
Endophytic Diaporthe Associated with Morinda officinalis in China

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of Endophytic Fungi
2.2. DNA Extraction and PCR Amplification
2.3. Phylogenetic Analysis
2.4. Morphological Characterisation
2.5. Pairwise Homoplasy Index (PHI)
3. Results
3.1. Isolation
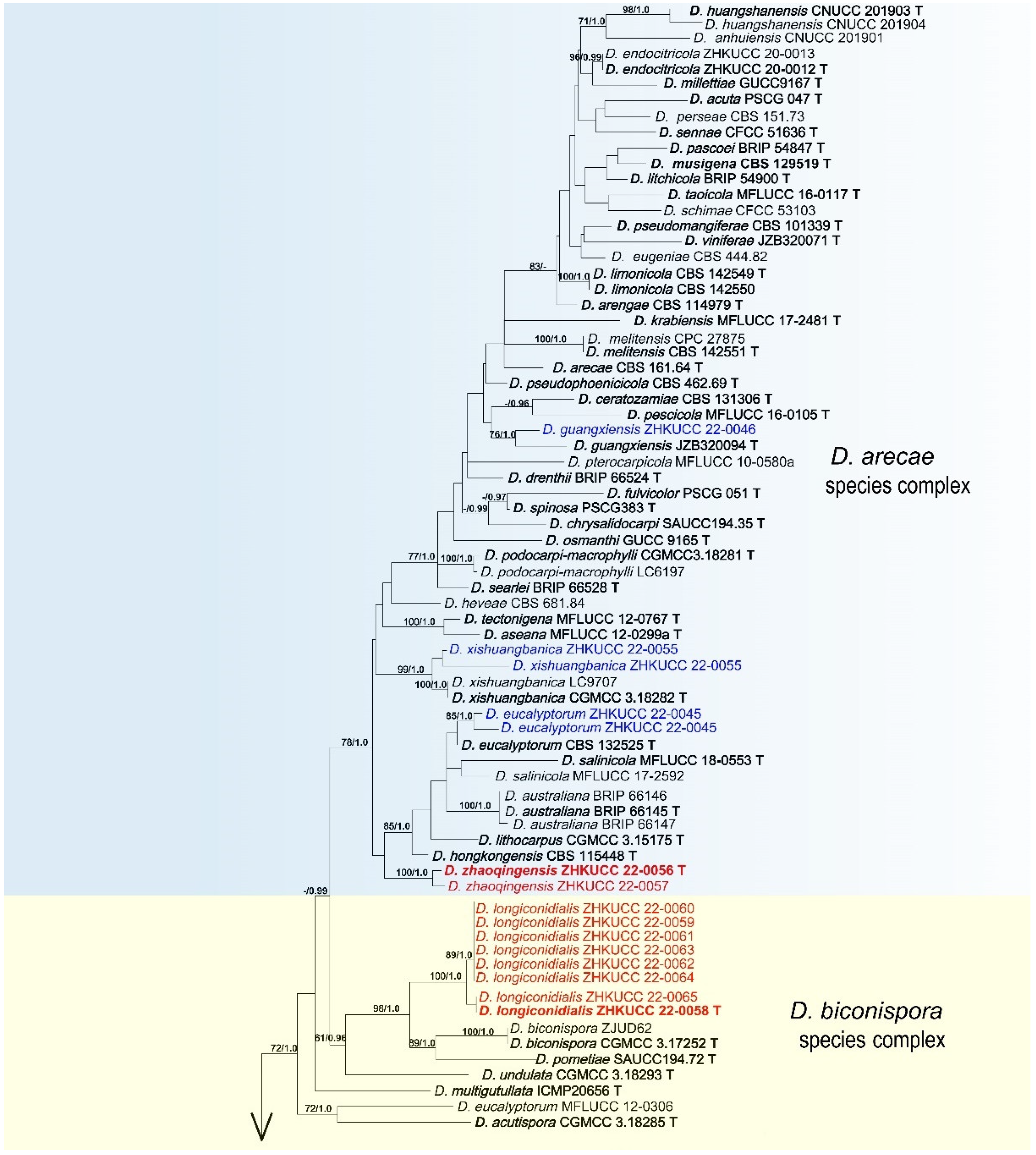
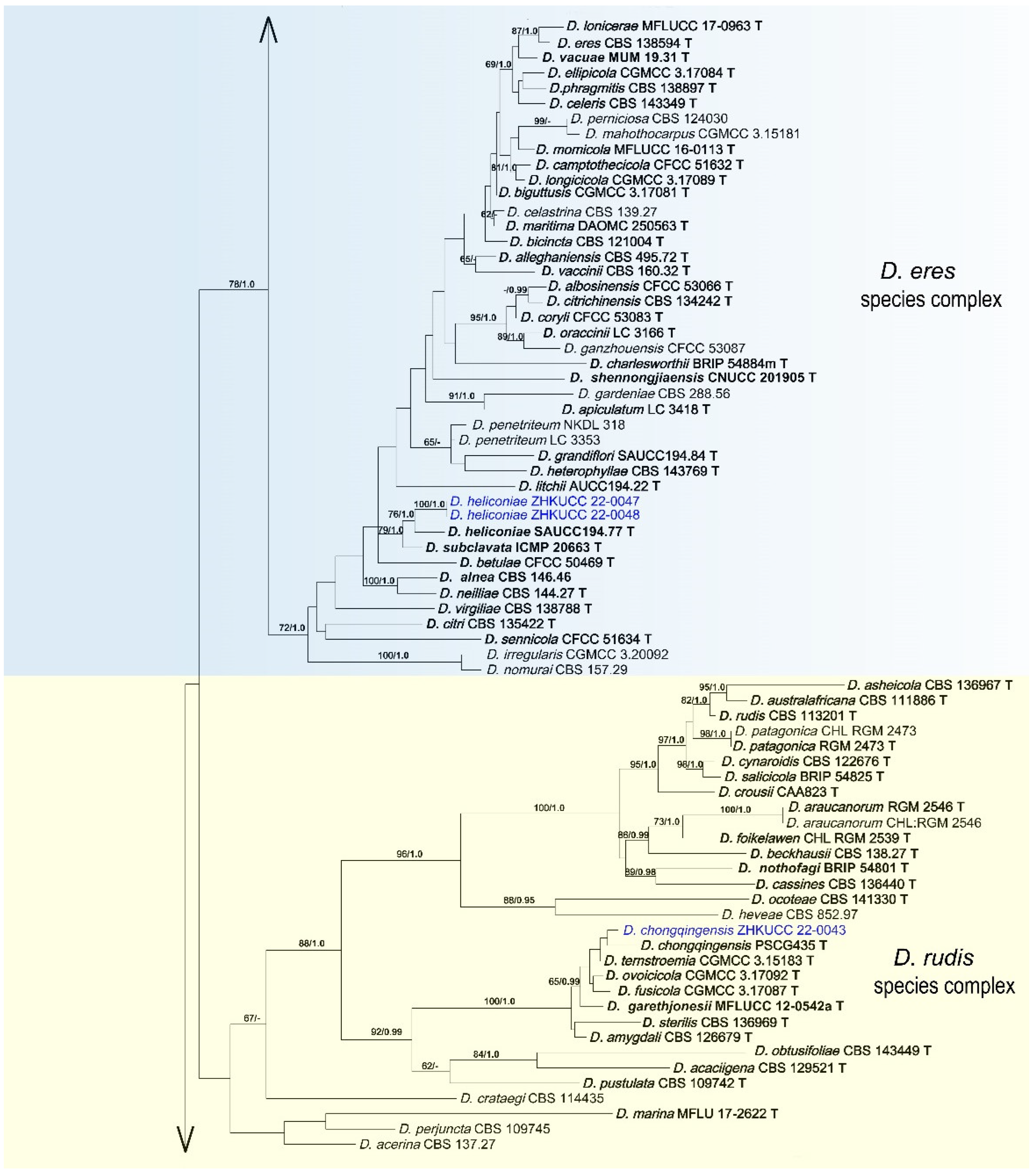
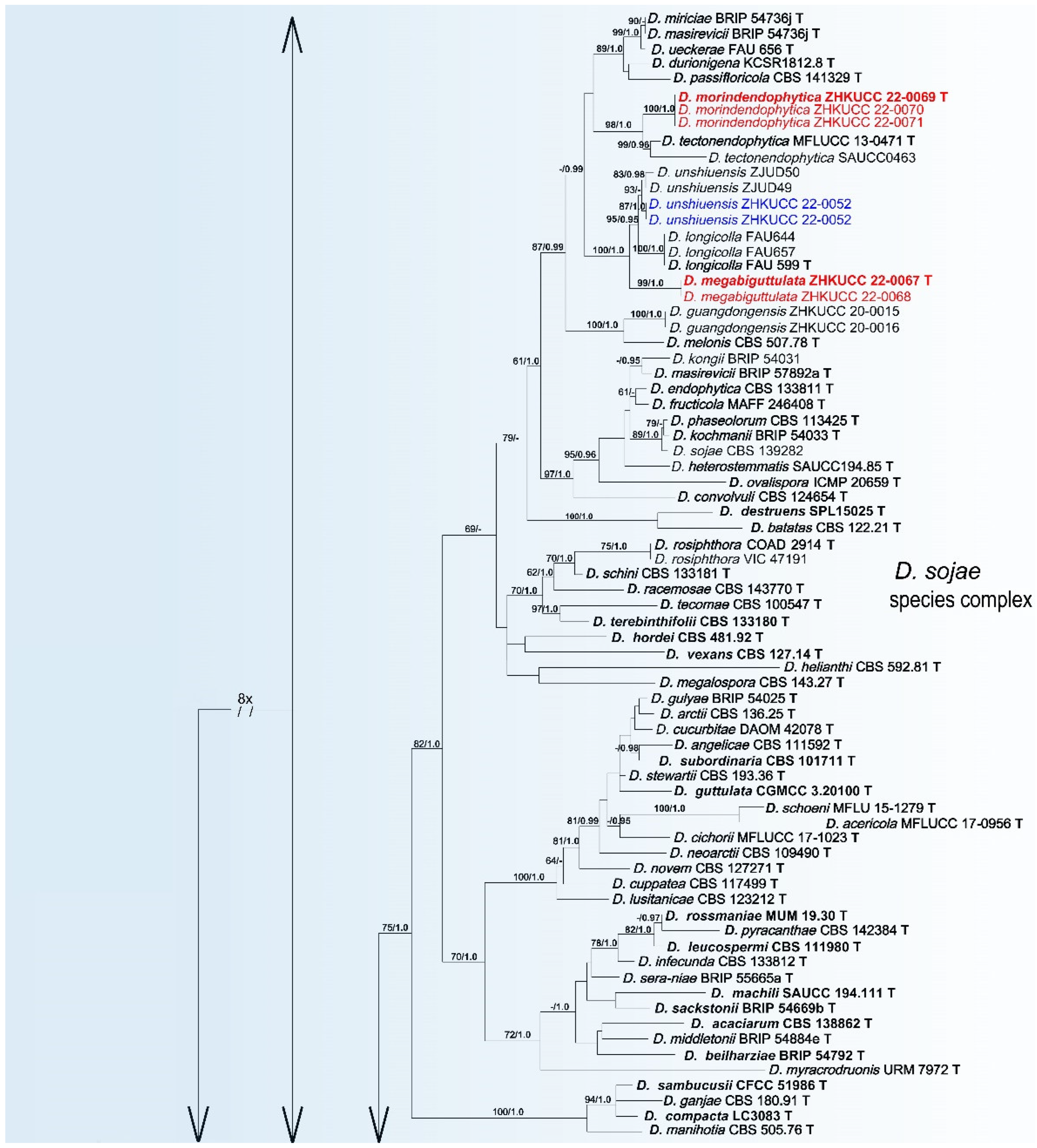
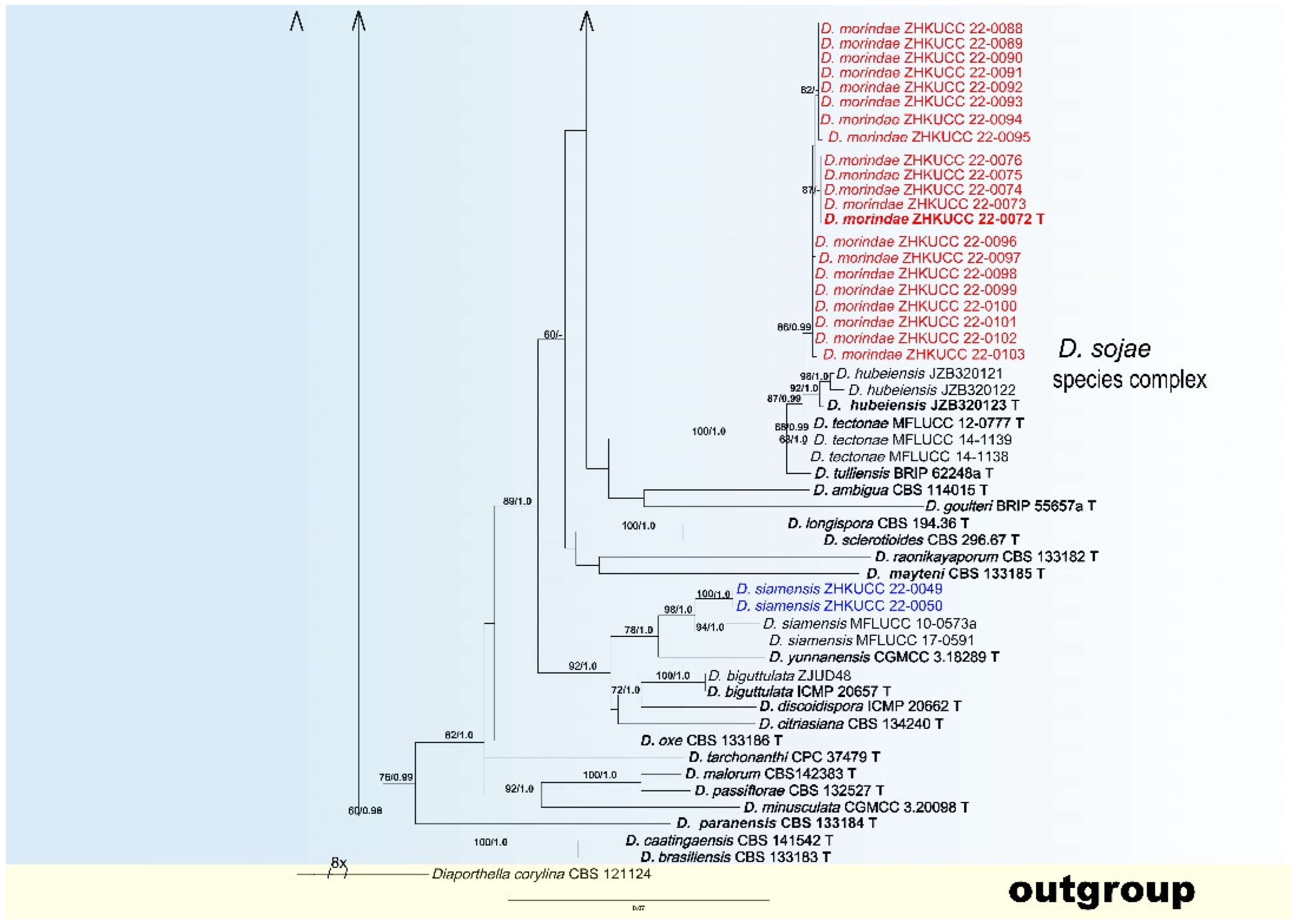
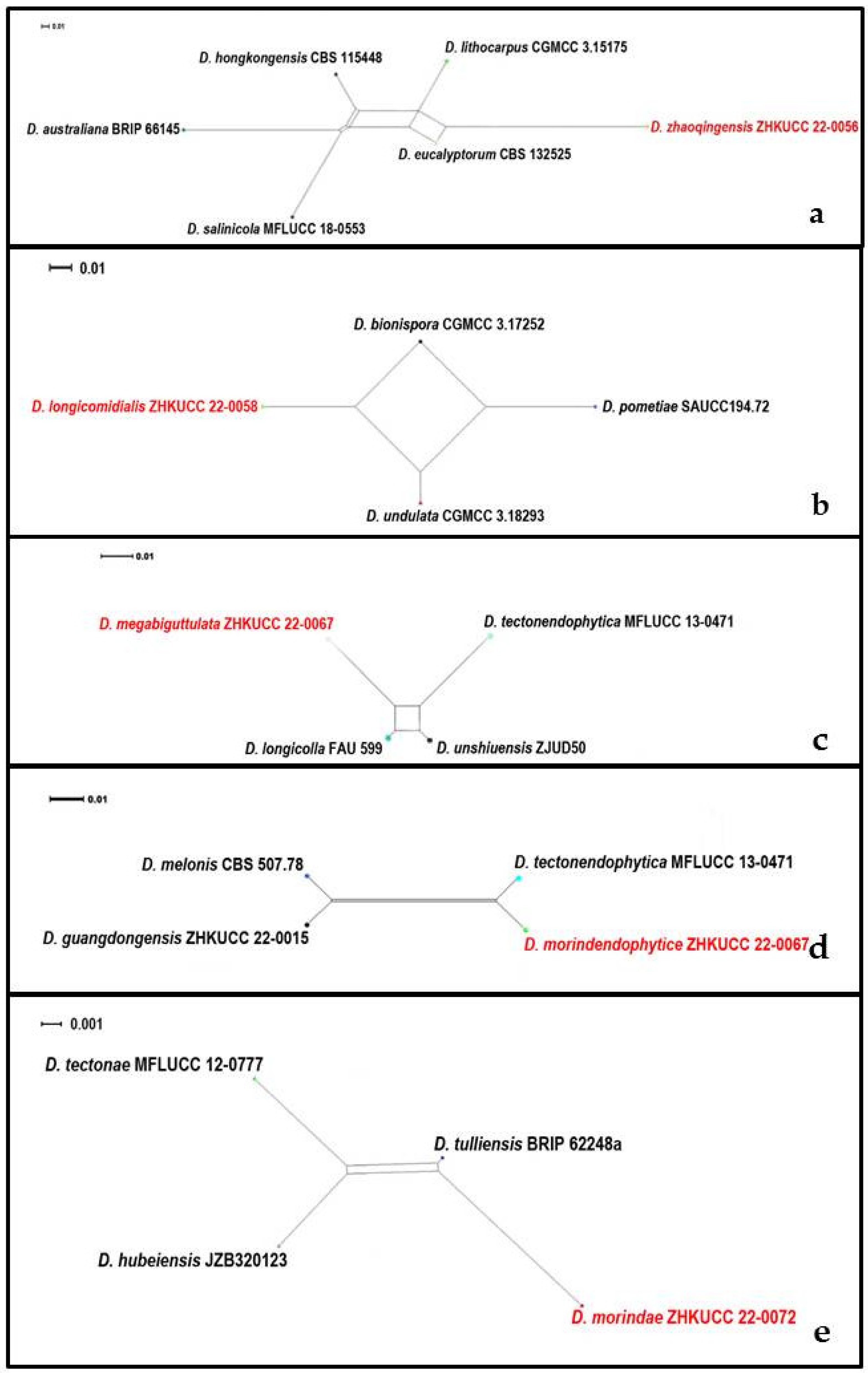
3.2. Phylogenetic Analyses
3.3. PHI Analysis
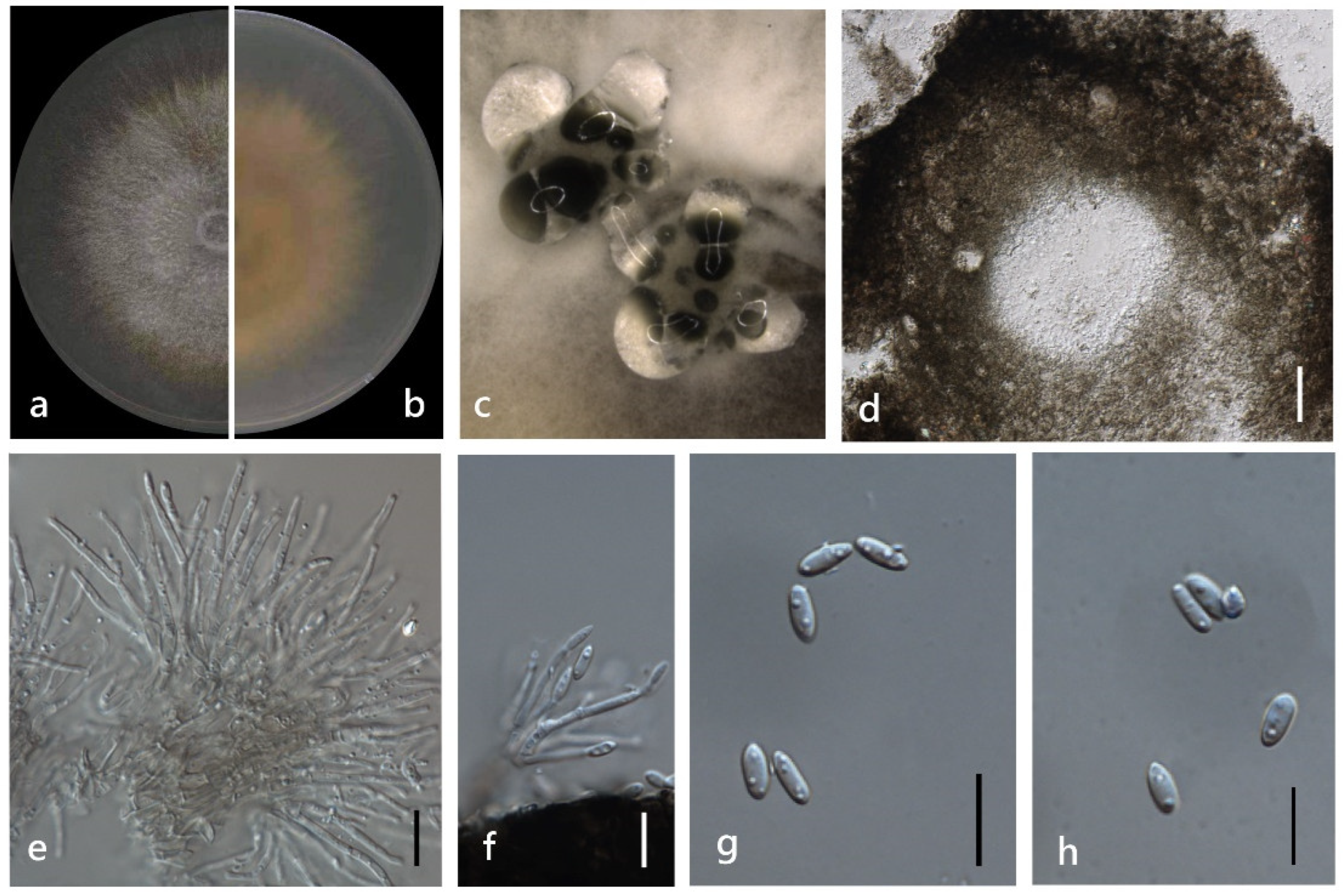
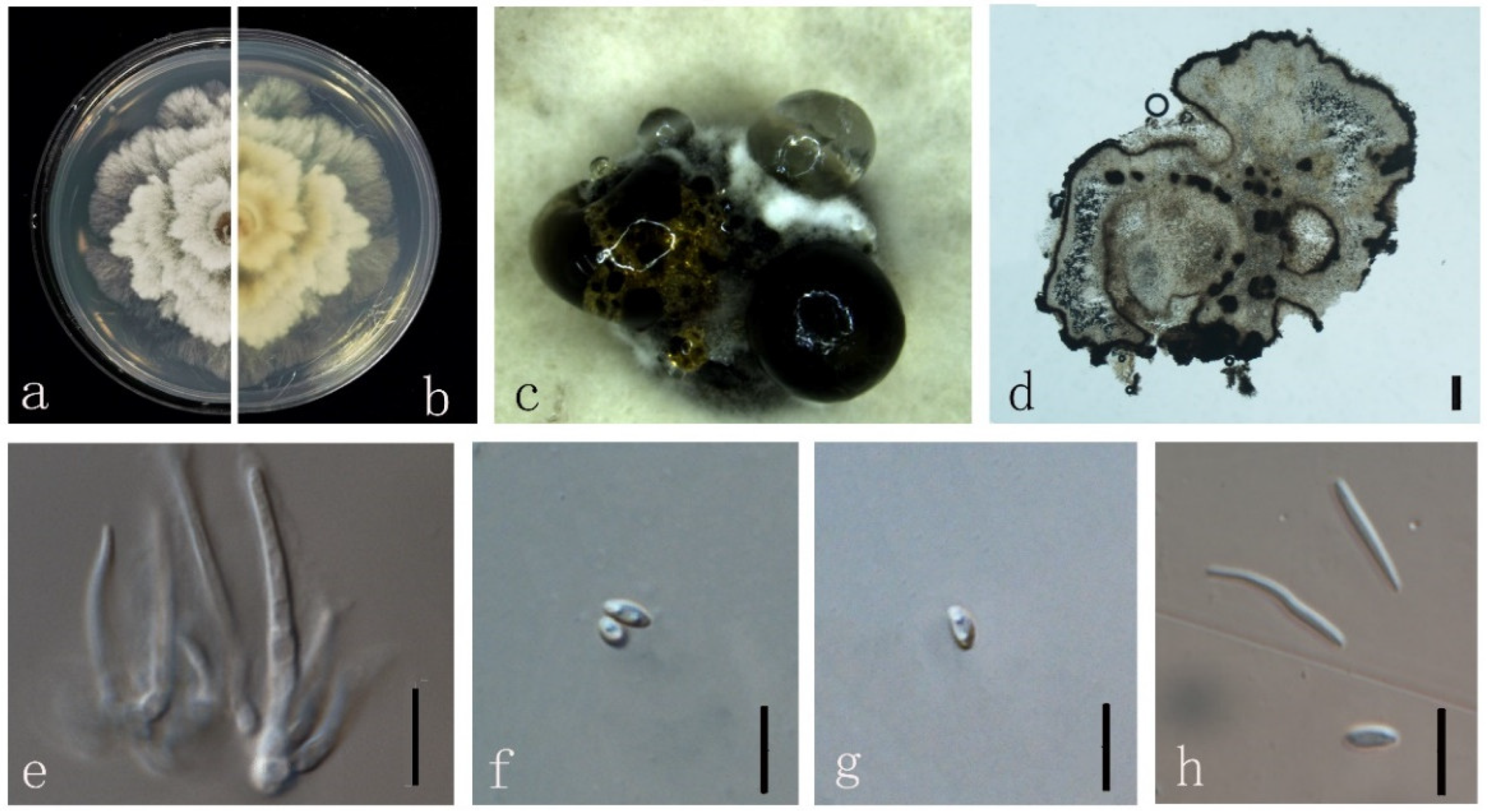
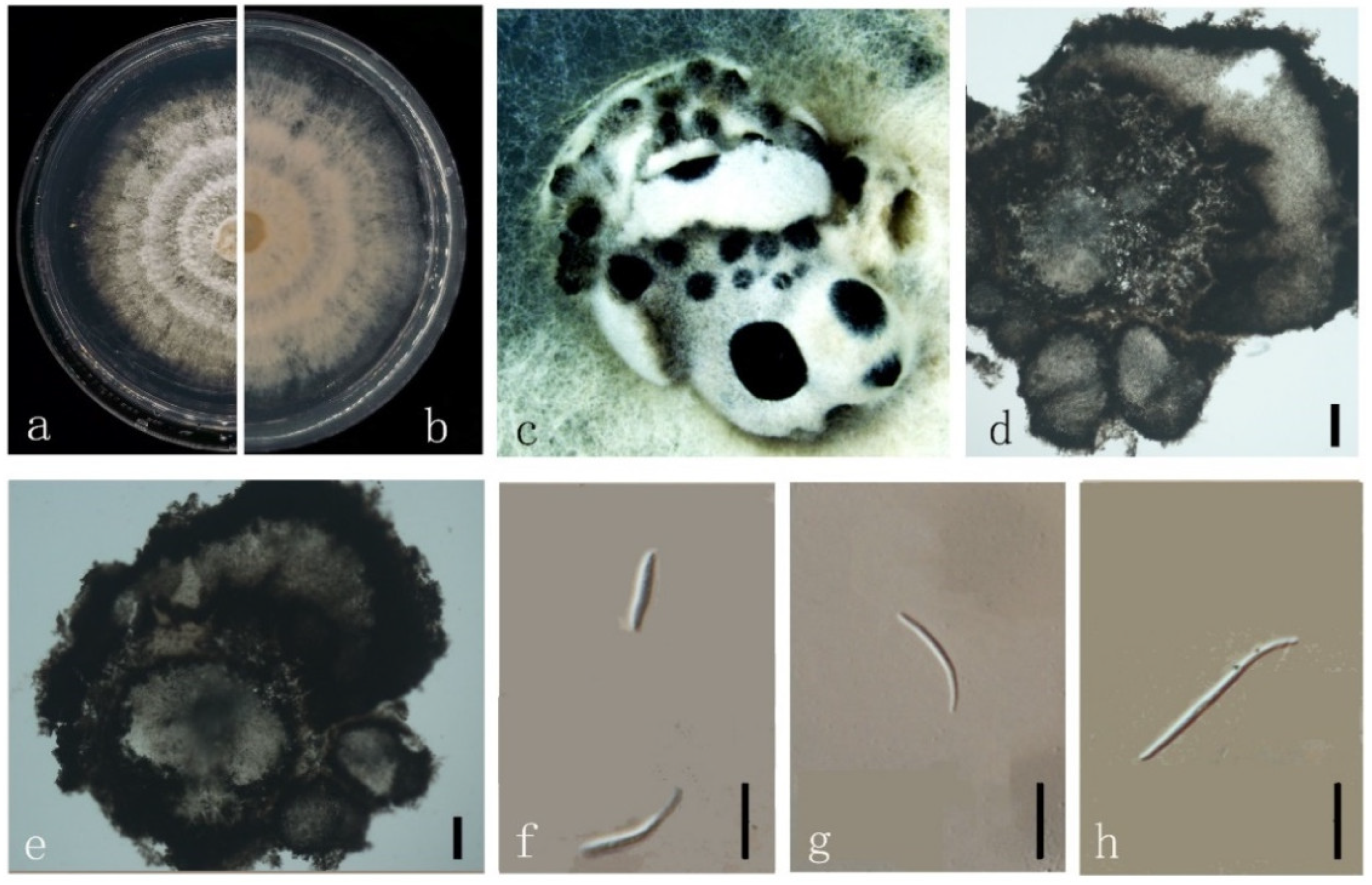
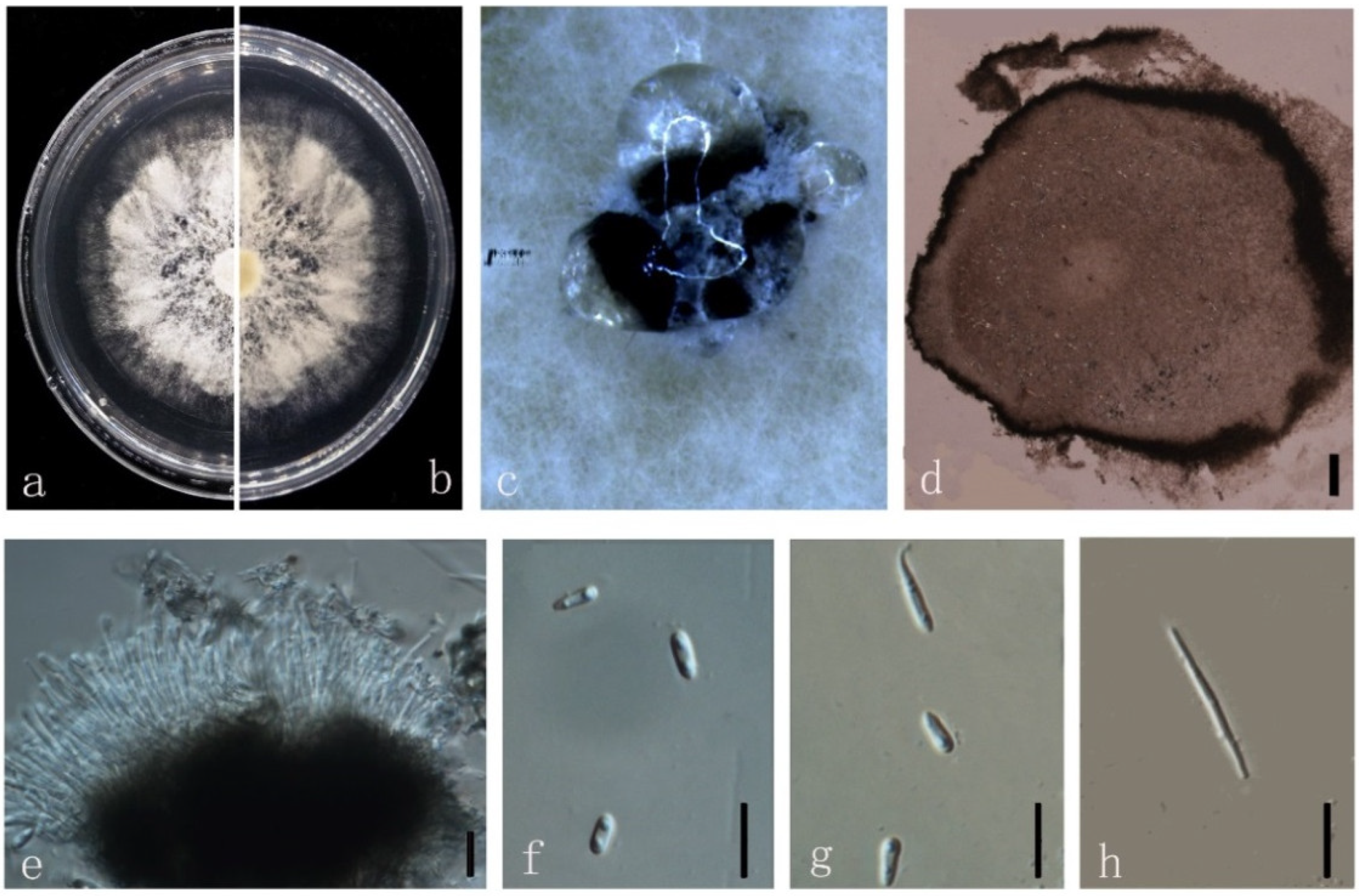
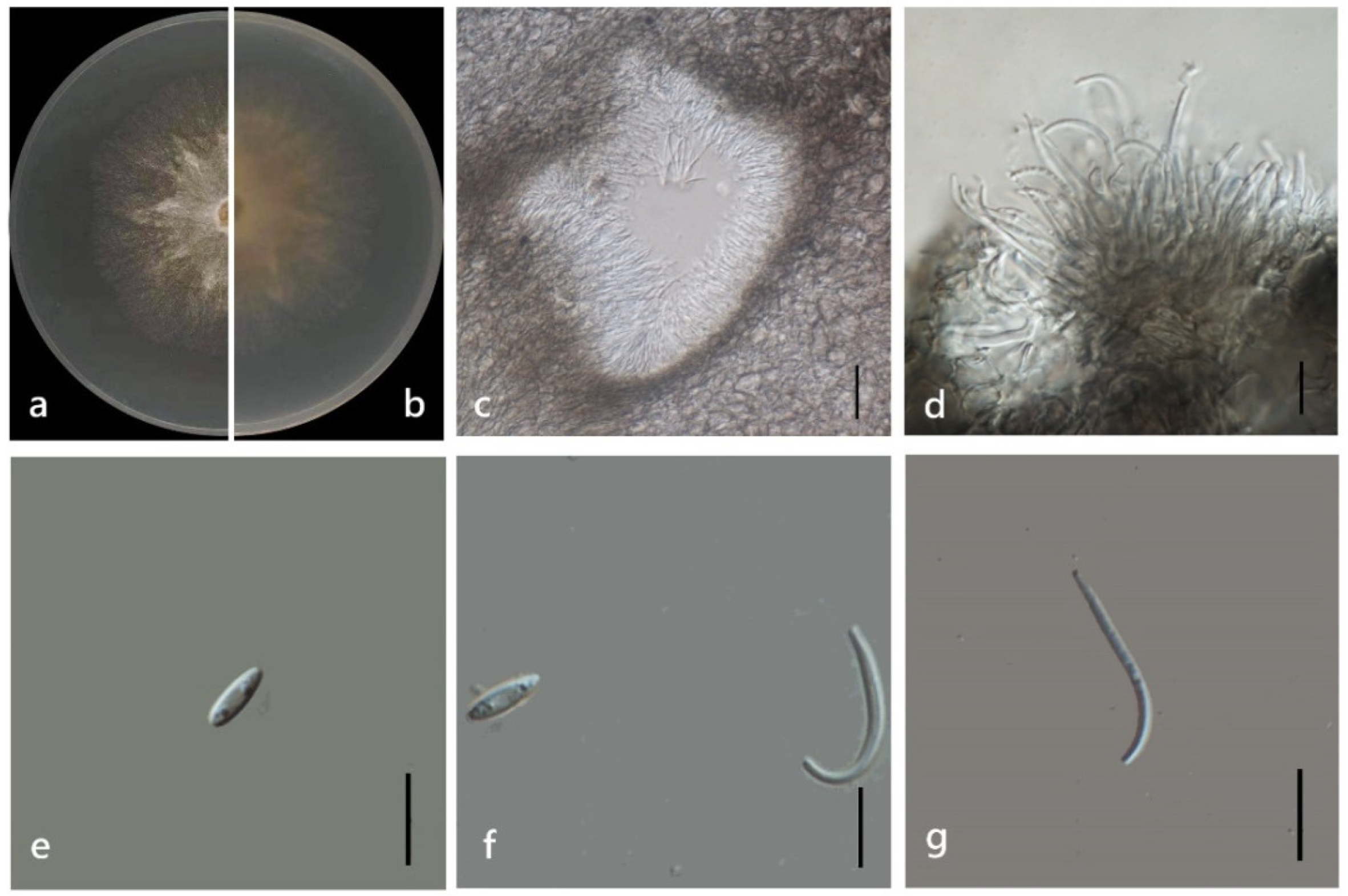
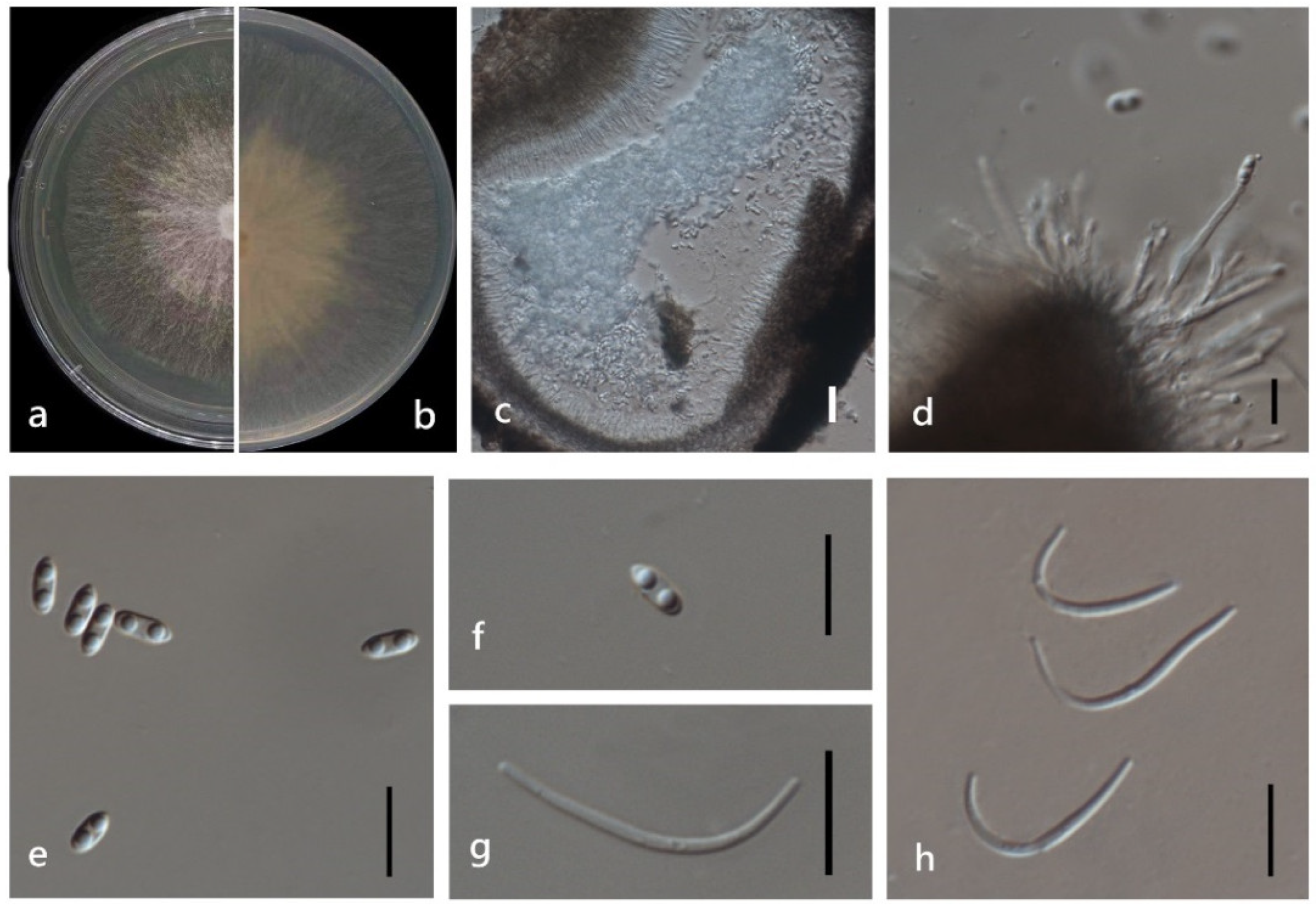
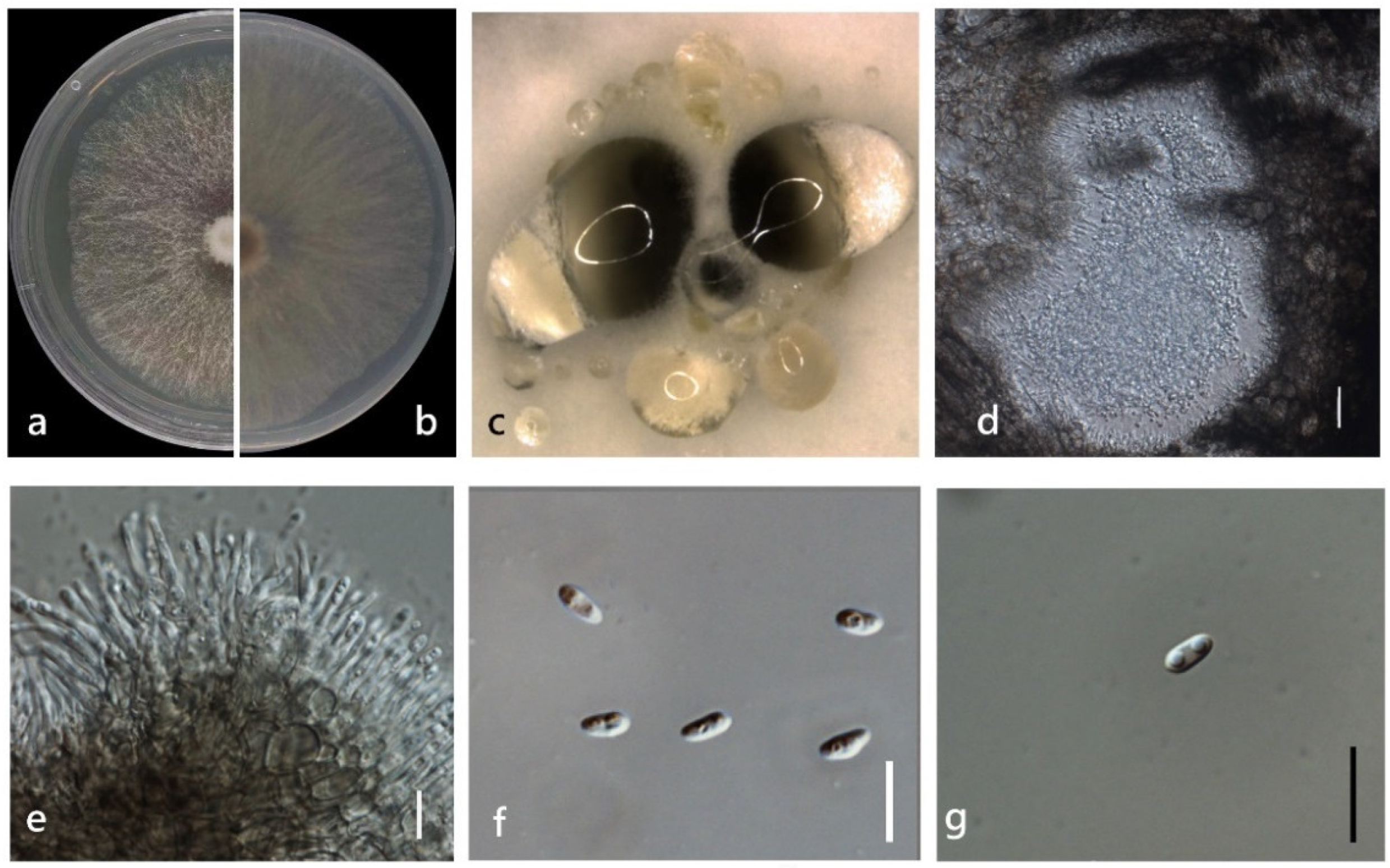
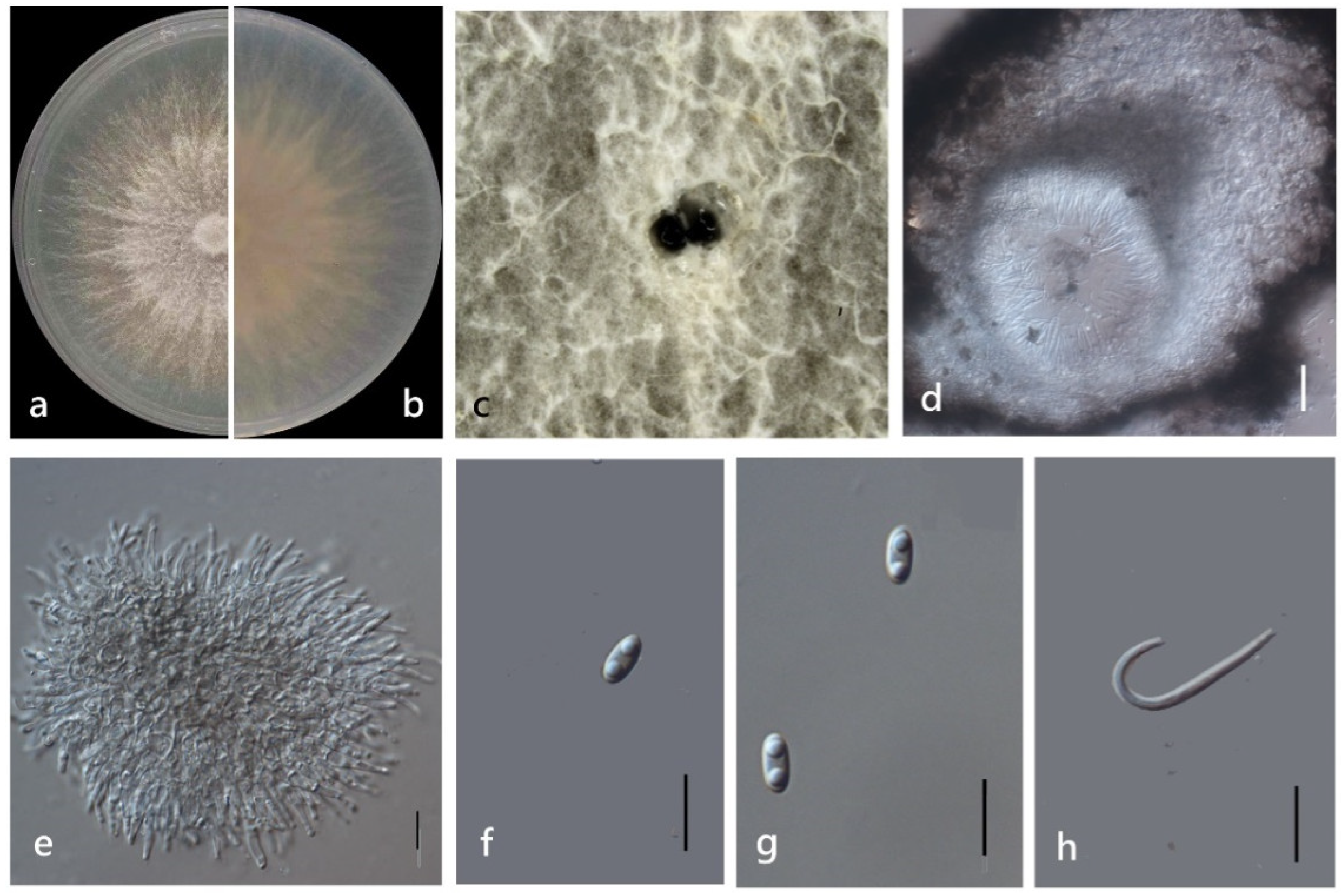
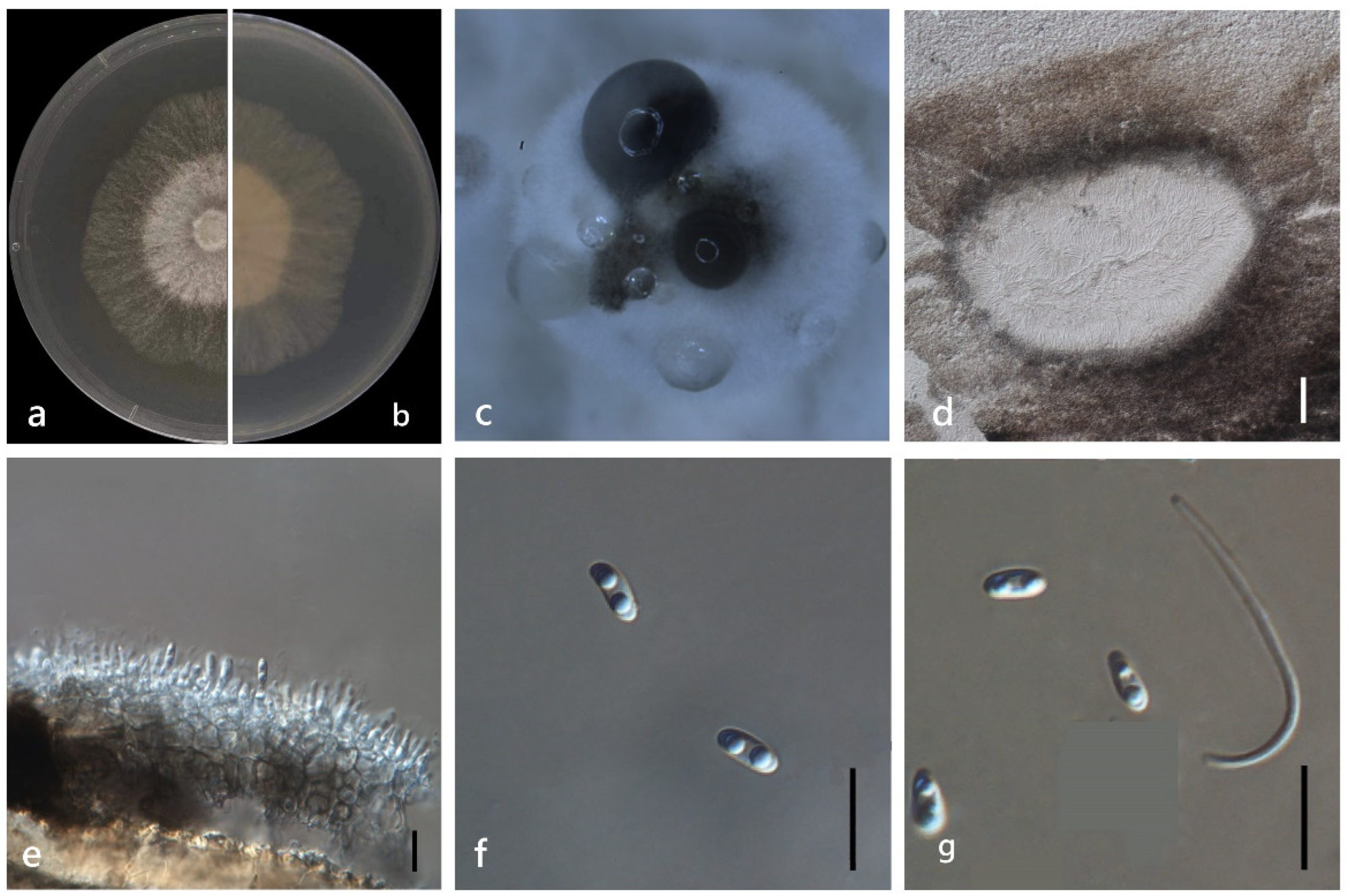
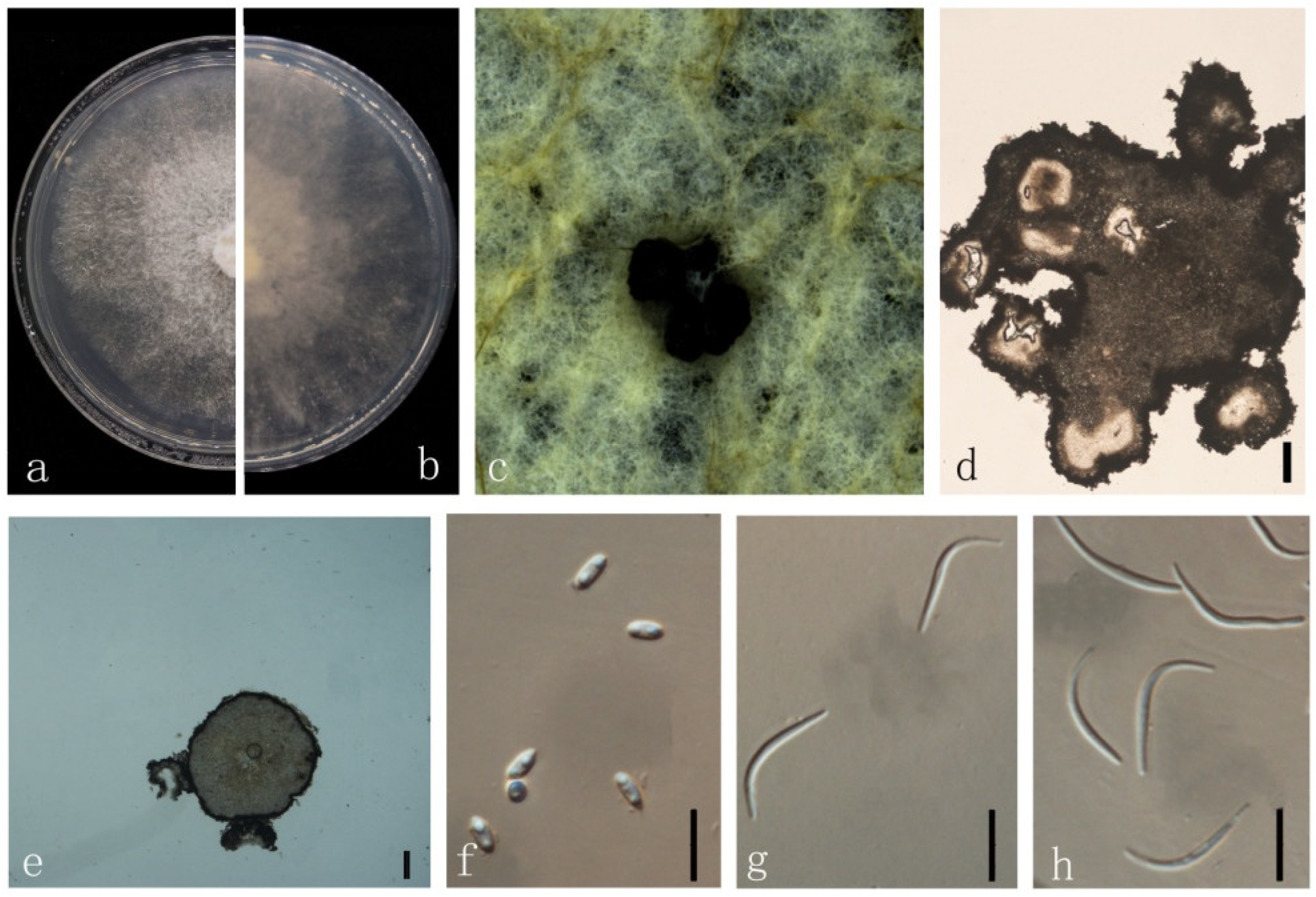
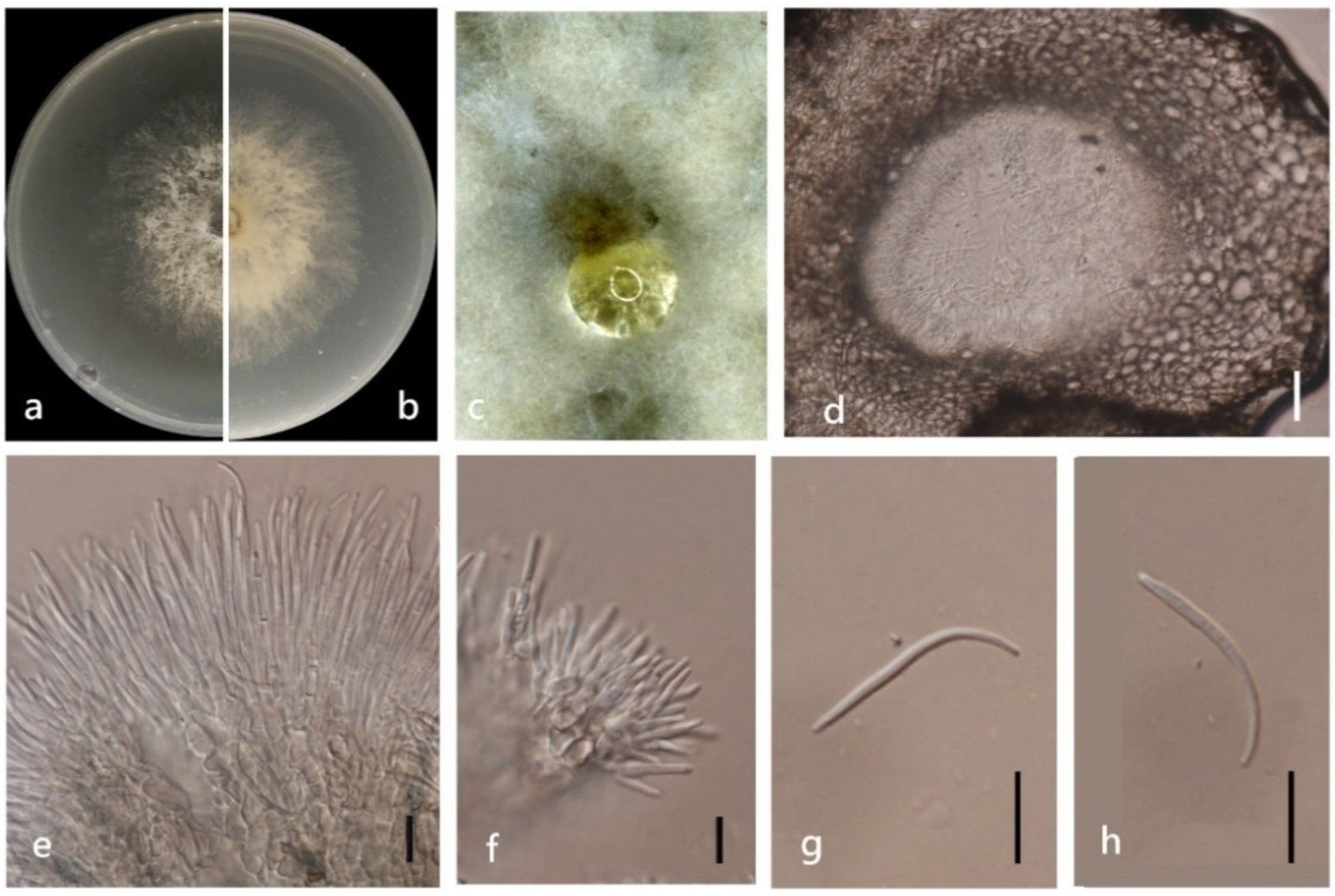
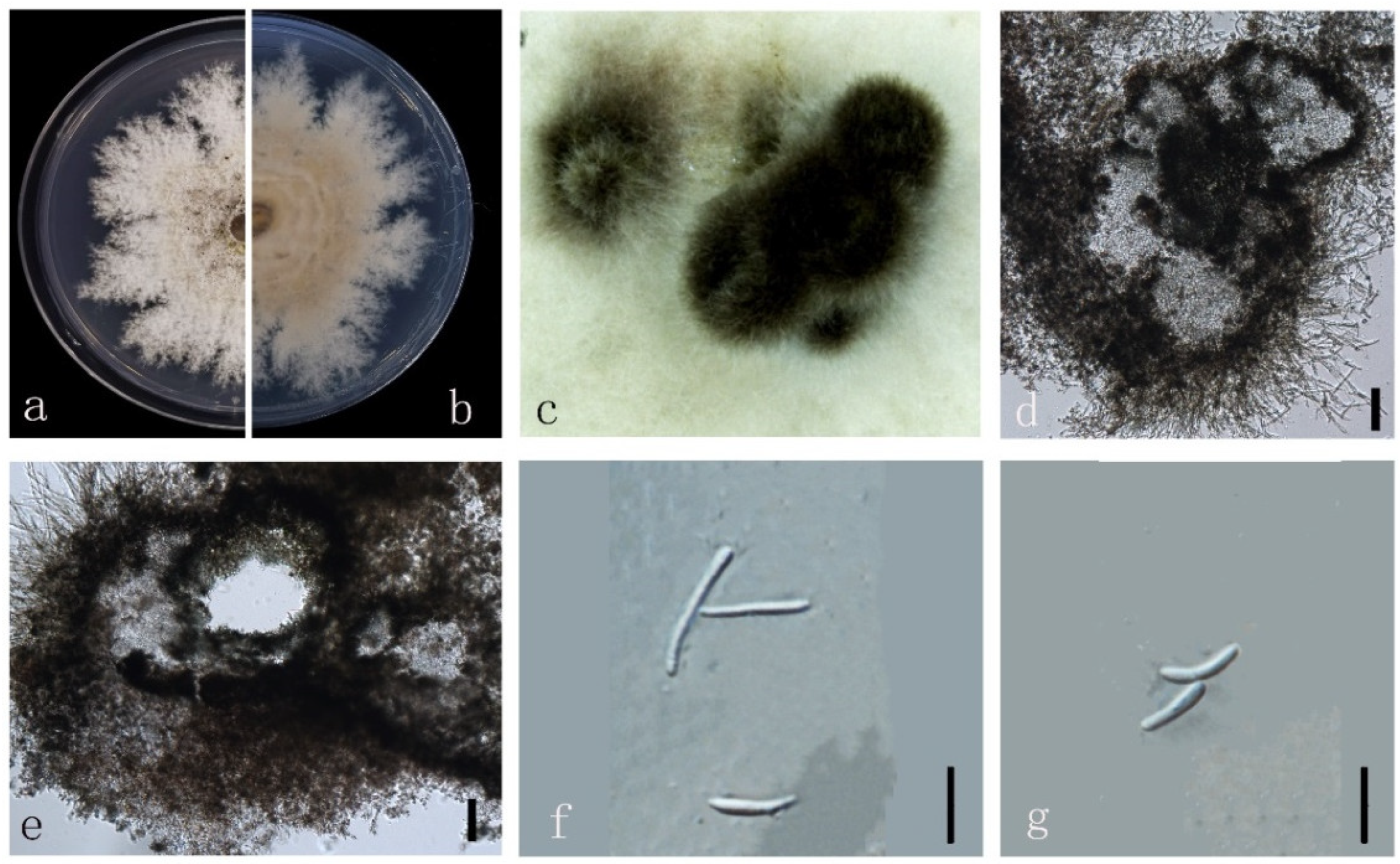
3.4. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Senanayake, I.C.; Crous, P.W.; Groenewald, J.Z.; Maharachchikumbura, S.S.; Jeewon, R.; Phillips, A.J.L.; Bhat, J.D.; Perera, R.H.; Li, Q.R.; Li, W.J.; et al. Families of Diaporthales based on morphological and phylogenetic evidence. Stud. Mycol. 