
Fungal Species from Rhododendron sp.: Discosia rhododendricola sp.nov, Neopestalotiopsis rhododendricola sp.nov and Diaporthe nobilis as a New Host Record.

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Morphological Observation, and Fungal Isolation
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Phylogenetic Analyses
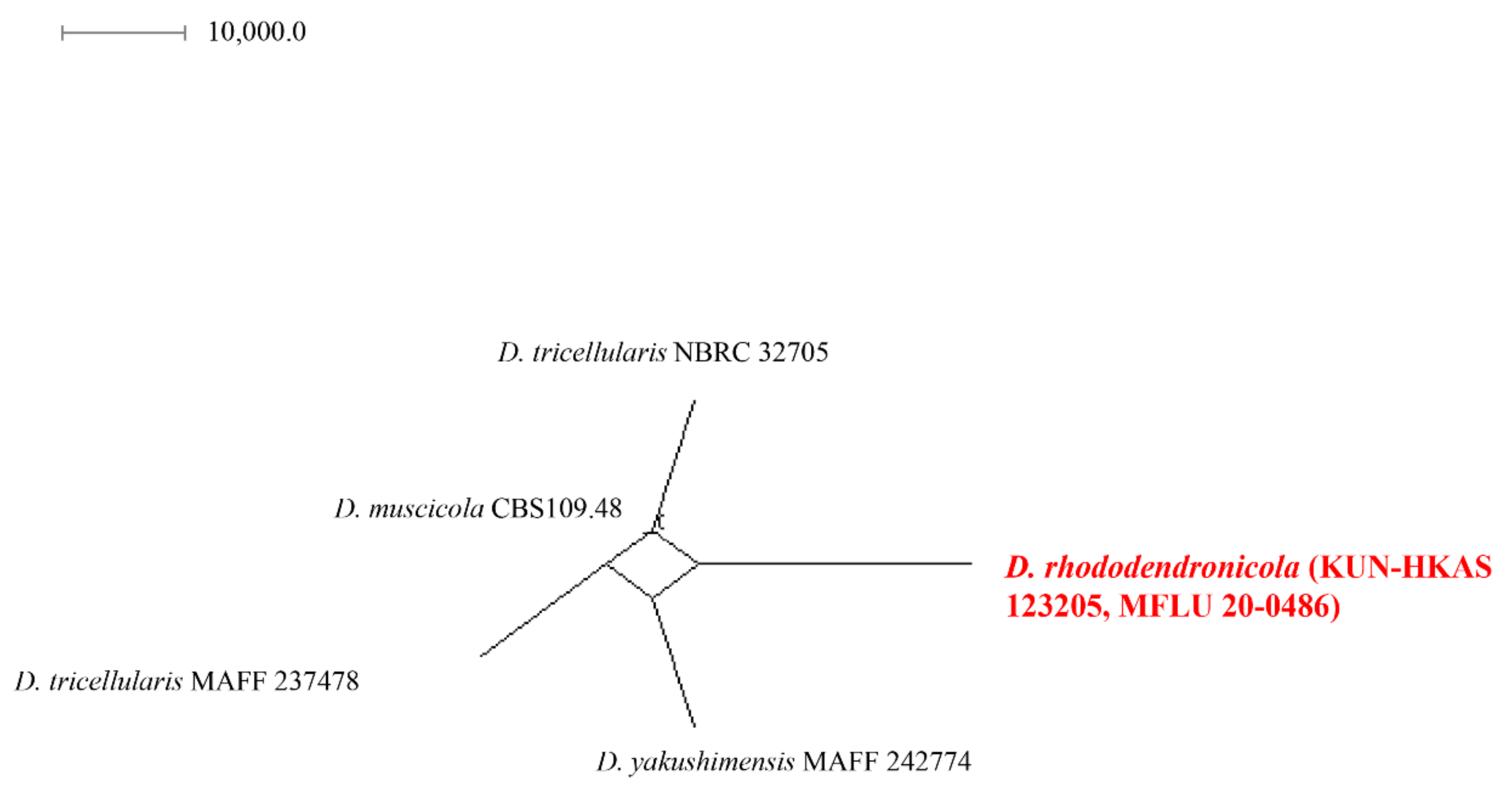
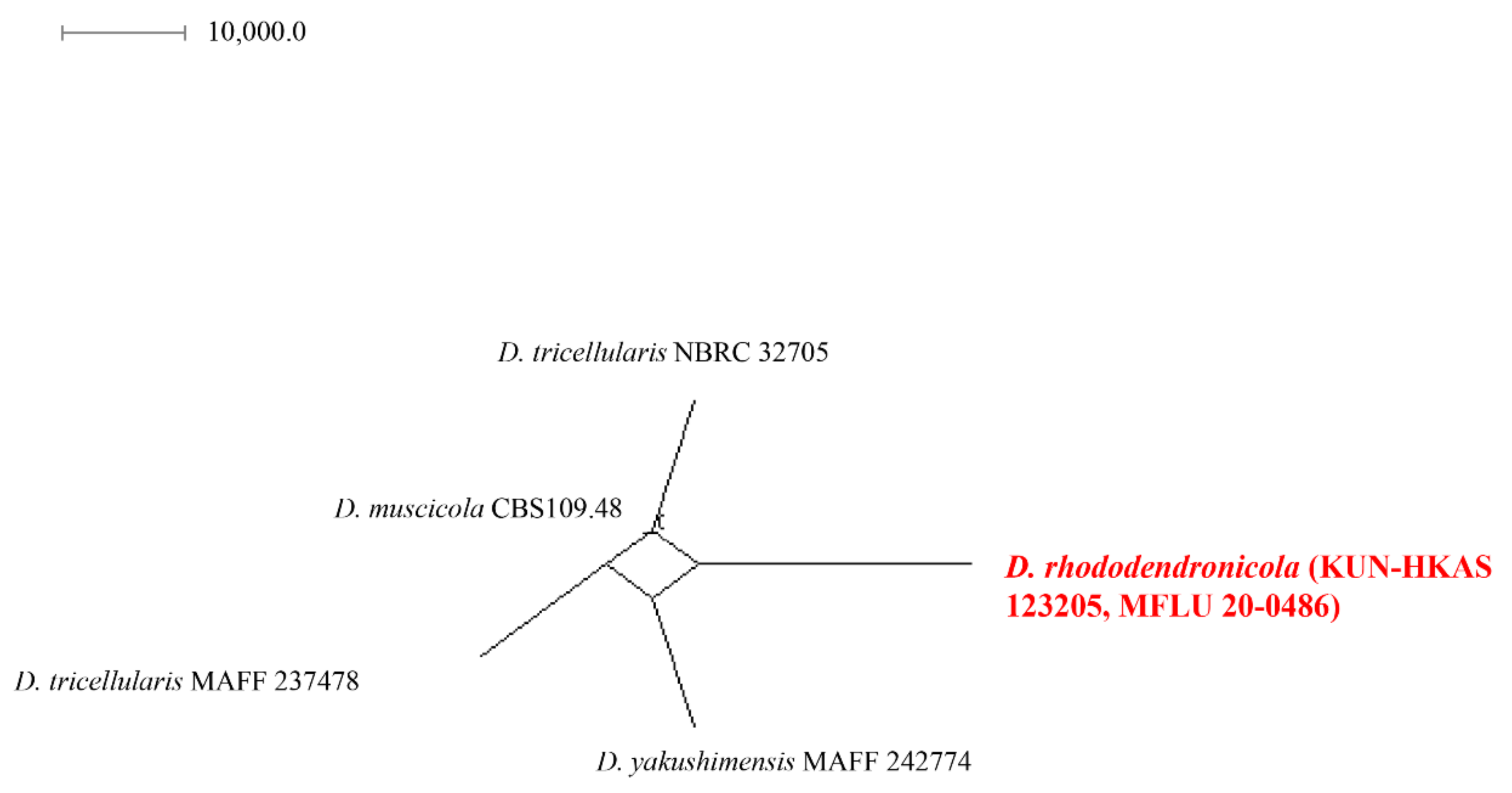
2.4. Genealogical Concordance Phylogenetic Species Recognition (GCPSR) Analysis
