
Morpho-Phylogenetic Evidence Reveals Novel Pleosporalean Taxa from Sichuan Province, China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Morphological Examination
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
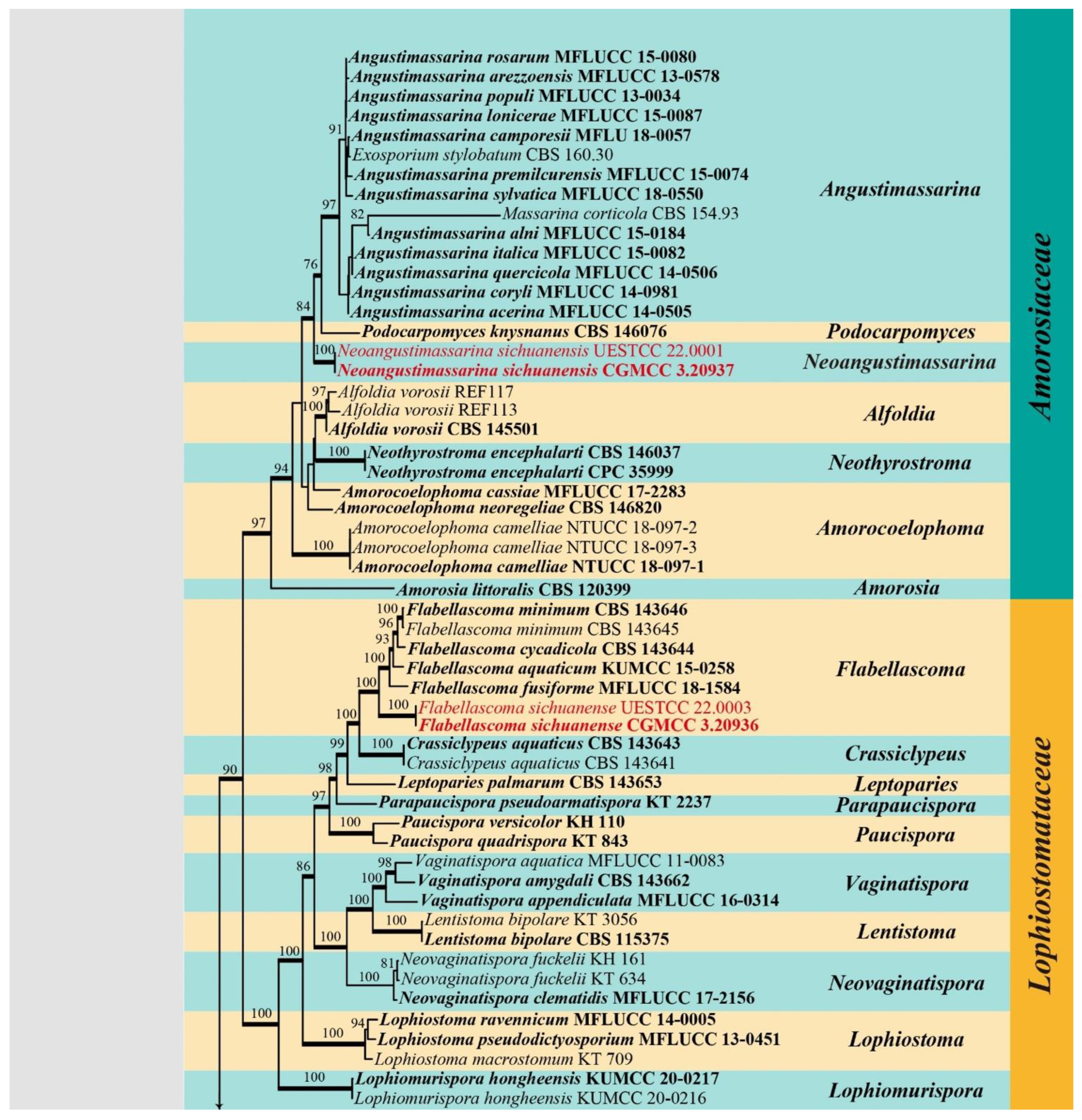
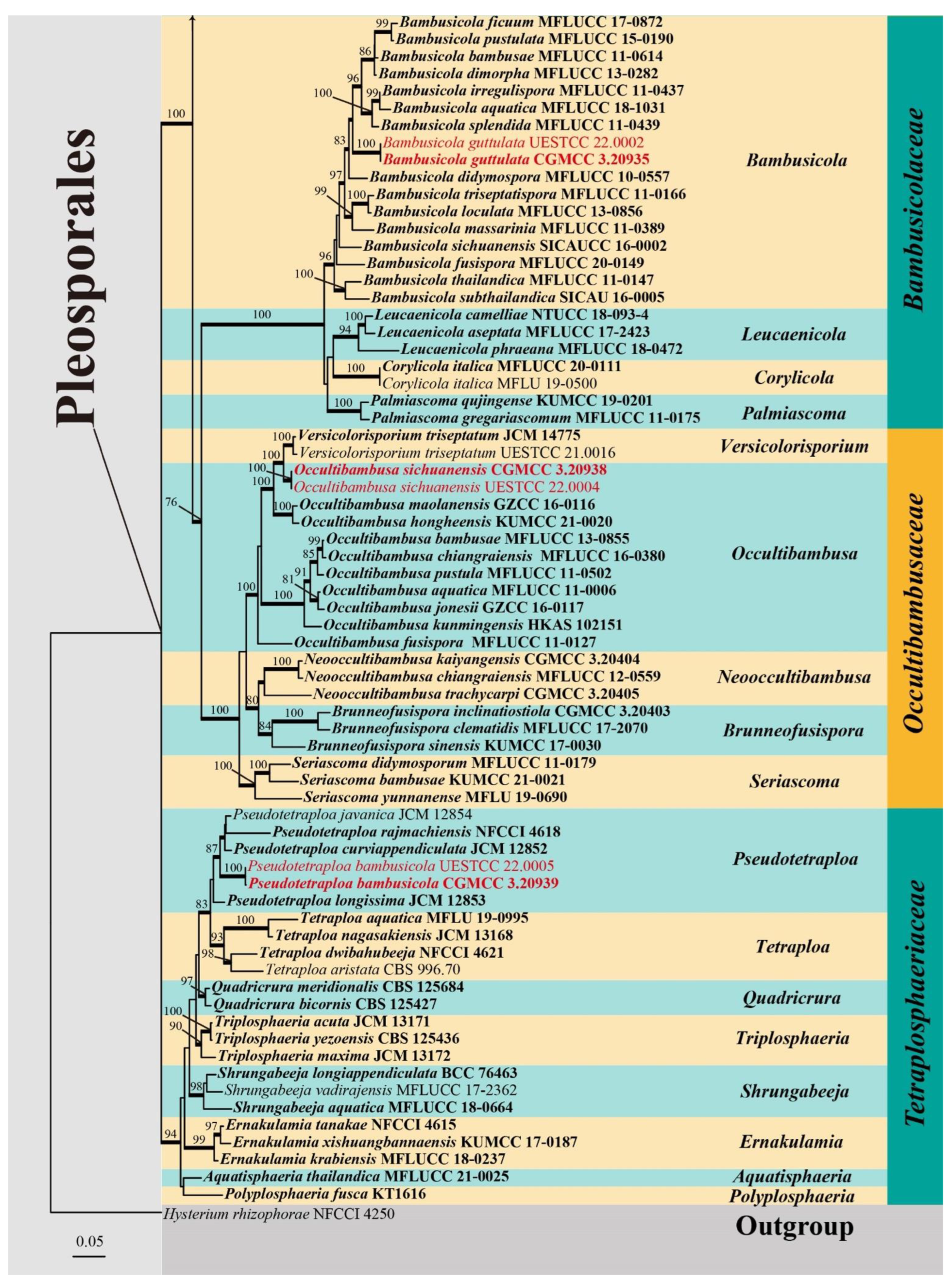
3. Results
3.1. Phylogenetic Analyses
3.2. Taxonomy
Pleosporales Luttr. ex M.E. Barr, Prodromus to class Loculoascomycetes: 67 (1987)
Amorosiaceae Thambug and K.D. Hyde, Fungal Diversity 74: 252 (2015)
Neoangustimassarina X.D. Yu and Jian K. Liu, gen. nov.
Type species: Neoangustimassarina sichuanensis X.D. Yu, S.N. Zhang and Jian K. Liu
MycoBank: MB 843716
Etymology: The name refers to the similarity to the genus Angustimassarina.
Neoangustimassarina sichuanensis X.D. Yu, S.N. Zhang and Jian K. Liu, sp. nov., Figure 2
MycoBank: MB 843717
Etymology: The epithet refers to Sichuan Province where the fungus was collected.
Holotype: HKAS 123092
Bambusicolaceae D.Q. Dai and K.D. Hyde, Fungal Diversity 63: 49 (2013)
Bambusicola D.Q. Dai and K.D. Hyde, Cryptogamie Mycologie 33: 367 (2012)
Bambusicola guttulata X.D. Yu, S.N. Zhang and Jian K. Liu, sp. nov., Figure 3.
