
Experimental Outcrossing in Agaricus bisporus Revealed a Major and Unexpected Involvement of Airborne Mycelium Fragments

Abstract
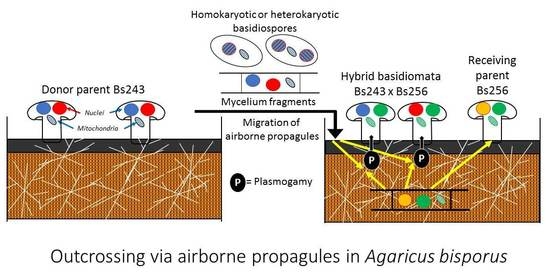
1. Introduction
2. Materials and Methods
2.1. Strains and Grain Spawn Preparation
2.2. Experimental Outcrossing
2.3. DNA Extraction, PCR Amplification and Sequencing
2.4. Nuclear Cleaved Amplified Polymorphic Sequence (CAPS) and Nuclear Haplotype Markers
2.5. Mitochondrial Marker
3. Results
3.1. Characterization of the Wild Heterokaryons Bs177, Bs243 and Bs256
3.1.1. Different Cap Colors Observed in the Three Parental Strains
3.1.2. Detection of 29 SNPs and 24 Heteromorphisms in the ITS DNA Region and the Fruk Gene
3.1.3. Development of CAPS Nuclear Markers and Genotypes of the Parent Strains
3.1.4. Development of Haplotype Nuclear Markers and Haplotypes Found in the Parent Strains
3.1.5. Haplotype Mitochondrial Marker with Different Lengths in the Three Parental Strains
3.2. Characterization of Basidiomata Collected on the Receiver Bs177
3.3. Characterization of Basidiomata Collected on the Receiver Bs256
3.3.1. Cap Color, Mitochondrial Haplotypes and CAPS Nuclear Genotypes of 32 Basidiomata Collected on Bs256
3.3.2. Haplotypes of Basidiomata Collected on Bs256 Reveals Many Hybrids but with Only Two Genotypes in Agreement with the Involvement of Airborne Mycelium Fragments
3.3.3. Different Results Confirming the Non-Involvement of the Spores in the Outcrossing
4. Discussion
4.1. Conditions for Outcrossing and Consequences on Genetic Diversity in Collection Sites
4.2. Genotypes and Cap Color of the Hybrids
4.3. How Did Outcrossing Occur?
4.4. Success of Airborne Mycelium Fragments
4.5. Possible Undetected Recombination and Hidden Outcrossing
4.6. A method for Strain Hybridization
4.7. Biotrophic Contribution of the Resident Mycelium
4.8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heneghan, M.N.; Burns, C.; Costa, A.M.S.B.; Burton, K.S.; Challen, M.P.; Bailey, A.M.; Foster, G.D. Functional analysis of Agaricus bisporus serine proteinase 1 reveals roles in utilization of humic rich substrates and adaptation to the leaf-litter ecological niche. Environ. Microbiol. 2016, 18, 4687–4696. [Google Scholar] [CrossRef]
- Callac, P.; Theochari, I.; Kerrigan, R.W. The germplasm of Agaricus bisporus: Main results after ten years of collecting in France, in Greece, and in North America. Acta Hortic. 2002, 579, 49–55. [Google Scholar] [CrossRef]
- Savoie, J.-M.; Foulongne-Oriol, M.; Barroso, G.; Callac, P. 1 Genetics and genomics of cultivated mushrooms, application to breeding of Agarics. In The Mycota; Agricultural Application; Kempken, F., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 11, pp. 3–33. [Google Scholar] [CrossRef]
- Callac, P.; Jacobe de Haut, I.; Imbernon, M.; Guinberteau, J.; Theochari, I. A novel homothallic variety of Agaricus bisporus comprises rare tetrasporic isolates from Europe. Mycologia 2003, 95, 222–231. [Google Scholar] [CrossRef]
- Callac, P.; Billette, C.; Imbernon, M.; Kerrigan, R.W. Morphological, genetic, and interfertility analyses reveal a novel, tetrasporic variety of Agaricus bisporus from the Sonoran Desert of California. Mycologia 1993, 85, 835–851. [Google Scholar] [CrossRef]
- Xu, J.; Kerrigan, R.W.; Horgen, P.A.; Anderson, J.B. Localization of the mating type gene in Agaricus bisporus. Appl. Environ. Microbiol. 1993, 59, 3044–3049. [Google Scholar] [CrossRef]
- Imbernon, M.; Callac, P.; Granit, S.; Pirobe, L. Allelic polymorphism at the mating type locus in Agaricus bisporus var. burnettii, and confirmation of the dominance of its tetrasporic trait. In Proceedings of the 14th International Congress on the Science and Cultivation of Edible Fungi, Oxford, UK, 17–22 September 1995; Elliott, T.J., Ed.; Balkema: Rotterdam, The Netherlands, 1995; Volume 1, pp. 11–19. Available online: https://hal.archives-ouvertes.fr/hal-03817251 (accessed on 6 November 2022).
