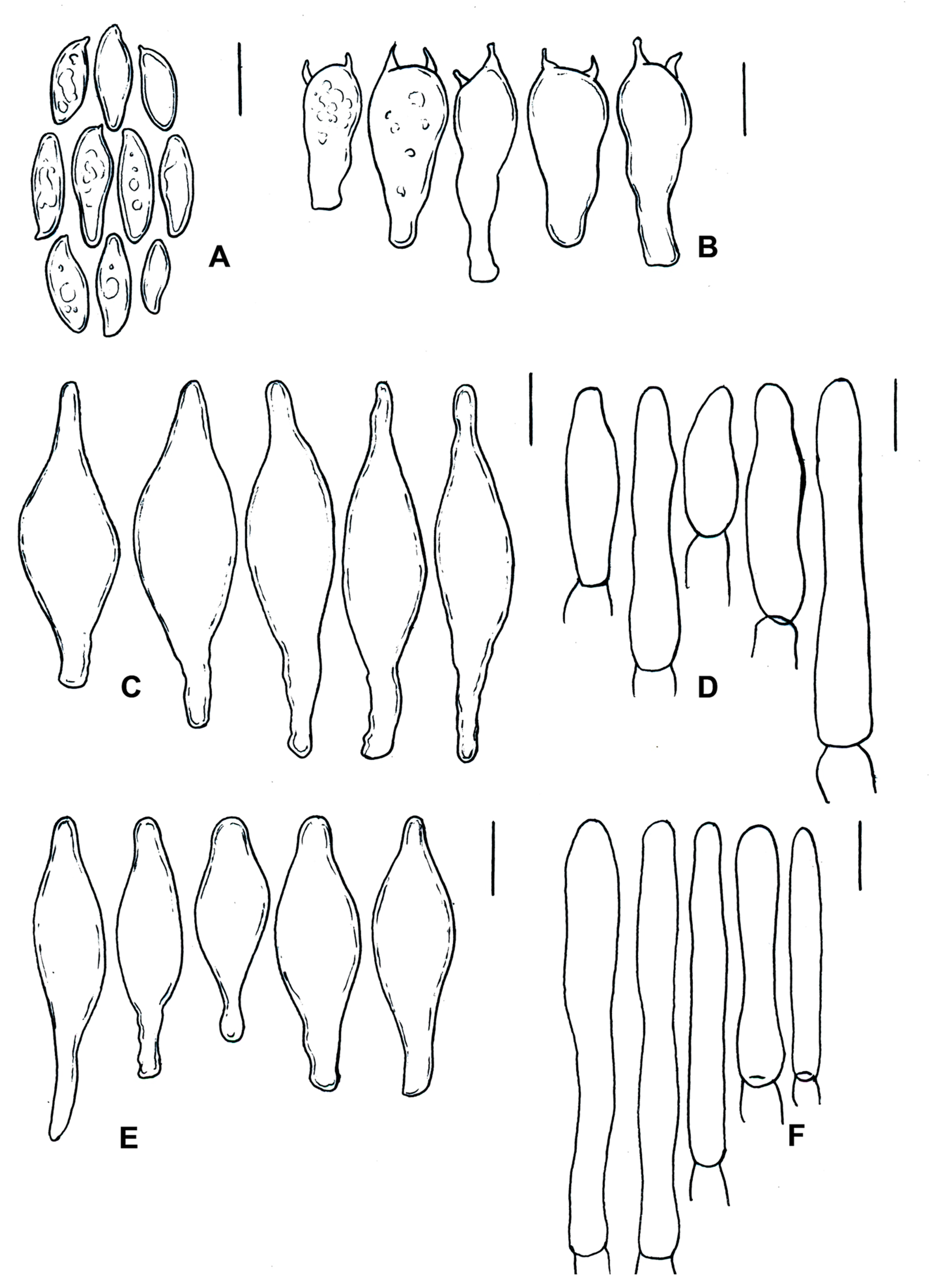
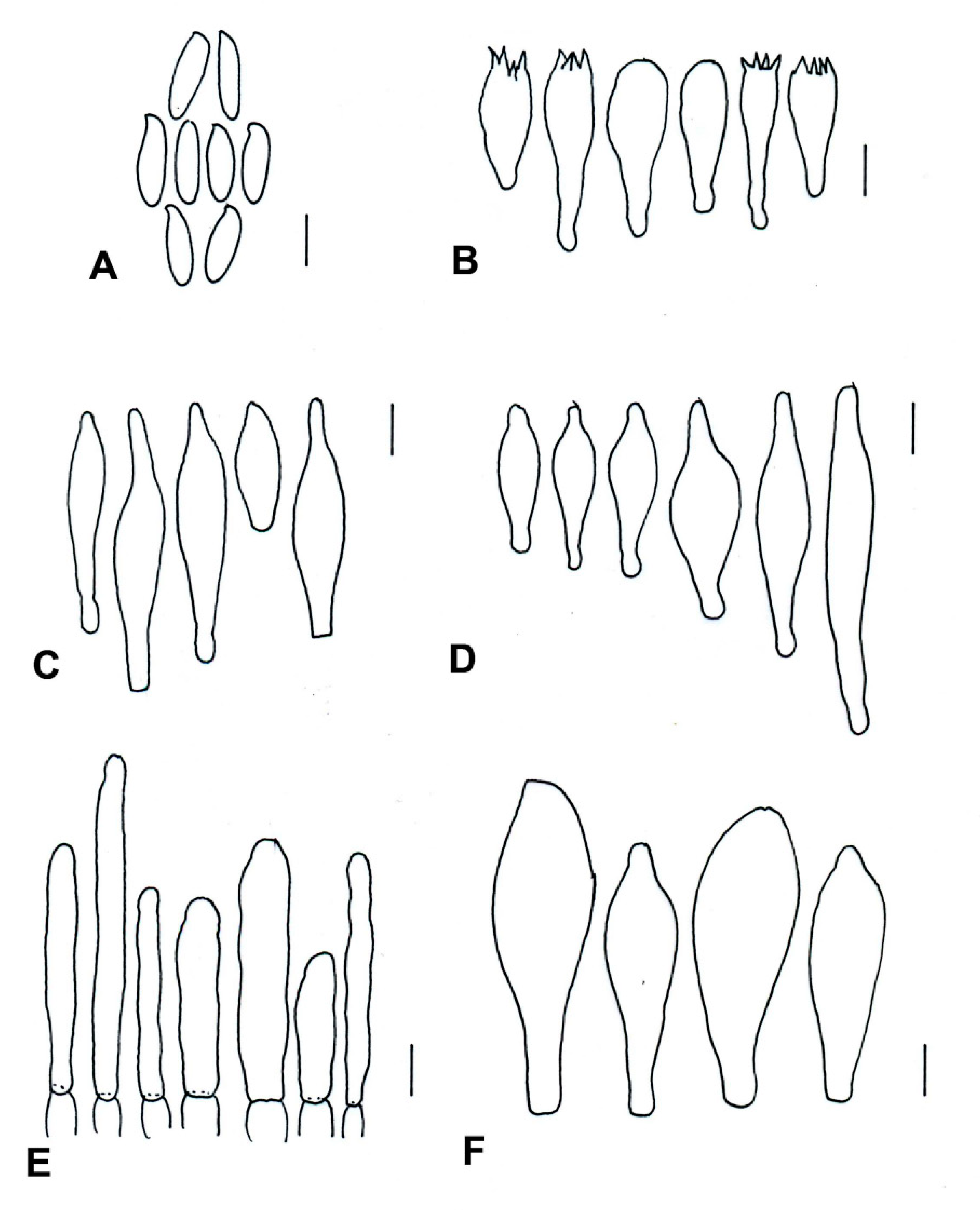
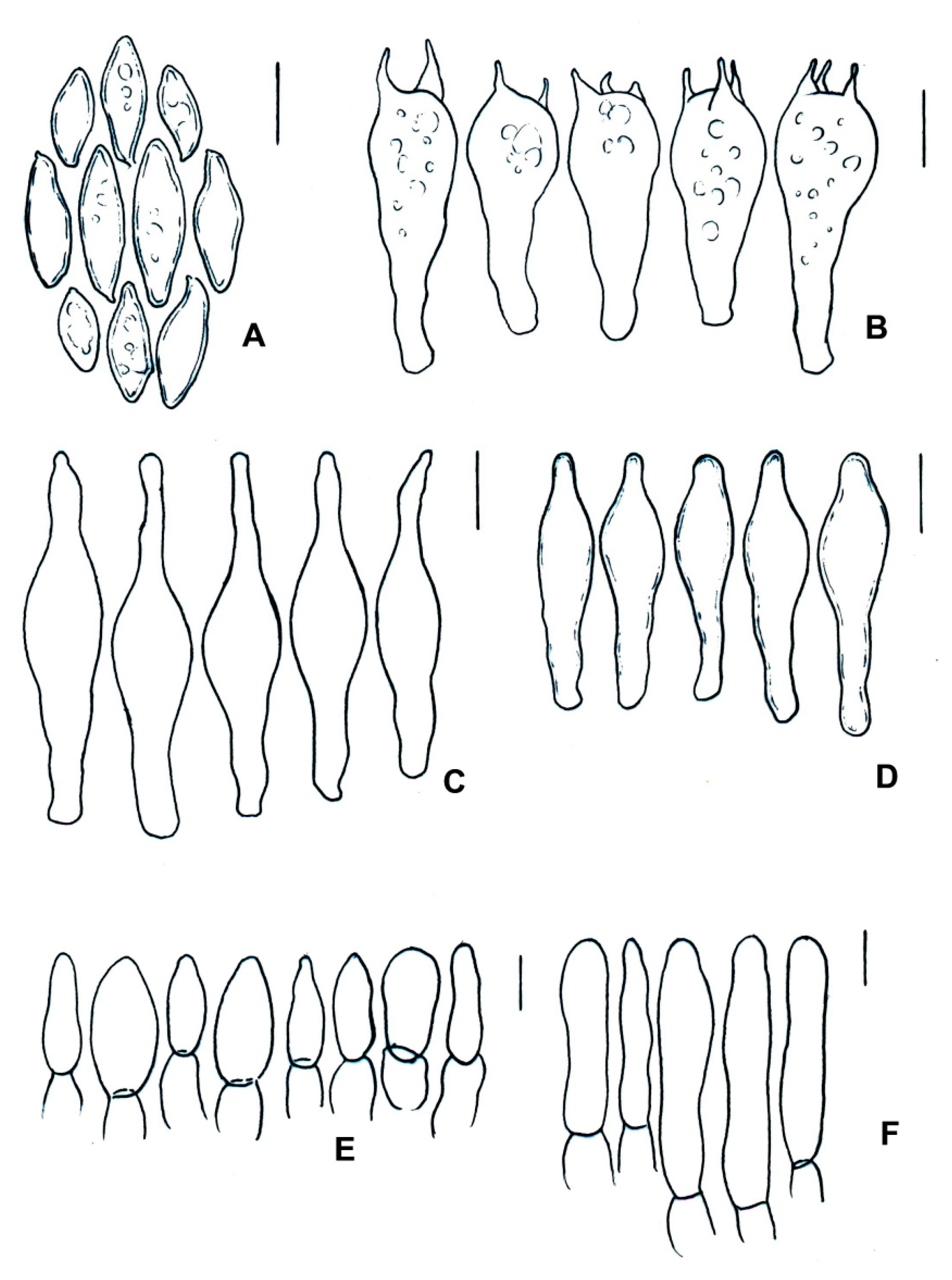
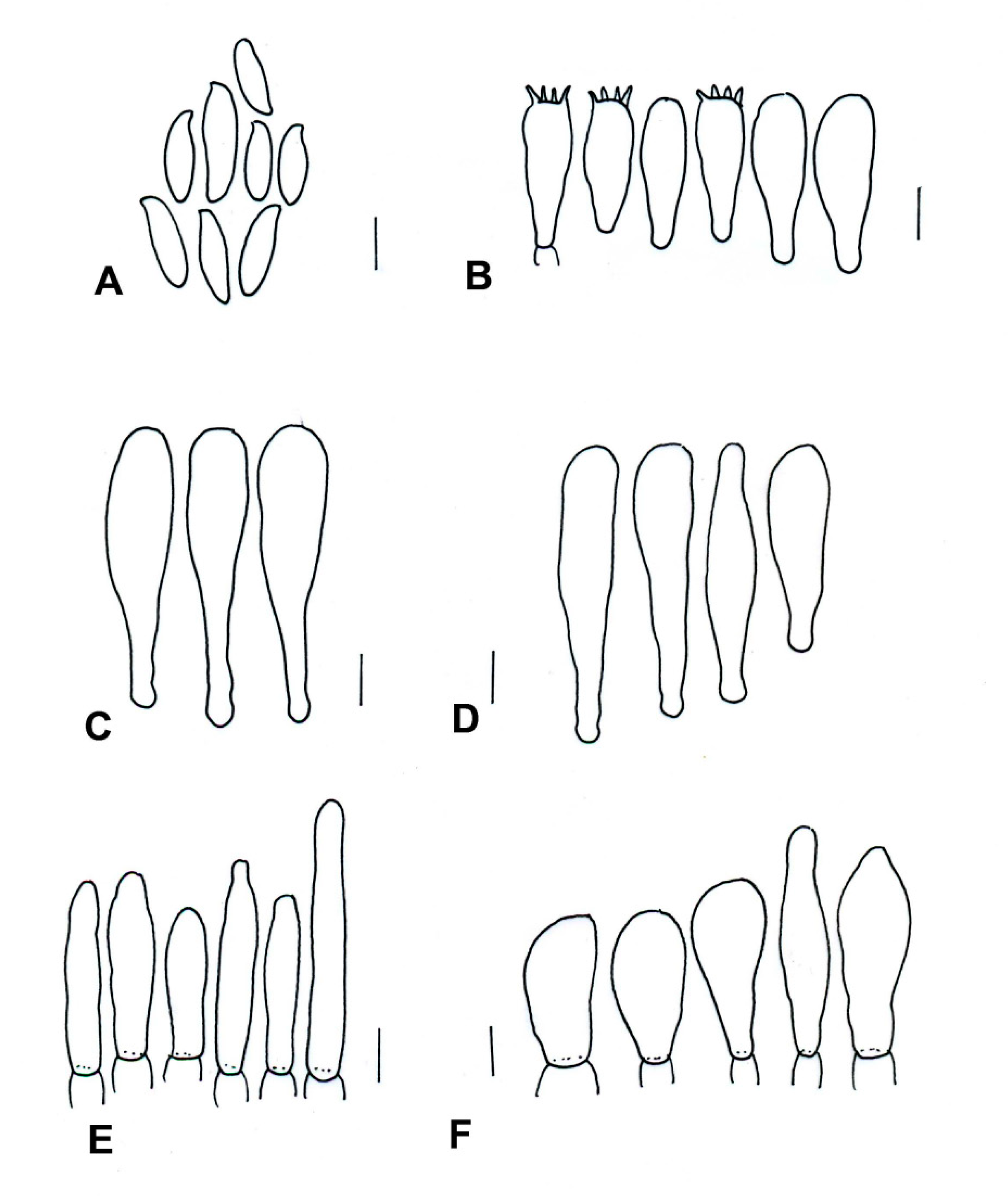
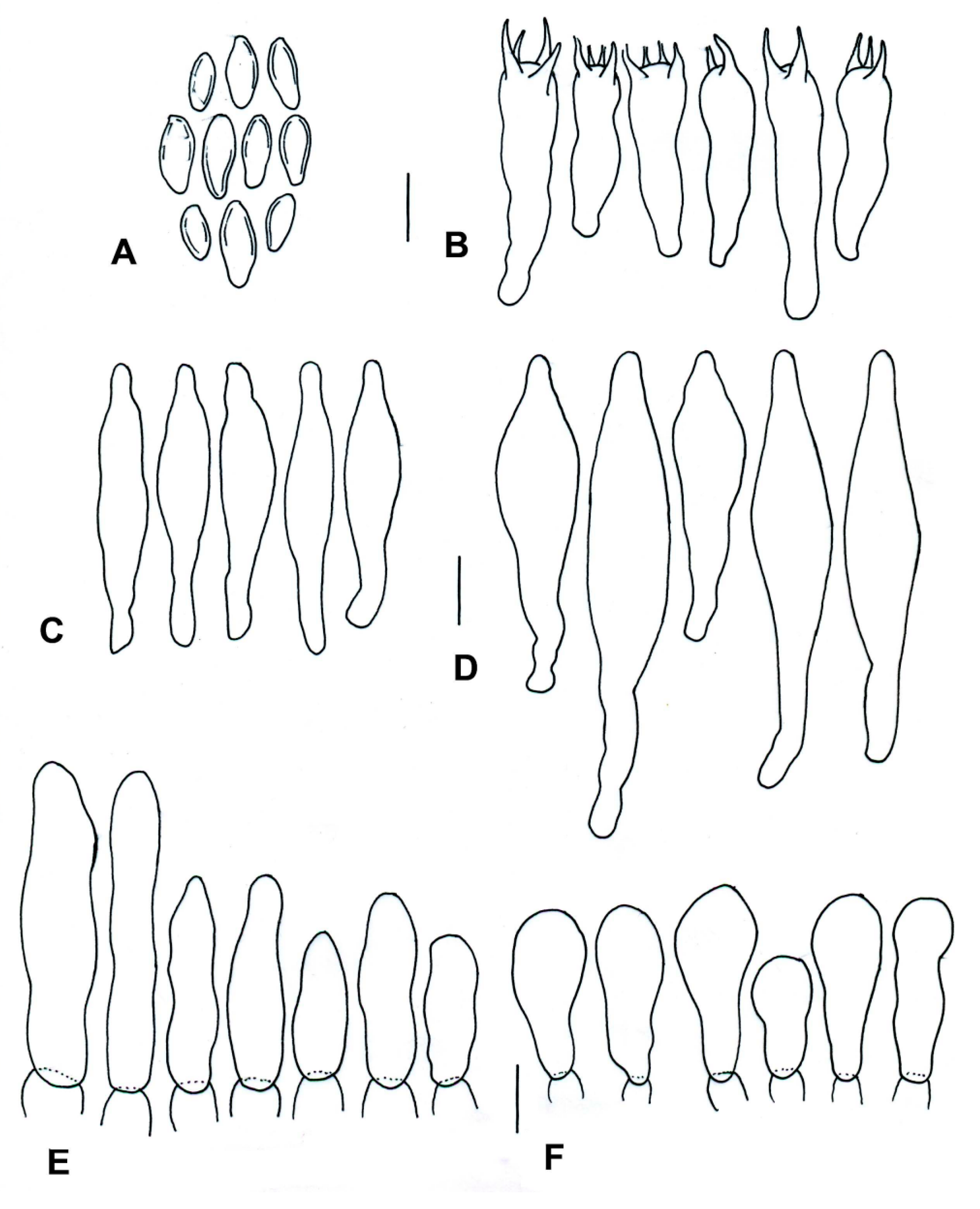
| Species | Basidiomata | Basidiospores | Cystidia or Pseudocystidia | Terminal Elements of the Pileipellis | Caulobasidia |
|---|---|---|---|---|---|
| T. balloui * | 40–85 mm. | 8.2–10.5 × 3–3.7 μm, ellipsoid, ovoid to fusoid. | 27–60 (−80) × 6– 21 μm, utriform. | 4–8 μm, elongated cylindrical. | Clavate dermatocystidia, 33 × 14 μm. |