MycoBank: MB 843718
Etymology: Referring to the conidia with large guttules.
Holotype: HKAS 123091
Lophiostomataceae Sacc., Sylloge Fungorum 2: 672 (1883)
Flabellascoma A. Hashim., K. Hiray. and Kaz. Tanaka, Studies in Mycology 90: 167 (2018)
Flabellascoma sichuanense X.D. Yu, S.N. Zhang and Jian K. Liu, sp. nov., Figure 4.
MycoBank: MB 843719
Etymology: The epithet refers to Sichuan Province where the fungus was collected.
Holotype: HKAS 123094
Occultibambusaceae D.Q. Dai and K.D. Hyde, Fungal Diversity 82: 25 (2017)
Occultibambusa D.Q. Dai and K.D. Hyde, Fungal Diversity 82: 25 (2017)
Occultibambusa sichuanensis X.D. Yu, S.N. Zhang and Jian K. Liu, sp. nov., Figure 5.
MycoBank: MB 843720
Etymology: The epithet refers to Sichuan Province where the fungus was collected.
Holotype: HKAS 123093
Tetraplosphaeriaceae Kaz. Tanaka and K. Hiray, Studies in Mycology 64: 177 (2009)
Pseudotetraploa Kaz. Tanaka and K. Hiray, Studies in Mycology 64: 193 (2009)
Pseudotetraploa bambusicola X.D. Yu, S.N. Zhang and Jian K. Liu, sp. nov., Figure 6.
MycoBank: MB 843721
Etymology: Refers to the bamboo host.
