
The Umbelopsis ramanniana Sensu Lato Consists of Five Cryptic Species


Abstract
1. Introduction
2. Materials and Methods
2.1. Cultures and Isolation
2.2. Media, Cultivation and Morphological Observation
2.3. DNA Extraction, Amplification and Sequence Analyses
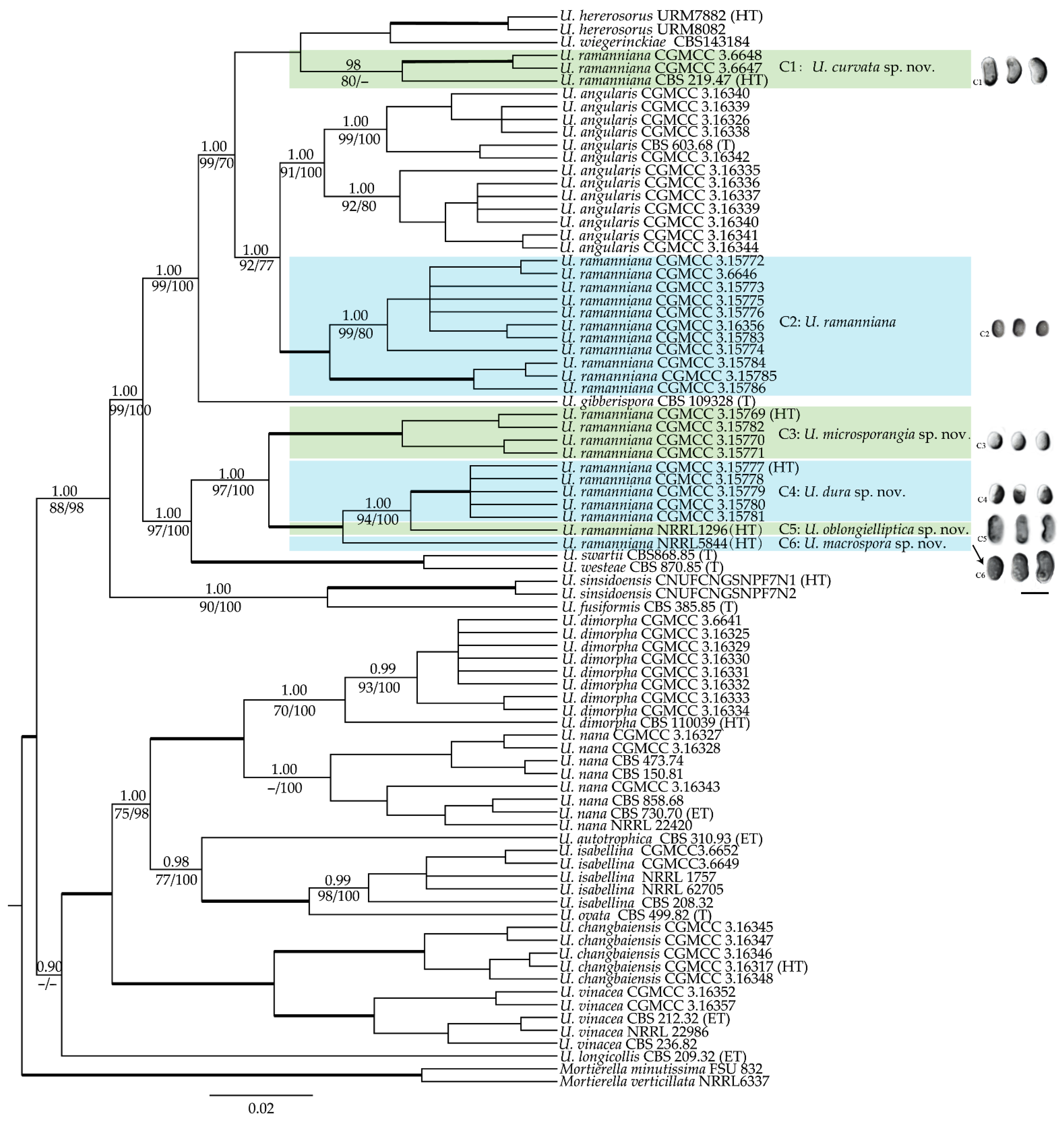
3. Results
3.1. Molecular Phylogenetic Analyses
3.2. Maximum Growth Temperature
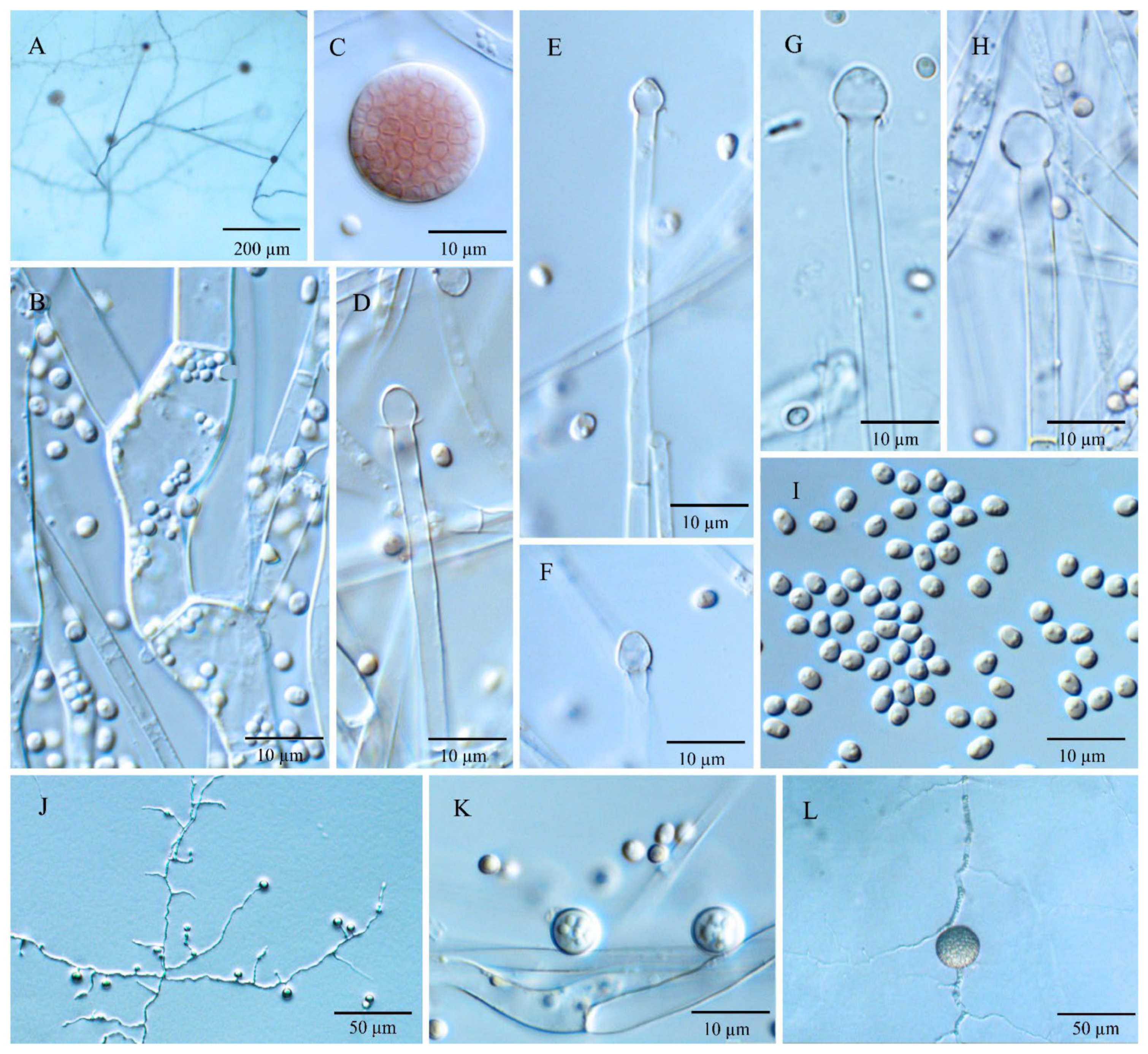
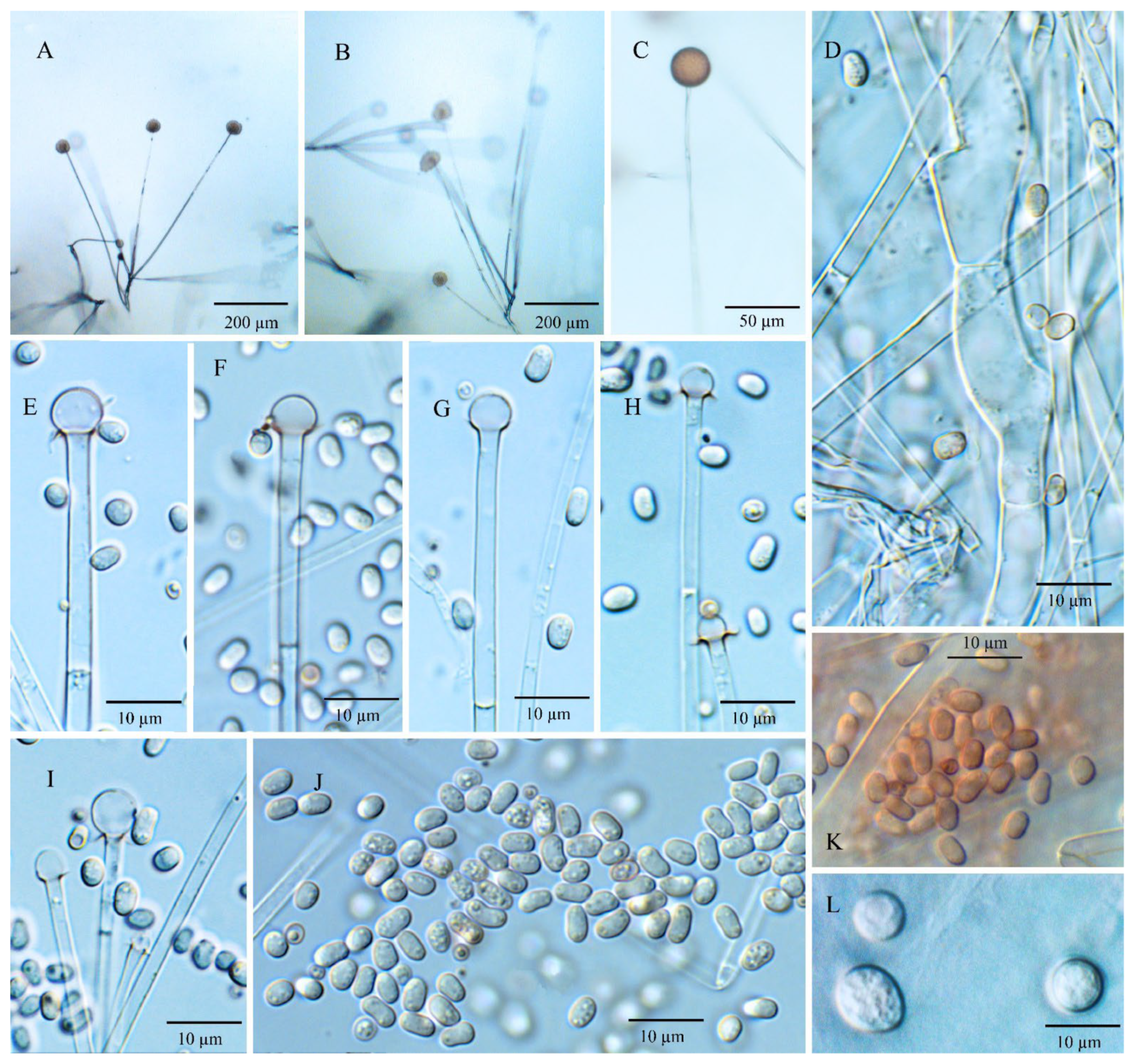
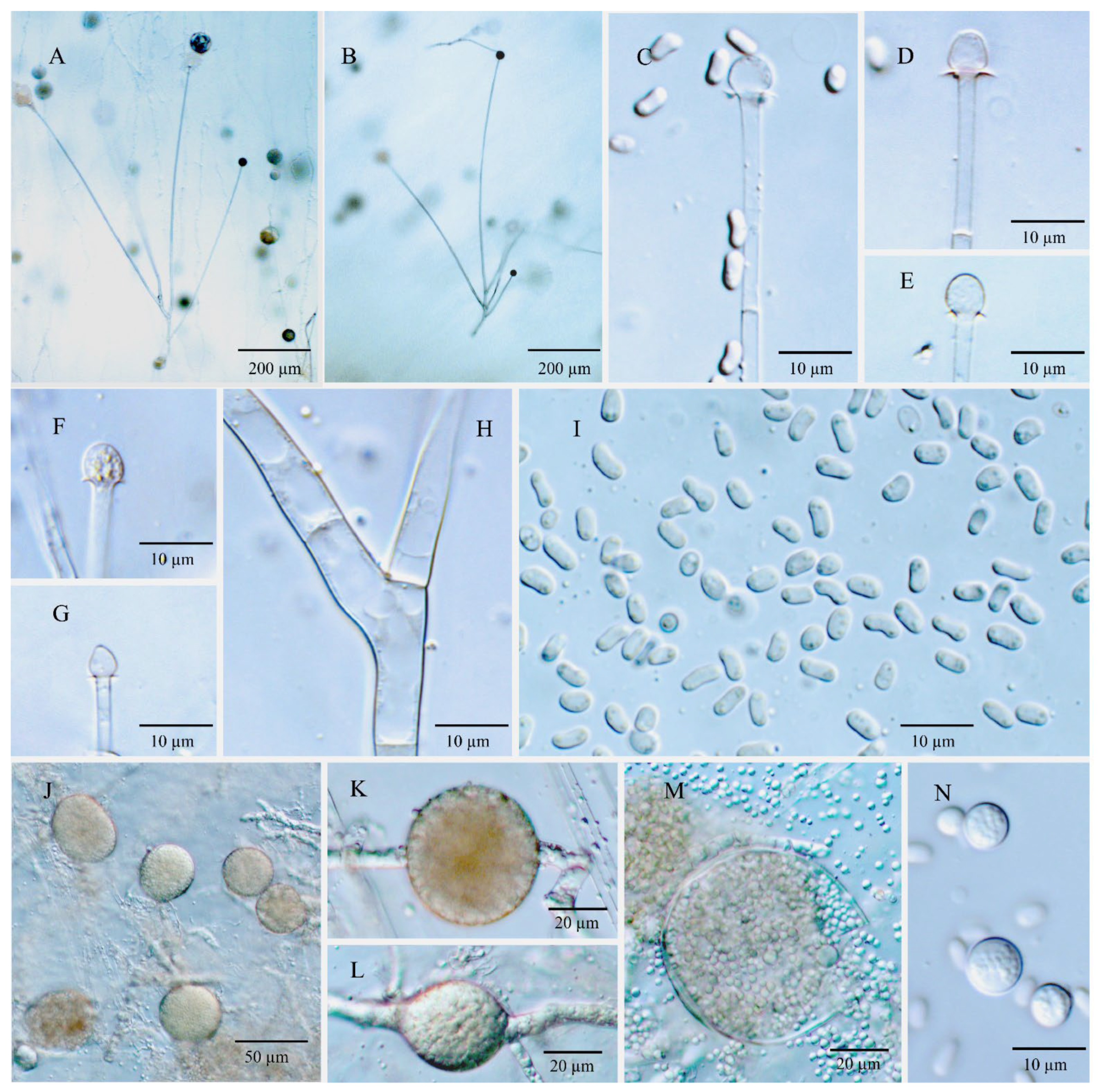
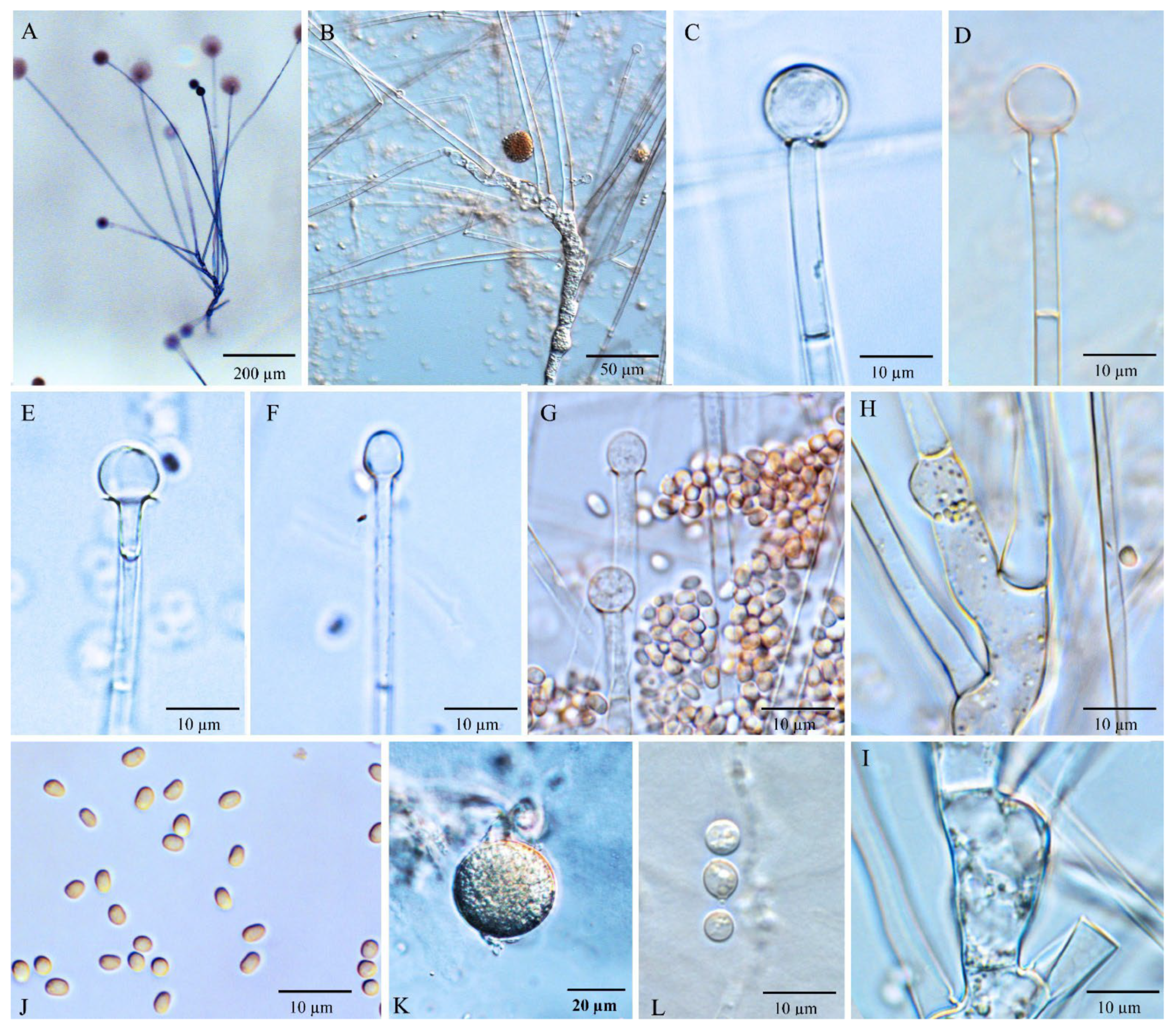
3.3. Morphology and Taxonomy
3.3.1. Umbelopsis curvata Y.N. Wang, X.Y. Liu and R.Y. Zheng, sp. nov.
3.3.2. Umbelopsis dura Y.N. Wang, X.Y. Liu and R.Y. Zheng, sp. nov.
3.3.3. Umbelopsis macrospora Y.N. Wang, X.Y. Liu and R.Y. Zheng, sp. nov.
3.3.4. Umbelopsis microsporangia Y.N. Wang, X.Y. Liu and R.Y. Zheng, sp. nov.
3.3.5. Umbelopsis oblongielliptica Y.N. Wang, X.Y. Liu and R.Y. Zheng, sp. nov.
3.3.6. Umbelopsis ramanniana (Möller) W. Gams, Mycol. Res. 107(3): 349 (2003)
- ≡
- Mucor ramannianus Möller, Z. Forst- u. Jagdw. 35: 330 (1903)
- ≡
- Mortierella ramanniana (Möller) Linnem., Mucor.-Gatt. Mortierella Coem.: 19 (1941)
- ≡
- Micromucor ramannianus (Möller) Arx, Sydowia 35: 19 (1984)
3.3.7. Key to Species of Umbelopsis
| 1. | Sporangia sometimes with neck-like base | Umbelopsis longicollis |
| – | Sporangia without neck-like base | 2 |
| 2. | Colonies drab grey when the sporangia are produced heavily | 3 |
| – | Colonies roseate to almost white when the sporangia are produced heavily | 4 |
| 3. | Sporangia obovate to ovate | U. ovata |
| – | Sporangia globose to subglobose | U. isabellina |
| 4. | With two kinds of sporangia, single- and multi-spored | U. dimorpha |
| – | With one kind of sporangia, single- or multi-spored | 5 |
| 5. | Colony color white to pale pinkish; sporangia single-spored with 4.5–8.0 µm in diam.; sporangiospores globose with the same size of sporangia | U. nana |
| – | Colony color roseate to pale pinkish; sporangia multi-spored larger than 10 µm; sporangiospore globose to oblong-ellipsoidal far smaller than sporangia | 6 |
