
Deep Genotypic Species Delimitation of Aspergillus Section Flavi Isolated from Brazilian Foodstuffs and the Description of Aspergillus annui sp. nov. and Aspergillus saccharicola sp. nov.
 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling and Molecular Methods
2.2. Phylogenetic Analysis
2.3. Coalescence Analysis
2.4. Genetic Distance Analysis
2.5. Morphological Analysis
2.6. Secondary Metabolite and Mycotoxin Analysis
3. Results and Discussion
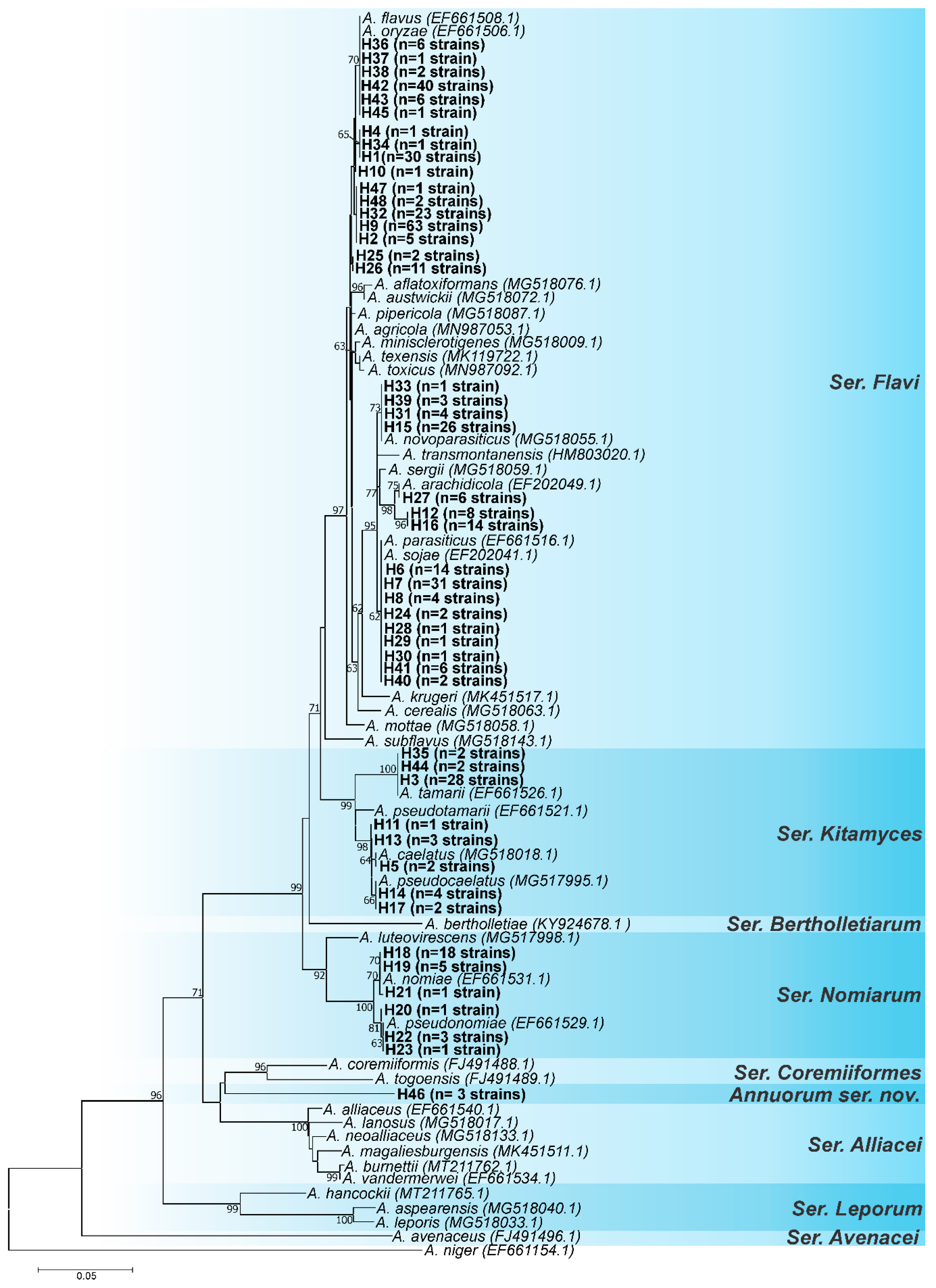
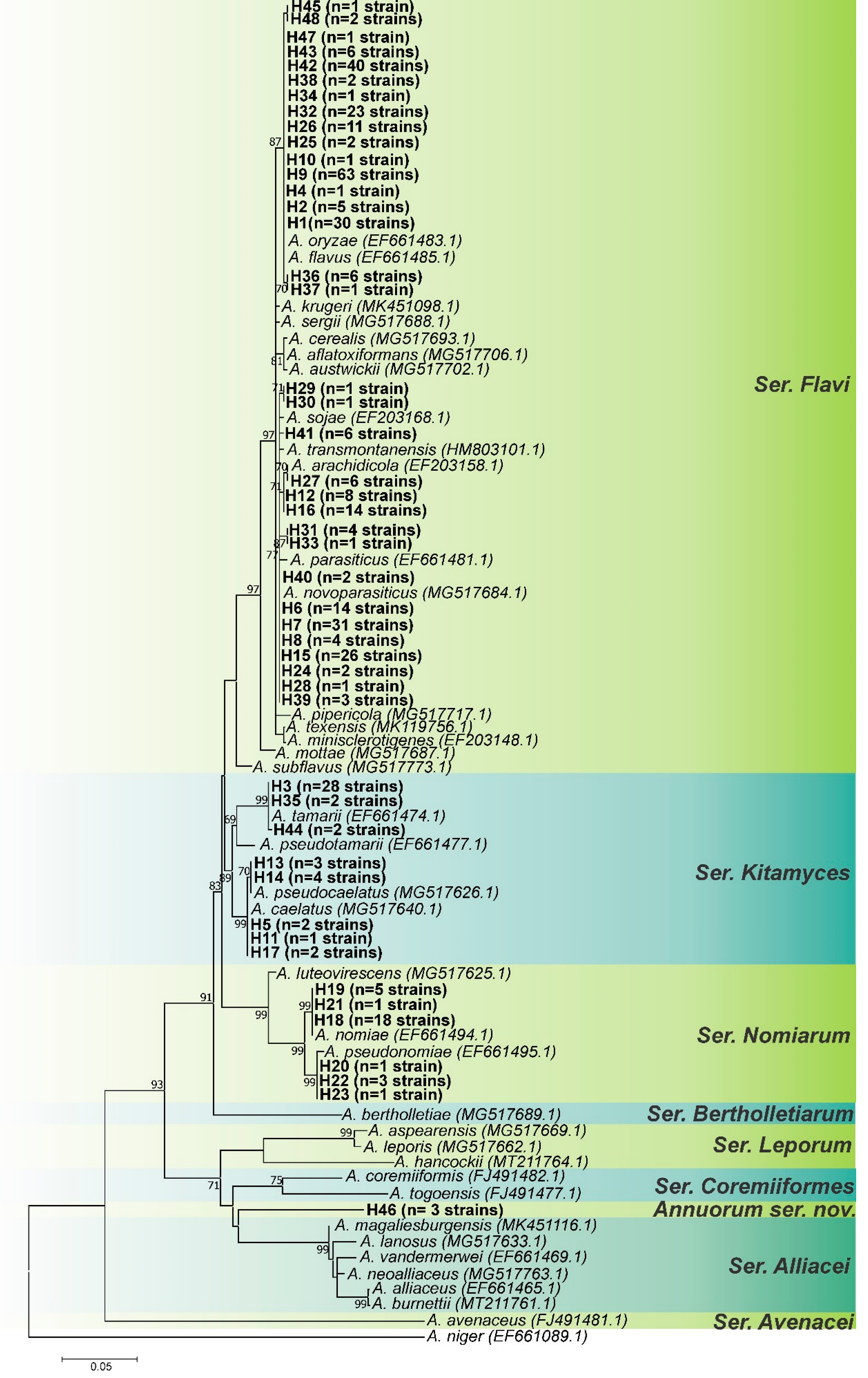
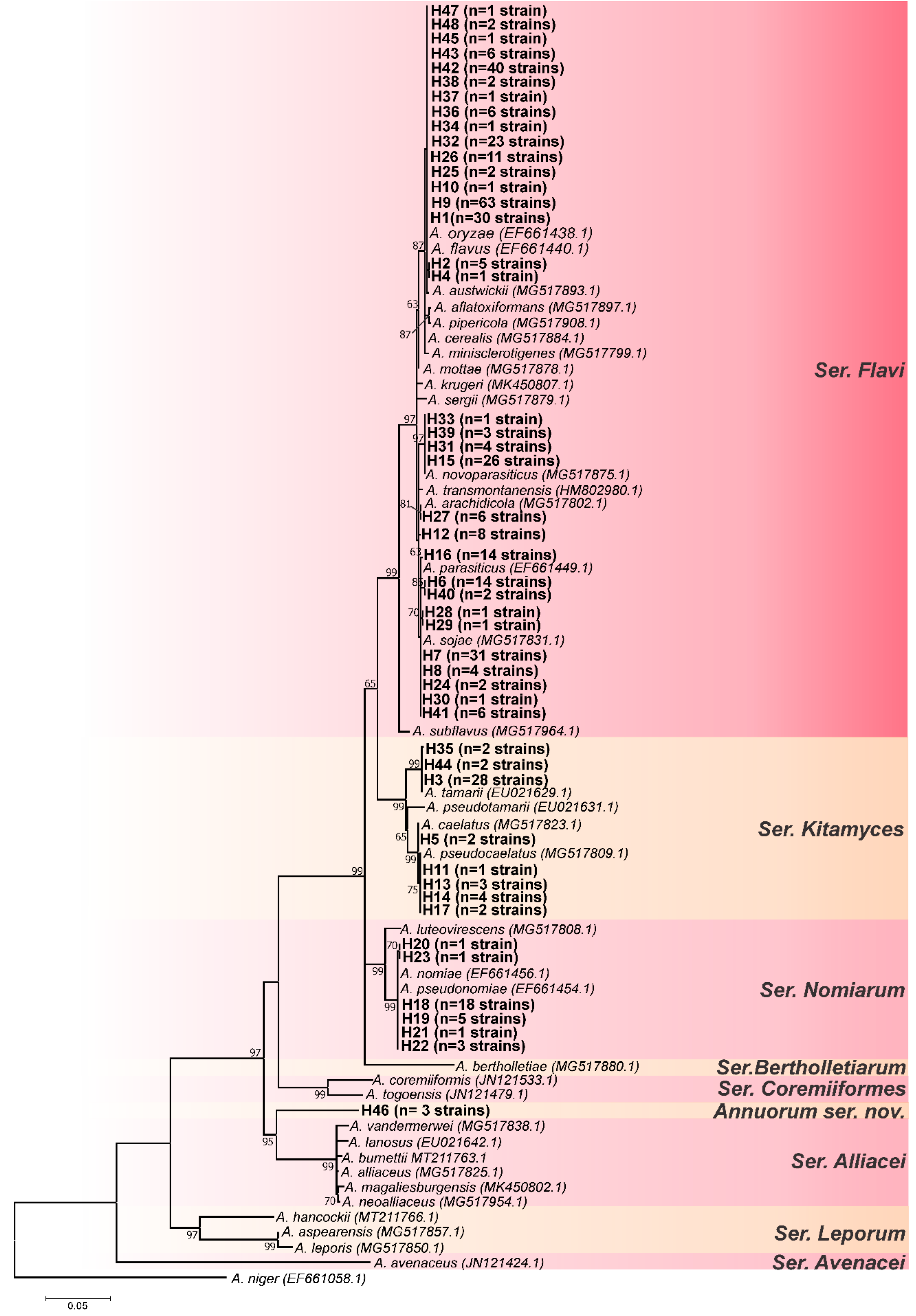
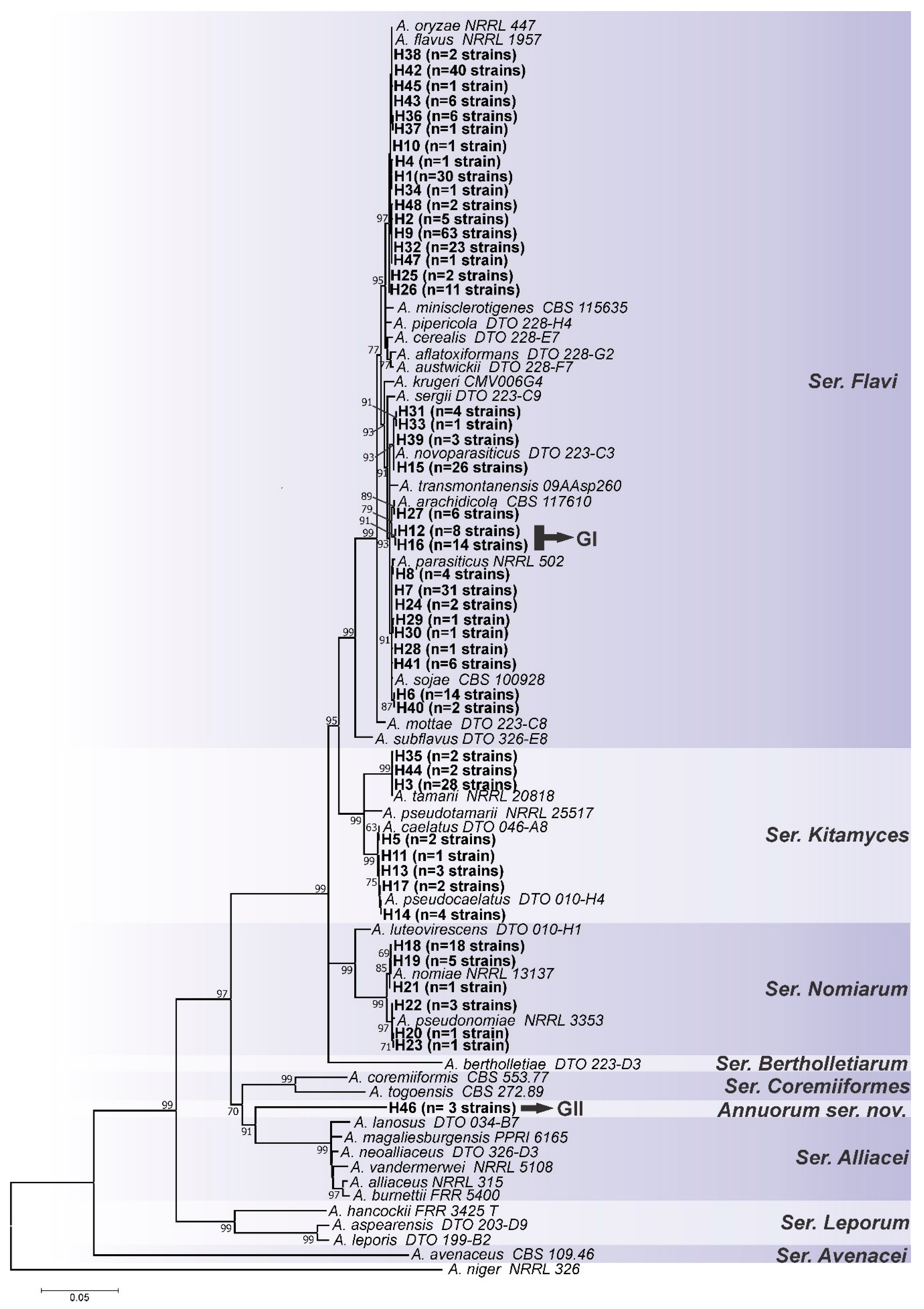
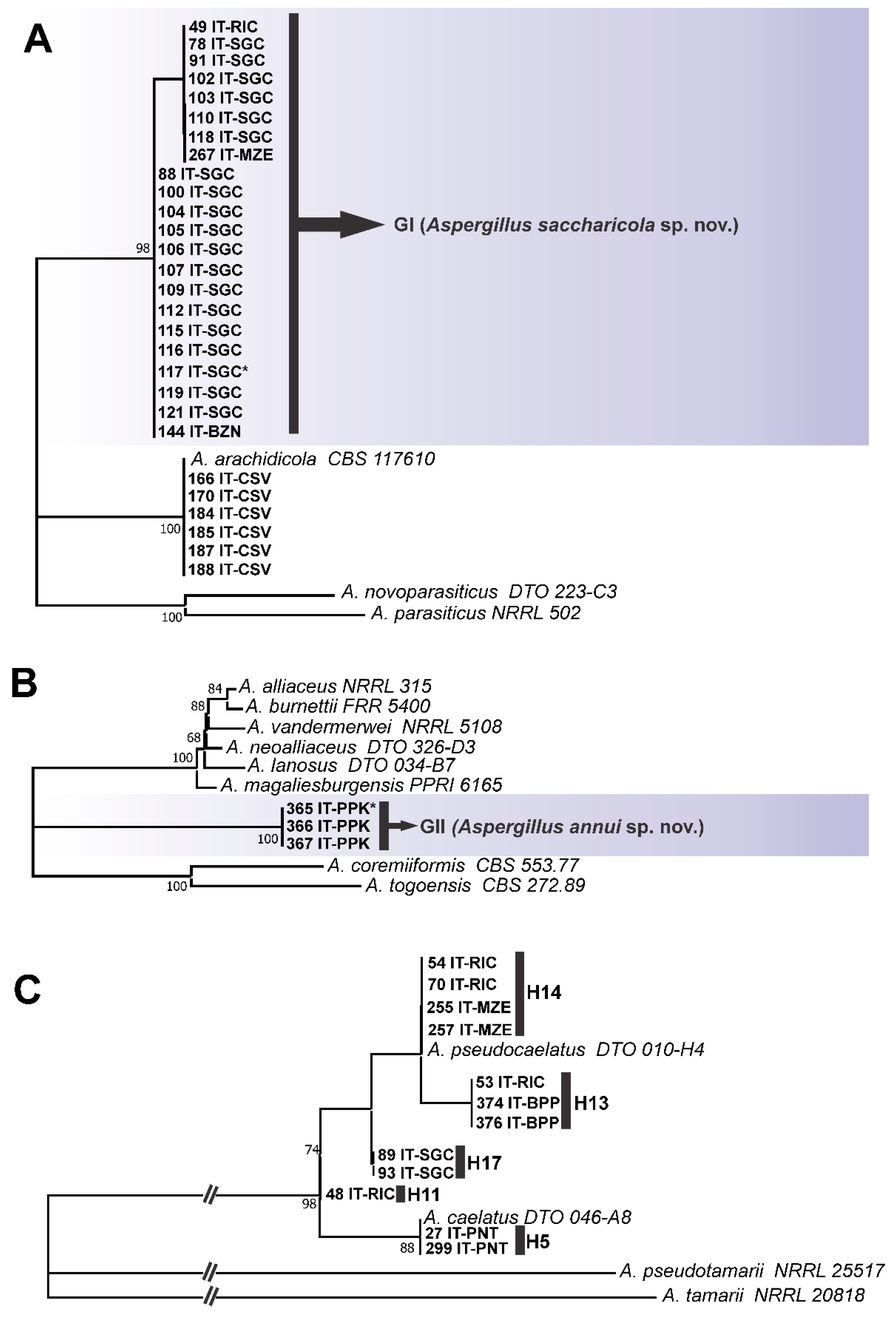
