On the Origins of a Species: What Might Explain the Rise of Candida auris?


Abstract
1. Introduction
2. First Detection and Known Global Distribution
3. Genomic Epidemiology
4. Distinctive Biological Properties of C. auris
5. Taxonomy and Phylogeny of C. auris and Close Relatives
- First described in 1961 as Torulopsis haemulonii after having been isolated from the gut of the blue-striped grunt fish (Haemulon sciurus) near the U.S. Florida coast [41].
- Other marine settings:
- ∘
- ∘
- Soft coral (Palythoa variabilis) in Brazil in 2016 [43]
- ∘
- Skin of a dolphin (species not reported) near Suriname [44]
- ∘
- Second most common yeast from pool water of captive bottlenose dolphins (Tursiops truncates) in the U.S. state of Connecticut [45]
- ∘
- Indian researchers applied C. haemulonii to giant tiger shrimp (Penaeus monodon)—widely farmed in aquaculture—finding that its presence boosted an immunostimulatory molecule in the shrimp, conferring protection against a viral infection that causes economic losses from shrimp die-off [42]. It is unclear whether C. haemulonii has become used more broadly in aquaculture since publication of the study in 2011. Further investigation of its use and of whether the isolate used was truly C. haemulonii would be of great interest.
- Terrestrial sources:
- It is important to note that reports of C. haemulonii without use of ribosomal DNA sequencing, and before naming of C. auris in 2009, must be examined with caution, since misclassification as other yeasts is possible.
- Isolated from sap from a rubber tree (Hevea brasiliensis) in Yunnan Province, China, in 2008 [56].
- Isolated from Ruellia sp. flowers in India in 2008 [49].
- Isolated from blood of a patient who died of aspiration pneumonia in Malaysia [57].
- Twice identified from human sources (CDC, unpublished data)
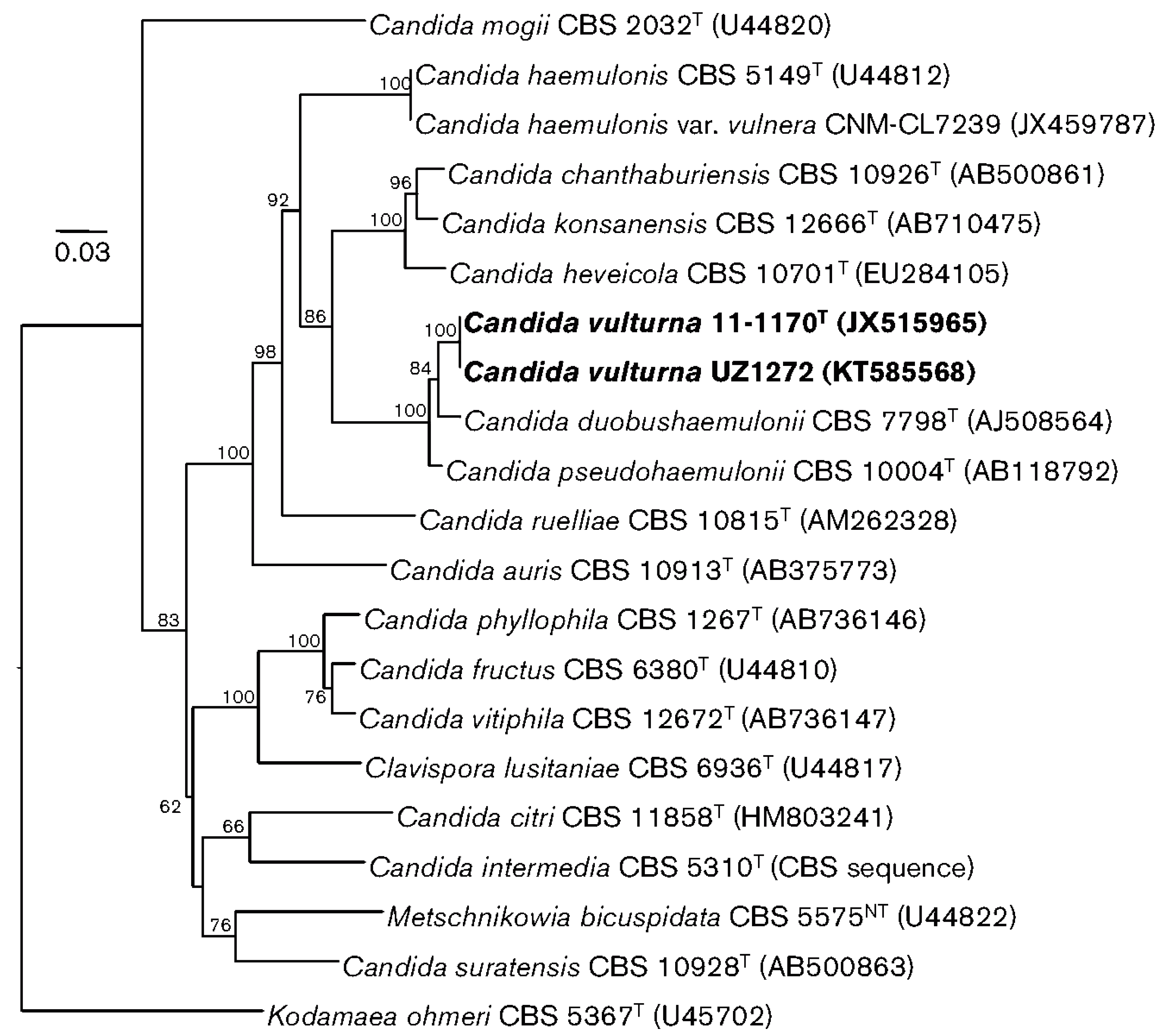
- Isolated from an unspecified flower collected on the outskirts of the city of Cagayan de Oro, Philippines, in 2016 [57]. The authors report, “as most of the other related species were originally described from plants, we presume that the natural habitat of C. vulturna is associated with plants but, occasionally, it can also infect humans” [57]. This report contains the most comprehensive phylogenic tree published to date about the species most closely related to C. auris by ribosomal DNA sequencing (Figure 1).
- Isolated from bark of tall-stilt mangrove (Rhizophora apiculata) in Thailand in 2004. The authors report that a wide variety of yeasts can be found in mangrove forests, which provide large quantities of organic matter [58].
- Isolated from bluegrape jasmine (Jasminum adenophyllum) in Thailand [59].