Holotype: HKAS 123095
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirk, P.; Cannon, P.; Minter, D.; Stalpers, J. Ainsworth and Bisby’s Dictionary of the Fungi, 10th ed.; CABI Publishing: Wallingford, UK, 2008; ISBN 978-0-85199-826-8. [Google Scholar]
- Mapook, A.; Boonmee, S.; Liu, J.K.; Jones, E.B.G.; Bahkali, A.H.; Hyde, K.D. Taxonomic and phylogenetic placement of Phaeodimeriella (Pseudoperisporiaceae, Pleosporales). Cryptogam. Mycol. 2016, 37, 157–176. [Google Scholar] [CrossRef]
- Ferdinandez, H.S.; Manamgoda, D.S.; Udayanga, D.; Deshappriya, N.; Munasinghe, M.S.; Castlebury, L.A. Molecular phylogeny and morphology reveal three novel species of Curvularia (Pleosporales, Pleosporaceae) associated with cereal crops and weedy grass hosts. Mycol. Prog. 2021, 20, 431–451. [Google Scholar] [CrossRef]
- Matsumura, M.; Kato, W.; Hashimoto, A.; Takahashi, Y.S.; Shirouzu, T.; Tanaka, K. Crassiperidium (Pleosporales, Dothideomycetes), a new ascomycetous genus parasitic on Fagus crenata in Japan. Mycosphere 2018, 9, 1256–1267. [Google Scholar] [CrossRef]
- Liu, Z.P.; Zhang, S.N.; Cheewangkoon, R.; Zhao, Q.; Liu, J.K. Crassoascoma gen. nov. (Lentitheciaceae, Pleosporales): Unrevealing Microfungi from the Qinghai-Tibet Plateau in China. Diversity 2022, 14, 15. [Google Scholar] [CrossRef]
- Li, W.L.; Ba, D.F.; Liu, N.G.; Hyde, K.D.; Liu, J.K. Aquatisphaeria thailandica gen. et sp. nov. (Tetraplosphaeriaceae, Pleosporales) from freshwater habitat in Thailand. Phytotaxa 2021, 513, 118–128. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Devadatha, B.; Abdel-Wahab, M.A.; Dayarathne, M.C.; Zhang, S.N.; Hyde, K.D.; Liu, J.K.; Bahkali, A.H.; Sarma, V.V.; Tibell, S.; et al. Phylogeny of new marine Dothideomycetes and Sordariomycetes from mangroves and deep-sea sediments. Bot. Mar. 2020, 63, 155–181. [Google Scholar] [CrossRef] [Green Version]
- Rupcic, Z.; Chepkirui, C.; Hernández-Restrepo, M.; Crous, P.W.; Luangsa-Ard, J.J.; Stadler, M. New nematicidal and antimicrobial secondary metabolites from a new species in the new genus, Pseudobambusicola thailandica. MycoKeys 2018, 33, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Luttrell, E.S. The Ascostromatic Ascomycetes. Mycologia 1955, 47, 511–532. [Google Scholar] [CrossRef]
- Barr, M.E. Prodromus to Class Loculoascomycetes; University of Massachusetts: Amherst, MA, USA, 1987. [Google Scholar]
- Barr, M.E. New taxa and combinations in the Loculoascomycetes. Mycotaxon 1987, 29, 501–505. [Google Scholar]
- Zhang, Y.; Crous, P.W.; Schoch, C.L.; Hyde, K.D. Pleosporales. Fungal Divers. 2012, 53, 1–221. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Jones, E.B.G.; Liu, J.K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.Q.; et al. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Sun, Y.R.; Liu, N.G.; Hyde, K.D.; Jayawardena, R.S.; Wang, Y. Pleocatenata chiangraiensis gen. et. sp. nov. (Pleosporales, Dothideomycetes) from medicinal plants in northern Thailand. MycoKeys 2022, 87, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Luecking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Wijayawardene, N.; Hyde, K.; Dai, D.Q.; Sánchez, G.; Tomio Goto, B.; Saxena, R.; Erdoğdu, M.; Selcuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Antonelli, A.; Smith, R.; Fry, C.; Simmonds, M.S.; Kersey, P.J.; Pritchard, H.; Abbo, M.; Acedo, C.; Adams, J.; Ainsworth, A. State of the World’s Plants and Fungi. Ph.D. Thesis, Royal Botanic Gardens (Kew). Sfumato Foundation, Richmond, UK, 2020. [Google Scholar]
- Wanasinghe, D.N.; Mortimer, P.E.; Xu, J. Insight into the Systematics of Microfungi Colonizing Dead Woody Twigs of Dodonaea viscosa in Honghe (China). J. Fungi 2021, 7, 180. [Google Scholar] [CrossRef]
- Cheek, M.; Nic Lughadha, E.; Kirk, P.; Lindon, H.; Carretero, J.; Looney, B.; Douglas, B.; Haelewaters, D.; Gaya, E.; Llewellyn, T. New scientific discoveries: Plants and fungi. Plants People Planet 2020, 2, 371–388. [Google Scholar] [CrossRef]
- Wang, S.; Li, H.; Niu, S. Empirical Research on Climate Warming Risks for Forest Fires: A Case Study of Grade I Forest Fire Danger Zone, Sichuan Province, China. Sustainability 2021, 13, 7773. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Wijayawardene, N.N.; Xu, J.C.; Cheewangkoon, R.; Mortimer, P.E. Taxonomic novelties in Magnolia-associated pleosporalean fungi in the Kunming Botanical Gardens (Yunnan, China). PLoS ONE 2020, 15, e0235855. [Google Scholar] [CrossRef]
- Lopez-Pujol, J.; Zhang, F.M.; Ge, S. Plant biodiversity in China: Richly varied, endangered, and in need of conservation. Biodivers. Conserv. 2006, 15, 3983–4026. [Google Scholar] [CrossRef]