3.1. Genotypic Analysis
3.2. Phenotypic Analysis
3.2.1. Secondary Metabolites Analysis
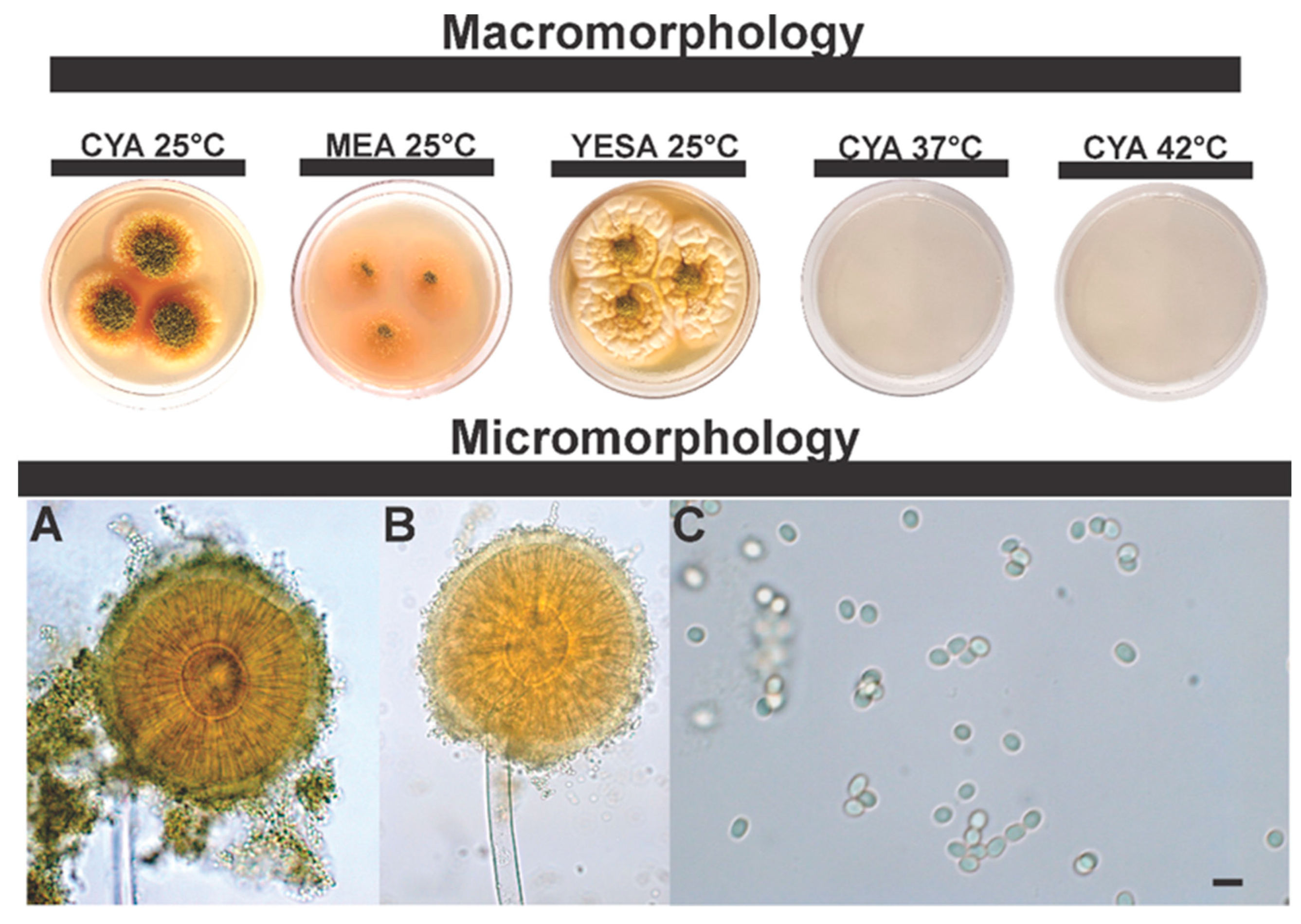
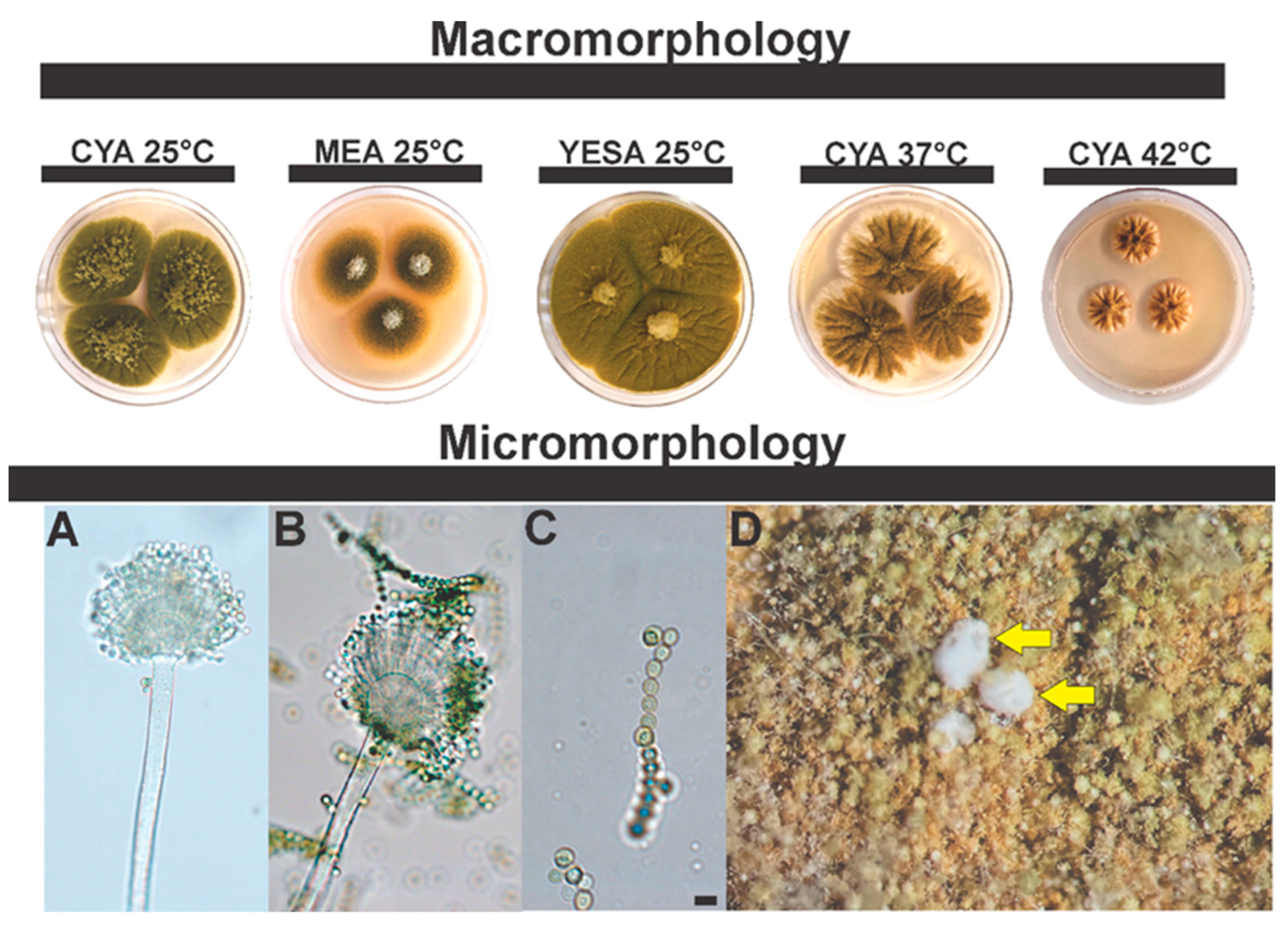
3.2.2. Morphological Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buszewska-Forajta, M. Mycotoxins, invisible danger of feedstuff with toxic effect on animals. Toxicon 2020, 182, 34–53. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.; Yu, J.H. Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed]
- RASFF—The Rapid Alert System for Food and Feed—Annual Report 2020. Available online: https://food.ec.europa.eu/safety/rasff-food-and-feed-safety-alerts_en (accessed on 1 September 2022).