2017, 86, 217–296. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Udayanga, D.; Liua, X.; Mckenzie, E.H.C.; Chukeatirote, E.; Hyde, K.D. Multi-locus Phylogeny Reveals Three new Species of Diaporthe from Thailand. Cryptogam. Mycol. 2012, 33, 295–309. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Hyde, K.D. Species limits in Diaporthe: Molecular reassessment of D. citri, D. cytosporella, D. foeniculina and D. rudis. Persoonia 2014, 32, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Guarnaccia, V.; Martino, I.; Tabone, G.; Brondino, L.; Gullino, M.L. Fungal pathogens associated with stem blight and dieback of blueberry in northern Italy. Phytopathol. Mediterr. 2020, 59, 229–245. [Google Scholar]
- Ariyawansa, H.A.; Tsai, I.; Wang, J.Y.; Withee, P.; Tanjira, M.; Lin, S.R.; Suwannarach, N.; Kumla, J.; Elgorban, A.M.; Cheewangkoon, R. Molecular Phylogenetic Diversity and Biological Characterization of Diaporthe Species Associated with Leaf Spots of Camellia sinensis in Taiwan. Plants 2021, 10, 1434. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by species in Diaporthe. IMA Fungus 2017, 8, 317–334. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, F.; Duan, W.; Crous, P.W.; Cai, L. Diaporthe is paraphyletic. IMA Fungus 2017, 8, 153–187. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Crous, P.W. Species of Diaporthe on Camellia and Citrus in the Azores Islands. Phytopathol. Medit. 2018, 57, 307–319. [Google Scholar]