- Foulongne-Oriol, M.; Taskent, O.; Kües, U.; Sonnenberg, A.S.M.; Van Peer, A.F.; Giraud, T. Mating-type locus organization and mating-type chromosome differentiation in the bipolar edible button mushroom Agaricus bisporus. Genes 2021, 12, 1079. [Google Scholar] [CrossRef]
- Raper, C.A.; Raper, J.R.; Miller, R.E. Genetic analysis of the life cycle of Agaricus bisporus. Mycologia 1972, 64, 1088–1117. [Google Scholar] [CrossRef]
- Sonnenberg, A.S.; Gao, W.; Lavrijssen, B.; Hendrickx, P.; Sedaghat-Tellgerd, N.; Foulongne-Oriol, M.; Kong, W.S.; Schijlen, E.G.; Baars, J.J.; Visser, R.G. A detailed analysis of the recombination landscape of the button mushroom Agaricus bisporus var. bisporus. Fungal Genet. Biol. 2016, 93, 35–45. [Google Scholar] [CrossRef]
- Xu, J.; Kerrigan, R.W.; Callac, P.; Horgen, P.A.; Anderson, J.B. Genetic structure of natural populations of Agaricus bisporus, the commercial button mushroom. J. Hered. 1997, 88, 482–488. [Google Scholar] [CrossRef]
- Xu, J.; Desmerger, C.; Callac, P. Fine-scale genetic analyses reveal unexpected spatial-temporal heterogeneity in two natural populations of the commercial mushroom Agaricus bisporus. Microbiology 2002, 148, 1253–1262. [Google Scholar] [CrossRef]
- Kerrigan, R.W.; Horgen, P.A.; Anderson, J.B. The California population of Agaricus bisporus comprises at least two ancestral elements. Syst. Bot. 1993, 18, 123–136. [Google Scholar] [CrossRef]
- Kerrigan, R.W. Evidence of genetic divergence in two populations of Agaricus bisporus. Mycol. Res. 1990, 94, 721–733. [Google Scholar] [CrossRef]
- Callac, P.; Imbernon, M.; Kerrigan, R.W.; Olivier, J.M. The two life cycles of Agaricus bisporus. Mushroom biology and mushroom product. In Proceedings of the Second International Conference on Mushroom Biology and Mushroom Products, University Park, PA, USA, 9–12 June 1996; Royse, E.D., Ed.; pp. 57–66. Available online: https://hal.archives-ouvertes.fr/hal-03817267 (accessed on 6 November 2022).
- Buller, A.H.R. The diploid cell and the diploidisation process in plants and animals, with special reference to the higher fungi. Bot. Rev. 1941, 7, 389–431. Available online: https://www.jstor.org/stable/4353254 (accessed on 6 November 2022). [CrossRef]
- Xu, J.; Horgen, P.A.; Anderson, J.B. Somatic recombination in the cultivated mushroom Agaricus bisporus. Mycol. Res. 1996, 100, 188–192. [Google Scholar] [CrossRef]
- Callac, P.; Gaubert, J.; Imbernon, M.; Guinberteau, J.; Desmerger, C.; Olivier, J.-M. Ressources génétiques chez les agarics: Résultats récents et premiers essais expérimentaux d’interfécondation libre chez le champignon de Paris. In Les Actes du BRG; Bureau des Ressources Génétiques: Paris, France, 2003; Volume 4, pp. 331–346. [Google Scholar]
- Callac, P.; Spataro, C.; Caille, A.; Imbernon, M. Evidence for outcrossing via the Buller phenomenon in a substrate simultaneously inoculated with spores and mycelium of Agaricus bisporus. Appl. Environ. Microbiol. 2006, 72, 2366–2372. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Callac, P.; Imbernon, M.; Savoie, J.-M. Outcrossing via the Buller phenomenon in a substrate simultaneously inoculated with spores and mycelium of Agaricus bisporus creates variability for agronomic traits. In Proceedings of the 6th International Conference on Mushroom Biology and Mushroom Products, Bonn, Germany, 29 September–3 October 2008; Lelley, J.I., Buswell, J.A., Eds.; pp. 113–119. Available online: https://hal.archives-ouvertes.fr/hal-03696361 (accessed on 6 November 2022).