6. Why Emerge Now?
6.1. Role of Healthcare
- The ability of C. auris to persistently and asymptomatically colonize human skin.
- The ability of C. auris to persistently contaminate indoor surfaces, combined with tolerance of some commonly used healthcare disinfectants.
- Concentrations of people with decreased immunity, invasive devices, and disrupted microbiomes, including from broad-spectrum antibacterial medications, antifungal medications, and underlying illness. Differences in bathing and skin care practices between the general population and those in long-term medical care, including the use of antiseptics, might also affect the skin microbiome.
6.2. Possible Changes in the Organism
6.3. New Human Activities—Increased Contact
6.4. New Human Activities—C. auris Amplification
6.5. Global Temperature Changes
7. Solving the Mystery Surrounding the Origins and Evolutionary Reservoirs of C. auris
8. Response to C. auris
Author Contributions
Funding
Conflicts of Interest
Disclaimer
Appendix A

{kind=link}
| Candida species 1 | Human | Animal | Plant | Aquatic Environment |
|---|---|---|---|---|
| Candida haemulonii | Numerous isolates from invasive and non-invasive sites 2 | Haemulon scirus (blue-striped grunt) off coast of Florida, USA; Laboratory tick colony in Czech Republic; Soft coral in Brazil; Applied to farmed shrimp in India | Cassava root in Brazil | Seawater near Portugal and India; pool water of captive bottlenose dolphins |
| Candida duobushaemulonii | Blood and foot ulcers in Spain and USA; vaginal samples in Brazil | Pyrrhocoris apterus (firebug) in Germany | Not reported | Not reported |
| Candida pseudohaemulonii | Blood in Thailand and South Korea; tentative identification from nail in Argentina | Not reported | Not reported | Not reported |
| Candida heveicola | Not reported | Not reported | Hevea brasiliensis (rubber tree) sap in China | Not reported |
| Candida ruelliae | Not reported | Not reported | Ruellia sp. flowers in India | Not reported |
| Candida vulturna | Blood in Malaysia; two human sources in USA | Not reported | Flower in Philippines | Not reported |
| Candida chanthaburiensis | Not reported | Not reported | Bark of Rhizophora apiculata (tall-stilt mangrove) in Thailand | Not reported |
| Candida konsanensis | Not reported | Not reported | Jasminum adenophyllum (bluegrape jasmine) in Thailand | Not reported |
References
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol. Immunol 2009, 53, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Clinical Alert to U.S. Healthcare Facilities—June 2016|CDC. Available online: https://www.cdc.gov/fungal/candida-auris/candida-auris-alert.html (accessed on 6 June 2019).