2.5. Discosia, Habitat and Known Distribution Checklist Associated with Rhododendron sp.
3. Results
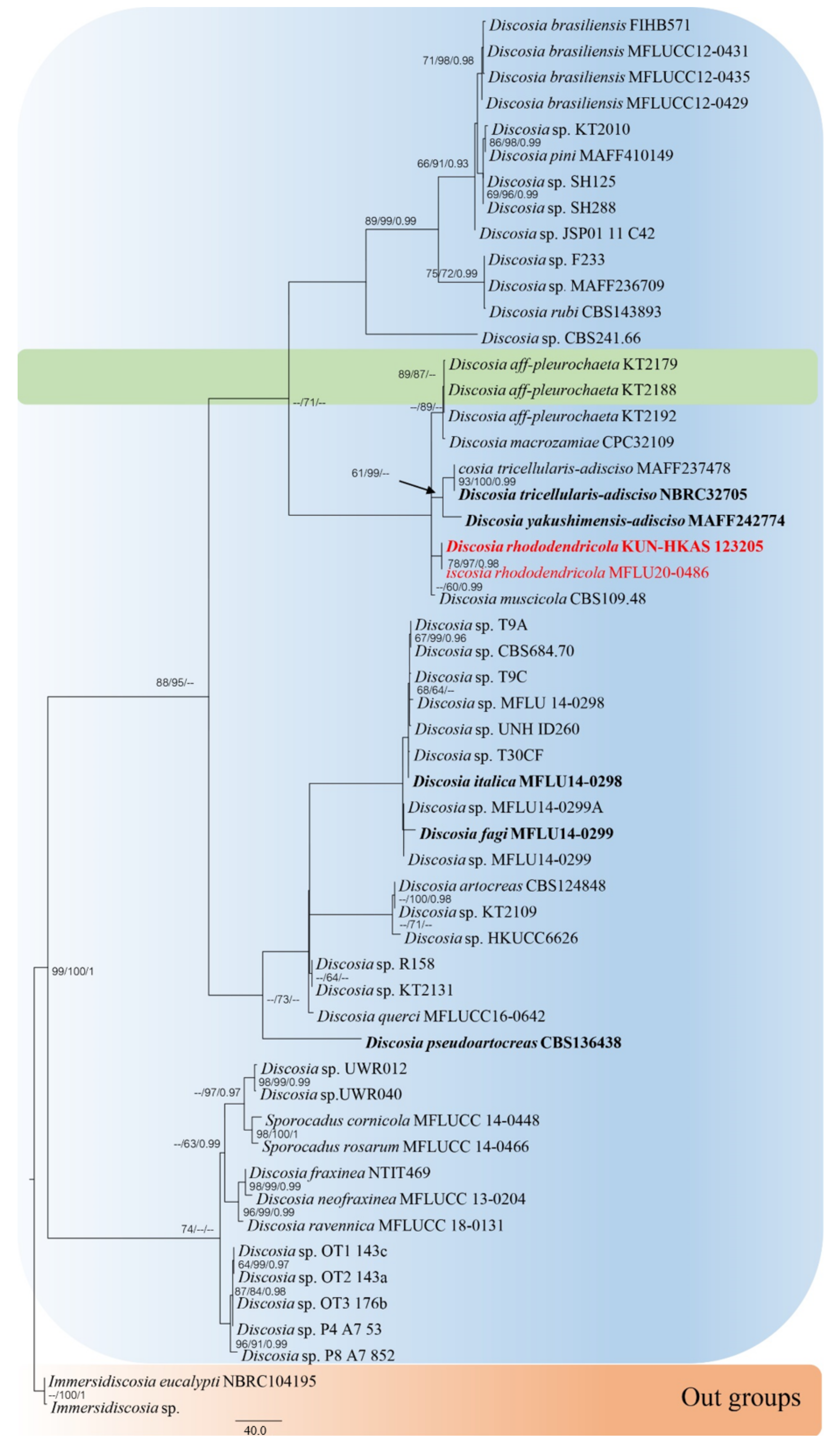
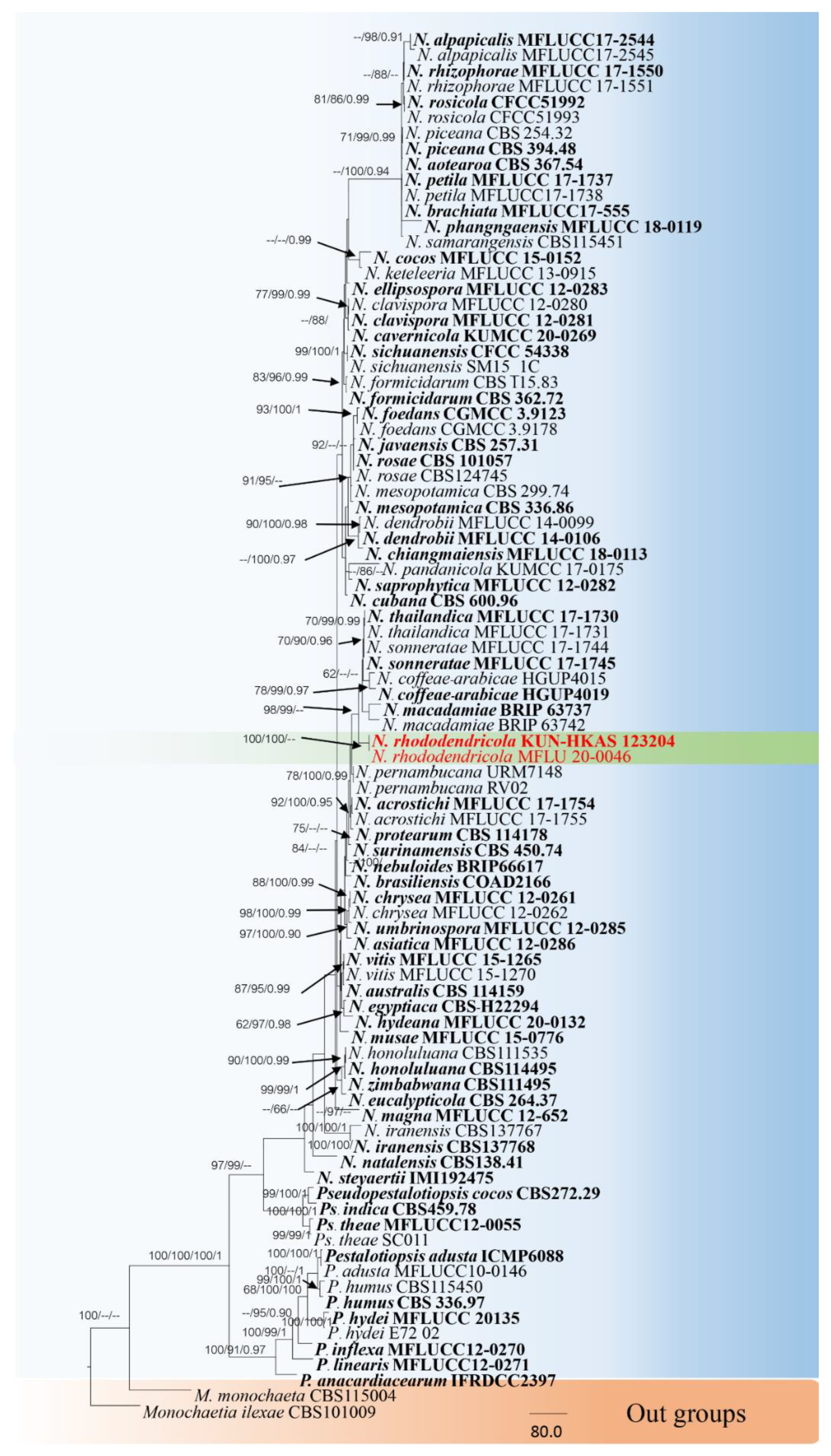
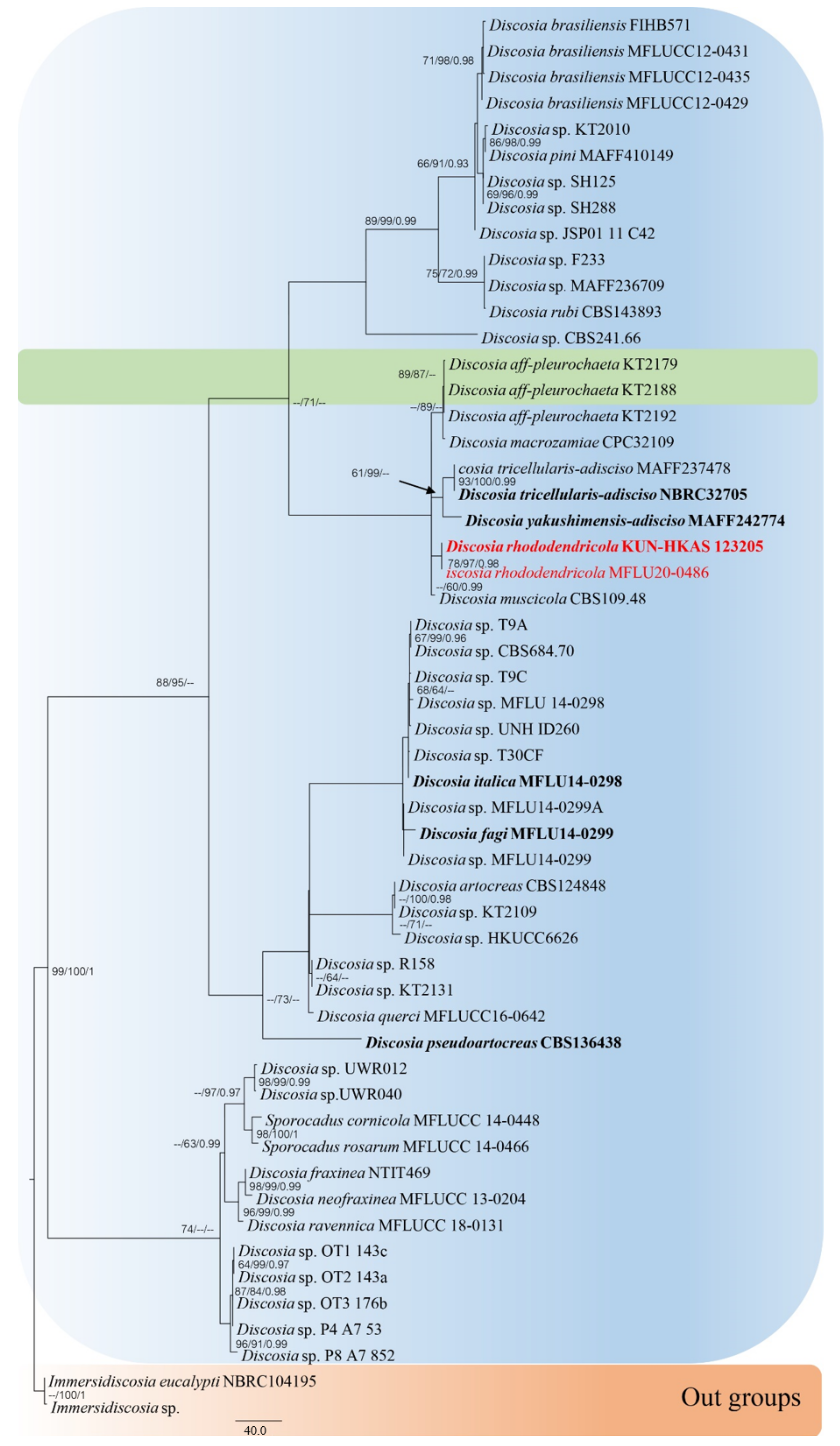
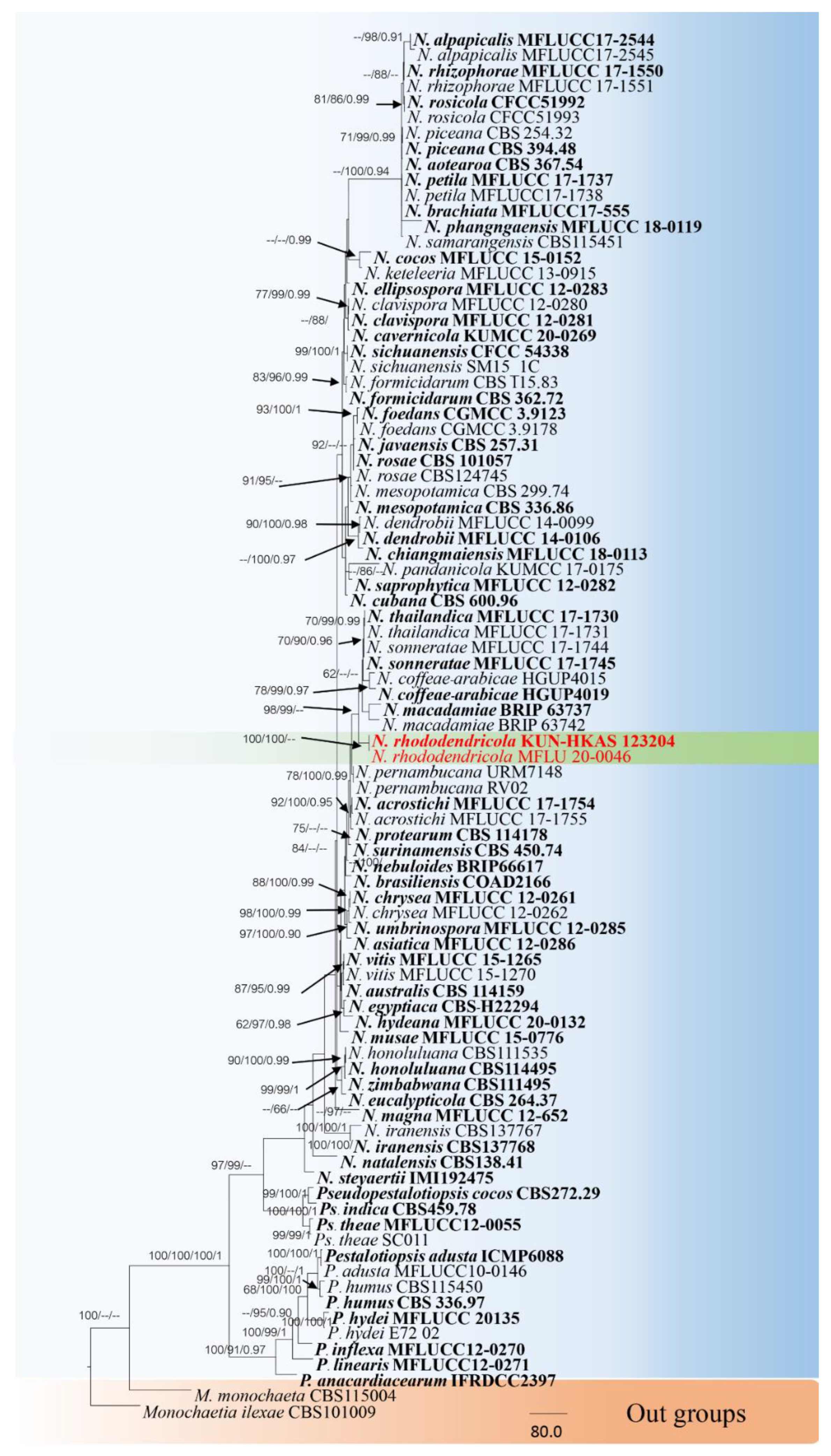
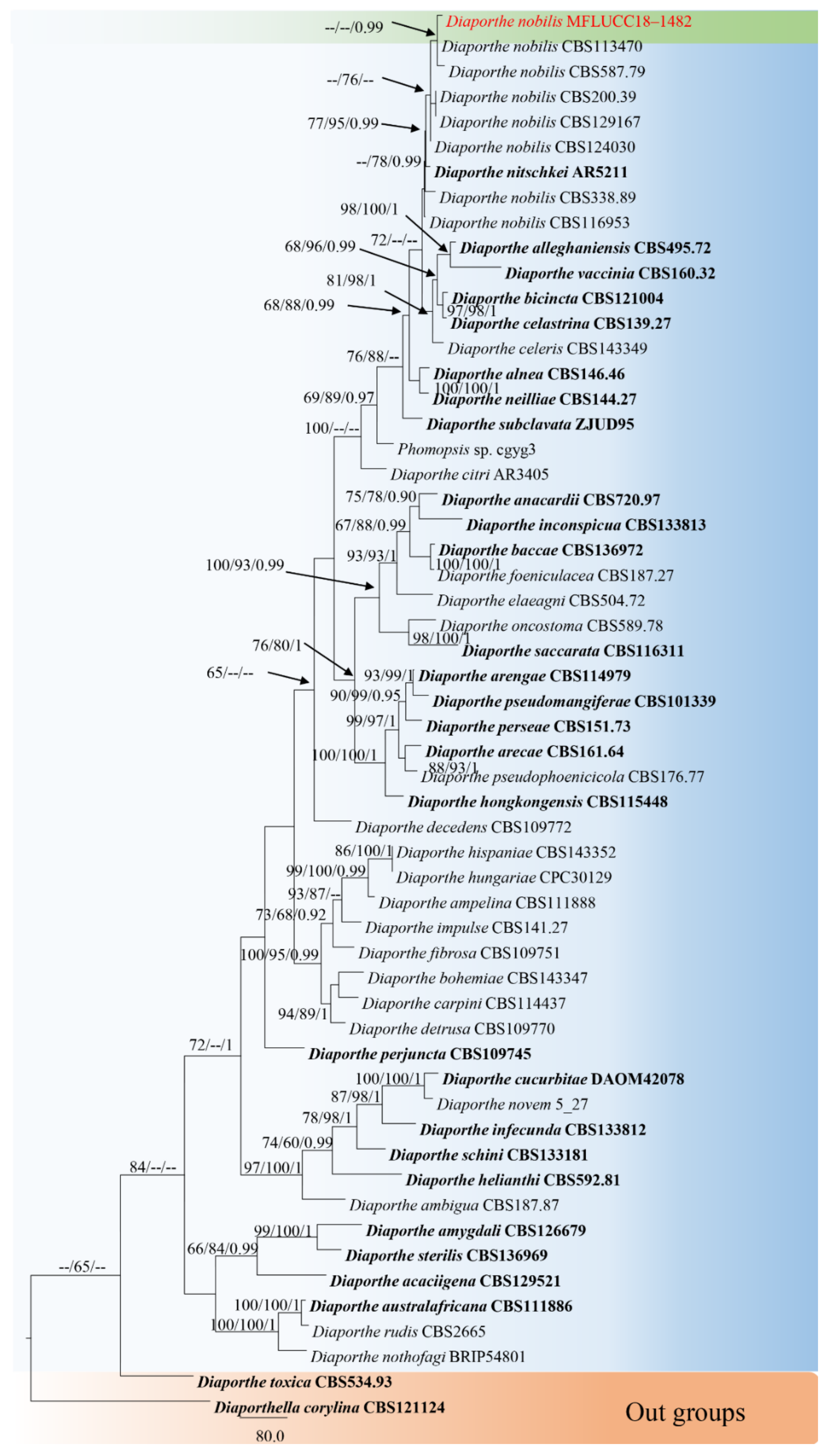
3.1. Phylogenetic Analyses
3.2. Taxonomy
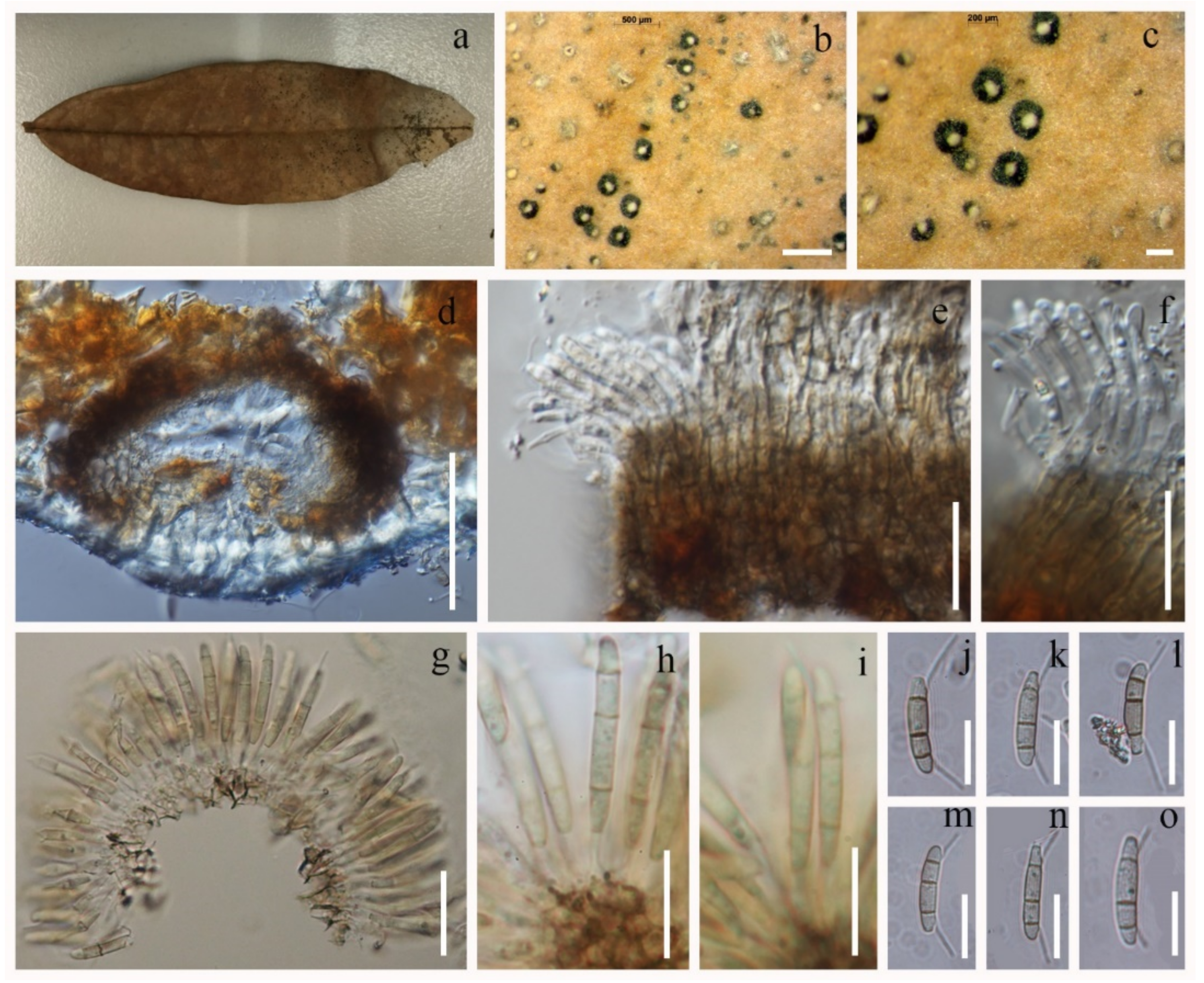
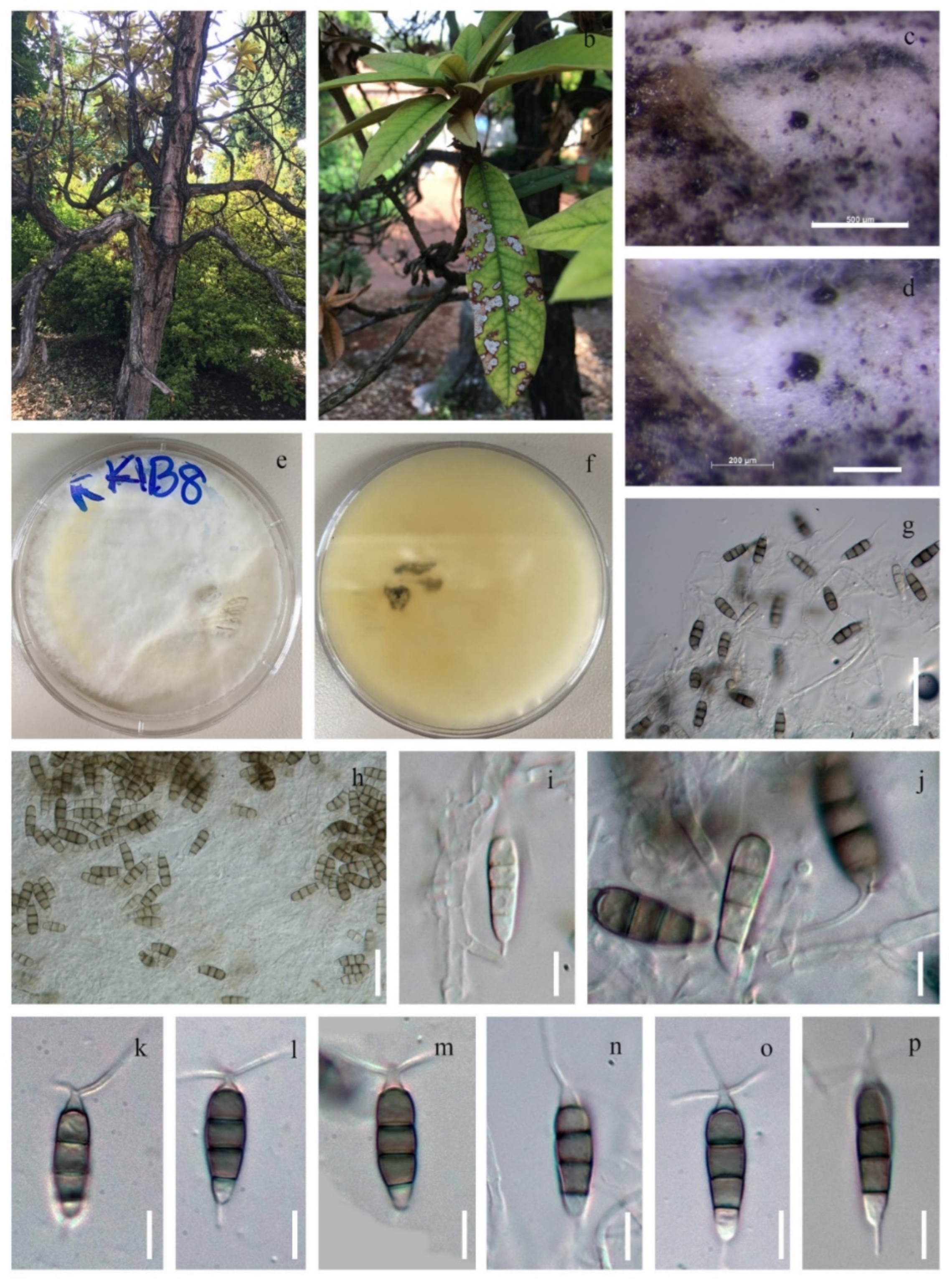
3.2.1. Discosia rhododendricola Chaiwan & K.D. Hyde, sp. Nov. (Figure 4)

3.2.2. Neopestalotiopsis Rhododendricola Chaiwan & K.D. Hyde, sp. Nov. (Figure 6)

3.2.3. Diaporthe nobilis Tanaka & S. Endô, in Endô, J. Pl. Prot. Japan 13: (1927) (Figure 7)

4. Discussion
5. Discosia Species Associated with Rhododendron sp.: Habitat, Known Distribution and Checklist
- Discosia artocreas (Tode) Fr., Summa veg. Scand., Sectio Post. (Stockholm): 423 (1849)= Sphaeria artocreas Tode, F. Meckl. 2: 77, 1791; Fries, Syst. Myc. 2: 523, 1823.