- Xu, J.; Kerrigan, R.W.; Sonnenberg, A.S.; Callac, P.; Horgen, P.A.; Anderson, J.B. Mitochondrial DNA variation in natural populations of the mushroom Agaricus bisporus. Mol. Ecol. 1998, 7, 19–33. [Google Scholar] [CrossRef]
- Barroso, G.; Blesa, S.; Labarère, J. Wide distribution of mitochondrial genome rearrangements in wild strains of the basidiomycete Agrocybe aegerita. Appl. Environ. Microbiol. 1995, 61, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Imbernon, M.; Callac, P.; Gasqui, P.; Kerrigan, R.W.; Velcko, A.J., Jr. BSN, the primary determinant of basidial spore number and reproductive mode in Agaricus bisporus, maps to chromosome I. Mycologia 1996, 88, 749–761. [Google Scholar] [CrossRef]
- Jalalzadeh, B.; Sare Carras, I.; Ferandon, C.; Callac, P.; Farsi, M.; Savoie, J.-M.; Barroso, G. The intraspecific variability of mitochondrial genes of Agaricus bisporus reveals an extensive group I intron mobility combined with low nucleotide substitution rates. Curr. Genet. 2015, 61, 87–102. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignments through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Bullock, W.O.; Fernandez, J.M.; Short, J.M. XL1-Blue—A high-efficiency plasmid transforming recA Escherichia coli strain with β-galactosidase selection. Biotechniques 1987, 5, 376–379. [Google Scholar]
- Hanahan, D. Studies on the transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A.S.M.; Baars, J.J.P.; Hendrikx, P.M.; Lavrijssen, B.; Gao, W.; Weijn, A.; Mes, J.J. Breading and strain protection in the button mushroom Agaricus bisporus. In Proceedings of the 7th International Conference on Mushroom Biology and Mushroom Product, Arcachon, France, 4–7 October 2011; Savoie, J.M., Foulongne-Oriol, M., Largeteau, M., Barroso, G., Eds.; Volume 1, pp. 7–15. Available online: https://library.wur.nl/WebQuery/wurpubs/fulltext/217696 (accessed on 6 November 2022).
- Sonnenberg, A.S.; de Groot, P.W.; Schaap, P.J.; Baars, J.J.; Visser, J.; Van Griensven, L.J. Isolation of expressed sequence tags of Agaricus bisporus and their assignment to chromosomes. Appl. Environ. Microbiol. 1996, 62, 4542–4547. [Google Scholar] [CrossRef] [PubMed]
- Callac, P.; Moquet, F.; Imbernon, M.; Ramos Guedes-Lafargue, M.; Mamoun, M.; Olivier, J.-M. Evidence for PPC1, a determinant of the pilei-pellis color of Agaricus bisporus fruit bodies. Fungal Genet. Biol. 1998, 23, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Foulongne-Oriol, M.; Rodier, A.; Rousseau, T.; Savoie, J.-M. Quantitative trait locus mapping of yield-related components and oligogenic control of the cap color of the button mushroom, Agaricus bisporus. Appl. Environ. Microbiol. 2012, 78, 2422–2434. [Google Scholar] [CrossRef]
- Lösel, D.M. The stimulation of spore germination in Agaricus bisporus by living mycelium. Ann. Bot. 1964, 28, 541–554. [Google Scholar] [CrossRef]