- Marroquín-Cardona, A.G.; Johnson, N.M.; Phillips, T.D.; Hayes, A.W. Mycotoxins in a changing global environment—A review. Food Chem. Toxicol. 2014, 69, 220–230. [Google Scholar] [CrossRef] [PubMed]
- IARC. Monographs on the evaluation of carcinogenic risks to humans: Chemical agents and related occupations. a review of human carcinogens. Int. Agency Res. Cancer 2012, 100, 224–248. [Google Scholar]
- Lewis, L.; Onsongo, M.; Njapau, H.; Schurz-Rogers, H.; Luber, G.; Kieszak, S.; Nyamongo, J.; Backer, L.; Dahiye, A.M.; Misore, A.; et al. Aflatoxin contamination of commercial maize products during an outbreak of acute aflatoxicosis in Eastern and Central Kenya. Environ. Health Perspect. 2005, 113, 1763. [Google Scholar] [CrossRef]
- Mohsenzadeh, M.S.; Hedayati, N.; Riahi-Zanjani, B.; Karimi, G. Immunosuppression following dietary aflatoxin b1 exposure: A review of the existing evidence. Toxin Rev. 2016, 35, 121–127. [Google Scholar] [CrossRef]
- Varga, J.; Baranyi, N.; Muthusamy, C.; Vágvölgyi, C.; Kocsubé, S. Mycotoxin producers in the Aspergillus genus: An update. Acta Biol. Szeged. 2015, 59, 151–167. [Google Scholar]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef]
- Zhai, S.; Zhu, Y.; Feng, P.; Li, M.; Wang, W.; Yang, L.; Yang, Y. Ochratoxin A: Its impact on poultry gut health and microbiota, an overview. Poult. Sci. 2021, 100, 101037. [Google Scholar] [CrossRef]
- Kabak, B. The Fate of Mycotoxins during thermal food processing. J. Sci. Food Agric. 2009, 89, 549–554. [Google Scholar] [CrossRef]
- Hathout, A.S.; Abel-Fattah, S.M.; Abou-Sree, Y.H.; Fouzy, A.S.M. Incidence and exposure assessment of aflatoxins and ochratoxin a in Egyptian wheat. Toxicol. Rep. 2020, 7, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Majeed, S.; Iqbal, M.; Asi, M.R.; Iqbal, S.Z. Aflatoxins and ochratoxin A contamination in rice, corn and corn products from Punjab, Pakistan. J. Cereal Sci. 2013, 58, 446–450. [Google Scholar] [CrossRef]
- Magnoli, C.; Hallak, C.; Astoreca, A.; Ponsone, L.; Chiacchiera, S.; Dalcero, A.M. Occurrence of ochratoxin A-producing fungi in commercial corn kernels in Argentina. Mycopathologia 2006, 161, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Zhihong, L.; Kunlun, H.; Yunbo, L. Ochratoxin A and ochratoxin-producing fungi on cereal grain in china: A review. Food Addit. Contam. Surveill. Part A Surveill 2015, 32, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Zaied, C.; Abid, S.; Bouaziz, C.; Chouchane, S.; Jomaa, M.; Bacha, H. Ochratoxin A levels in spices and dried nuts consumed in Tunisia. Food Addit. Contam. B Surveill. Surveill 2010, 3, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Mateo, R.; Medina, Á.; Mateo, E.M.; Mateo, F.; Jiménez, M. An overview of ochratoxin A in beer and wine. Int. J. Food Microbiol. 2007, 119, 79–83. [Google Scholar] [CrossRef]
- Battilani, P.; Magan, N.; Logrieco, A. European research on ochratoxin A in grapes and wine. Int. J. Food Microbiol. 2006, 111, S2–S4. [Google Scholar] [CrossRef]
- Mondani, L.; Palumbo, R.; Tsitsigiannis, D.; Perdikis, D.; Mazzoni, E.; Battilani, P. Pest Management and ochratoxin A contamination in grapes: A review. Toxins 2020, 12, 303. [Google Scholar] [CrossRef]
- Veras, F.F.; Dachery, B.; Manfroi, V.; Welke, J.E. Colonization of Aspergillus carbonarius and accumulation of ochratoxin A in Vitis Vinifera, Vitis Labrusca, and hybrid grapes—Research on the most promising alternatives for organic viticulture. J. Sci. Food Agric. 2021, 101, 2414–2421. [Google Scholar] [CrossRef]
- Pakshir, K.; Dehghani, A.; Nouraei, H.; Zareshahrabadi, Z.; Zomorodian, K. Evaluation of fungal contamination and ochratoxin A detection in different types of coffee by HPLC-based method. J. Clin. Lab. Anal. 2021, 35, e24001. [Google Scholar] [CrossRef]
- Pires, P.N.; Vargas, E.A.; Gomes, M.B.; Vieira, C.B.M.; Santos, E.A.D.; Bicalho, A.A.C.; Silva, S.D.C.; Rezende, R.P.; Oliveira, I.S.D.; Luz, E.D.M.N.; et al. Aflatoxins and ochratoxin A: Occurrence and contamination levels in cocoa beans from Brazil. Food Addit. Contam. Part A Surveill. 2019, 36, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Kabak, B. Aflatoxins and ochratoxin A in chocolate products in turkey. Food Addit. Contam. Part B Surveill. 2019, 12, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Iha, M.H.; Rodrigues, M.L.; de Cássia Briganti, R. Survey of aflatoxins and ochratoxin A in spices from brazilian market. Braz. Arch. Biol. Technol. 2022, 64, e21210244. [Google Scholar] [CrossRef]
- Freke, J.; Pleadin, J.; Mitak, M.; Lešić, T.; Jakopović, Ž.; Perkovic, I.; Markov, K.; Zadravec, M. Toxicogenic fungi and the occurrence of mycotoxins in traditional meat products. Croat. J. Food Sci. Technol. 2019, 11, 272–281. [Google Scholar] [CrossRef]
- Perši, N.; Pleadin, J.; Kovačević, D.; Scortichini, G.; Milone, S. Ochratoxin A in raw materials and cooked meat products made from ota-treated pigs. Meat Sci. 2014, 96, 203–210. [Google Scholar] [CrossRef]
- Kemboi, D.C.; Antonissen, G.; Ochieng, P.E.; Croubels, S.; Okoth, S.; Kangethe, E.K.; Faas, J.; Lindahl, J.F.; Gathumbi, J.K. A review of the impact of mycotoxins on dairy cattle health: Challenges for food safety and dairy production in Sub-Saharan Africa. Toxins 2020, 12, 222. [Google Scholar] [CrossRef]
- Gumus, R.; Ercan, N.; Imik, H. Determination of ochratoxin A levels in mixed feed and feed stuffs used in some laying hens and ruminant enterprises of Sivas city. Braz. J. Poult. Sci. 2018, 20, 85–90. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef]
- Aragão, A.; Contini, E.O. Agro Brasileiro Alimenta 800 Milhões de Pessoas. Available online: https://www.embrapa.br/busca-de-noticias/-/noticia/59784047/o-agro-brasileiro-alimenta-800-milhoes-de-pessoas-diz-estudo-da-embrapa (accessed on 25 June 2022).