- Discosia blumencronii Bubák, in Handel-Mazzetti, Annln K. K. naturh. Hofmus. Wien 23: 106 (1910)Habitat: Rhododendron ponticum (on dead leaves) [101]Known distribution: Turkey [101]
- Discosia himalayensis Died., Annls mycol. 14(3/4): 218 (1916)= Discosia strobilina Lib. ex Sacc., Syll. Fung. (Abellini) 3: 656 (1884)
- Discosia rhododendri (Speschnew, Monit. Jard. Bot. Tiflis 4: 10 (1906)
- Discosia rhododendricola (This study *)Habitat: Rhododendron sp. (on dead leaves) (This study *)Known distribution: China (This study *)
- Discosia sp.Habitat: Rhododendron sp.* [104]Known distribution: Japan* [104]
- Discosia tricellularis (Okane, Nakagiri & Tad. Ito) F. Liu, L. Cai & Crous, in Liu, Bonthond, Groenewald, Cai & Crous, Stud. Mycol. 92: 322 (2018) (2019)Habitat: Rhododendron indicum [105]Known distribution: Japan [105]
- Discosia vagans De Not., Atti Acad. Tor.: 354 (1849)
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mainra, A.; Badola, H.K.; Mohanty, B. Rhododendrons: Conservation and Sustainable Use International Conference; Government of Sikkim: Gangtok-Sikkim, India, 2010.
- Sekar, K.C.; Srivastava, S.K. Rhododendrons in Indian Himalayan Region: Diversity and Conservation. Am. J. Plant Sci. 2010, 1, 131–137. [Google Scholar] [CrossRef]
- Samant, S.S.; Dhar, U. Diversity, endemism and economic potential of wild edibles plants of Indian Himalaya. Int. J. Sustain. Dev. World Ecol. 1997, 4, 179–191. [Google Scholar] [CrossRef]
- Negi, V.S.; Maikhuri, R.K.; Rawat, L.S.; Chandra, A. Bioprospecting of Rhododendron arboreum for Livelihood Enhancement in Central Himalaya. Indian J. Sci. Technol. 2013, 8, 61–70. [Google Scholar]
- Crane, P.E. Rust fungi on rhododendrons in Asia: Diaphanopellis forrestii gen. et sp. nov., new species of Caeoma, and expanded descriptions of Chrysomyxa dietelii and C. succinea. Mycologia 2005, 97, 534–548. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Viderira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe, a genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef]
- Kierpiec-Baran, B.; Zolna, M.; Kowalik, M. Diversity of fungi colonizing leaves of rhododendron (Rhododendron L.) cuttings. Acta Agrobot. 2014, 67, 21–26. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Huang, S.K.; Abdel-Wahab, M.A.; Daranagama, D.A.; Dayarathne, M.; D’souza, M.J.; Goonasekara, I.D.; et al. Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Tanaka, K.; Endo, M.; Hirayama, K.; Okeanae, I.; Hosoya, T.; Sato, T. Phylogeny of Discosia and Seimatosporium, and introduction of Adisciso and Immersidiscosia genera nova. Persoonia 2011, 26, 85–98. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Bhat, J.D.; Gareth Jones, E.B.; McKenzie, E.H.C.; Dai, D.Q.; Daranagama, D.A.; Dayarathne, M.C.; Goonasekara, I.D.; et al. Towards unraveling relationships in Xylariomycetidae (Sordariomycetes). Fungal Divers. 2015, 73, 73–144. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Gardiennet, A.; Voglmayr, H. Resolution of morphology-based taxonomic delusions: Acrocordiella, Basiseptospora, Blogiascospora, Clypeosphaeria, Hymenopleella, Lepteutypa, Pseudapiospora, Requienella, Seiridium and Strickeria. Persoonia 2016, 37, 82–105. [Google Scholar] [CrossRef]
- Libert, M.A. Plantae Cryptogamae, quas in Arduenna collegit. Fasc 1837, 4, 301–400. [Google Scholar]
- Liu, F.; Bonthond, G.; Groenewald, J.Z.; Cai, L.; Crous, P.W. Sporocadaceae, a family of coelomycetous fungi with appendagebearing conidia. Stud. Mycol. 2019, 92, 287–415. [Google Scholar] [CrossRef]
- Index Fungorum. 2022. Available online: http://www.indexfungorum.org/names/Names.asp (accessed on 26 June 2022).
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Gareth Jones, E.B.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Gerardo-Lugo, S.S.; Tovar-Pedraza, J.M.; Maharachchikumbura, S.S.N.; Apodaca-Sánchez, M.A.; Correia, K.C.; Sauceda-Acosta, C.P.; Camacho-Tapia, M.; Hyde, K.D.; Marraiki, N.; Elgorban, A.M.; et al. Characterization of Neopestalotiopsis species associated with mango grey leaf spot disease in Sinaloa, Mexico. Pathogens 2020, 9, 788. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Bhat, D.J.; Dayarathne, M.C.; Huang, S.K.; Norphanphoun, C.; Senanayake, I.C.; Perera, R.H.; et al. Families of Sordariomycetes. Fungal Divers. 2016, 79, 1–317. [Google Scholar] [CrossRef]
- Hyfe, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar]
- Norphanphoun, C.; Jayawardena, R.S.; Chen, Y.; Wen, T.C.; Meepol, W.; Hyde, K.D. Morphological and phylogenetic characterization of novel pestalotioid species associated with mangroves in Thailand. Mycosphere 2019, 10, 531–578. [Google Scholar] [CrossRef]
- Liu, X.; Tibpromma, S.; Zhang, F.; Xu, J.; Kandawatte, T.C.; Karunarathna, S.C.; Mortimer, P.E. Neopestalotiopsis cavernicola sp. nov. from Gem Cave in Yunnan Province, China. Phytotaxa 2021, 512, 1–27. [Google Scholar] [CrossRef]
- Nitschke, T. Pyrenomycetes Germanici; Eduard Trewendt Breslau: Germany, 1870; Volume 2, pp. 161–320. [Google Scholar]
- Kishi, K. (Ed.) Plant Diseases in Japan; Zenkoku Noson Kyoiku Kyokai: Tokyo, Japan, 1998. (In Japanese) [Google Scholar]
- Kajitani, Y.; Kanematsu, S. Diaporthe kyushuensis sp. nov., the teleomorph of the causal fungus of grapevine swelling arm in Japan, and its anamorph Phomopsis vitimegaspora. Mycoscience 2000, 41, 111–114. [Google Scholar] [CrossRef]
- Katsumoto, K. List of Fungi Recorded in Japan; Kanto Branch of the Mycological Society of Japan: Chiba, Japan, 2010. (In Japanese) [Google Scholar]
- Ando, Y.; Masuya, H.; Aikawa, T.; Ichihara, M. Diaporthe toxicodendri sp. nov., a causal fungus of the canker disease on Toxicodendron vernicifluum in Japan. Mycosphere 2017, 8, 1157–1167. [Google Scholar] [CrossRef]
- Li, Y.; Tan, P.; Zhao, D.G. Diaporthe nobilis, a new record on Camellia sinensis in Guizhou Province, China. Mycosphere 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Groenewald, J.Z.; Woodhall, J.; Armengol, J.; Cinelli, T.; Eichmerier, A.; Ezra, D.; Fontaine, F.; Gramaje, D.; Gutierrez-Aguirregabiria, A. Diaporthe diversity and pathogenicity revealed from a broad survey of grapevine diseases in Europe. Persoonia 2018, 40, 135–153. [Google Scholar] [CrossRef]
- Sessa, L.; Abreo, E.; Bettucci, L.; Lupo, S. Diversity and virulence of Diaporthe species associated with wood disease symptoms in deciduous fruit trees in Uruguay. Phytopathol. Mediterr. 2017, 56, 431–444. [Google Scholar]
- Udayanga, D.; Liu, X.Z.; Crous, P.W.; McKenzie, E.H.C.; Chukeatirote, E.; Hyde, K.D. A multi-locus phylogenetic evaluation of Diaporthe (Phomopsis). Fungal Divers. 2012, 56, 157–171. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Camporesi, E.; Hyde, K.D.; Zhang, W.; Yan, J.Y.; Li, X.H. Molecular phylogenetic analysis reveals seven new Diaporthe species from Italy. Mycosphere 2017, 8, 853–877. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernandez-Restrepo, M.; Wingfield, M.J.; Akulov, A.; Carnegie, A.J.; Cheewangkoon, R.; Gramaje, D.; Groenewald, J.Z.; Guarnaccia, V.; Halleen, F. Genera of phytopathogenic fungi: GOPHY 2. Stud. Mycol. 2019, 92, 47–133. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Pem, D.; Dissanayake, L.; Wijesinghe, S.N.; Bundhun, D.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, D.J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The faces of fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosp 2016, 11, 1669–1677. [Google Scholar] [CrossRef]
- White, T.J.; Burns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Chaiwan, N.; Maharachchikumbura, S.S.N.; Wanasinghe, D.N.; Doilom, M.; Jayaeardena, R.S.; Hyde, K.D. First sexual morph record of Sarcopodium vanilla. Mycotaxon 2020, 134, 707–717. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.N.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2653–2677. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acid Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2017, 30, 772–780. [Google Scholar] [CrossRef]
- Bundhun, D.; Jeewon, R.; Senanayake, I.C.; Camporesi, E.; Aluthmuhandiram, J.V.S.; Tang, A.M.C.; Kang, J.C.; Bhoyroo, V.; Hyde, K.D. Morpho-molecular characterization of Discosia ravennica sp. nov. and a new host record for Sporocadus rosigena. MycoKeys 2021, 79, 173–192. [Google Scholar] [CrossRef]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Kishino, H.; Hasegawa, M. Evaluation of the maximum likelihood estimate of the evolutionary tree topologies from DNA sequence data, and the branching order in hominoidea. J. Mol. Evol. 1989, 29, 170–179. [Google Scholar] [CrossRef]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analyses. BMC Genom. 2002, 3, 4. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J. Tracer v1.4. 2007. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 30 June 2022).