| 6. | Sporangiospores globose, maximum growth temperature 40 °C | U. autotrophica |
| – | Sporangiospores subglobose, ovoid to oblong-ellipsoidal, angular or irregular; maximum growth temperature no more than 39 °C | 7 |
| 7. | Sporangiophores no more than 100 µm in length | 8 |
| – | Sporangiophores mainly more than 100 µm in length | 10 |
| 8. | Colonies reaching 20–21 mm in diam. on MEA after 7 days at 20 °C; sporangiospores globose, ellipsoidal or angular | U. sinsidoensis |
| – | Colonies reaching more than 30 mm in diam. on MEA after 7 days at 20 °C; sporangiospores angular | 9 |
| 9. | Colonies russet; sporangiophores umbellately branched from distinct vesicles on agar surface, (16–) 23–63 (−80) µm in length; columellae small but distinct 3.2–5.5 × 1.6–3.8; without micro-chlamydospores | U. changbaiensis |
| – | Colonies light russet-vinaceous pinkish; sporangiophores simply branched from slightly swollen stalks, (27.7–) 47.4–94.8 µm in length; columellae absent or slightly convex up to 1 µm diam.; with micro-chlamydospores | U. vinacea |
| 10. | Sporangia fusiform; without columella | U. fusiformis |
| – | Sporangia globose to subglobose; with distinct columellae | 11 |
| 11. | Columellae distinct to inconspicuous and variable in shape and size; sporangiospores variable in shape and size up to 3.5–5 × 5–11 µm | U. heterosporus |
| – | Columellae distinct and uniform; sporangiospores mostly uniform and no more than 6 µm in diam | 12 |
| 12. | Sporangiophores mainly umbellately branched from swollen portion of the subtended stalk | U. wiegerinckiae |
| – | Sporangiophores sympodial branched from slightly swollen stalks | 13 |
| 13. | Sporangiospores angular | U. angularis |
| – | Sporangiospores not angular | 14 |
| 14. | Sporangiospores with appendage | 15 |
| – | Sporangiospores smooth | 17 |
| 15. | Sporangiophores straight, sporangiospores without a narrowed end, walls thickened unilaterally | U. gibberispora |
| – | Sporangiophores sometimes recurved; sporangiospores with a narrowed end, walls thickened or not | 19 |
| 16. | Sporangiospores tear-shaped with 4.0–6.0 ×2.0–2.5 µm | U. swartii |