- FAOSTAT. Food and Agriculture Organization of the United Nations—Statistics. 2022. Production-Crops and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 25 March 2022).
- Hong, S.B.; Cho, H.S.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Novel neosartorya species isolated from soil in Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 477–486. [Google Scholar] [CrossRef]
- Peterson, S.W.; Vega, F.E.; Posada, F.; Nagai, C. Penicillium coffeae, a new endophytic species isolated from a coffee plant and its phylogenetic relationship to P. fellutanum, P. thiersii and P. brocae based on parsimony analysis of multilocus DNA sequences. Mycologia 2005, 97, 659–666. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Houbraken, J.; Spierenburg, H.; Frisvad, J.C. Rasamsonia, a new genus comprising thermotolerant and thermophilic Talaromyces and Geosmithia species. Antonie Van Leeuwenhoek 2011, 101, 403–421. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.J.; Iamanaka, B.T.; Ferranti, L.S.; Massi, F.P.; Taniwaki, M.H.; Puel, O.; Lorber, S.; Frisvad, J.C.; Fungaro, M.H.P. Diversity within Aspergillus niger clade and description of a new species: Aspergillus vinaceus sp. nov. J. Fungi 2020, 6, 371. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Mirarab, S.; Warnow, T. ASTRAL-II: Coalescent-Based species tree estimation with many hundreds of taxa and thousands of genes. Bioinformatics 2015, 31, i44–i52. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop-GCE 2010, New Orleans, LA, USA, 14 November 2010. [Google Scholar] [CrossRef]
- Raper, K.B.; Thom, C. A Manual of the Penicillia; Williams & Wilkins Co.: Baltimore, MD, USA, 1949. [Google Scholar]
- Pitt, J.I.; Hocking, A.D.; Glenn, D.R. An improved medium for the detection of Aspergillus flavus and A. parasiticus. J. Appl. Bacteriol. 1983, 54, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.F.; Månsson, M.; Rank, C.; Frisvad, J.C.; Larsen, T.O. Dereplication of microbial natural products by LC-DAD-TOFMS. J. Nat. Prod. 2011, 74, 2338–2348. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Subko, K.; Kildgaard, S.; Frisvad, J.C.; Larsen, T.O. Mass spectrometry-based network analysis reveals new insights into the chemodiversity of 28 species in Aspergillus section Flavi. Front. Fungal Biol. 2021, 2, 719420. [Google Scholar] [CrossRef]
- Kildgaard, S.; Mansson, M.; Dosen, I.; Klitgaard, A.; Frisvad, J.C.; Larsen, T.O.; Nielsen, K.F. Accurate dereplication of bioactive secondary metabolites from marine-derived fungi by UHPLC-DAD-QTOFMS and a MS/HRMS library. Mar. Drugs 2014, 12, 3681–3705. [Google Scholar] [CrossRef]
- Singh, P.; Orbach, M.J.; Cotty, P.J. Aspergillus texensis: A novel aflatoxin producer with s morphology from the United States. Toxins 2018, 10, 513. [Google Scholar] [CrossRef]
- Singh, P.; Callicott, K.A.; Orbach, M.J.; Cotty, P.J. Molecular analysis of s-morphology aflatoxin producers from the United States reveals previously unknown diversity and two new taxa. Front. Microbiol. 2020, 11, 1236. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J. updating the taxonomy of Aspergillus in South Africa. Stud. Mycol. 2020, 95, 253–292. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Lacey, H.J.; Vuong, D.; Pitt, J.I.; Lange, L.; Lacey, E.; Pilgaard, B.; Chooi, Y.H.; Piggott, A.M. Comprehensive chemotaxonomic and genomic profiling of a biosynthetically talented Australian fungus, Aspergillus burnettii sp. nov. Fungal Genet. Biol. 2020, 143, 103435. [Google Scholar] [CrossRef] [PubMed]
- Aime, M.C.; Miller, A.N.; Aoki, T.; Bensch, K.; Cai, L.; Crous, P.W.; Hawksworth, D.L.; Hyde, K.D.; Kirk, P.M.; Lücking, R.; et al. How to publish a new fungal species, or name, version 3.0. IMA Fungus 2021, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, J.R.; Roe, A.D.; Sperling, F.A.H. Multi-locus species delimitation in closely related animals and fungi: One marker is not enough. Mol. Ecol. 2012, 21, 4422–4436. [Google Scholar] [CrossRef]
- Hime, P.M.; Hotaling, S.; Grewelle, R.E.; O’Neill, E.M.; Voss, S.R.; Shaffer, H.B.; Weisrock, D.W. The influence of locus number and information content on species delimitation: An empirical test case in an endangered mexican salamander. Mol. Ecol. 2016, 25, 5959–5974. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Suarez, M.; García, D.; Cano-Lira, J.F.; Guarro, J.; Gené, J. Species diversity in Penicillium and Talaromyces from herbivore dung, and the proposal of two new genera of Penicillium-like fungi in Aspergillaceae. Fungal Syst. Evol. 2020, 5, 39–75. [Google Scholar] [CrossRef] [PubMed]
- Tanney, J.B.; McMullin, D.R.; Green, B.D.; Miller, J.D.; Seifert, K.A. Production of antifungal and antiinsectan metabolites by the Picea endophyte Diaporthe maritima sp. nov. Fungal Biol. 2016, 120, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.W.; Jurjević, Z. Talaromyces columbinus sp. nov., and genealogical concordance analysis in Talaromyces Clade 2a. PLoS ONE 2013, 8, e78084. [Google Scholar] [CrossRef]
- Gutiérrez, E.E.; Garbino, G.S.T. Species delimitation based on diagnosis and monophyly, and its importance for advancing mammalian taxonomy. Zool. Res. 2018, 39, 301–308. [Google Scholar] [CrossRef]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef]