- Candida auris Identified in England. Available online: https://www.gov.uk/government/publications/candida-auris-emergence-in-england/candida-auris-identified-in-england (accessed on 6 June 2019).
- 3 October 2016: Candida auris Outbreaks in Health Care Services—Epidemiological Alert|PAHO WHO. Available online: https://www.paho.org/hq/index.php?option=com_content&view=article&id=12565%3A3-october-2016-candida-auris-outbreaks-in-health-care-services-epidemiological-alert&catid=2103%3Arecent-epidemiological-alerts-updates&Itemid=42346&lang=en (accessed on 6 June 2019).
- Magobo, R.E.; Naicker, S.D.; Wadula, J.; Nchabeleng, M.; Coovadia, Y.; Hoosen, A.; Lockhart, S.R.; Govender, N.P. TRAC-South Africa group. Detection of neonatal unit clusters of Candida parapsilosis fungaemia by microsatellite genotyping: Results from laboratory-based sentinel surveillance, South Africa, 2009–2010. Mycoses 2017, 60, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Escandón, P.; Chow, N.A.; Caceres, D.H.; Gade, L.; Berkow, E.L.; Armstrong, P.; Rivera, S.; Misas, E.; Duarte, C.; Moulton-Meissner, H.; et al. Molecular epidemiology of Candida auris in Colombia reveals a highly-related, country-wide colonization with regional patterns in Amphotericin B resistance. Clin. Infect. Dis. 2019, 68, 15–21. [Google Scholar] [PubMed]
- Welsh, R.M.; Bentz, M.L.; Shams, A.; Houston, H.; Lyons, A.; Rose, L.J.; Litvintseva, A.P. Survival, Persistence, and Isolation of the Emerging Multidrug-Resistant Pathogenic Yeast Candida auris on a Plastic Health Care Surface. J. Clin. Microbiol. 2017, 55, 2996–3005. [Google Scholar] [CrossRef] [PubMed]
- Cadnum, J.L.; Shaikh, A.A.; Piedrahita, C.T.; Sankar, T.; Jencson, A.L.; Larkin, E.L.; Ghannoum, M.A.; Donskey, C.J. Effectiveness of Disinfectants Against Candida auris and Other Candida Species. Infect. Control. Hosp. Epidemiol. 2017, 38, 1240–1243. [Google Scholar] [CrossRef] [PubMed]
- Tracking Candida auris|CDC. Available online: https://www.cdc.gov/fungal/candida-auris/tracking-c-auris.html (accessed on 6 June 2019).
- Forsberg, K.; Woodworth, K.; Walters, M.; Berkow, E.L.; Jackson, B.; Chiller, T.; Vallabhaneni, S. Candida auris: The recent emergence of a multidrug-resistant fungal pathogen. Med. Mycol. 2018, 57, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.G.; Shin, J.H.; Uh, Y.; Kang, M.G.; Kim, S.H.; Park, K.H.; Jang, H.-C. First three reported cases of nosocomial fungemia caused by Candida auris. J. Clin. Microbiol. 2011, 49, 3139–3142. [Google Scholar] [CrossRef]
- Iguchi, S.; Mizushima, R.; Kamada, K.; Itakura, Y.; Yoshida, A.; Uzawa, Y.; Arai, Y.; Takaoka, M.; Sato, S.; Goto, A. The Second Candida auris Isolate from Aural Discharge in Japan. Jpn. J. Infect. Dis. 2018, 71, 174–175. [Google Scholar] [CrossRef]
- Pemán, J.; Ruiz-Gaitán, A. How should we manage the C. auris outbreak? Eur. Congr. Clin. Microbiol. Infect. Dis. 2018, S0067. Available online: https://www.escmid.org/escmid_publications/escmid_elibrary/ (accessed on 5 July 2019).