| – | Sporangiospores oval to clavate with distinctively thickened wall at the narrower end with 4.0–6.0–8.0 × 2.5–3.5 µm | U. westeae |
| 17. | Colony color deep roseate, Indian red to dark vinaceous-brown; sporangial walls slowly deliquescent | U. dura |
| – | Colony color roseate to pinkish, brownish vinaceous to light russet-vinaceous; sporangial walls quickly deliquescent | 18 |
| 18. | Sporangiospores ovoid to ellipsoidal no curved, mostly 2.4–3.5 µm in length | 19 |
| – | Sporangiospores ellipsoidal to oblong-ellipsoidal and sometimes slightly curved to one side, mainly 3.5–4.9 µm in length | 20 |
| 19. | Colony diam. on CMA reaching 35–40 mm in diam. after 10 days at 18 °C; maximum growth temperature 31–32 °C; columellae mostly depressed globose (3.0–) 4.0–6.5 (–10.0) × (2.5–) 3.5–5.5 (–9.0) µm | U. microsporangia |
| – | Colony diam. on CMA reaching 40–44 mm in diam. after 10 days at 18 °C, maximum growth temperature higher than 32 °C; columellae mostly subglobose (4.0–) 6.0–8.0 (–10.0) µm | U. ramanniana |
| 20. | Colony diam. on CMA reaching 60–62 mm in diam. after 10 days at 18 °C; colonies 1 mm in high; sporangiophores(120–) 270–475 (–980) µm in length | U. oblongielliptica |
| – | Colony diam. on CMA reaching 43–48 mm in diam. after 10 days at 18 °C; colonies up to 2 or 3 mm; sporangiophores commonly more than 500 µm in length | 21 |
| 21. | Sporangiospores oblong-ellipsoidal to ellipsoidal, (3.6–) 4.0–4.9 (–5.5) × (1.7–) 2.4–2.8 (–3.2) µm; macro-chlamydospore not formed on CMA and MEA | U. macrospora |
| – | Sporangiospores ellipsoidal (2.7–) 3.2–4.5 (–5.7) × (1.6–) 2.0–2.4 (–2.8) µm in size; macro-chlamydospore abundant on CMA and MEA | U. curvata |
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamisaka, Y.; Yokochi, T.; Nakahara, T.; Suzuki, O. Characterization of the diacylglycerol acyltransferase activity in the membrane fraction from a fungus. Lipids 1993, 28, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Pillai, M.G.; Certik, M.; Nakahara, T.; Kamisaka, Y. Characterization of triacylglycerol biosynthesis in subcellular fractions of an oleaginous fungus, Mortierella ramanniana var. angulispora. Biochim. Biophys. Acta 1998, 1393, 128–136. [Google Scholar] [CrossRef]