- Rambaut, A. FigTree v1.4: Tree Figure Drawing Tool. 2014. Available online: http://treebio.ed.ac.uk/software/figtree/ (accessed on 30 June 2022).
- Bruen, T.C.; Philippe, H.; Bryant, D. A simple and robust statistical test for detecting the presence of recombination. Genetics 2006, 172, 2665–2681. [Google Scholar] [CrossRef]
- Huson, D.H. Splits Tree: Analyzing and visualizing evolutionary data. Bioinformatics 1998, 14, 68–73. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, Systematic Mycology and Microbiology Laboratory, ARS, USDA. 2022. Available online: http://nt.arsgrin.gov/fungaldatabases (accessed on 10 June 2022).
- Tangthirasunun, N.; Silar, P.; Bhat, D.J.; Chukeatirote, E.; Wijayawardene, D.N.N.; Maharachchikumbura, S.S.N.; Hyde, K.D. Morphology and phylogeny of Pseudorobillarda eucalypti sp. nov., from Thailand. Phytotaxa 2014, 176, 251–259. [Google Scholar] [CrossRef]
- Li, W.J.; Liu, J.K.; Bhat, D.J.; Camporesi, E.; Dai, D.Q.; Mortimer, P.E.; Xu, J.; Hyde, K.D.; Chomnunti, P. Molecular phylogenetic analysis reveals two new species of Discosia from Italy. Phytotaxa 2015, 203, 37–46. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.S.J.; Gene, J.; Guarro, J.; Baseia, I.G.; Garcia, D.; Gusmao, L.F.P.; Souza-Motta, C.M.; et al. Fungal Planet description sheets: 716–784. Persoonia 2018, 40, 240–393. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Cheewangkoon, R.; van der Bank, M.; Swart, W.J.; Stchigel, A.M.; Cano-Lira, J.F.; Roux, J.; Madrid, H.; et al. Fungal Planet description sheets: 154–213. Persoonia 2013, 31, 188–296. [Google Scholar] [CrossRef]
- Hyde, K.D.; Chaiwan, N.; Norphanphoun, C.; Boonmee, S.; Camporesi, E.; Chethana, K.W.T.; Dayarathne, M.C.; de Silva, N.I.; Dissanayake, A.J.; Ekanayaka, A.H.; et al. Mycosphere notes 169–224. Mycosphere 2018, 9, 271–430. [Google Scholar] [CrossRef]
- Greengarten, P.J.; Tuininga, A.R.; Morath, S.U.; Falco, R.C.; Norelus, H.; Daniels, T.J. Occurrence of Soil- and Tick-Borne Fungi and Related Virulence Tests for Pathogenicity to Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2011, 48, 337–344. [Google Scholar] [CrossRef]
- Rahi, P.; Vyas, P.; Sharma, S.; Gulati, A.; Gulati, A. Plant growth promoting potential of the fungus Discosia sp. FIHB 571 from tea rhizosphere tested on chickpea, maize and pea. Indian J. Microbiol. 2009, 49, 128–133. [Google Scholar] [CrossRef]
- Jeewon, R.; Liew, E.C.Y.; Hyde, K.D. Phylogenetic relationships of Pestalotiopsis and allied genera inferred from ribosomal DNA sequences and morphological characters. Mol. Phylogenet. Evol. 2002, 25, 378–392. [Google Scholar] [CrossRef]
- Pereira, J.S.; Costa, R.R.; Nagamoto, N.S.; Forti, L.C.; Pagnocca, F.C.; Rodrigues, A. Comparative analysis of fungal communities in colonies of two leafcutting ant species with different substratum preferences. Fungal Ecol. 2016, 21, 68–75. [Google Scholar] [CrossRef]
- Marčiulynas, A.; Marčiulynienė, D.; Lynikiene, J.; Gedminas, A.; Vaiciukyne, M.; Menkis, A. Fungi and Oomycetes in the Irrigation Water of Forest Nurseries. Forests 2020, 11, 459. [Google Scholar] [CrossRef]
- Wijayawardene, D.N.N.; Goonasekara, I.D.; Camporesi, E.; Wang, Y.; An, Y.L. Two new Seimatosporium species from Italy. Mycosphere 2016, 7, 204–213. [Google Scholar] [CrossRef]
- Kumar, V.; Cheewangkoon, R.; Gentekaki, E.; Maharachchikumbura, S.S.N.; Brahmanage, R.S.; Hyde, K.D. Neopestalotiopsis alpapicalis sp. nov. a new endophyte from tropical mangrove trees in Krabi Province (Thailand). Phytotaxa 2019, 393, 251–262. [Google Scholar] [CrossRef]
- Bezerra, J.D.P.; Machado, A.R.; Firmino, A.L.; Rosado, A.W.C.; de Souza, C.A.F.; de Souza-Motta, C.M.; de Sousa Freire, K.T.L.; Paiva, L.M.; Magalhaes, O.M.C.; Pereira, O.L. Mycological diversity description I. Acta Bot. Bras. 2018, 32, 656–666. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840-928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Lei, C.; Chukeatirote, E.; Wu, W.; Sun, X.; Crous, P.W.; Bhat, D.J.; McKenzie, E.; Bahkali, A. A multi-locus backbone tree for Pestalotiopsis, with a polyphasic characterization of 14 new species. Fungal Divers. 2012, 56, 95–129. [Google Scholar] [CrossRef]
- Song, Y.; Geng, N.; Hyde, K.D.; Zhao, W.S.; Wei, J.G.; Kang, J.C.; Wang, Y. Two new species of Pestalotiopsis from Southern China. Phytotaxa 2013, 126, 22–30. [Google Scholar] [CrossRef]
- Ma, X.Y.; Maharachchikumbura, S.S.N.; Chen, B.W.; Hyde, K.D.; McKenzie, E.H.C.; Chomnunti, P.; Kang, J.C. Endophytic pestaloid taxa in Dendrobium orchids. Phytotaxa 2019, 419, 268–286. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Le Roux, J.J.; Richardson, D.M.; Strasberg, D.; Shivas, R.G.; Alvardo, P.; Edwards, J.; Moreno, G.; Sharma, R.; et al. Fungal Planet description sheets: 371–399. Persoonia 2015, 35, 264–327. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Chukeatirote, E.; Hyde, K.D. Improving the backbone tree for the genus Pestalotiopsis; addition of P. steyaertii and P. magna sp. nov. Mycol. Prog. 2014, 13, 617–624. [Google Scholar] [CrossRef]
- Huanluek, N.; Jjayawardena, R.S.; Maharachchikumbura, S.S.N.; Harishchandra, D.L. Additions to pestalotioid fungi in Thailand: Neopestalotiopsis hydeana sp. nov. and Pestalotiopsis hydei sp. nov. Phytotaxa 2021, 479, 23–43. [Google Scholar] [CrossRef]
- Ayoubi, N.; Soleimani, M.J. Strawberry fruit rot caused by Neopestalotiopsis iranensis sp. nov. and N. mesopotamica. Curr. Microbiol. 2016, 72, 329–336. [Google Scholar] [CrossRef]
- Akinsanmi, O.A.; Nisa, S.; Jeff-Ego, O.S.; Shivas, R.G.; Drenth, A. Dry flower disease of Macadamia in Australia caused by Neopestalotiopsis macadamiae sp. nov. and Pestalotiopsis macadamiae sp. nov. Plant Dis. 2016, 101, 45–53. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.H.; Gilchrist, C.L.M.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano-Lira, J.F.; Valenzuela-Lopez, N.; et al. Fungal Planet description sheets: 1042–1111. Persoonia 2020, 44, 301–459. [Google Scholar] [CrossRef]
- Silvério, M.L.; Cavalcanti, M.A.Q.; Silva, G.A.; Oliveira, R.J.V.; Bezerra, J.L. A new epifoliar species of Neopestalotiopsis from Brazil. Agrotropica 2016, 28, 151–158. [Google Scholar] [CrossRef]
- Jiang, N.; Bonthond, G.; Fan, X.L.; Tian, C.M. Neopestalotiopsis rosicola sp. nov. causing stem canker of Rosa chinensis in China. Mycotaxon 2018, 133, 271–283. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Chukeatirote, E.; Mckenzie, E.H.C.; Hyde, K.D. A destructive new disease of Syzygium samarangense in Thailand caused by the new species Pestalotiopsis amarangensis. Trop. Plant Pathol. 2013, 38, 227–235. [Google Scholar] [CrossRef]
- Jiang, N.; Fan, X.L.; Tian, C.M. Identification and Characterization of Leaf-Inhabiting Fungi from Castanea Plantations in China. J. Fungi 2021, 7, 64. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Liu, M.; Maharachchikumbura, S.S.N.; Zhang, W.; Xing, Q.; Hyde, K.D.; Nilthong, S.; Li, X.; Yan, J. Neopestalotiopsis vitis sp. nov. causing grapevine leaf spot in China. Phytotaxa 2016, 258, 63–74. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Zhang, Y.M.; Wang, Y.; Hyde, K.D. Pestalotiopsis anacardiacearum sp. nov. (Amphisphaeriaceae) has an intricate relationship with Penicillaria jocosatrix, the mango tip borer. Phytotaxa 2013, 99, 49–57. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Hyde, K.D. Species limits in Diaporthe: Molecular re-assessment of D. citri, D. cytosporella, D. foeniculina and D. rudis. Persoonia 2014, 32, 83–101. [Google Scholar] [CrossRef]
- Tan, Y.P.; Edwards, J.; Grice, K.R.E.; Shivas, R.G. Molecular phylogenetic analysis reveals six new species of Diaporthe from Australia. Fungal Divers. 2013, 61, 251–260. [Google Scholar] [CrossRef]
- Santos, J.M.; Vrandecic, K.; Cosic’, J.; Duvnjak, T.; Phillips, A.J.L. Resolving the Diaporthe species occurring on soybean in Croatia. Persoonia 2011, 27, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Lombard, L.; van Leeuwen, G.C.M.; Guarnaccia, V.; Polizzi, G.; van Rijswick, P.C.J.; Rosendahl, K.; Gabler, J.; Crous, P.W. Diaporthe species associated with Vaccinium, with specific reference to Europe. Phytopathol. Mediterr. 2014, 53, 85–97. [Google Scholar]
- Huang, F.; Hou, X.; Dewdney, M.M.; Fu, Y.; Chen, G.; Hyde, K.D.; Li, H. Diaporthe species occurring on citrus in China. Fungal Divers. 2013, 61, 237–250. [Google Scholar] [CrossRef]
- Toulouse, N. Morelet in Ann. Soc. Sci. nat. 1968, Archeol, Toulon and Var, 20, p. 104. Available online: http://sftp.kew.org/pub/data-repositories/LibriFungorum/IXF3/IXF3-550.jpg (accessed on 26 June 2022).