- Guo, Y.H.; Crous, P.W.; Bai, Q.; Fu, M.; Yang, M.M.; Wang, X.H.; Du, Y.M.; Hong, N.; Xu, W.X.; Wang, G.P. High diversity of Diaporthe species associated with pear shoot canker in China. Persoonia 2020, 45, 132–162. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, X.L.; Du, Z.; Tian, C.M. Diaporthe species occurring on Senna bicapsularis in southern China, with descriptions of two species. Phytotaxa 2017, 302, 145–155. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Dissanayake, A.J.; Li, X.; Liu, M.; Wanasinghe, D.N.; Xu, J.; Zhao, W.; Zhang, W.; Zhou, Y.; Hyde, K.D.; et al. High genetic diversity and species complexity of Diaporthe associated with grapevine dieback in China. Front. Microbiol. 2019, 10, 1936. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Udayanga, D.; Wang, X.H.; Hou, X.; Mei, X.F.; Fu, Y.S.; Hyde, K.D.; Li, H. Endophytic Diaporthe associated with Citrus: A phylogenetic reassessment with seven new species from China. Fungal Biol. 2015, 119, 331–347. [Google Scholar] [CrossRef]
- Dong, Z.; Manawasinghe, I.S.; Huang, Y.; Shu, Y.; Phillips, A.J.L.; Dissanayake, A.J.; Hyde, K.D.; Xiang, M.M.; Luo, M. Endophytic Diaporthe Associated With Citrus grandis cv. Tomentosa in China. Front. Microbiol. 2021, 11, 609387. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z. Hosts, species and genotypes: Opinions versus data—Presented as a keynote address at the 15th biennial conference of the Australasian plant pathology society, Geelong, Australia, 26–29 September 2005. Australas. Plant Pathol. 2005, 34, 463–470. [Google Scholar] [CrossRef]
- Promputtha, I.; Jeewon, R.; Lumyong, S.; Mckenzie, E.; Hyde, K. Ribosomal DNA fingerprinting in the identification of non sporulating endophytes from Magnolia liliifera (Magnoliaceae). Fungal Divers. 2005, 20, 167–186. [Google Scholar]
- Huang, W.; Cai, Y.; Hyde, K.; Corke, H.; Sun, M. Biodiversity of endophytic fungi associated with 29 Traditional Chinese Medicinal plants. Fungal Divers. 2008, 33, 61–75. [Google Scholar]
- Zhang, J.H.; Xin, H.L.; Xu, Y.M.; Shen, Y.; He, Y.Q.; Lin, B.; Song, H.T.; Yang, H.Y.; Qin, L.P.; Zhang, Q.Y.; et al. Morinda officinalis How.—A comprehensive review of traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2018, 213, 230–255. [Google Scholar] [CrossRef]
- Dong, Z.; Shu, Y.; Luo, M.; Zhang, W.; Xiang, M.M. First report of black root rot disease on Morinda officinalis caused by Lasiodiplodia pseudotheobromae in China. Plant Dis. 2019, 103, 2693. [Google Scholar] [CrossRef]