- Dettman, J.R.; Jacobson, D.J.; Taylor, J.W. A Multilocus genealogical approach to phylogenetic species recognition in the model eukaryote Neurospora. Evolution 2003, 57, 2703–2720. [Google Scholar] [CrossRef] [PubMed]
- Rintoul, T.L.; Eggertson, Q.A.; Lévesque, C.A. Multigene phylogenetic analyses to delimit new species in fungal plant pathogens. Methods Mol. Biol. 2012, 835, 549–569. [Google Scholar] [CrossRef]
- Caceres, I.; al Khoury, A.; el Khoury, R.; Lorber, S.; Oswald, I.P.; el Khoury, A.; Atoui, A.; Puel, O.; Bailly, J.D. Aflatoxin biosynthesis and genetic regulation: A review. Toxins 2020, 12, 150. [Google Scholar] [CrossRef]
- Wendel, J.F.; Doyle, J.J. Phylogenetic incongruence: Window into genome history and molecular evolution. In Molecular Systematics of Plants II, 1st ed.; Soltis, D.E., Soltis, P.S., Doyle, J.J., Eds.; Springer: Boston, MA, USA, 1998; pp. 265–296. [Google Scholar] [CrossRef]
- Yan, Z.; Smith, M.L.; Du, P.; Hahn, M.W.; Nakhleh, L. Species tree inference methods intended to deal with incomplete lineage sorting are robust to the presence of paralogs. Syst. Biol. 2022, 71, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Steenkamp, E.T.; Wingfield, M.J.; McTaggart, A.R.; Wingfield, B.D. Fungal species and their boundaries matter—Definitions, mechanisms and practical implications. Fungal Biol. Rev. 2018, 32, 104–116. [Google Scholar] [CrossRef]
- Baum, D.A.; Smith, S.D. Tree Thinking: An introduction to Phylogenetic Biology, 1st ed.; Macmillan Learning: New York, NY, USA, 2012; p. 496. [Google Scholar]
- Maddison, W.P. Gene trees in species trees. Syst. Biol. 1997, 46, 523–536. [Google Scholar] [CrossRef]
- Yu, Y.; Than, C.; Degnan, J.H.; Nakhleh, L. Coalescent Histories on phylogenetic networks and detection of hybridization despite incomplete lineage sorting. Syst. Biol. 2011, 60, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Avise, J.C.; Robinson, T.J. Hemiplasy: A new term in the lexicon of phylogenetics. Syst. Biol. 2008, 57, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, M.; Damm, U.; Crous, P.W.; Cai, L. Species boundaries in plant pathogenic fungi: A Colletotrichum case study. BMC Evol. Biol. 2016, 16, 81. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Pastor, J.L.; Vargas, P.; Pfeil, B.E. Coalescent simulations reveal hybridization and incomplete lineage sorting in Mediterranean Linaria. PLoS ONE 2012, 7, e39089. [Google Scholar] [CrossRef]
- Rosenberg, N.A.; Nordborg, M. Genealogical trees, coalescent theory and the analysis of genetic polymorphisms. Nat. Rev. Genet. 2002, 3, 380–390. [Google Scholar] [CrossRef]
- Liu, L.; Yu, L.; Kubatko, L.; Pearl, D.K.; Edwards, S.V. Coalescent methods for estimating phylogenetic trees. Mol. Phylogenet. Evol. 2009, 53, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Degnan, J.H.; Rosenberg, N.A. Gene tree discordance, phylogenetic inference and the multispecies coalescent. Trends Ecol. Evol. 2009, 24, 332–340. [Google Scholar] [CrossRef]
- Knowles, L.L.; Kubatko, L.S. Estimating Species Trees: Practical and Theoretical Aspects, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; p. 232. [Google Scholar]
- Saag, L.; Mark, K.; Saa, A.; Randlane, T. Species delimitation in the lichenized fungal genus Vulpicida (Parmeliaceae, Ascomycota) using gene concatenation and coalescent-based species tree approaches. Am. J. Bot. 2014, 101, 2169–2182. [Google Scholar] [CrossRef]
- Stukenbrock, E.H.; Banke, S.; Javan-Nikkhah, M.; McDonald, B.A. Origin and domestication of the fungal wheat pathogen Mycosphaerella graminicola via sympatric speciation. Mol. Biol. Evol. 2007, 24, 398–411. [Google Scholar] [CrossRef] [PubMed]
- Looney, B.P.; Adamčík, S.; Matheny, P.B. Coalescent-Based delimitation and species-tree estimations reveal appalachian origin and neogene diversification in Russula subsection Roseinae. Mol. Phylogenet. Evol. 2020, 147, 106787. [Google Scholar] [CrossRef]
- Singh, G.; Dal Grande, F.; Divakar, P.K.; Otte, J.; Leavitt, S.D.; Szczepanska, K.; Crespo, A.; Rico, V.J.; Aptroot, A.; da Silva Cáceres, M.E.; et al. Coalescent-Based species delimitation approach uncovers high cryptic diversity in the cosmopolitan lichen-forming fungal genus Protoparmelia (Lecanorales, Ascomycota). PLoS ONE 2015, 10, e0124625. [Google Scholar] [CrossRef] [PubMed]
- Hubka, V.; Nováková, A.; Jurjević, Ž.; Sklenář, F.; Frisvad, J.C.; Houbraken, J.; Arendrup, M.C.; Jørgensen, K.M.; Siqueira, J.P.Z.; Gené, J.; et al. Polyphasic data support the splitting of Aspergillus candidus into two species; proposal of Aspergillus dobrogensis sp. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 995–1011. [Google Scholar] [CrossRef]
- Geiser, D.M.; Pitt, J.I.; Taylor, J.W. Cryptic speciation and recombination in the aflatoxin-producing fungus Aspergillus flavus. Proc. Natl. Acad. Sci. USA 1998, 95, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Machida, M.; Asai, K.; Sano, M.; Tanaka, T.; Kumagai, T.; Terai, G.; Kusumoto, K.I.; Arima, T.; Akita, O.; Kashiwagi, Y.; et al. Genome sequencing and analysis of Aspergillus oryzae. Nature 2005, 438, 1157–1161. [Google Scholar] [CrossRef]
- Rokas, A.; Payne, G.; Fedorova, N.D.; Baker, S.E.; Machida, M.; Yu, J.; Georgianna, D.R.; Dean, R.A.; Bhatnagar, D.; Cleveland, T.E.; et al. What can comparative genomics tell us about species concepts in the genus Aspergillus? Stud. Mycol. 2007, 59, 11–17. [Google Scholar] [CrossRef]