- Shaw, C.G. Host fungus index for the Pacific Northwest. I. Hosts. Wash. State Univ. Agric. Exp. Sta. Bull. 1973, 765, 1–121. [Google Scholar]
- Eglitis, M.; Gould, C.J.; Johnson, F. Fungi found on Ericaceae in the Pacific coastal area. Wash. State Univ. Agric. Exp. Sta. Bull. 1966, 675, 21. [Google Scholar]
- United States Agricultural Research Service; Crops Research Division. Index of Plant Diseases in the United States; Agriculture handbook; US Department of Agriculture: Annapolis, MD, USA, 1960; pp. 1–531.
- Tassi, F. Origine e sviluppo delle Leptostromacee e loro rapporti con le famiglie affini. Bull. Lab. Orto Bot. Reale Univ. Siena 1904, 6, 3–124. [Google Scholar]
- Huseyinov, E.; Selcuk, F. Contribution to study of mycoflora of Turkey I. Coelomycetes of orders Melanconiales and Sphaeropsidales on forest trees and shrubs in the Black Sea coast (Rize and Trabzon Provinces). Mikol. Fitopatol. 2001, 35, 28–33. [Google Scholar]
- Dennis, R.W.G. Fungi of the Hebrides; Royal Botanic Gardens, Kew: Richmond, UK, 1986; pp. 1–383. [Google Scholar]
- Nag Raj, T.R. Coelomycetous Anamorphs with Appendage-Bearing Conidia; Mycologue Publications: Waterloo, ON, Canada, 1993; pp. 1–1101. [Google Scholar]
- Sydow, P.; Sydow, H.; Butler, E.J. Fungi Indiae Orientalis. Ann. Mycol. 1916, 14, 177–220. [Google Scholar]
- Mathur, R.S. The Coelomycetes of India; Bishen Singh Mahendra Pal Singh: Delhi, India, 1979; pp. 1–460. [Google Scholar]
- Kobayashi, T. Index of Fungi Inhabiting Woody Plants in Japan; Host, Distribution and Literature; Zenkoku-Noson-Kyoiku Kyokai Publishing Co., Ltd.: Tokyo, Japan, 2007; pp. 1–1227. [Google Scholar]
- Kirk, P.M.; Spooner, B.M. An account of the fungi of Arran, Gigha and Kintyre. Kew Bull. 1984, 38, 503–597. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Culture Collection No. | Substrate/Host | Country | GenBank Accession No | References | ||||
|---|---|---|---|---|---|---|---|---|---|
| LSU | ITS | TUB2 | TEF1 | RPB2 | |||||
| Discosia pleurochaeta | KT 2179 | - | - | KT281912 | KT284775 | - | - | - | [9] |
| Discosia pleurochaeta | KT 2188 | - | - | AB593713 | AB594781 | AB594179 | - | - | [9] |
| Discosia pleurochaeta | KT 2192 | - | - | AB593714 | AB594782 | AB594180 | - | - | [9] |
| Discosia artocreas | CBS 124848 | Fagus sylvatica | Germany | MH554213 | MH553994 | MH554662 | MH554420 | MH554903 | [13] |
| Discosia brasiliensis | MFLUCC 12-0429 | Dead leaf | Thailand | KF827436 | KF827432 | KF827469 | KF827465 | KF827473 | [59] |
| Discosia brasiliensis | MFLUCC 12-0431 | Dead leaf | Thailand | KF827437 | KF827433 | KF827470 | KF827466 | KF827474 | [59] |
| Discosia brasiliensis | MFLUCC 12-0435 | Dead leaf | Thailand | KF827438 | KF827434 | KF827471 | KF827467 | KF827475 | [59] |
| Discosia fagi | MFLU 14-0299A | Fagus sylvatica | Italy | KM678048 | KM678040 | - | - | - | [60] |
| Discosia fagi | MFLU 14-0299B | Fagus sylvatica | Italy | KM678047 | KM678039 | - | - | - | [60] |
| Discosia fagi | MFLU 14-0299C | Fagus sylvatica | Italy | KM678048 | KM678040 | - | - | - | [60] |
| Discosia italica | MFLU 14-0298B | Fagus sylvatica | Italy | KM678045 | KM678042 | - | - | - | [60] |
| Discosia italica | MFLU 14-0298C | Fagus sylvatica | Italy | KM678044 | KM678041 | - | - | - | [60] |
| Discosia macrozamiae | CPC 32109 | - | - | MH327856 | MH327820 | MH327895 | MH327884 | - | [61] |
| Discosia muscicola | CBS 109.48 | - | - | MH867828 | - | - | - | - | [62] |
| Discosia neofraxinea | NTIT 469 | Fagus sylvatica | Italy | KF827439 | KF827435 | KF827472 | KF827468 | KF827476 | [59] |
| Discosia neofraxinea | MFLUCC 13-0204 | Fagus sylvatica | Italy | KR072672 | KR072673 | - | - | - | [10] |
| Discosia pseudoartocreas | CBS 136438 | Tilia sp. | Austria | KF777214 | KF777161 | MH554672 | MH554430 | MH554913 | [63] |
| Discosia pini | MAFF 410149 | Pinus densiflora | Japan | AB593708 | AB594776 | AB594174 | - | - | [9] |
| Discosia querci | MFLUCC 16-0642 | - | - | MG815830 | MG815829 | - | - | - | [64] |
| Discosia ravennica | MFLU 18-0131 | Pyrus sp. | Italy | MT376617 | MT376615 | MT393594 | - | - | [46] |
| Discosia rhododendricola | KUN-HKAS 123205 | Rhododendron sp. | China | MT741963 | MT741959 | - | - | MW143037 | This study |
| MFLU20-0486 | Rhododendron sp. | China | OP162409 | OP162414 | - | - | OP169687 | This study | |
| Discosia rubi | CBS 143893 | Rubus phoenicolasius | USA | MH554334 | MH554131 | MH554804 | MH554566 | MH555038 | [13] |
| Discosia sp. | F 233 | - | - | - | KU751876 | - | - | - | [13] |
| Discosia sp. | 3T30CF | - | - | - | FJ861385 | - | - | - | [65] |
| Discosia sp. | 3T9A | - | - | - | FJ861386 | - | - | - | [65] |
| Discosia sp. | 3T9C | - | - | - | FJ861387 | - | - | - | [65] |
| Discosia sp. | FIHB 571 | - | - | - | DQ536523 | - | - | - | [66] |
| Discosia sp. | HKUCC 6626 | - | - | AF382381 | AF405303 | - | - | - | [67] |
| Discosia sp. | JSP0111c42 | - | - | - | KR093849 | - | - | - | [68] |
| Discosia sp. | KT 2193 | - | - | AB593706 | AB594774 | - | - | - | [9] |
| Discosia sp. | OT1 143c | - | - | - | KT804147 | - | - | - | [13] |
| Discosia sp. | OT2 143a | - | - | - | KT804075 | - | - | - | [13] |
| Discosia sp. | OT3 176b | - | - | - | KT804146 | - | - | - | [13] |
| Discosia sp. | P4 A7 53 | - | - | - | KU325138 | - | - | - | [13] |
| Discosia sp. | P8 A7-852 | - | - | - | KU325418 | - | - | - | [13] |
| Discosia sp. | R 158 | - | - | - | JN689956 | - | - | - | [13] |
| Discosia sp. | UNH ID260 | - | - | - | KX459431 | - | - | - | [13] |
| Discosia sp. | UWR 012 | - | - | - | KX426948 | - | - | - | [13] |
| Discosia sp. | UWR 040 | - | - | - | KX426977 | - | - | - | [13] |
| Discosia sp. | KT 2109 | - | - | - | MT236494 | - | - | - | [69] |
| Discosia sp. | SH 125 | - | - | - | JF449727 | - | - | - | [13] |
| Discosia sp. | SH 288 | - | - | - | AB594783 | - | - | - | [9] |
| Discosia sp. | MAFF 236709 | - | - | - | KU751876 | - | - | - | [13] |
| Discosia sp. | CBS 241.66 | Acacia karroo | South Africa | MH554244 | MH554022 | MH554698 | - | - | [13] |
| Discosia sp. | CBS 684.70 | Aesculus hippocastanum | Netherlands | MH554277 | MH554064 | MH554740 | - | - | [13] |
| Discosia tricellularis | MAFF 237478 | - | - | AB593730 | AB594798 | AB594189 | - | - | [9] |
| Discosia tricellularis | NBRC 32705 | Rhododendron indicum | Japan | AB593728 | AB594796 | AB594188 | - | - | [9] |
| Discosia yakushimensis | MAFF 242774 | Symplocos prunifolia | Japan | AB593721 | AB594789 | AB594187 | - | - | [9] |
| Sporocadus cornicola | MFLUCC 14-0448 | Cornus sanguinea | Italy | - | KU974967 | - | - | - | [70] |
| Sporocadus rosarum | MFLUCC 14-0466 | Rosa canina | Italy | KT281912 | KT284775 | - | - | - | [70] |
| Species Name | Culture Collection No. | Substrate/Host | Country | GenBank Accession No | References | ||
|---|---|---|---|---|---|---|---|
| ITS | TUB2 | TEF1 | |||||
| Monochaetia ilexae | CBS 101009 | Air | Japan | MH55395 | MH554612 | MH554371 | [13] |
| M. monochaeta | CBS 115004 | Quercus robur | Netherlands | AY853243 | MH554639 | MH554398 | [13] |
| Neopestalotiopsis acrostichi | MFLUCC 17-1754 | Acrostichum aureum | Thailand | MK764272 | MK764338 | MK764316 | [21] |
| N. acrostichi | MFLUCC 17-1755 | Acrostichum aureum | Thailand | MK764273 | MK764339 | MK764317 | [21] |
| N. alpapicalis | MFLUCC 17-2544 | Rhizophora mucronata | Thailand | MK357772 | MK463545 | MK463547 | [71] |
| N. alpapicalis | MFLUCC 17-2545 | Symptomatic Rhizophora apiculata leaves | Thailand | MK357773 | MK463546 | MK463548 | [71] |
| N. aotearoa | CBS 367.54 | Canvas | New Zealand | KM199369 | KM199454 | KM199526 | [15] |
| N. asiatica | MFLUCC 12-0286 | Prunus dulcis | China | JX398983 | JX399018 | JX399049 | [15] |
| N. australis | CBS 114159 | Telopea sp. | Australia | KM199348 | KM199432 | KM199537 | [15] |
| N. brachiata | MFLUCC 17-555 | Rhizophora apiculata | Thailand | MK764274 | MK764340 | MK764318 | [21] |
| N. brasiliensis | COAD 2166 | Psidium guajava | Brazil | MG686469 | MG692400 | MG692402 | [72] |
| N. cavernicola | KUMCC 20-0269 | Cave | China | MW545802 | MW557596 | MW550735 | [22] |