- Senanayake, I.; Rathnayake, A.; Marasinghe, D.; Calabon, M.; Gentekaki, E.; Lee, H. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Liu, J.K.; Chomnunti, P.; Cai, L.; Phookamsak, R.; Chukeatirote, E.; Jones, E.B.G.; Moslem, M.; Hyde, K.D. Phylogeny and morphology of Neodeightonia palmicola sp. nov. from palms. Sydowia 2010, 62, 261–276. [Google Scholar]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Kazutaka, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Dissanayake, A.; Bhunjun, C.; Maharachchikumbura, S.; Liu, J. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2652–2676. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2011, 12, 335–337. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, UK, 2003. [Google Scholar]
- Nylander, J. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thambugala, K.M.; Hyde, K.D.; Tanaka, K.; Tian, Q.; Wanasinghe, D.N.; Ariyawansa, H.A.; Jayasiri, S.C.; Boonmee, S.; Camporesi, E.; Hashimoto, A.; et al. Towards a natural classification and backbone tree for Lophiostomataceae, Floricolaceae, and Amorosiaceae fam. nov. Fungal Divers. 2015, 74, 199–266. [Google Scholar] [CrossRef]
- Mantle, P.G.; Hawksworth, D.L.; Pazoutova, S.; Collinson, L.M.; Rassing, B.R. Amorosia littoralis gen. sp. nov., a new genus and species name for the scorpinone and caffeine-producing hyphomycete from the littoral zone in The Bahamas. Mycol. Res. 2006, 110, 1371–1378. [Google Scholar] [CrossRef]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef] [PubMed]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; Liu, J.K.; Lu, Y.Z.; et al. Diversity, morphology and molecular phylogeny of Dothideomycetes on decaying wild seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Lombard, L.; Roets, F.; Swart, W.J.; Alvarado, P.; Carnegie, A.J.; Moreno, G.; Luangsa-ard, J.; Thangavel, R.; et al. Fungal Planet description sheets: 951–1041. Persoonia 2019, 43, 223–425. [Google Scholar] [CrossRef]
- Dai, D.Q.; Bhat, D.J.; Liu, J.K.; Zhao, R.L.; Hyde, K.D. Bambusicola, a new genus from bamboo with asexual and sexual morphs. Cryptogam. Mycol. 2012, 33, 363–379. [Google Scholar] [CrossRef]
- Wijesinghe, S.N.; Wang, Y.; Camporesi, E.; Wanasinghe, D.N.; Boonmee, S.; Hyde, K.D. A new genus of Bambusicolaceae (Pleosporales) on Corylus avellana (Fagales) from Italy. Biodivers. Data J. 2020, 8, e55957. [Google Scholar] [CrossRef]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; Mckenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Dai, D.Q.; Bahkali, A.H.; Li, W.J.; Bhat, D.J.; Zhao, R.L.; Hyde, K.D. Bambusicola loculata sp. nov.(Bambusicolaceae) from bamboo. Phytotaxa 2015, 213, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Wanasinghe, D.N.; Phillips, A.J.L.; Camporesi, E.; Bulgakov, T.S.; Phukhamsakda, C.; Ariyawansa, H.A.; Goonasekara, I.D.; Phookamsak, R.; Dissanayake, A.; et al. Mycosphere notes 1–50: Grass (Poaceae) inhabiting Dothideomycetes. Mycosphere 2017, 8, 697–796. [Google Scholar] [CrossRef]
- Yang, C.L.; Xu, X.L.; Liu, Y.G. Two new species of Bambusicola (Bambusicolaceae, Pleosporales) on Phyllostachys heteroclada from Sichuan, China. Nova Hedwig. 2019, 108, 527–545. [Google Scholar] [CrossRef]
- Monkai, J.; Wanasinghe, D.N.; Jeewon, R.; Promputtha, I.; Phookamsak, R. Morphological and phylogenetic characterization of fungi within Bambusicolaceae: Introducing two new species from the Greater Mekong Subregion. Mycol. Prog. 2021, 20, 721–732. [Google Scholar] [CrossRef]
- Brahmanage, R.S.; Dayarathne, M.C.; Wanasinghe, D.N.; Thambugala, K.M.; Jeewon, R.; Chethana, K.W.T.; Samarakoon, M.C.; Tennakoon, D.S.; De Silva, N.I.; Camporesi, E.; et al. Taxonomic novelties of saprobic Pleosporales from selected dicotyledons and grasses. Mycosphere 2020, 11, 2481–2541. [Google Scholar] [CrossRef]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes. Fungal Divers. 2020, 105, 319–575. [Google Scholar] [CrossRef]
- Nitschke, T. Grundlage eines systems der Pyrenomyceten. Verh. Des Nat. Ver. Der Preuss. Rheinl. Westfal. Und Des Regier. Osnabrück 1869, 26, 70–77. [Google Scholar]
- Saccardo, P.A. Sylloge Fungorum Omnium Hucusque Cognitorum. Sylloge Fungorum 1883, 2, 672. [Google Scholar]
- Maharachchikumbura, S.S.; Wanasinghe, D.N.; Cheewangkoon, R.; Al-Sadi, A.M. Uncovering the hidden taxonomic diversity of fungi in Oman. Fungal Divers. 2021, 106, 229–268. [Google Scholar] [CrossRef]
- Hashimoto, A.; Hirayama, K.; Takahashi, H.; Matsumura, M.; Okada, G.; Chen, C.Y.; Huang, J.W.; Kakishima, M.; Ono, T.; Tanaka, K. Resolving the Lophiostoma bipolare complex: Generic delimitations within Lophiostomataceae. Stud. Mycol. 2018, 90, 161–189. [Google Scholar] [CrossRef]
- Bao, D.F.; Su, H.Y.; Maharachchikumbura, S.S.N.; Liu, J.K.; Nalumpang, S.; Luo, Z.L.; Hyde, K.D. Lignicolous freshwater fungi from China and Thailand: Multi-gene phylogeny reveals new species and new records in Lophiostomataceae. Mycosphere 2019, 10, 1080–1099. [Google Scholar] [CrossRef]