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef]
- Tsay, S.; Welsh, R.M.; Adams, E.H.; Chow, N.A.; Gade, L.; Berkow, E.L.; Poirot, E.; Lutterloh, E.; Quinn, M.; Chaturvedi, S.; et al. Notes from the Field: Ongoing Transmission of Candida auris in Health Care Facilities—United States, June 2016–May 2017. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 514–515. [Google Scholar] [CrossRef] [PubMed]
- Chow, N.A.; Gade, L.; Tsay, S.V.; Forsberg, K.; Greenko, J.A.; Southwick, K.L.; Barrett, P.M.; Kerins, J.L.; Lockhart, S.R.; Chiller, T.M.; et al. Multiple introductions and subsequent transmission of multidrug-resistant Candida auris in the USA: A molecular epidemiological survey. Lancet Infect. Dis. 2018, 18, 1377–1384. [Google Scholar] [CrossRef]
- Chow, N.; de Groot, T.; Badali, H.; Abastabar, M.; Chiller, T.M.; Meis, J.F. A Fifth Clade: New Insights into the Population Structure of Candida auris, Iran, 2018. Emerg. Infect. Dis. 2019, in press. [Google Scholar]
- Abastabar, M.; Haghani, I.; Ahangarkani, F.; Rezai, M.S.; Taghizadeh Armaki, M.; Roodgari, S.; Kiakojuri, K.; Al-Hatmi, A.M.S.; Meis, J.F.; Badali, H. Candida auris otomycosis in Iran and review of recent literature. Mycoses 2019, 62, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Gabaldón, T.; Carreté, L. The birth of a deadly yeast: Tracing the evolutionary emergence of virulence traits in Candida glabrata. FEMS Yeast Res. 2016, 16, fov110. [Google Scholar] [CrossRef]
- Muñoz, J.F.; Gade, L.; Chow, N.A.; Loparev, V.N.; Juieng, P.; Berkow, E.L.; Farrer, R.A.; Litvintseva, A.P.; Cuomo, C.A. Genomic insights into multidrug-resistance, mating and virulence in Candida auris and related emerging species. Nat. Commun. 2018, 9, 5346. [Google Scholar] [CrossRef] [PubMed]
- Daniel, H.-M.; Lachance, M.-A.; Kurtzman, C.P. On the reclassification of species assigned to Candida and other anamorphic ascomycetous yeast genera based on phylogenetic circumscription. Antonie Van Leeuwenhoek 2014, 106, 67–84. [Google Scholar] [CrossRef]
- Asner, S.A.; Giulieri, S.; Diezi, M.; Marchetti, O.; Sanglard, D. Acquired Multidrug Antifungal Resistance in Candida lusitaniae during Therapy. Antimicrob. Agents Chemother. 2015, 59, 7715–7722. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Iqbal, N.; Cleveland, A.A.; Farley, M.M.; Harrison, L.H.; Bolden, C.B.; Baughman, W.; Stein, B.; Hollick, R.; Park, B.J.; et al. Species identification and antifungal susceptibility testing of Candida bloodstream isolates from population-based surveillance studies in two U.S. cities from 2008 to 2011. J. Clin. Microbiol. 2012, 50, 3435–3442. [Google Scholar] [CrossRef]
- Chatterjee, S.; Alampalli, S.V.; Nageshan, R.K.; Chettiar, S.T.; Joshi, S.; Tatu, U.S. Draft genome of a commonly misdiagnosed multidrug resistant pathogen Candida auris. BMC Genom. 2015, 16, 686. [Google Scholar] [CrossRef]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; NIH Intramural Sequencing Center Comparative Sequencing Program; et al. Topographic diversity of fungal and bacterial communities in human skin. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Southwick, K.; Adams, E.H.; Greenko, J.; Ostrowsky, B.; Fernandez, R.; Patel, R.; Quinn, M.; Vallabhaneni, S.; Denis, R.J.; Erazo, R.; et al. 2039. New York State 2016–2018: Progression from Candida auris Colonization to Bloodstream Infection. Open Forum Infect. Dis. 2018, 5, S594–S595. [Google Scholar] [CrossRef]