| N. chiangmaiensis | MFLUCC 18-0113 | Pandanus sp. | Thailand | - | MH412725 | MH388404 | [73] |
| N. chrysea | MFLUCC 12-0261 | Dead leaves | China | JX398985 | JX399020 | JX399051 | [74] |
| N. chrysea | MFLUCC 12-0262 | Dead plant | China | JX398986 | JX399021 | JX399052 | [74] |
| N. clavispora | MFLUCC 12-0280 | Magnolia sp. | China | JX398978 | JX399013 | JX399044 | [74] |
| N. clavispora | MFLUCC 12-0281 | Magnolia sp. | China | JX398979 | JX399014 | JX399045 | [74] |
| N. cocoës | MFLUCC 15-0152 | Cocos nucifera | Thailand | KX789687 | - | KX789689 | [17] |
| N. coffeae-arabicae | HGUP4015 | Coffea arabica | China | KF412647 | KF412641 | KF412644 | [75] |
| N. coffeae-arabicae | HGUP4019 | Coffea arabica | China | KF412649 | KF412643 | KF412646 | [75] |
| N. cubana | CBS 600.96 | Leaf Litter | Cuba | KM199347 | KM199438 | KM199521 | [15] |
| N. dendrobii | MFLUCC 14-0099 | Dendrobium cariniferum | Thailand | MK993570 | MK975834 | MK975828 | [76] |
| N. dendrobii | MFLUCC 14-0106 | Dendrobium cariniferum | Thailand | MK993571 | MK975835 | MK975829 | [76] |
| N. egyptiaca | CBS H 22294 | Mangifera indica | Egypt | KP943747 | KP943746 | KP943748 | [77] |
| N. ellipsospora | MFLUCC 12-0283 | Dead plant materials | China | JX398980 | JX399016 | JX399047 | [74] |
| N. eucalypticola | CBS 264.37 | Eucalyptus globulus | - | KM199376 | KM199431 | KM199551 | [15] |
| N. foedans | CGMCC 3.9123 | Mangrove plant | China | JX398987 | JX399022 | JX399053 | [74] |
| N. foedans | CGMCC 3.9178 | Neodypsis decaryi | China | JX398989 | JX399024 | JX399055 | [74] |
| N. formicidarum | CBS 115.83 | Plant debris | Cuba | KM199344 | KM199444 | KM199519 | [78] |
| N. formicidarum | CBS 362.72 | Dead Formicidae (ant) | Cuba | KM199358 | KM199455 | KM199517 | [78] |
| N. honoluluana | CBS 111535 | Telopea sp. | USA | KM199363 | KM199461 | KM199546 | [15] |
| N. honoluluana | CBS 114495 | Telopea sp. | USA | KM199364 | KM199457 | KM199548 | [15] |
| N. hydeana | MFLUCC 20-0132 | Artocarpus heterophyllus | Thailand | MW266069 | MW251119 | MW251129 | [79] |
| N. iranensis | CBS 137767 | Fragaria ananassa | Iran | KM074045 | KM074056 | KM074053 | [80] |
| N. iranensis | CBS 137768 | Fragaria ananassa | Iran | KM074048 | KM074057 | KM074051 | [81] |
| N. javaensis | CBS 257.31 | Cocos nucifera | Java | KM199357 | KM199437 | KM199543 | [15] |
| N. keteleeria | MFLUCC 13-0915 | Keteleeria pubescens | China | KJ023087 | KJ023088 | KJ023089 | [75] |
| N. macadamiae | BRIP 63737c | Macadamia integrifolia | Australia | KX186604 | KX186654 | KX186627 | [81] |
| N. macadamiae | BRIP 63742a | Macadamia integrifolia | Australia | KX186599 | KX186657 | KX186629 | [82] |
| N. magna | MFLUCC 12-652 | Pteridium sp. | France | KF582795 | KF582793 | KF582791 | [15] |
| N. mesopotamica | CBS 299.74 | Eucalyptus sp. | Turkey | KM199361 | KM199435 | KM199541 | [15] |
| N. mesopotamica | CBS 336.86 | Pinus brutia | Iraq | KM199362 | KM199441 | KM199555 | [15] |
| N. musae | MFLUCC 15-0776 | Musa sp. | Thailand | KX789683 | KX789686 | KX789685 | [17] |
| N. natalensis | CBS 138.41 | Acacia mollissima | South Africa | KM199377 | KM199466 | KM199552 | [15] |
| N. nebuloides | BRIP 66617 | Sporobolus elongatus | Australia | MK966338 | MK977632 | MK977633 | [82] |
| N. pandanicola | KUMCC 17-0175 | Pandanus sp. | China | - | MH412720 | MH388389 | [73] |
| N. pernambucana | URM7148 | Vismia guianensis | Brazil | KJ792466 | - | KU306739 | [83] |
| N. pernambucana | RV02 | Vismia guianensis | Brazil | KJ792467 | - | KU306740 | [83] |
| N. petila | MFLUCC 17-1737 | Rhizophora mucronata | Thailand | MK764275 | MK764341 | MK764319 | [21] |
| N. petila | MFLUCC 17-1738 | Rhizophora mucronata | Thailand | MK764276 | MK764342 | MK764320 | [21] |
| N. phangngaensis | MFLUCC 18-0119 | Pandanus sp. | Thailand | MH388354 | MH412721 | MH388390 | [73] |
| N. piceana | CBS 254.32 | Cocos nucifera | Indonesia | KM199372 | KM199452 | KM199529 | [15] |
| N. piceana | CBS 394.48 | Picea sp. | UK | KM199368 | KM199453 | KM199527 | [15] |
| N. protearum | CBS 114178 | Leucospermum cuneiforme cv. “Sunbird” | Zimbabwe | JN712498 | KM199463 | LT853201 | [15] |
| N. rhizophorae | MFLUCC 17-1550 | Rhizophora mucronata | Thailand | MK764277 | MK764343 | MK764321 | [21] |
| N. rhizophorae | MFLUCC 17-1551 | Rhizophora mucronata | Thailand | MK764278 | MK764344 | MK764322 | [21] |
| N. rhododendricola | KUN-HKAS 123204 | Rhododendron sp. | China | OK283069 | OK274147 | OK274148 | This study |
| MFLU20-0046 | Rhododendron sp. | China | OP11897554 | OP169689 | OP169688 | This study | |
| N. rosae | CBS 101057 | Rosa sp. | New Zealand | KM199359 | KM199429 | KM199523 | [15] |
| N. rosae | CBS 124745 | Paeonia suffruticosa | USA | KM199360 | KM199430 | KM199524 | [15] |
| N. rosicola | CFCC 51992 | Rosa chinensis | China | KY885239 | KY885245 | KY885243 | [84] |
| N. rosicola | CFCC 51993 | Rosa chinensis | China | KY885240 | KY885246 | KY885244 | [84] |
| N. samarangensis | CBS 115451 | Unidentified tree | China | KM199365 | KM199447 | KM199556 | [85] |
| N. saprophytica | MFLUCC 12-0282 | Magnolia sp. | China | JX398982 | JX399017 | JX399048 | [74] |
| N. sichuanensis | CFCC 54338 | Castanea mollissima | China | MW166231 | MW218524 | MW199750 | [86] |
| N. sichuanensis | SM15-1C | Castanea mollissima | China | MW166232 | MW218525 | MW199751 | [86] |
| N. sonneratae | MFLUCC 17-1744 | Sonneronata alba | Thailand | MK764279 | MK764345 | MK764323 | [21] |
| N. sonneratae | MFLUCC 17-1745 | Sonneronata alba | Thailand | MK764280 | MK764346 | MK764324 | [21] |
| N. steyaertii | IMI 192475 | Eucalyptus viminalis | Australia | KF582796 | KF582794 | KF582792 | [15] |
| N. surinamensis | CBS 450.74 | Soil under Elaeis guineensis | Suriname | KM199351 | KM199465 | KM199518 | [15] |
| N. thailandica | MFLUCC 17-1730 | Rhizophora mucronata | Thailand | MK764281 | MK764347 | MK764325 | [21] |
| N. thailandica | MFLUCC 17-1731 | Rhizophora mucronata | Thailand | MK764282 | MK764348 | MK764326 | [21] |
| N. umbrinospora | MFLUCC 12-0285 | Unidentified plant | China | JX398984 | JX399019 | JX399050 | [74] |
| N. vitis | MFLUCC 15-1265 | Vitis vinifera cv. “Summer black” | China | KU140694 | KU140685 | KU140676 | [87] |
| N. vitis | MFLUCC 15-1270 | Vitis vinifera cv. “Kyoho” | China | KU140699 | KU140690 | KU140681 | [87] |
| N. zimbabwana | CBS 111495 | Leucospermum cunciforme | Zimbabwe | JX556231 | KM199456 | KM199545 | [15] |
| Pestalotiopsis adusta | ICMP6088 | On refrigerator door PVC gasket | Fiji | JX399006 | JX399037 | JX399070 | [74] |
| P. adusta | MFLUCC10-0146 | Syzygium sp. | Thailand | JX399007 | JX399038 | JX399071 | [74] |
| P. anacardiacearum | IFRDCC 2397 | Mangifera indica | China | KC247154 | KC247155 | KC247156 | [88] |
| P. humus | CBS 115450 | Ilex cinerea | China | KM199319 | KM199418 | KM199487 | [15] |
| P. humus | CBS 336.97 | Soil | Papua New Guinea | KM199317 | KM199420 | KM199484 | [15] |
| P. hydei | MFLUCC 20135 | Litsea petiolata | Thailand | MW266063 | MW251112 | MW251113 | [79] |
| N. thailandica | MFLUCC 17-1730 | Rhizophora mucronata | Thailand | MK764281 | MK764347 | MK764325 | [21] |
| N. thailandica | MFLUCC 17-1731 | Rhizophora mucronata | Thailand | MK764282 | MK764348 | MK764326 | [21] |
| N. umbrinospora | MFLUCC 12-0285 | Unidentified plant | China | JX398984 | JX399019 | JX399050 | [74] |
| P. hydei | E-72-02 | Eucalyptus grandis | Brazil | KU926708 | KU926716 | KU926712 | [79] |
| P. inflexa | MFLUCC12-0270 | Unidentifified tree | China | JX399008 | JX399039 | JX399072 | [74] |
| P. linearis | MFLUCC12-0271 | Trachelospermumsp. | China | JX398992 | JX399027 | JX399058 | [74] |
| Pseudopestalotiopsis cocos | CBS 272.29 | Cocos nucifera | Indonesia | KM199378 | KM199467 | KM199553 | [15] |
| Ps. indica | CBS 459.78 | Hibiscus rosa-sinensis | India | KM199381 | KM199470 | KM199560 | [15] |
| Ps. theae | MFLUCC12-0055 T | Camellia sinensis | Thailand | JQ683727 | JQ683711 | JQ683743 | [15] |