- Mysyakina, I.S.; Sergeeva, Y.E.; Bokareva, D.A. Lipid composition of the spores of zygomycetous and ascomycetous fungi during cessation of the exogenous dormancy state. Microbiology 2018, 87, 51–59. [Google Scholar] [CrossRef]
- Özşen, Ö.; Kıran, İ.; Dağ, İ.; Atlı, Ö.; Çiftçi, G.A.; Demirci, F. Biotransformation of abietic acid by fungi and biological evaluation of its metabolites. Process Biochem. 2017, 52, 130–140. [Google Scholar] [CrossRef]
- Lardizabal, K.; Effertz, R.; Levering, C.; Mai, J.; Pedroso, M.C.; Jury, T.; Aasen, E.; Gruys, K.; Bennett, K. Expression of Umbelopsis ramanniana DGAT2A in seed increases oil in soybean. Plant Physiol. 2008, 148, 89–96. [Google Scholar] [CrossRef]
- Domsch, K.H.; Gams, W.; Anderson, T.-H. Compendium of Soil Fungi; Academic Press: London, UK, 1980. [Google Scholar]
- Parshikov, I.A.; Freeman, J.P.; Lay, J.O.; Beger, R.D.; Williams, A.J.; Sutherland, J.B. Regioselective transformation of ciprofloxacin to N-acetylciprofloxacin by the fungus Mucor ramannianus. FEMS Microbiol. Lett. 1999, 177, 131–135. [Google Scholar] [CrossRef]
- Parshikov, I.A.; Freeman, J.P.; Lay, J.O.; Beger, R.D.; Williams, A.J.; Sutherland, J.B. Microbiological transformation of enrofloxacin by the fungus Mucor ramannianus. Appl. Environ. Microbiol. 2000, 66, 2664–2667. [Google Scholar] [CrossRef]
- Seo, J.; Jeon, J.; Kim, S.D.; Kang, S.; Han, J.; Hur, H.G. Fungal biodegradation of carbofuran and carbofuran phenol by the fungus Mucor ramannianus: Identification of metabolites. Water Sci. Technol. 2007, 55, 163–167. [Google Scholar] [CrossRef]
- Kang, S.-I.; Kang, S.-Y.; Hur, H.-G. Identification of fungal metabolites of anticonvulsant drug carbamazepine. Appl. Microbiol. Biotechnol. 2008, 79, 663. [Google Scholar] [CrossRef]
- Murphy, C.D. Microbial degradation of fluorinated drugs: Biochemical pathways, impacts on the environment and potential applications. Appl. Microbiol. Biotechnol. 2016, 100, 2617–2627. [Google Scholar] [CrossRef]