- Doilom, M.; Dissanayake, A.J.; Wanasinghe, D.N.; Boonmee, S.; Liu, J.K.; Bhat, D.J.; Taylor, J.E.; Bahkali, A.H.; Mckenzie, E.H.C.; Hyde, K.D. Microfungi on Tectona grandis (teak) in Northern Thailand. Fungal Divers. 2017, 82, 107–182. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.B.; Phookamsak, R.; Hyde, K.D.; Mortimer, P.E.; Xu, J.C.; Kakumyan, P.; Karunarathna, S.C.; Kumla, J. A Taxonomic Appraisal of Bambusicolous Fungi in Occultibambusaceae (Pleosporales, Dothideomycetes) with New Collections from Yunnan Province, China. Life 2021, 11, 932. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.D.; Zhang, S.N.; Cheewangkoon, R.; Liu, J.K. Additions to Occultibambusaceae (Pleosporales, Dothideomycetes): Unrevealing Palmicolous Fungi in China. Diversity 2021, 13, 516. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.; Jones, E.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Zhang, J.F.; Liu, J.K.; Hyde, K.D.; Yang, W.; Liu, Z.Y. Fungi from Asian Karst formations II. Two new species of Occultibambusa (Occultibambusaceae, Dothideomycetes) from karst landforms of China. Mycosphere 2017, 8, 550–559. [Google Scholar] [CrossRef]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Hatakeyama, S.; Harada, Y.; Sano, T.; Shirouzu, T.; Hosoya, T. Molecular taxonomy of bambusicolous fungi: Tetraplosphaeriaceae, a new pleosporalean family with Tetraploa-like anamorphs. Stud. Mycol. 2009, 64, 175–209. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Kelchner, S.A. Higher level phylogenetic relationships within the bamboos (Poaceae: Bambusoideae) based on five plastid markers. Mol. Phylogenet. Evol. 2013, 67, 404–413. [Google Scholar] [CrossRef]
- Feng, Y.; Liu, J.K.; Lin, C.G.; Chen, Y.Y.; Xiang, M.M.; Liu, Z.Y. Additions to the Genus Arthrinium (Apiosporaceae) From Bamboos in China. Front. Microbiol. 2021, 720, 12. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.Q.; Tang, L.Z.; Wang, H.B. A Review of Bambusicolous Ascomycetes. Bamboo Curr. Future Prospect. 2018, 165, 10. [Google Scholar] [CrossRef] [Green Version]
- Dlamini, L.C.; Fakudze, S.; Makombe, G.G.; Muse, S.; Zhu, J. Bamboo as a Valuable Resource and its Utilization in Historical and Modern-day China. BioResources 2022, 17, 1926–1938. [Google Scholar] [CrossRef]
- Sun, Y.R.; Goonasekara, I.D.; Thambugala, K.M.; Jayawardena, R.S.; Wang, Y.; Hyde, K.D. Distoseptispora bambusae sp. nov. (Distoseptisporaceae) on bamboo from China and Thailand. Biodivers. Data J. 2020, 8, e53678. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.B.; Zhang, S.J.; Phookamsak, R.; Promputtha, I.; Kakumyan, P.; Xu, J.C. Amphibambusa hongheensis sp. nov., a novel bambusicolous ascomycete from Yunnan, China. Phytotaxa 2021, 505, 201–212. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | SSU | ITS, LSU, RPB2 | TEF1α | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Temperature | Time | Cycles | Temperature | Time | Cycles | Temperature | Time | Cycles | |
| Initial Denaturation | 98 °C | 2 min | 1 | 98 °C | 2 min | 1 | 98 °C | 2 min | 1 |
| Denaturation | 98 °C | 10 s | 35 | 98 °C | 10 s | 35 | 98 °C | 10 s | 35 |
| Annealing | 47 °C | 10 s | 56 °C | 10 s | 61.7 °C | 10 s | |||
| Extension | 72 °C | 10 s | 72 °C | 10 s | 72 °C | 10 s | |||
| Final Extension | 72 °C | 5 min | 1 | 72 °C | 5 min | 1 | 72 °C | 5 min | 1 |
| Hold | 4 °C | - | - | 4 °C | - | - | 4 °C | - | - |
| Species | Voucher/Strain/Isolate | GenBank Accession Number | ||||
|---|---|---|---|---|---|---|
| SSU | ITS | LSU | RPB2 | TEF1α | ||
| Alfoldia vorosii | CBS 145501 | MK589346 | JN859336 | MK589354 | N/A | MK599320 |
| Alfoldia vorosii | REF113 | MK589345 | JN859333 | MK589353 | N/A | MK599319 |
| Alfoldia vorosii | REF117 | MK589347 | JN859337 | MK589355 | N/A | MK599321 |
| Amorocoelophoma camelliae | NTUCC 18-097-1 | MT071230 | MT112303 | MT071279 | MT459143 | MT743271 |
| Amorocoelophoma camelliae | NTUCC 18-097-2 | MT071231 | MT112304 | MT071280 | MT459141 | MT743272 |
| Amorocoelophoma camelliae | NTUCC 18-097-3 | MT071232 | MT112305 | MT071281 | MT459142 | MT743273 |
| Amorocoelophoma cassiae | MFLUCC 17-2283 | MK347847 | MK347739 | MK347956 | MK434894 | MK360041 |
| Amorocoelophoma neoregeliae | CBS 146820 | N/A | MZ064410 | MZ064467 | MZ078193 | MZ078247 |
| Amorosia littoralis | CBS 120399 | AM292056 | AM292047 | AM292055 | N/A | N/A |
| Angustimassarina acerina | MFLUCC 14-0505 | KP899123 | KP899132 | KP888637 | N/A | KR075168 |
| Angustimassarina arezzoensis | MFLUCC 13-0578 | KY501113 | KY496743 | KY496722 | N/A | KY514392 |
| Angustimassarina camporesii | MFLU 18-0057 | MN244173 | MN244197 | MN244167 | N/A | N/A |