- Thongklang, N.; Hoang, E.; Estrada, A.E.R.; Sysouphanthong, P.; Moinard, M.; Hyde, D.K.; Kerrigan, R.W.; Foulongne-Oriol, M.; Callac, P. Evidence for amphithallism and broad geographical hybridization potential among Agaricus subrufescens isolates from Brazil, France and Thailand. Fungal Biol. 2014, 118, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Largeteau, M.; Callac, P.; Navarro-Rodriguez, A.M.; Savoie, J.-M. Diversity in the ability of Agaricus bisporus wild isolates to fruit at high temperature (25 °C). Fungal Biol. 2011, 115, 1186–1195. [Google Scholar] [CrossRef]
- Dickhardt, R. Homokaryotization of Agaricus bitorquis (Quel.) Sacc. and Agaricus bisporus (Lange) Imb. Theor. Appl. Genet. 1985, 70, 52–56. [Google Scholar] [CrossRef]
- O’Connor, E.; Doyle, S.; Amini, A.; Grogan, H.; Fitzpatrick, D.A. Transmission of mushroom virus X and the impact of virus infection on the transcriptomes and proteomes of different strains of Agaricus bisporus. Fungal Biol. 2021, 125, 704–717. [Google Scholar] [CrossRef]
- Kerrigan, R.W.; Callac, P.; Xu, J.; Noble, R. Population and phylogenetic structure within the Agaricus subfloccosus complex. Mycol. Res. 1999, 103, 1515–1523. [Google Scholar] [CrossRef]
- Zhou, J.L.; Xu, J.; Jiao, A.-G.; Yang, L.; Chen, J.; Callac, P.; Liu, Y.; Wang, S. Patterns of PCR amplification artifacts of the fungal barcode marker in a hybrid mushroom. Front. Microbiol. 2019, 10, 2686. [Google Scholar] [CrossRef] [PubMed]
- Callac, P.; Kabalan, R.; Férandon, C.; Barroso, G. Mitochondrial inheritance during outcrossing in substrate simultaneously inoculated with spores and mycelium of Agaricus bisporus. In Proceedings of the 9th European Conference on Fungal Genetics, Edinbrugh, UK, 5–8 April 2008; Available online: https://hal.archives-ouvertes.fr/hal-03691349 (accessed on 6 November 2022).
- Kamzolkina, O.; Volkova, V.; Kozlova, M.; Pancheva, E.; Dyakov, Y.; Callac, P. Karyological evidence for meiosis in the three different types of life cycles existing in Agaricus bisporus. Mycologia 2006, 98, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A.S.M.; Baars, J.J.P.; Gao, W.; Visser, R.G.F. Developments in breeding of Agaricus bisporus var. bisporus: Progress made and technical and legal hurdles to take. Appl. Microbiol. Biotechnol. 2017, 101, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, P. Biology and ecology of mycoparasitism. Can. J. Bot. 1995, 73, 1264–1300. [Google Scholar] [CrossRef]
- Sinden, J.W.; Tschierpe, H.J.; Hauser, E. Transplantation of sporophores as a new method for studying growth and nutritional factors of mushrooms. In Proceedings of the 5th International Congress on the Science and Cultivation of Edible Fungi, Philadelphia, PA, USA, 1962; pp. 250–266. Available online: https://www.isms.biz/ (accessed on 6 November 2022).