- Summerbell, R.C. Root endophyte and mycorrhizosphere fungi of black spruce, Picea mariana, in a boreal forest habitat: Influence of site factors on fungal distributions. Stud. Mycol. 2005, 53, 121–145. [Google Scholar] [CrossRef]
- Ikeda, H.; Fukuda, T.; Yokoyama, J. Endophytic fungi associated with a holoparasitic pant, Balanophora japonica (Balanophoraceae). Am. J. Plant Sci. 2016, 7, 152–158. [Google Scholar] [CrossRef]
- Möller, A. Untersuchungen über ein- und zweijährige Kiefern im märkischen Sandbode. Z. Forst Jagdwes. 1903, 35, 257–321. [Google Scholar]
- Sugiyama, M.; Tokumasu, S.; Gams, W. Umbelopsis gibberispora sp. nov. from Japanese leaf litter and a clarification of Micromucor ramannianus var. angulisporus. Mycoscience 2003, 44, 217–226. [Google Scholar] [CrossRef]
- Linnemann, G. Die Mucorineen-Gattung Mortierella Coemans; Gustav Fischer: Jena, Germany, 1941. [Google Scholar]
- Mil’ko, A.A. Opredeltiel’ Mukoral’nykh Gribov [Key to the identification of Mucorales]; Naukova Dumka: Kyiv, Ukraine, 1974. [Google Scholar]
- Gams, W. Some new or noteworthy species of Mortierella. Persoonia 1976, 9, 111–140. [Google Scholar]
- Von Arx, J. On Mucoraceae s. str. and other families of the Mucorales. Sydowia 1984, 35, 10–26. [Google Scholar]
- Meyer, W.; Gams, W. Delimitation of Umbelopsis (Mucorales, Umbelopsidaceae fam. nov.) based on ITS sequence and RFLP data. Mycol. Res. 2003, 107, 339–350. [Google Scholar] [CrossRef]
- Chalabuda, T. Systematics of the family Mortierellaceae. Novosti Sist. Nizsh. Rast. 1968, 1968, 120–131. [Google Scholar]
- Evans, E.H. Studies on Mortierella ramanniana: I. Relationship between morphology and cultural behaviour of certain isolates. Trans. Br. Mycol. Soc. 1971, 56, 201–216. [Google Scholar] [CrossRef]
- Wang, Y.N.; Liu, X.Y.; Zheng, R.Y. Umbelopsis changbaiensis sp. nov. from China and the typification of Mortierella vinacea. Mycol. Prog. 2014, 13, 657–669. [Google Scholar] [CrossRef]
- Turner, M. Studies in the genus Mortierella: I. Mortierella isabellina and related species. Trans. Br. Mycol. Soc. 1963, 46, 262–272. [Google Scholar] [CrossRef]