| Angustimassarina italica | MFLUCC 15-0082 | KY501124 | KY496756 | KY496736 | N/A | KY514400 |
| Angustimassarina lonicerae | MFLUCC 15-0087 | N/A | KY496759 | KY496724 | N/A | N/A |
| Angustimassarina populi | MFLUCC 13-0034 | KP899128 | KP899137 | KP888642 | N/A | KR075164 |
| Angustimassarina premilcurensis | MFLUCC 15-0074 | N/A | KY496745 | KY496725 | KY514404 | N/A |
| Angustimassarina quercicola | MFLUCC 14-0506 | KP899124 | KP899133 | KP888638 | N/A | KR075169 |
| Angustimassarina rosarum | MFLUCC 15-0080 | N/A | MG828869 | MG828985 | N/A | N/A |
| Angustimassarina sylvatica | MFLUCC 18-0550 | MK314097 | MK307843 | MK307844 | N/A | MK360181 |
| Angustimassarina alni | MFLUCC 15-0184 | KY548098 | KY548099 | KY548097 | N/A | N/A |
| Angustimassarina coryli | MFLUCC 14-0981 | N/A | MF167431 | MF167432 | N/A | MF167433 |
| Aquatisphaeria thailandica | MFLUCC 21-0025 | MW890967 | MW890969 | MW890763 | N/A | N/A |
| Bambusicola aquatica | MFLUCC 18-1031 | MT864293 | MT627729 | MN913710 | MT878462 | MT954392 |
| Bambusicola bambusae | MFLUCC 11-0614 | JX442039 | JX442031 | JX442035 | KP761718 | KP761722 |
| Bambusicola didymospora | MFLUCC 10-0557 | KU872110 | KU940116 | KU863105 | KU940163 | KU940188 |
| Bambusicola dimorpha | MFLUCC 13-0282 | KY038354 | KY026582 | KY000661 | KY056663 | N/A |
| Bambusicola ficuum | MFLUCC 17-0872 | MT215581 | N/A | MT215580 | N/A | MT199326 |
| Bambusicola fusispora | MFLUCC 20-0149 | MW076529 | MW076532 | MW076531 | MW034589 | N/A |
| Bambusicola guttulata * | CGMCC 3.20935 | ON332919 | ON332909 | ON332927 | ON383985 | ON381177 |
| Bambusicola guttulata * | UESTCC 22.0002 | ON332920 | ON332910 | ON332928 | ON383986 | ON381178 |
| Bambusicola irregulispora | MFLUCC 11-0437 | JX442040 | JX442032 | JX442036 | KP761719 | KP761723 |
| Bambusicola loculata | MFLUCC 13-0856 | KP761735 | KP761732 | KP761729 | KP761715 | KP761724 |
| Bambusicola massarinia | MFLUCC 11-0389 | JX442041 | JX442033 | JX442037 | KP761716 | KP761725 |
| Bambusicola pustulata | MFLUCC 15-0190 | KU872112 | KU940118 | KU863107 | KU940165 | KU940190 |
| Bambusicola sichuanensis | SICAUCC 16-0002 | MK253528 | MK253473 | MK253532 | MK262830 | MK262828 |
| Bambusicola splendida | MFLUCC 11-0439 | JX442042 | JX442034 | JX442038 | KP761717 | KP761726 |
| Bambusicola subthailandica | SICAU 16-0005 | MK253529 | MK253474 | MK253533 | MK262831 | MK262829 |
| Bambusicola thailandica | MFLUCC 11-0147 | N/A | KU940119 | KU863108 | KU940166 | KU940191 |
| Bambusicola triseptatispora | MFLUCC 11-0166 | N/A | KU940120 | KU863109 | KU940167 | N/A |
| Brunneofusispora clematidis | MFLUCC 17-2070 | MT226685 | MT310615 | MT214570 | MT394692 | MT394629 |
| Brunneofusispora inclinatiostiola | CGMCC 3.20403 | MZ964884 | MZ964866 | MZ964875 | OK061075 | OK061069 |
| Brunneofusispora sinensis | KUMCC 17-0030 | MH393556 | MH393558 | MH393557 | N/A | MH395329 |
| Corylicola italica | MFLU 19-0500 | MT554923 | MT554925 | MT554926 | MT590776 | N/A |
| Corylicola italica | MFLUCC 20-0111 | MT633084 | MT633085 | MT626713 | MT635596 | MT590777 |
| Crassiclypeus aquaticus | CBS 143641 | LC312470 | LC312499 | LC312528 | LC312586 | LC312557 |
| Crassiclypeus aquaticus | CBS 143643 | LC312472 | LC312501 | LC312530 | LC312588 | LC312559 |
| Ernakulamia krabiensis | MFLUCC 18–0237 | MK347880 | MK347773 | MK347990 | N/A | N/A |
| Ernakulamia tanakae | NFCCI 4615 | N/A | MN937229 | MN937211 | N/A | N/A |
| Ernakulamia xishuangbannaensis | KUMCC 17-0187 | MH260354 | MH275080 | MH260314 | N/A | N/A |
| Exosporium stylobatum | CBS 160.30 | N/A | JQ044428 | JQ044447 | N/A | N/A |
| Flabellascoma aquaticum | KUMCC 15-0258 | MN304832 | MN304827 | MN274564 | MN328895 | MN328898 |
| Flabellascoma cycadicola | CBS 143644 | LC312473 | LC312502 | LC312531 | LC312589 | LC312560 |
| Flabellascoma fusiforme | MFLUCC 18-1584 | N/A | MN304830 | MN274567 | N/A | MN328902 |
| Flabellascoma minimum | CBS 143645 | LC312474 | LC312503 | LC312532 | LC312590 | LC312561 |
| Flabellascoma minimum | CBS 143646 | LC312475 | LC312504 | LC312533 | LC312591 | LC312562 |
| Flabellascoma sichuanense * | CGMCC 3.20936 | ON332921 | ON332911 | ON332929 | ON383987 | ON381179 |
| Flabellascoma sichuanense * | UESTCC 22.0003 | ON332922 | ON332912 | ON332930 | ON383988 | ON381180 |
| Hysterium rhizophorae | NFCCI-4250 | MG844280 | MG844284 | MG844276 | MG968956 | N/A |
| Lentistoma bipolare | CBS 115375 | LC312477 | LC312506 | LC312535 | LC312593 | LC312564 |
| Lentistoma bipolare | KT 3056 | LC312484 | LC312513 | LC312542 | LC312600 | LC312571 |
| Leptoparies palmarum | CBS 143653 | LC312485 | LC312514 | LC312543 | LC312601 | LC312572 |
| Leucaenicola aseptata | MFLUCC 17-2423 | MK347853 | MK347746 | MK347963 | MK434891 | MK360059 |