| Ps. theae | SC011 | Camellia sinensis | Thailand | JQ683726 | JQ683710 | JQ683742 | [15] |
| Species Name | Culture Collection No. | Substrate/Host | Country | GenBank Accession No | References | ||
|---|---|---|---|---|---|---|---|
| ITS | TUB2 | TEF1 | |||||
| Diaporthe acaciigena | CBS 129521 | Acacia retinodes | - | KC343005 | KC343973 | KC343731 | [6] |
| Diaporthe alleghaniensis | CBS 495.72 | Betula alleghaniensis | - | KC343007 | KC343975 | KC343733 | [6] |
| Diaporthe alnea | CBS 146.46 | Alnus sp. | - | KC343008 | KC343976 | KC343734 | [6] |
| Diaporthe ambigua | CBS 187.87 | Helianthus annuus | Italy | KC343015 | KC343983 | KC343741 | [6] |
| Diaporthe ampelina | CBS 111888 | Vaccinium vinifera | USA | KC343016 | KC343984 | KC343742 | [6] |
| Diaporthe amygdali | CBS 126679 | Prunus dulcis | - | KC343022 | KC343990 | AY343748 | [6] |
| Diaporthe anacardii | CBS 720.97 | Anacardium ocidentale | - | KC343024 | KC343992 | KC343750 | [6] |
| Diaporthe arecae | CBS 161.64 | Areca catechu | - | KC343032 | KC344000 | KC343758 | [6] |
| Diaporthe arengae | CBS 114979 | Arenga engleri | - | KC343034 | KC344002 | KC343760 | [6] |
| Diaporthe australafricana | CBS 111886 | Vaccinium vinifera | Australia | KC343038 | KC344006 | KC343764 | [6] |
| Diaporthe baccae | CBS 136972 | Vaccinium corymbosum | - | KJ160565 | - | KJ160597 | [45] |
| Diaporthe bicincta | CBS 121004 | Juglans sp. | - | KC343134 | KC344102 | KC343860 | [6] |
| Diaporthe bohemiae | CBS 143347 | Vitis spp. | Czech Republic | MG281015 | MG281188 | MG281536 | [29] |
| Diaporthe carpini | CBS 114437 | Carpinus betulus | Sweden | KC343044 | KC344012 | KC343770 | [6] |
| Diaporthe celastrina | CBS 139.27 | Celastrus scandens | - | KC343047 | KC344015 | KC343773 | [6] |
| Diaporthe celeris | CBS 143349 | Vaccinium vinifera | UK | MG281017 | MG281190 | MG281538 | [29] |
| Diaporthella corylina | CBS 121124 | Corylus sp. | - | KC343004 | KC343972 | KC343730 | [6] |
| Diaporthe citri | AR 3405 | - | - | KC843311 | KC843187 | KC843071 | [89] |
| Diaporthe cucurbitae | DAOM 42078 | Cucumis sativus | - | KM453210 | KP118848 | KM453211 | [89] |
| Diaporthe decedens | CBS 109772 | Corylus avellana | Austria | KC343059 | KC344027 | KC343785 | [6] |
| Diaporthe detrusa | CBS 109770 | Berberis vulgaris | Austria | KC343061 | KC344029 | KC343787 | [6] |
| Diaporthe elaeagni | CBS 504.72 | Eleagnus sp. | Netherlands | KC343064 | KC344032 | KC343790 | [6] |
| Diaporthe nobilis | KUN-HKAS 123203 | Rhododendron sp. | China | MT741962 | MW150988 | MW248138 | This study |
| Diaporthe nobilis | CBS 338.89 | Hedera helix | - | KC343152 | KC344120 | KC343878 | [6] |
| Diaporthe nobilis | CBS 200.39 | Laurus nobilis | Germany | KC343151 | KC344119 | KC343877 | [6] |
| Diaporthe nobilis | CBS 113470 | - | - | KC343146 | - | - | [6] |
| Diaporthe nobilis | CBS 116953 | - | - | KC343147 | - | - | [6] |
| Diaporthe nobilis | CBS 124030 | - | - | KC343149 | - | - | [6] |
| Diaporthe nobilis | CBS 129167 | - | - | KC343150 | - | - | [6] |
| Diaporthe nobilis | CBS 587.79 | Pinus pantepella | - | KC343153 | KC344121 | KC343879 | [6] |
| Diaporthe fibrosa | CBS 109751 | - | - | KC343099 | KC344067 | KC343825 | [6] |
| Diaporthe foeniculacea | CBS 187.27 | - | - | KC343107 | KC344075 | KC343833 | [6] |
| Diaporthe helianthi | CBS 592.81 | Helianthus annuus | - | KC343115 | KC344083 | KC343841 | [6] |
| Diaporthe nitschkei | AR 5211 | Hedera helix | - | KJ210538 | KJ420828 | KJ210559 | [89] |
| Diaporthe hispaniae | CBS 143351 | - | - | MG281124 | MG281296 | MG281645 | [29] |
| Diaporthe hongkongensis | CBS 115448 | Dichroa febrífuga | - | KC343119 | KC344087 | KC343845 | [6] |
| Diaporthe hungariae | CPC 30129 | - | - | - | - | MG281646 | [29] |
| Diaporthe impulse | CBS 114434 | - | - | KC343122 | KC344089 | KC343847 | [6] |
| Diaporthe inconspicua | CBS 133813 | Maytenus ilicifolia | - | KC343123 | KC344091 | KC343849 | [6] |
| Diaporthe infecunda | CBS 133812 | Schinus terebinthifolius | - | KC343126 | KC344094 | KC343852 | [6] |
| Diaporthe neilliae | CBS 144. 27 | Spiraea sp. | - | KC343144 | KC344112 | KC343870 | [90] |
| Diaporthe nothofagi | BRIP 54801 | Nothofagus cunninghamii | - | JX862530 | KF170922 | JX862536 | [91] |
| Diaporthe novem | CBS 127271 | - | - | HM347710 | - | HM347698 | [6] |
| Diaporthe oncostoma | CBS 589.78 | - | - | KC343162 | KC344130 | KC343888 | [6] |
| Diaporthe perjuncta | CBS 109745 | Ulmus glabra | - | KC343172 | KC344140 | KC343898 | [6] |
| Diaporthe perseae | CBS 151.73 | Persea gratissima | - | KC343173 | KC344141 | KC343899 | [6] |
| Diaporthe pseudomangiferae | CBS 101339 | Mangifera indica | - | KC343181 | KC344149 | KC343907 | [6] |
| Diaporthe pseudophoenicicola | CBS 462.69 | Phoenix dactylifera | - | KC343183 | KC344151 | KC343909 | [6] |
| Diaporthe rudis | CBS 2665 | - | - | - | KM396309 | KM396311 | [6] |
| Diaporthe saccarata | CBS 116311 | Protea repens | - | KC343190 | KC344158 | KC343916 | [6] |
| Diaporthe schini | CBS 133181 | Schinus terebinthifolius | - | KC343191 | KC344159 | KC343917 | [6] |
| Diaporthe sterilis | CBS 136969 | Vaccinium corymbosum | - | KJ160579 | KJ160528 | KJ160611 | [92] |
| Diaporthe subclavata | ZJUD 95 | - | - | KJ490630 | KJ490451 | KJ490509 | [93] |
| Diaporthe toxica | CBS 534.93 | Lupinus angustifolius | - | KC343220 | KC344188 | KC343946 | [6] |
| Diaporthe vaccinii | CBS 160.32 | Vaccinium macrocarpon | - | AF317578 | JX270436 | GQ250326 | [92] |
| Phomopsis sp. | FH 2012b | - | - | JQ954649 | - | JQ954667 | [93] |
| LSU | ITS | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Base Pair Positions | Base Pair Positions | ||||||||||||||||
| 70 | 369 | 379 | 407 | 502 | 646 | 872 | 873 | 939 | 959 | 1003 | 1004 | 1019 | 1039 | 1303 | 1402 | 1404 | |
| D. rhododendricola (KUN-HKAS 123205) | G | A | T | T | G | A | C | T | C | T | A | C | A | T | A | C | T |
| D. macrozamiae CPC 32109 | A | A | T | T | G | G | - | - | C | G | G | T | T | T | A | G | A |
| D. muscicola CBS 109.48 | G | G | T | C | A | A | - | - | - | - | - | - | - | - | - | - | - |
| D. pleurochaeta KT2179 | A | A | C | T | G | G | T | C | C | G | G | T | T | T | A | G | A |
| D. pleurochae KT 2188 | A | A | T | T | G | G | T | C | C | G | G | T | T | T | A | G | A |
| D. pleurochae KT 2192 | A | A | T | T | G | G | T | C | C | G | G | T | T | T | A | G | A |
| D. tricellularis MAFF237478 | - | - | - | - | - | - | - | - | G | G | G | T | T | A | G | G | A |
| D. tricellularis NCBR32705 | - | - | - | - | - | - | - | - | G | G | G | T | T | A | G | G | A |
| D. yakushimensis MAFF 242774 | - | - | - | - | - | - | - | - | C | G | G | T | T | T | A | G | A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaiwan, N.; Jeewon, R.; Pem, D.; Jayawardena, R.S.; Nazurally, N.; Mapook, A.; Promputtha, I.; Hyde, K.D. Fungal Species from Rhododendron sp.: Discosia rhododendricola sp.nov, Neopestalotiopsis rhododendricola sp.nov and Diaporthe nobilis as a New Host Record. J. Fungi 2022, 8, 907. https://doi.org/10.3390/jof8090907
Chaiwan N, Jeewon R, Pem D, Jayawardena RS, Nazurally N, Mapook A, Promputtha I, Hyde KD. Fungal Species from Rhododendron sp.: Discosia rhododendricola sp.nov, Neopestalotiopsis rhododendricola sp.nov and Diaporthe nobilis as a New Host Record. Journal of Fungi. 2022; 8(9):907. https://doi.org/10.3390/jof8090907
Chicago/Turabian StyleChaiwan, Napalai, Rajesh Jeewon, Dhandevi Pem, Ruvishika Shehali Jayawardena, Nadeem Nazurally, Ausana Mapook, Itthayakorn Promputtha, and Kevin D. Hyde. 2022. "Fungal Species from Rhododendron sp.: Discosia rhododendricola sp.nov, Neopestalotiopsis rhododendricola sp.nov and Diaporthe nobilis as a New Host Record." Journal of Fungi 8, no. 9: 907. https://doi.org/10.3390/jof8090907
APA StyleChaiwan, N., Jeewon, R., Pem, D., Jayawardena, R. S., Nazurally, N., Mapook, A., Promputtha, I., & Hyde, K. D. (2022). Fungal Species from Rhododendron sp.: Discosia rhododendricola sp.nov, Neopestalotiopsis rhododendricola sp.nov and Diaporthe nobilis as a New Host Record. Journal of Fungi, 8(9), 907. https://doi.org/10.3390/jof8090907