- Ruiz-Gaitan, A.; Moret, A.M.; Tasias-Pitarch, M.; Aleixandre-Lopez, A.I.; Martínez-Morel, H.; Calabuig, E.; Salavert-Lletí, M.; Ramírez, P.; López-Hontangas, J.L.; Hagen, F.; et al. An outbreak due to Candida auris with prolonged colonization and candidemia in a tertiary care European hospital. Mycoses 2018, 61, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Schelenz, S.; Hagen, F.; Rhodes, J.L.; Abdolrasouli, A.; Chowdhary, A.; Hall, A.; Ryan, L.; Shackleton, J.; Trimlett, R.; Meis, J.F.; et al. First hospital outbreak of the globally emerging Candida auris in a European hospital. Antimicrob. Resist. Infect. Control. 2016, 5, 35. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Sharma, C.; Duggal, S.; Agarwal, K.; Prakash, A.; Singh, P.K.; Jain, S.; Kathuria, S.; Randhawa, H.S.; Hagen, F.; et al. New clonal strain of Candida auris, Delhi, India. Emerg. Infect. Dis. 2013, 19, 1670–1673. [Google Scholar] [CrossRef] [PubMed]
- Emara, M.; Ahmad, S.; Khan, Z.; Joseph, L.; Al-Obaid, I.; Purohit, P.; Bafna, R. Candida auris Candidemia in Kuwait, 2014. Emerg. Infect. Dis. 2015, 21, 1091–1092. [Google Scholar] [CrossRef] [PubMed]
- Robert, V.A.; Casadevall, A. Vertebrate Endothermy Restricts Most Fungi as Potential Pathogens. J. Infect. Dis. 2009, 200, 1623–1626. [Google Scholar] [CrossRef]
- Ekowati, Y.; Ferrero, G.; Kennedy, M.D.; de Roda Husman, A.M.; Schets, F.M. Potential transmission pathways of clinically relevant fungi in indoor swimming pool facilities. Int. J. Hyg. Environ. Health 2018, 221, 1107–1115. [Google Scholar] [CrossRef]
- Limtong, S.; Nasanit, R. Phylloplane Yeasts in Tropical Climates. In Yeasts in Natural Ecosystems: Diversity; Springer: Cham, Switzerland, 2017; pp. 199–223. [Google Scholar]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef]
- Kalan, L.; Loesche, M.; Hodkinson, B.P.; Heilmann, K.; Ruthel, G.; Gardner, S.E.; Grice, E.A. Redefining the Chronic-Wound Microbiome: Fungal Communities Are Prevalent, Dynamic, and Associated with Delayed Healing. mBio 2016, 7. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Global species numbers of fungi: Are tropical studies and molecular approaches contributing to a more robust estimate? Biodivers. Conserv. 2012, 21, 2425–2433. [Google Scholar] [CrossRef]
- Cendejas-Bueno, E.; Kolecka, A.; Alastruey-Izquierdo, A.; Theelen, B.; Groenewald, M.; Kostrzewa, M.; Cuenca-Estrella, M.; Gomez-Lopez, A.; Boekhout, T. Reclassification of the Candida haemulonii Complex as Candida haemulonii (C. haemulonii Group I), C. duobushaemulonii sp. nov. (C. haemulonii Group II), and C. haemulonii var. vulnera var. nov.: Three Multiresistant Human Pathogenic Yeasts. J. Clin. Microbiol. 2012, 50, 3641–3651. [Google Scholar] [PubMed]
- Hou, X.; Xiao, M.; Chen, S.C.-A.; Wang, H.; Cheng, J.-W.; Chen, X.-X.; Xu, Z.-P.; Fan, X.; Kong, F.; Xu, Y.-C. Identification and Antifungal Susceptibility Profiles of Candida haemulonii Species Complex Clinical Isolates from a Multicenter Study in China. J. Clin. Microbiol. 2016, 54, 2676–2680. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prakash, A.; Singh, A.; Kumar, H.; Hagen, F.; Meis, J.F.; Chowdhary, A. Candida haemulonii species complex: An emerging species in India and its genetic diversity assessed with multilocus sequence and amplified fragment-length polymorphism analyses. Emerg. Microbes Infect. 2016, 5, e49. [Google Scholar] [CrossRef] [PubMed]
- Search on: MycoBank_Advanced. Available online: http://www.mycobank.org/Biolomics.aspx?Table=Mycobank_Advanced&Page=200&ViewMode=Basic (accessed on 24 June 2019).