- Nguyen, D.T.; Hieu, N.C.; Hung, N.V.; Hoat, T.X. Biological control of Fusarium root rot of indian mulberry (Morinda officinalis how.) with consortia of agriculturally important microorganisms in vietnam. Chem. Biol. Technol. Agric. 2019, 6, 27. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Yang, J.; Huang, J.H.; Xiang, M.M. Study on biodiversity of endophytic fungi of Amomum villosum. J. Zhongkai Univ. Agric. Eng. 2010, 23, 15–17. (In Chinese) [Google Scholar]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; MInnis, A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z.; Risède, J.; Simoneau, P.; Hywel-Jones, N.L. Calonectria species and their cylindrocladium anamorphs species with sphaeropedunculate vesicles. Stud. Mycol. 2004, 50, 415–430. [Google Scholar]
- Hall, T. Bioedit. Department of Microbiology, North Carolina State University. 2006. Available online: http://www.mbio.ncsu.edu/BioEdit/page2.html (accessed on 1 July 2021).
- Yang, Q.; Jiang, N.; Tian, C.M. New species and records of Diaporthe from Jiangxi Province, China. MycoKeys 2021, 77, 41–64. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2010, 9, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, D.; Michalak, I. RaxmlGUI: A Graphical Front-End for RAxML. 2010. Available online: http://sourceforgenet/projects/raxmlgui/ (accessed on 25 February 2021).