| Leucaenicola camelliae | NTUCC 18-093-4 | MT071229 | MT112302 | MT071278 | MT743283 | MT374091 |
| Leucaenicola phraeana | MFLUCC 18-0472 | MK347892 | MK347785 | MK348003 | MK434867 | MK360060 |
| Lophiomurispora hongheensis | KUMCC 20-0216 | MW264227 | MW264218 | MW264197 | MW256810 | MW256819 |
| Lophiomurispora hongheensis | KUMCC 20-0217 | MW264225 | MW264216 | MW264195 | MW256808 | MW256817 |
| Lophiostoma macrostomum | KT 709/HHUF 27293 | AB521732 | AB433276 | AB433274 | JN993493 | LC001753 |
| Lophiostoma pseudodictyosporium | MFLUCC 13-0451 | N/A | KR025858 | KR025862 | N/A | N/A |
| Lophiostoma ravennicum | MFLUCC 14-0005 | KP698415 | KP698413 | KP698414 | N/A | N/A |
| Massarina corticola | CBS 154.93 | FJ795491 | N/A | FJ795448 | FJ795465 | N/A |
| Neoangustimassarina sichuanensis * | CGMCC 3.20937 | ON332917 | ON332907 | ON332925 | ON383983 | ON381175 |
| Neoangustimassarina sichuanensis * | UESTCC 22.0001 | ON332918 | ON332908 | ON332926 | ON383984 | ON381176 |
| Neooccultibambusa chiangraiensis | MFLUCC 12-0559 | KU712458 | KU712442 | KU764699 | N/A | KU872761 |
| Neooccultibambusa kaiyangensis | CGMCC 3.20404 | MZ964886 | MZ964868 | MZ964877 | OK061077 | OK061071 |
| Neooccultibambusa trachycarpi | CGMCC 3.20405 | MZ964888 | MZ964870 | MZ964879 | OK061079 | OK061073 |
| Neothyrostroma encephalarti | CBS 146037 | N/A | MN562104 | MN567612 | N/A | MN556830 |
| Neothyrostroma encephalarti | CPC 35999 | N/A | MN562105 | MN567613 | N/A | MN556831 |
| Neovaginatispora clematidis | MFLUCC 17–2156 | MT226676 | MT310606 | MT214559 | N/A | MT394738 |
| Neovaginatispora fuckelii | KH 161 | AB618689 | LC001731 | AB619008 | N/A | LC001749 |
| Neovaginatispora fuckelii | KT 634 | AB618690 | LC001732 | AB619009 | N/A | LC001750 |
| Occultibambusa aquatica | MFLUCC 11-0006 | KX698112 | KX698114 | KX698110 | N/A | N/A |
| Occultibambusa bambusae | MFLUCC 13-0855 | KU872116 | KU940123 | KU863112 | KU940170 | KU940193 |
| Occultibambusa chiangraiensis | MFLUCC 16-0380 | KX655551 | N/A | KX655546 | KX655566 | KX655561 |
| Occultibambusa fusispora | MFLUCC 11-0127 | N/A | KU940125 | KU863114 | KU940172 | KU940195 |
| Occultibambusa hongheensis | KUMCC 21-0020 | MZ329029 | MZ329037 | MZ329033 | N/A | MZ325467 |
| Occultibambusa jonesii | GZCC 16-0117 | KY628324 | N/A | KY628322 | KY814758 | KY814756 |
| Occultibambusa kunmingensis | HKAS 102151 | MT864342 | MT627716 | MN913733 | MT878453 | MT954407 |
| Occultibambusa maolanensis | GZCC 16-0116 | KY628325 | N/A | KY628323 | KY814759 | KY814757 |
| Occultibambusa pustula | MFLUCC 11-0502 | KU872118 | KU940126 | KU863115 | N/A | N/A |
| Occultibambusa sichuanensis * | CGMCC 3.20938 | N/A | ON332913 | ON332931 | ON383989 | ON381181 |
| Occultibambusa sichuanensis * | UESTCC 22.0004 | N/A | ON332914 | ON332932 | ON383990 | ON381182 |
| Palmiascoma gregariascomum | MFLUCC 11-0175 | KP753958 | KP744452 | KP744495 | KP998466 | N/A |
| Palmiascoma qujingense | KUMCC 19-0201 | MT477186 | MT477183 | MT477185 | MT495782 | N/A |
| Parapaucispora pseudoarmatispora | KT 2237 | LC100018 | LC100021 | LC100026 | N/A | LC100030 |
| Paucispora quadrispora | KT 843 | AB618692 | LC001734 | AB619011 | N/A | LC001755 |
| Paucispora versicolor | KH 110 | LC001721 | AB918731 | AB918732 | N/A | LC001760 |
| Podocarpomyces knysnanus | CBS 146076 | N/A | MN562155 | MN567662 | MN556816 | MN556836 |
| Polyplosphaeria fusca | KT1616 | AB524463 | AB524789 | AB524604 | N/A | N/A |
| Pseudotetraploa curviappendiculata | JCM 12852 | AB524467 | AB524792 | AB524608 | N/A | N/A |
| Pseudotetraploa javanica | JCM 12854 | AB524470 | AB524795 | AB524611 | N/A | N/A |
| Pseudotetraploa longissima | JCM 12853 | AB524471 | AB524796 | AB524612 | N/A | N/A |
| Pseudotetraploa rajmachiensis | NFCCI 4618 | N/A | MN937222 | MN937204 | N/A | N/A |
| Pseudotetraploa bambusicola * | CGMCC 3.20939 | ON332923 | ON332915 | ON332933 | ON383991 | ON381183 |
| Pseudotetraploa bambusicola * | UESTCC 22.0005 | ON332924 | ON332916 | ON332934 | ON383992 | ON381184 |
| Quadricrura bicornis | CBS 125427 | AB524472 | AB524797 | AB524613 | N/A | N/A |
| Quadricrura meridionalis | CBS 125684 | AB524473 | AB524798 | AB524614 | N/A | N/A |
| Seriascoma bambusae | KUMCC 21-0021 | MZ329031 | MZ329039 | MZ329035 | MZ325470 | MZ325468 |
| Seriascoma didymosporum | MFLUCC 11-0179 | KU872119 | KU940127 | KU863116 | KU940173 | KU940196 |
| Seriascoma yunnanense | MFLU 19-0690 | MN174694 | N/A | MN174695 | MN210324 | MN381858 |
| Shrungabeeja aquatica | MFLUCC 18-0664 | N/A | MT627722 | MT627663 | N/A | N/A |
| Shrungabeeja longiappendiculata | BCC 76463 | KT376471 | KT376474 | KT376472 | N/A | N/A |
| Shrungabeeja vadirajensis | MFLUCC 17-2362 | N/A | MT627681 | MN913685 | N/A | N/A |