- Woolston, B.M.; Schlagnhaufer, C.; Wilkinson, J.; Larsen, J.; Shi, Z.; Mayer, K.; Walters, D.S.; Curtis, W.R.; Romaine, C.P. Long-distance translocation of protein during morphogenesis of the fruiting body in the filamentous fungus, Agaricus bisporus. PLoS ONE 2011, 6, e28412. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polymorphic Positions | Genotyping Based on ITS Sequences Obtained after PCR Cloning 1 | Genotyping at its:541 CAPS Marker after HaeIII Digestion 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ITS Haplotypes of the Nuclei Based on PCR Clones | Genotypes of the Strains | Genotypes of the Strains | |||||||||
| its-1 Bs177-Clone1 OP518685 | its-2 Bs177-Clone2 OP518686 | its-3 Bs243-Clone1 OP518687 | its-4 Bs243-Clone2 OP518688 | its-5 Bs256 OP520895 | its-1/2 Bs177 KF848690 | its-3/4 Bs243 KF848693 | its-5/5 Bs256 KF848696 | its:541-2/2 Bs177 | its:541-1/2 Bs243 | its:541-1/1 Bs256 | |
| 51 | G | A | A | A | A | A/G | A/A | A/A | |||
| 169 | T | C | C | T | C | C/T | C/T | C/C | |||
| 172 | C | T | T | C | T | C/T | C/T | T/T | |||
| 503 | G | A | G | G | G | A/G | G/G | G/G | |||
| 541 | C | C | T | C | T | C/C | C/T | T/T | 205, 244, 317 (C/C) | 205, 244, 317, 522 (C/T) | 244, 522 (T/T) |
| 547 | C | A | C | C | C | A/C | C/C | C/C | |||
| 582 | C | C | C | C | T | C/C | C/C | T/T | |||
| Polymorphic Position | Genotyping Based on fruk Sequences Obtained after PCR Cloning 1 | Genotyping at fruk:655 CAPS Marker after HaeIII Digestion 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| fruk Haplotypes of Parental Nuclei | Genotype of the Strains | Genotypes of the Strains | |||||||||
| fruk-1 Bs177 OP555287 | fruk-2 Bs243-clone1 OP555288 | fruk-3 Bs243-clone2 OP555289 | fruk-4 Bs256-clone1 OP55529 | fruk-5 Bs256-clone2 OP555291 | fruk-1/1 Bs177 KF848701 | fruk-2/3 Bs243 KF848704 | fruk-4/5 Bs256 KF848707 | fruk:655-1/1 Bs177 | fruk:655-1/2 Bs243 | fruk:655-1/2 Bs256 | |
| 144 | C | C | T | C | C | C/C | C/T | C/C | |||
| 242 | C | G | C | G | G | C/C | G/C | G/G | |||
| 247 | G | A | A | A | A | G/G | A/A | A/A | |||
| 279 | C | T | T | T | T | C/C | T/T | T/T | |||
| 289 | A | A | A | G | A | A/A | A/A | A/G | |||
| 435 | C | C | T | T | C | C/C | C/T | C/T | |||
| 463 | C | T | T | T | T | C/C | T/T | T/T | |||
| 490 | C | T | T | T | T | C/C | T/T | T/T | |||
| 640 | A | G | G | G | G | A/A | G/G | G/G | |||
| 655 | T | T | C | C | T | T/T | C/T | C/T | 879 (T/T) | 205, 674, 879 (C/T) | 205, 674, 879 (C/T) |
| 671 | C | T | T | T | T | C/C | T/T | T/T | |||
| 674 | A | G | G | G | G | A/A | G/G | G/G | |||
| 676 | C | C | A | C | C | C/C | C/A | C/C | |||
| 677 | G | G | A | G | G | G/G | A/G | G/G | |||
| 683 | G | A | G | G | A | G/G | A/G | A/G | |||
| 710 | G | G | G | A | A | G/G | G/G | A/A | |||
| 730 | C | G | G | C | C | C/C | G/G | C/C | |||
| 752 | C | C | T | C | T | C/C | C/T | C/T | |||
| 788 | G | A | A | G | A | G/G | A/A | A/G | |||
| 809 | A | G | G | G | G | A/A | G/G | G/G | |||
| 815 | G | G | G | A | G | G/G | G/G | A/G | |||
| 823 | T | C | C | C | T | T/T | C/C | C/T | |||
| Material | Ploidy Level | Expected and Observed its and fruk Genotypes at CAPS Markers in Mycelia and Spores | Mitochondrial Haplotype |
|---|---|---|---|
| Donor mycelium Bs243 | Heterokaryotic mycelium | its:541-1 fruk:655-1 + its:541-2 fruk:655-2 or its:541-1 fruk:655-2 + its:541-2 fruk:655-1 | iAbi11-0 |