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest 2.0. Program Distributed by the Author; Evolutionary Biology Centre Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; Institute of Electrical and Electronics Engineers: New Orleans, LA, USA, 2010. [Google Scholar]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The faces of fungi database: Fungal names linked with morphology, molecular and human attributes. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; de Farias, A.R.G.; Bhunjun, C.S.; Ferdinandez, H.S.; Manamgoda, D.S.; Udayanga, D.; Herath, I.S.; Thambugala, K.M.; Manawasinghe, I.S.; et al. What is a species in fungal plant pathogens? Fungal Divers. 2021, 109, 239–266. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.L.; Xu, J.; Balasuriya, A.; Hyde, K.D.; Tępień, L.; Harischandra, D.L.; Karunarathna, A.; Yan, J.; Weerasinghe, J.; et al. Defining a species in plant pathology: Beyond the species level. Fungal Divers. 2021, 109, 267–282. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute: Kew, UK, 1970. [Google Scholar]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Norphanphoun, C.; Gentekaki, E.; Hongsanan, S.; Jayawardena, R.; Senanayake, C.; Manawasinghe, I.; Abeywickrama, P.; Bhunjun, C.S.; Hyde, K.D. Diaporthe: Formalizing species-group 26 concepts. Mycosphere 2022, unpublished. [Google Scholar]
- Crous, P.W.; Summerell, B.A.; Shivas, R.G.; Burgess, T.I.; Decock, C.A.; Dreyer, L.L.; Granke, L.L.; Guest, D.I.; Hardy, G.E.S.J.; Hausbeck, M.K.; et al. Fungal Planet description sheets: 107–127. Persoonia 2012, 28, 138–182. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Huang, S.; Xia, J.; Zhang, X.; Li, Z. Morphological and molecular identification of Diaporthe species in south-western China, with description of eight new species. MycoKeys 2021, 77, 65–95. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.T.; Xia, J.W.; Zhang, X.G.; Sun, W.X. Morphological and phylogenetic analyses reveal three new species of Diaporthe from Yunnan, China. MycoKeys 2021, 78, 49–77. [Google Scholar] [CrossRef]
- Santos, J.M.; Vrandecic, K.; Cosic, J.; Duvnjak, T.; Phillips, A.J.L. Resolving the Diaporthe species occuring on soybean. Persoonia 2011, 27, 9–19. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity: Up to 96% of fungi in northern Thailand may be novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Retrieved 27 March 2022. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 20 January 2022).