- Van Uden, N.; Kolipinski, M.C. Torulopsis haemulonii nov. spec., a yeast from the Atlantic Ocean. Antonie Van Leeuwenhoek 1962, 28, 78–80. [Google Scholar] [CrossRef]
- Antony, S.P.; Singh, I.S.B.; Sudheer, N.S.; Vrinda, S.; Priyaja, P.; Philip, R. Molecular characterization of a crustin-like antimicrobial peptide in the giant tiger shrimp, Penaeus monodon, and its expression profile in response to various immunostimulants and challenge with WSSV. Immunobiology 2011, 216, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Pagani, D.M.; Heidrich, D.; Paulino, G.V.B.; Alves, K.; de, O.; Dalbem, P.T.; de Oliveira, C.F.; Andrade, Z.M.M.; Silva, C.; Correia, M.D.; et al. Susceptibility to antifungal agents and enzymatic activity of Candida haemulonii and Cutaneotrichosporon dermatis isolated from soft corals on the Brazilian reefs. Arch. Microbiol. 2016, 198, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Candida haemulonii. Available online: http://www.mycobank.org/BioloMICS.aspx?TableKey=14682616000000089&Rec=246&Fields=All (accessed on 6 June 2019).
- Buck, J.D. Occurrence of human-associated yeasts in the feces and pool waters of captive bottlenosed dolphins (Tursiops truncatus). J. Wildl. Dis. 1980, 16, 141–149. [Google Scholar] [CrossRef]
- Ferreira, N.; Belloch, C.; Querol, A.; Manzanares, P.; Vallez, S.; Santos, A. Yeast microflora isolated from brazilian cassava roots: Taxonomical classification based on molecular identification. Curr. Microbiol. 2010, 60, 287–293. [Google Scholar] [CrossRef]
- Loosová, G.; Jindrák, L.; Kopácek, P. Mortality caused by experimental infection with the yeast Candida haemulonii in the adults of Ornithodoros moubata (Acarina: Argasidae). Folia Parasitol. 2001, 48, 149–153. [Google Scholar] [CrossRef]
- Singh, A.; Rani Garg, A.; Garcha, S.; Kamaldeep, K.; Magotra, A. Characterization and optimization of physicochemical properties of dairy effluents. Pollut. Res. 2017, 36, 306–311. [Google Scholar]
- Saluja, P.; Prasad, G.S. Candida ruelliae sp. nov., a novel yeast species isolated from flowers of Ruellia sp. (Acanthaceae). FEMS Yeast Res. 2008, 8, 660–666. [Google Scholar]
- Gargeya, I.B.; Pruitt, W.R.; Meyer, S.A.; Ahearn, D.G. Candida haemulonii from clinical specimens in the USA. J. Med. Vet. Mycol. 1991, 29, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ami, R.; Berman, J.; Novikov, A.; Bash, E.; Shachor-Meyouhas, Y.; Zakin, S.; Maor, Y.; Tarabia, J.; Schechner, V.; Adler, A.; et al. Multidrug-Resistant Candida haemulonii and C. auris, Tel Aviv, Israel. Emerg. Infect. Dis. 2017, 23, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Ramos, L.S.; Figueiredo-Carvalho, M.H.G.; Barbedo, L.S.; Ziccardi, M.; Chaves, A.L.S.; Zancopé-Oliveira, R.M.; Pinto, M.R.; Sgarbi, D.B.G.; Dornelas-Ribeiro, M.; Branquinha, M.H.; et al. Candida haemulonii complex: Species identification and antifungal susceptibility profiles of clinical isolates from Brazil. J. Antimicrob. Chemother. 2015, 70, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.-Y.; Wei, K.-C.; Chen, W.-C.; Lee, S.-J.; Yang, K.-C.; Wu, C.-S.; Sun, P.-L. Primary deep cutaneous candidiasis caused by Candida duobushaemulonii in a 68-year-old man: The first case report and literature review. Mycoses 2016, 59, 818–821. [Google Scholar]
- Boatto, H.F.; Cavalcanti, S.D.B.; Del Negro, G.M.; Girão, M.J.B.; Francisco, E.C.; Ishida, K.; Gompertz, O.F. Candida duobushaemulonii: An emerging rare pathogenic yeast isolated from recurrent vulvovaginal candidiasis in Brazil. Mem. Inst. Oswaldo Cruz 2016, 111, 407–410. [Google Scholar] [CrossRef]
- Isla, G.; Taverna, C.G.; Szusz, W.; Vivot, W.; García-Effron, G.; Davel, G. Candida haemulonii sensu lato: Update of the Determination of Susceptibility Profile in Argentina and Literature Review. Curr. Fungal Infect. Rep. 2017, 11, 203–208. [Google Scholar] [CrossRef]