- Peberdy, J.F.; Turner, M. The esterases of Mortierella ramanniana in relation to taxonomy. J. Gen. Microbiol. 1968, 51, 303–312. [Google Scholar] [CrossRef][Green Version]
- Nagy, Á.; Pesti, M.; Galgóczy, L.; Vágvölgyi, C. Electrophoretic karyotype of two Micromucor species. J. Basic Microbiol. 2004, 44, 36–41. [Google Scholar] [CrossRef]
- Ogawa, Y.; Suda, A.; Kusama-Eguchi, K.; Watanabe, K.; Tokumasu, S. Intraspecific groups of Umbelopsis ramanniana inferred from nucleotide sequences of nuclear rDNA internal transcribed spacer regions and sporangiospore morphology. Mycoscience 2005, 46, 343–351. [Google Scholar] [CrossRef]
- Ogawa, Y.; Sugiyama, M.; Hirose, D.; Kusama-Eguchi, K.; Tokumasu, S. Polyphyly of intraspecific groups of Umbelopsis ramanniana and their genetic and morphological variation. Mycoscience 2011, 52, 91–98. [Google Scholar] [CrossRef]
- Chen, F.-J. The genus Mortierella Coemans in China; Graduate University of Chinese Academy of Sciences: Beijing, China, 1986. [Google Scholar]
- Zheng, R.-Y.; Chen, G.-Q.; Huang, H.; Liu, X.-Y. A monograph of Rhizopus. Sydowia 2007, 59, 273–372. [Google Scholar]
- Ridgway, R. Color Standards and Color Nomenclature; Published by the Author: Washington, DC, USA, 1912. [Google Scholar]
- Wang, Y.-N.; Liu, X.-Y.; Zheng, R.-Y. Umbelopsis longicollis comb. nov. and the synonymy of U. roseonana and U. versiformis with U. nana. Mycologia 2015, 107, 1023–1032. [Google Scholar] [CrossRef]
- Schmitt, I.; Crespo, A.; Divakar, P.K.; Fankhauser, J.D.; Herman-Sackett, E.; Kalb, K.; Nelsen, M.P.; Nelson, N.A.; Rivas-Plata, E.; Shimp, A.D.; et al. New primers for promising single-copy genes in fungal phylogenetics and systematics. Persoonia 2009, 23, 35–40. [Google Scholar] [CrossRef]
- Tretter, E.D.; Johnson, E.M.; Wang, Y.; Kandel, P.; White, M.M. Examining new phylogenetic markers to uncover the evolutionary history of early-diverging fungi: Comparing MCM7, TSR1 and rRNA genes for single- and multi-gene analyses of the Kickxellomycotina. Persoonia 2013, 30, 106–125. [Google Scholar] [CrossRef] [PubMed]
- Molitor, C.; Inthavong, B.; Sage, L.; Geremia, R.A.; Mouhamadou, B. Potentiality of the cox1 gene in the taxonomic resolution of soil fungi. FEMS Microbiol. Lett. 2010, 302, 76–84. [Google Scholar] [CrossRef]