| Tetraploa aquatica | MFLU 19-0995 | N/A | MT530448 | MT530452 | N/A | N/A |
| Tetraploa aristata | CBS 996.70 | AB524486 | AB524805 | AB524627 | N/A | N/A |
| Tetraploa dwibahubeeja | NFCCI 4621 | N/A | MN937226 | MN937208 | N/A | N/A |
| Tetraploa nagasakiensis | JCM 13168 | AB524489 | AB524806 | AB524630 | N/A | N/A |
| Triplosphaeria acuta | JCM 13171 | AB524492 | AB524809 | AB524633 | N/A | N/A |
| Triplosphaeria maxima | JCM 13172 | AB524496 | AB524812 | AB524637 | N/A | N/A |
| Triplosphaeria yezoensis | CBS 125436 | AB524497 | AB524813 | AB524638 | N/A | N/A |
| Vaginatispora amygdali | CBS 143662 | LC312495 | LC312524 | LC312553 | LC312611 | LC312582 |
| Vaginatispora appendiculata | MFLUCC 16-0314 | KU743219 | KU743217 | KU743218 | N/A | KU743220 |
| Vaginatispora aquatica | MFLUCC 11-0083 | KJ591575 | KJ591577 | KJ591576 | N/A | N/A |
| Versicolorisporium triseptatum | JCM 14775 | AB524501 | AB365596 | AB330081 | N/A | N/A |
| Versicolorisporium triseptatum | UESTCC 21.0016 = NMX1222 | OL741381 | OL741378 | OL741318 | N/A | N/A |
| Taxa | Ascomata (μm) | Hamathecium (μm) | Asci (μm) | Ascospores (μm) | Sheath (μm) | References |
|---|---|---|---|---|---|---|
| F. aquaticum | 280–440 × 260–390 | 1.2–2.0 | 48.0–72.0 × 8–9 | 16–18 × 4.3–5.3 | 4.7–7.0 μm wide | [57] |
| F. cycadicola | 490–530 × 600–620 | 1.0–3.0 | 67.5–88.0 × 9–12 | 17–23 × 4.5–7.0 | 7.0–10 μm long | [56] |
| F. fusiforme | 310–420 × 320–380 | 1.5–3.0 | 66.0–80.0 × 10–12 | 15–18 × 4.0–5.0 | 5.4–8.0 μm wide | [57] |
| F. minimumT | 250–320 × 350–500 | 1.5–3.0 | 45.0–77.5 × 7.5–12 | 12–17.5 × 3.5–5 | 5.5–8.0 μm long | [56] |
| F. sichuanense | 150–350 × 190–280 | 1.5–3.5 | 55.0–75.0 × 9.5–13 | 15–18 × 5.0–7.0 | 3.0–7.0 μm long, 1.5–2.5 μm wide | This study |
| Taxa | Ascomata (μm) | Asci | Ascospores | References | ||
|---|---|---|---|---|---|---|
| Morphology | Size (μm) | Morphology | Size (μm) | |||
| O. aquatica | 100–250 × 180–280 | Clavate | 73–86 × 9–13 | Narrowly fusiform, 1-septate, brownish, with a sheath | 19–25 × 3.5–6.5 | [62] |
| O. bambusaeT | 150–200 × 400–550 | Broadly cylindrical | (50–)60–80(−90) × (9.5–)11.5–14.5(−15) | Slightly broad and fusiform, 1-septate, dark brown, with a sheath | (22–)23.5–27.5× 4.5–7 | [47] |
| O. chiangraiensis | 195–295 × 352–520 | Clavate-oblong | 47–92 × 12–16 | Fusiform, (1–)3-septate, hyaline when immature, pale brown to red-brown at maturity, without a sheath | 16–24 × 5–7 | [62] |
| O. fusispora | 135–185 × 240–275 | Clavate to cylindric-clavate | (60–)65–90(−110) × (11–)12–14(−15)(−16) | Fusiform, mostly 1-septate, rarely 2–3-septate, light brown, without a sheath | (20–)22–25(−26) × 5–6(−6.5) | [47] |
| O. hongheensis | 180–340 × 400–550 | Cylindric-clavate to clavate | (78–)80–130(–137) × (18–)19–23(–25) | Fusiform, 1-septate, hyaline when young and becoming pale brown when mature, with a sheath | (25–)27–30 × (5.5–)8–9(–10) | [60] |
| O. jonesii | 196–236 × 200–260 | Broadly cylindrical to clavate | (65–)75–89(–105) × 13.5–19 | Inequilateral-fusiform, 2-celled, hyaline when young and becoming brown to grayish when mature, without a sheath | 27–33.5 × 5.5–6.5 | [63] |
| O. kunmingensis | 110–150 × 220–260 | Clavate or cylindric-clavate | 110–140(–160) × 13–16.5 | Fusiform, 1-septate, brown, without a sheath | 32–40 × 5–6.5 | [52] |
| O. maolanensis | 544–600 diameter | Broadly cylindrical to clavate | (66–)77–85(–94) × 17–20(–24) | Inequality-fusiform, 2-celled, hyaline when young and become light brown when mature, without a sheath | 25–31 × 8–10 | [63] |
| O. pustula | 150–200 × 200–300 | Cylindrical | 80–105 × 8–12 | Slightly broad-fusiform, 1-septate, hyaline to pale brown, with a sheath | 22–25 × 5–5.5 | [47] |
| O. sichuanensis | 130–180 ×340–440 | Obovoid to pyriform | 70–100 × 22–27 | Fusiform, 1-septate, brown, with a sheath | 31–41 × 6.5–10 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.-D.; Zhang, S.-N.; Liu, J.-K. Morpho-Phylogenetic Evidence Reveals Novel Pleosporalean Taxa from Sichuan Province, China. J. Fungi 2022, 8, 720. https://doi.org/10.3390/jof8070720
Yu X-D, Zhang S-N, Liu J-K. Morpho-Phylogenetic Evidence Reveals Novel Pleosporalean Taxa from Sichuan Province, China. Journal of Fungi. 2022; 8(7):720. https://doi.org/10.3390/jof8070720
Chicago/Turabian StyleYu, Xian-Dong, Sheng-Nan Zhang, and Jian-Kui Liu. 2022. "Morpho-Phylogenetic Evidence Reveals Novel Pleosporalean Taxa from Sichuan Province, China" Journal of Fungi 8, no. 7: 720. https://doi.org/10.3390/jof8070720
APA StyleYu, X.-D., Zhang, S.-N., & Liu, J.-K. (2022). Morpho-Phylogenetic Evidence Reveals Novel Pleosporalean Taxa from Sichuan Province, China. Journal of Fungi, 8(7), 720. https://doi.org/10.3390/jof8070720