| T. leucomycelinus ** | (36–) 43–72 (–110) mm. | 6.6 (–8.2) × (3.9–) (–5.5) μm, allantoid, phaseoliform, ovoid, subglobose. | (38–) 42–75 (–83) × 7–16 (–19) μm, fusiform to ventricose fusiform or lageniform to sublageniform. | 24–115 × 3–8 μm, cylindrical. | 40–75 (–88) × 7–18 (–20) μm, fusiform to ventricose fusiform or lageniform to sublageniform. |
| T. pseudoleucomycelinus *** | 19–28 (40) mm. | 5–7 (–10) × 4–5 µm ovoid to lachrymoid, ellipsoid. | 30–35 (−66) × 7–8 (−12) µm, fusiform, subclavate to piriform. | 30–40 × 7–9 µm, cylindrical. | Caulocystidia 25–60 × 7–11 µm, fusiform, ventricose-rostrate. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayala-Vásquez, O.; Pérez-Moreno, J.; Pinzón, J.P.; Garibay-Orijel, R.; García-Jiménez, J.; de la Fuente, J.I.; Venegas-Barrera, C.S.; Martínez-Reyes, M.; Montoya, L.; Bandala, V.; et al. Broadening the Knowledge of Mexican Boletes: Addition of a New Genus, Seven New Species, and Three New Combinations. J. Fungi 2023, 9, 1126. https://doi.org/10.3390/jof9121126
Ayala-Vásquez O, Pérez-Moreno J, Pinzón JP, Garibay-Orijel R, García-Jiménez J, de la Fuente JI, Venegas-Barrera CS, Martínez-Reyes