- Katoh, K.; Kuma, K.-I.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Page, R.D.M. TreeView: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Alfaro, M.E.; Zoller, S.; Lutzoni, F. Bayes or bootstrap? A simulation study comparing the performance of Bayesian Markov chain Monte Carlo sampling and bootstrapping in assessing phylogenetic confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef]
- Mahoney, D.; Gams, W.; Meyer, W.; Starink-Willemse, M. Umbelopsis dimorpha sp. nov., a link between U. vinacea and U. versiformis. Mycol. Res. 2004, 108, 107–111. [Google Scholar] [CrossRef]
- Lendner, A. Les Mucorinées de La Suisse. In Materiaux pour la Flore Cryptogamique Suisse; K.-J. Wyss: Bern, Switzerland, 1908; Volume III. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.S.J.; Smith, D.; Summerell, B.A.; Cano-Lira, J.F.; Guarro, J.; Houbraken, J. Fungal Planet description sheets: 625–715. Persoonia 2017, 39, 270. [Google Scholar] [CrossRef]
- Yuan, H.-S.; Lu, X.; Dai, Y.-C.; Hyde, K.D.; Kan, Y.-H.; Kušan, I.; He, S.-H.; Liu, N.-G.; Sarma, V.V.; Zhao, C.-L. Fungal diversity notes 1277–1386: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2020, 104, 1–266. [Google Scholar] [CrossRef]
- Buyck, B.; Kauff, F.; Eyssartier, G.; Couloux, A.; Hofstetter, V. A multilocus phylogeny for worldwide Cantharellus (Cantharellales, Agaricomycetidae). Fungal Divers. 2014, 64, 101–121. [Google Scholar] [CrossRef]
- Xu, J. Fungal DNA barcoding. Genome 2016, 59, 913–932. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Colony Diam. (mm) | Sporangial Diam. (µm) | Sporangial Walls DLQ 1 | Collars | Columellae (µm) | Sporangiospores (µm) | Macro-Chlamydospores |
|---|---|---|---|---|---|---|---|
| C1: U. curvata | 43–48 | (11.0–) 15.5–22.5 (–24.5) | rapidly | small but conspicuous | depressed globose (4.0–) 6.0–8.5 (–10) × (3.5–) 5.0–7.0 (–8.5); or subglobose 4.0–6.0 (–7.0) | ellipsoidal and often curving to one side (2.7–) 3.2–4.5 (–5.7) × (1.6–) 2.0–2.4 (–2.8) | abundant on CMA and MEA |