| Basidiospores of Bs243 (possible genotypes) | Homokaryotic spores | its:541-1 fruk:655-1 | iAbi11-0 |
| its:541-1 fruk:655-2 | iAbi11-0 | ||
| its:541-2 fruk:655-1 | iAbi11-0 | ||
| its:541-2 fruk:655-2 | iAbi11-0 | ||
| Heterokaryotic spores 1 | its:541-1 fruk:655-1 + its:541-2 fruk:655-2 | iAbi11-0 | |
| its:541-1 fruk:655-2 + its:541-2 fruk:655-1 | iAbi11-0 | ||
| its:541-1 fruk:655-1 + its:541-1 fruk:655-2 infrequent 2 | iAbi11-0 | ||
| its:541-2 fruk:655-2 + its:541-2 fruk:655-1 infrequent 2 | iAbi11-0 | ||
| Receiver mycelium Bs256 | Heterokaryotic mycelium | its:541-1 fruk:655-2 + its:541-1 fruk:655-1 (medium brown cap) | iAbi11-L |
| Basidiomata collected on Bs256 (32) | Heterokaryon of unknown type (14) | its:541-1 fruk:655-2 + its:541-1 fruk:655-1 (medium brown cap) 3 | iAbi11-L |
| Heterokaryon of hybrid type (18) | its:541-1 fruk:655-2 + its:541-2 fruk:655-2 (cream cap) | iAbi11-L |
| Material | Ploidy Level | Expected and Observed its and fruk Nuclear Haplotypes in Mycelia and Spores | Mitochondrial Haplotype |
|---|---|---|---|
| Donor mycelium Bs243 | Heterokaryotic mycelium | its-3 fruk-2 + its-4 fruk-3 or its-3 fruk-3 + its-4 fruk-2 | iAbi11-0 |
| Basidiospores of Bs243 (possible genotypes) | Homokaryotic spores | its-3 fruk-2 | iAbi11-0 |
| its-4 fruk-3 | iAbi11-0 | ||
| its-3 fruk-3 | iAbi11-0 | ||
| its-4 fruk-2 | iAbi11-0 | ||
| Heterokaryotic spores 1 | its-3 fruk-2 + its-4 fruk-3 | iAbi11-0 | |
| its-4 fruk-2 + its-3 fruk-3 | iAbi11-0 | ||
| (its-3 fruk-2 + its-3 fruk-3) infrequent 2 | iAbi11-0 | ||
| (its-4 fruk-2 + its-4 fruk-3) infrequent 2 | iAbi11-0 | ||
| Receiver mycelium Bs256 | Heterokaryotic mycelium | its-5 fruk-4 + its-5 fruk-5 | iAbi11-L |
| Basidiomata collected on receiver trays (12) | Heterokaryon of parental type (2) | its-5 fruk-4 + its-5 fruk-5 (2 with medium brown cap) | iAbi11-L |
| Heterokaryon of hybrid type (10) | its-5 fruk-4 + its-3 fruk-2 (4 with medium brown cap) example: hybrid Bs256xBs243-H17 its 3/5 GB 3: OP704222, fruk 2/4 GB: OP750462 | iAbi11-L | |
| its-5 fruk-4 + its-4 fruk-3 (6 with cream cap) example: hybrid Bs256xBs243-H10 its 4/5 GB 3: OP704221, fruk 3/4 GB: OP750461 | iAbi11-L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jalalzadeh, B.; Barroso, G.; Savoie, J.-M.; Callac, P. Experimental Outcrossing in Agaricus bisporus Revealed a Major and Unexpected Involvement of Airborne Mycelium Fragments. J. Fungi 2022, 8, 1278. https://doi.org/10.3390/jof8121278
Jalalzadeh B, Barroso G, Savoie J-M, Callac P. Experimental Outcrossing in Agaricus bisporus Revealed a Major and Unexpected Involvement of Airborne Mycelium Fragments. Journal of Fungi. 2022; 8(12):1278. https://doi.org/10.3390/jof8121278
Chicago/Turabian StyleJalalzadeh, Banafsheh, Gérard Barroso, Jean-Michel Savoie, and Philippe Callac. 2022. "Experimental Outcrossing in Agaricus bisporus Revealed a Major and Unexpected Involvement of Airborne Mycelium Fragments" Journal of Fungi 8, no. 12: 1278. https://doi.org/10.3390/jof8121278
APA StyleJalalzadeh, B., Barroso, G., Savoie, J.-M., & Callac, P. (2022). Experimental Outcrossing in Agaricus bisporus Revealed a Major and Unexpected Involvement of Airborne Mycelium Fragments. Journal of Fungi, 8(12), 1278. https://doi.org/10.3390/jof8121278