M, Montoya L, Bandala V, et al. Broadening the Knowledge of Mexican Boletes: Addition of a New Genus, Seven New Species, and Three New Combinations. Journal of Fungi. 2023; 9(12):1126. https://doi.org/10.3390/jof9121126
Chicago/Turabian StyleAyala-Vásquez, Olivia, Jesús Pérez-Moreno, Juan Pablo Pinzón, Roberto Garibay-Orijel, Jesús García-Jiménez, Javier Isaac de la Fuente, Crystian Sadiel Venegas-Barrera, Magdalena Martínez-Reyes, Leticia Montoya, Víctor Bandala, and et al. 2023. "Broadening the Knowledge of Mexican Boletes: Addition of a New Genus, Seven New Species, and Three New Combinations" Journal of Fungi 9, no. 12: 1126. https://doi.org/10.3390/jof9121126
APA StyleAyala-Vásquez, O., Pérez-Moreno, J., Pinzón, J. P., Garibay-Orijel, R., García-Jiménez, J., de la Fuente, J. I., Venegas-Barrera, C. S., Martínez-Reyes, M., Montoya, L., Bandala, V., Aguirre-Acosta, C. E., Martínez-González, C. R., & Hernández-Del Valle, J. F. (2023). Broadening the Knowledge of Mexican Boletes: Addition of a New Genus, Seven New Species, and Three New Combinations. Journal of Fungi, 9(12), 1126. https://doi.org/10.3390/jof9121126