| C2: U. ramanniana | 40–44 | (11.0–) 15.0–23.0 (–29.5) | rapidly | small but conspicuous | ovoid to oblong-ovoid 4.0–6.0 (–8.0) × (4.5–) 5.0–7.0 (–9.5) µm | ellipsoidal 2.4–3.6 (–4.6) × 1.6–2.4 (–2.6) | less abundant on CMA or MEA |
| C3: U. microsporangia | 35–40 | (10.0–) 14.0–18.5 (–22.5) | rapidly | small or no | depressed globose (3.0–) 4.0–6.5 (–10.0) × (2.5–) 3.5–5.5 (–9.0); or subglobose (2.5–) 4.0–6.0 | ovoid to subglobose (2.4–) 2.6–3.2 (–4.0) × 1.8–2.5 (–2.8) | rare on CMA and less abundant on MEA |
| C4: U. dura | 41–45 | (10.0–) 15.0–23.5 (–31.5) | slowly | small or no | roundish conical (2.5–) 3.5–6.0 (–7.0) × (3.5–) 4.5–7.0 (–8.0) | ovoid and sometimes slightly narrowing on one end (2.4–) 2.8–3.3 (–4.6) × (1.6–) 1.9–2.4 (–2.8) | undiscovered on CMA and less abundant on MEA |
| C5: U. oblongielliptica | 60–62 | (11.5–) 15.5–21.5 (–26.0) | rapidly | small but conspicuous | roundish conical 3.5–6.0 × 4–6.5 | oblong-ellipsoidal and sometimes slightly curved to one side (3.2–) 4.0–5.0 (–5.5) × 1.6–2.0 (–2.5) | abundant on CMA and MEA |
| C6: U. macrospora | 46 | (13.5–) 16.0–23.0 (–25.5) | rapidly | small but conspicuous | depressed globose (4.0–) 6.0–8.0 (–10) × (3.5–) 4.0–6.0 (–7.0); or subglobose (3.0–) 5.0–7.0 (–8.0) | oblong-ellipsoidal to ellipsoidal and sometimes slightly curving to one side (3.6–) 4.0–4.9 (–5.5) × (1.7–) 2.4–2.8 (–3.2) | undiscovered on CMA or MEA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-N.; Liu, X.-Y.; Zheng, R.-Y. The Umbelopsis ramanniana Sensu Lato Consists of Five Cryptic Species. J. Fungi 2022, 8, 895. https://doi.org/10.3390/jof8090895
Wang Y-N, Liu X-Y, Zheng R-Y. The Umbelopsis ramanniana Sensu Lato Consists of Five Cryptic Species. Journal of Fungi. 2022; 8(9):895. https://doi.org/10.3390/jof8090895
Chicago/Turabian StyleWang, Ya-Ning, Xiao-Yong Liu, and Ru-Yong Zheng. 2022. "The Umbelopsis ramanniana Sensu Lato Consists of Five Cryptic Species" Journal of Fungi 8, no. 9: 895. https://doi.org/10.3390/jof8090895
APA StyleWang, Y.-N., Liu, X.-Y., & Zheng, R.-Y. (2022). The Umbelopsis ramanniana Sensu Lato Consists of Five Cryptic Species. Journal of Fungi, 8(9), 895. https://doi.org/10.3390/jof8090895