- Kim, K.M.; Lim, J.; Lee, J.J.; Hurh, B.S.; Lee, I. Characterization of Aspergillus sojae isolated from Meju, Korean traditional fermented soybean brick. J. Microbiol. Biotechnol. 2017, 27, 251–261. [Google Scholar] [CrossRef]
- Hua, S.S.T.; Parfitt, D.E.; Sarreal, S.B.L.; Lee, B.G.; Wood, D.F. First report of an atypical new Aspergillus parasiticus isolates with nucleotide insertion in AflR gene resembling to A. sojae. Mycotoxin Res. 2018, 34, 151–157. [Google Scholar] [CrossRef]
- Li, M.F.; Li, G.H.; Zhang, K.Q. Non-volatile metabolites from Trichoderma spp. Metabolites 2019, 9, 58. [Google Scholar] [CrossRef]
- Yang, S.X.; Wang, H.P.; Gao, J.M.; Zhang, Q.; Laatsch, H.; Kuang, Y. Fusaroside, a unique glycolipid from Fusarium sp., an endophytic fungus isolated from Melia Azedarach. Org. Biomol. Chem. 2012, 10, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Sandjo, L.; Yun, K.; Leutou, A.S.; Kim, G.D.; Choi, H.D.; Kang, J.S.; Hong, J.; Son, B.W. Flavusides A and B, antibacterial cerebrosides from the marine-derived fungus Aspergillus flavus. Chem. Pharm. Bull. 2011, 59, 1174–1177. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Sanada, M. Studies on fluorescent substances produced by Aspergillus fungi (VIII). Purification and isolation of asperopterin B and chemical properties of asperopterin A and B. J. Ferment. Technol. 1969, 47, 8–19. [Google Scholar]
- Matsuura, S.; Yamamoto, M.; Kaneko, Y. The structure of the pteridine glycoside from Aspergillus oryzae. Bull. Chem. Soc. Jpn. 2006, 45, 492–495. [Google Scholar] [CrossRef]
- Pildain, M.B.; Frisvad, J.C.; Vaamonde, G.; Cabral, D.; Varga, J.; Samson, R.A. Two novel aflatoxin-producing Aspergillus species from Argentinean peanuts. Int. J. Syst. Evol. Microbiol. 2008, 58, 725–735. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substitution Model | Alignment Size | Parsimony-Informative Sites | Polymorphic Sites | Nucleotide Diversity | |
|---|---|---|---|---|---|
| CaM | GTR+GI | 474 | 150 | 214 | 0.072 |
| BenA | K80+GI | 499 | 139 | 195 | 0.071 |
| RPB2 | GTR+G | 622 | 179 | 213 | 0.065 |
| Concatenated data | 1595 | 468 | 622 | 0.069 |
| Species | Isolate | Primary Metabolites | Secondary Metabolites |
|---|---|---|---|
| Aspergillus annui | 365 IT-PPK * = IBT 36122 | Glycocholic acid, flavin, pantothenic acid, phytosphingosine, sphinganine | Ergokonin B, kojic acid, nidulanin X3, nidulanin X5 |
| Aspergillus annui | 366 IT-PPK = IBT 36123 | Glycocholic acid, flavin, pantothenic acid, phytosphingosine, sphinganine | Ergokonin B, kojic acid, nidulanin X3, nidulanin X5 |
| Aspergillus annui | 367 IT-PPK = IBT 36124 | Glycocholic acid, flavin, pantothenic acid, phytosphingosine, sphinganine | Ergokonin B, kojic acid, nidulanin X3, nidulanin X5 |
| Aspergillus saccharicola | 78 IT-SGC = IBT 36125 | Phytosphingosine | Aflatoxin B1, B2, G1, G2, M1, anthranilic acid, aspergillic acid, chrysogine, cyclopiazonic acid, desertorin A, ergokonine B, erythroglaucin, kojic acid, O-methylsterigmatocystin, nidulanin X6, parasiticolide A, sterigmatocystin, tenuazonic acid, valine-tenuazonic acid |
| Aspergillus saccharicola | 117 IT-SGC * = IBT 36126 | Phytosphingosine | Aflatoxin B1, B2, G1, G2, M1, anthranilic acid, aspergillic acid, chrysogine, cyclopiazonic acid, desertorin A, ergokonine B, erythroglaucin, kojic acid, O-methylsterigmatocystin, nidulanin X6, parasiticolide A, sterigmatocystin, tenuazonic acid, valine-tenuazonic acid |
| Aspergillus saccharicola | 121 IT-SGC = IBT 36127 | Phytosphingosine | Aflatoxin B1, B2, G1, G2, M1, anthranilic acid, aspergillic acid, chrysogine, cyclopiazonic acid, desertorin A, ergokonine B, erythroglaucin, kojic acid, O-methylsterigmatocystin, nidulanin X6, parasiticolide A, sterigmatocystin, tenuazonic acid, valine-tenuazonic acid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, J.J.; Fungaro, M.H.P.; Wang, X.; Larsen, T.O.; Frisvad, J.C.; Taniwaki, M.H.; Iamanaka, B.T. Deep Genotypic Species Delimitation of Aspergillus Section Flavi Isolated from Brazilian Foodstuffs and the Description of Aspergillus annui sp. nov. and Aspergillus saccharicola sp. nov. J. Fungi 2022, 8, 1279. https://doi.org/10.3390/jof8121279
Silva JJ, Fungaro MHP, Wang X, Larsen TO, Frisvad JC, Taniwaki MH, Iamanaka BT. Deep Genotypic Species Delimitation of Aspergillus Section Flavi Isolated from Brazilian Foodstuffs and the Description of Aspergillus annui sp. nov. and Aspergillus saccharicola sp. nov. Journal of Fungi. 2022; 8(12):1279. https://doi.org/10.3390/jof8121279
Chicago/Turabian StyleSilva, Josué J., Maria H. P. Fungaro, Xinhui Wang, Thomas O. Larsen, Jens C. Frisvad, Marta H. Taniwaki, and Beatriz T. Iamanaka. 2022. "Deep Genotypic Species Delimitation of Aspergillus Section Flavi Isolated from Brazilian Foodstuffs and the Description of Aspergillus annui sp. nov. and Aspergillus saccharicola sp. nov." Journal of Fungi 8, no. 12: 1279. https://doi.org/10.3390/jof8121279
APA StyleSilva, J. J., Fungaro, M. H. P., Wang, X., Larsen, T. O., Frisvad, J. C., Taniwaki, M. H., & Iamanaka, B. T. (2022). Deep Genotypic Species Delimitation of Aspergillus Section Flavi Isolated from Brazilian Foodstuffs and the Description of Aspergillus annui sp. nov. and Aspergillus saccharicola sp. nov. Journal of Fungi, 8(12), 1279. https://doi.org/10.3390/jof8121279