- Wrona, C.J.; Mohankumar, V.; Schoeman, M.H.; Tan, Y.P.; Shivas, R.G.; Jeff-Ego, O.S.; Akinsanmi, O.A. Phomopsis husk rot of macadamia in Australia and South Africa caused by novel Diaporthe species. Plant Pathol. 2020, 69, 911–921. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Hyde, K.D.; Soytong, K. The fungal endophyte dilemma. Fungal Divers. 2008, 33, 163–173. [Google Scholar]
- Ghimire, S.R.; Charlton, N.D.; Bell, J.D.; Krishnamurthy, Y.L.; Craven, K.D. Biodiversity of fungal endophyte communities inhabiting switchgrass (Panicum virgatum L.) growing in the native tall grass prairie of northern Oklahoma. Fungal Divers. 2011, 47, 19–27. [Google Scholar] [CrossRef]
- Skaltsas, D.; Castlebury, L.; Chaverri, P. Delimitation of tropical endophytic Phomopsis species from three euphorbiaceous hosts: Hevea brasiliensis, H. guianensis, and Micandra sp. Inoculum 2011, 62, 41. [Google Scholar]
- Rhoden, S.A.; Garcia, A.; Rubin Filho, C.J.; Azevedo, J.L.; Pamphile, J.A. Phylogenetic diversity of endophytic leaf fungus isolates from the medicinal tree Trichilia elegans (meliaceae). Genet. Mol. Res. GMR 2012, 11, 2513–2522. [Google Scholar] [CrossRef]
- Doilom, M.; Dissanayake, A.J.; Wanasinghe, D.N.; Boonmee, S.; Liu, J.K.; Bhat, D.J.; Taylor, J.E.; Bahkali, A.H.; McKenzie, E.H.C.; Hyde, K.D. Microfungi on Tectona grandis (teak) in Northern Thailand. Fungal Divers. 2016, 82, 107–182. [Google Scholar] [CrossRef]
- Choi, J.; Lee, K.T.; Choi, M.Y.; Nam, J.H.; Jung, H.J.; Park, S.K.; Park, H.J. Antinociceptive antiinflammatory effect of monotropein isolated from the root of Morinda officinalis. Biol. Pharm. Bull. 2005, 28, 1915–1918. [Google Scholar] [CrossRef]
- Wu, Y.B.; Zheng, C.J.; Qin, L.P.; Sun, L.N.; Han, T.; Jiao, L.; Zhang, Q.Y.; Wu, J.Z. Antiosteoporotic activity of anthraquinones from Morinda officinalis on osteoblasts and osteoclasts. Molecules 2009, 14, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Li, J.; Li, G.; Wang, D.M.; Zhu, L.P.; Yang, D.P. Structural characterization and anti-fatigue activity of polysaccharides from the roots of Morinda officinalis. Int. J. Biol. Macromol. 2009, 44, 257–261. [Google Scholar] [CrossRef]
- Wang, F.; Wu, L.; Li, L.; Chen, S. Monotropein exerts protective effects against IL-1β-induced apoptosis and catabolic responses on osteoarthritis chondrocytes. Int. Immunopharmacol. 2014, 23, 575–580. [Google Scholar] [CrossRef]
- Wu, Z.Q.; Chen, D.L.; Lin, F.H.; Lin, L.; Shuai, O.; Wang, J.Y.; Qi, L.K.; Zhang, P. Effect of bajijiasu isolated from Morinda officinalis F. C. How on sexual function in male mice and its antioxidant protection of human sperm. J. Ethnopharmacol. 2015, 164, 283–292. [Google Scholar] [CrossRef]
- Carvalho, C.R.; Gonalves, V.N.; Pereira, C.B.; Johann, S.; Galliza, I.V.; Alves, T.M.A.; Rabello, A.; Sobral, M.E.G.; Zani, C.; Rosa, C.A.; et al. The diversity, antimicrobial and anticancer activity of endophytic fungi associated with the medicinal plant Stryphnodendron adstringens (mart.) coville (fabaceae) from the brazilian savannah. Symbiosis 2012, 57, 95–107. [Google Scholar] [CrossRef]
- Carrión, V.J.; Perez-Jaramillo, J.; Cordovez, V.; Tracanna, V.; de Hollander, M.; Ruiz-Buck, D.; Mendes, L.W.; Van Ijcken, W.F.J.; Gomez-Exposito, R.; Elsayed, S.S.; et al. Pathogen-induced activation of disease-suppressive functions in the endophytic root microbiome. Science 2019, 366, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Wonglom, P.; Ito, S.I.; Sunpapao, A. Volatile organic compounds emitted from endophytic fungus Trichoderma asperellum t1 mediate antifungal activity, defense response and promote plant growth in lettuce (Lactuca sativa). Fungal Ecol. 2020, 43, 100867. [Google Scholar] [CrossRef]
- Estrada, C.; Wcislo, W.T.; Van Bael, S.A. Symbiotic fungi alter plant chemistry that discourages leaf-cutting ants. New Phytol. 2013, 198, 241–251. [Google Scholar] [CrossRef]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical ecology of endophytic fungi: Origins of secondary metabolites. Chem. Biol. 2012, 19, 792–798. [Google Scholar] [CrossRef]
- Ying, W.; Liu, H.X.; Chen, Y.C.; Sun, Z.H.; Li, H.H.; Li, S.N.; Yan, M.L.; Zhang, W.M. Two new metabolites from the endophytic fungus Alternaria sp. A744 derived from Morinda officinalis. Molecules 2017, 22, 765. [Google Scholar]
- Purahong, W.; Hyde, K.D. Effects of fungal endophytes on grass and non-grass litter decomposition rates. Fungal Divers. 2011, 47, 1–7. [Google Scholar] [CrossRef]
- Chen, S.; Liu, H.; Liu, Z.; Li, S.; Chen, Y.; Li, H.; Li, D.; Zhang, W. Two New Polyketide Compounds from the Endophytic Fungus Trichoderma spirale A725 of Morinda officinalis. Chin. J. Org. Chem. 2020, 40, 209–214. [Google Scholar] [CrossRef]
- Gond, S.K.; Verma, V.C.; Kumar, A.; Kumar, V.; Kharwar, R.N. Study of endophytic fungal community from different parts of Aegle marmelos correae (rutaceae) from varanasi (India). World J. Microbiol. Biotechnol. 2007, 23, 1371–1375. [Google Scholar] [CrossRef]
- Rana, B.K.; Singh, U.P.; Taneja, D. Antifungal activity and kinetics of inhibition by essential oil isolated from leaves of Aegle marmelos. J. Ethnopharmacol. 1997, 57, 29–34. [Google Scholar] [CrossRef]
- Zhou, S.Y.; Zhou, X.; Zhang, Z.F.; Liang, J.M.; Cai, L. Attempt to isolate hitherto uncultured fungi. Mycosystema 2020, 39, 766–776. [Google Scholar]
- Begoude, B.A.D.; Slippers, B.; Wingfield, M.J.; Roux, J. The pathogenic potential of endophytic Botryosphaeriaceous fungi on Terminalia species in Cameroon. For. Pathol. 2011, 41, 281–292. [Google Scholar] [CrossRef][Green Version]
- de Silva, D.D.; Ades, P.K.; Crous, P.W.; Taylor, P.W.J. Colletotrichum species associated with chili anthracnose in Australia. Plant Pathol. 2017, 66, 254–267. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, X.L.; Guarnaccia, V.; Tian, C.M. High diversity of Diaporthe species associated with dieback diseases in China, with twelve new species described. MycoKeys 2018, 39, 97–149. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, A.J.; Zhang, W.; Liu, M.; Hyde, K.D.; Zhao, W.S.; Li, X.H.; Yan, J.Y. Diaporthe species associated with peach tree dieback in Hubei, China. Mycosphere 2017, 8, 533–549. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Region | Primers | Optimized PCR Protocols | References |
|---|---|---|---|
| ITS | ITS1 | (94 °C: 30 s, 55 °C: 50 s, 72 °C: 1 min) × 35 cycles | [22] |
| ITS4 | |||
| tef1-α | EF1-728F | (95 °C: 30 s, 58 °C: 30 s, 72 °C: 1 min) × 35 cycles | [23] |
| EF1-986R | |||
| cal | CAL-228F | (95 °C: 30 s, 55 °C: 50 s, 72 °C: 1 min) × 35 cycles | [23] |
| CAL-737R | |||
| his | CYLH4F | (95 °C: 30 s, 58 °C: 30 s, 72 °C: 1 min) × 35 cycles | [24] |
| H3-1b | [25] | ||
| β-tubulin | BT2a | (94 °C: 30 s, 58 °C: 50 s, 72 °C: 1 min) × 35 cycles | [24] |
| Bt2b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, M.; Guo, W.; Zhao, M.; Manawasinghe, I.S.; Guarnaccia, V.; Liu, J.; Hyde, K.D.; Dong, Z.; You, C. Endophytic Diaporthe Associated with Morinda officinalis in China. J. Fungi 2022, 8, 806. https://doi.org/10.3390/jof8080806
Luo M, Guo W, Zhao M, Manawasinghe IS, Guarnaccia V, Liu J, Hyde KD, Dong Z, You C. Endophytic Diaporthe Associated with Morinda officinalis in China. Journal of Fungi. 2022; 8(8):806. https://doi.org/10.3390/jof8080806
Chicago/Turabian StyleLuo, Mei, Wei Guo, Minping Zhao, Ishara S. Manawasinghe, Vladimiro Guarnaccia, Jiawei Liu, Kevin D. Hyde, Zhangyong Dong, and Chunping You. 2022. "Endophytic Diaporthe Associated with Morinda officinalis in China" Journal of Fungi 8, no. 8: 806. https://doi.org/10.3390/jof8080806
APA StyleLuo, M., Guo, W., Zhao, M., Manawasinghe, I. S., Guarnaccia, V., Liu, J., Hyde, K. D., Dong, Z., & You, C. (2022). Endophytic Diaporthe Associated with Morinda officinalis in China. Journal of Fungi, 8(8), 806. https://doi.org/10.3390/jof8080806