- Wang, S.-A.; Jia, J.-H.; Bai, F.-Y. Candida alocasiicola sp. nov., Candida hainanensis sp. nov., Candida heveicola sp. nov. and Candida musiphila sp. nov., novel anamorphic, ascomycetous yeast species isolated from plants. Antonie Van Leeuwenhoek 2008, 94, 257–265. [Google Scholar] [CrossRef]
- Sipickzi, M.; Tap, R.M. Candida vulturna pro tempore sp. nov., a dimorphic yeast species related to the Candida haemulonis species complex isolated from flowers and clinical sample. Int. J. Syst. Evol. Microbiol. 2016, 66, 4009–4015. [Google Scholar]
- Limtong, S.; Yongmanitchai, W. Candida chanthaburiensis sp. nov., Candida kungkrabaensis sp. nov. and Candida suratensis sp. nov., three novel yeast species from decaying plant materials submerged in water of mangrove forests. Antonie Van Leeuwenhoek 2010, 98, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Sarawan, S.; Mahakhan, P.; Jindamorakot, S.; Vichitphan, K.; Vichitphan, S.; Sawaengkaew, J. Candida konsanensis sp. nov., a new yeast species isolated from Jasminum adenophyllum in Thailand with potentially carboxymethyl cellulase-producing capability. World J. Microbiol. Biotechnol. 2013, 29, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Lachance, M.A. Metschnikowia: Half tetrads, a regicide and the fountain of youth. Yeast 2016, 33, 563–574. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Polme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [PubMed]
- Bensasson, D.; Dicks, J.; Ludwig, J.M.; Bond, C.J.; Elliston, A.; Roberts, I.N.; James, S.A. Diverse Lineages of Candida albicans Live on Old Oaks. Genetics 2019, 211, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.; Yang, J.; Vogan, A.; Maganti, H.; Yamamura, D.; Xu, J. Clinical and tree hollow populations of human pathogenic yeast in Hamilton, Ontario, Canada are different. Mycoses 2014, 57, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Gabaldón, T.; Fairhead, C. Genomes shed light on the secret life of Candida glabrata: Not so asexual, not so commensal. Curr. Genet. 2019, 65, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Kemler, M.; Witfeld, F.; Begerow, D.; Yurkov, A. Phylloplane Yeasts in Temperate Climates. In Yeasts in Natural Ecosystems: Diversity; Buzzini, P., Lachance, M.-A., Yurkov, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 171–197. [Google Scholar]
- Madden, A.A.; Epps, M.J.; Fukami, T.; Irwin, R.E.; Sheppard, J.; Sorger, D.M.; Dunn, R.R. The ecology of insect–yeast relationships and its relevance to human industry. Proc. Biol. Sci. 2018, 285, 20172733. [Google Scholar] [CrossRef]
- Stefanini, I.; Dapporto, L.; Legras, J.L.; Calabretta, A.; Paola, M.D.; Filippo, C.D.; Viola, R.; Capretti, P.; Polsinelli, M.; Turillazzi, S.; et al. Role of social wasps in Saccharomyces cerevisiae ecology and evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13398–13403. [Google Scholar] [CrossRef]
- Adams, E.; Quinn, M.; Tsay, S.; Poirot, E.; Chaturvedi, S.; Southwick, K.; Greenko, J.; Fernandez, R.; Kallen, A.; Vallabhaneni, S.; et al. Candida auris in Healthcare Facilities, New York, USA, 2013–2017. Emerg. Infect. Dis. 2018, 24, 1816–1824. [Google Scholar] [CrossRef]
- Won, S.Y.; Munoz-Price, L.S.; Lolans, K.; Hota, B.; Weinstein, R.A.; Hayden, M.K.; Centers for Disease Control and Prevention Epicenter Program. Emergence and rapid regional spread of Klebsiella pneumoniae carbapenemase-producing Enterobacteriaceae. Clin. Infect. Dis. 2011, 53, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Eyre, D.W.; Sheppard, A.E.; Madder, H.; Moir, I.; Moroney, R.; Quan, T.P.; Griffiths, D.; George, S.; Butcher, L.; Morgan, M.; et al. A Candida auris Outbreak and Its Control in an Intensive Care Setting. N. Engl. J. Med. 2018, 379, 1322–1331. [Google Scholar] [CrossRef] [PubMed]
- Jeffery-Smith, A.; Taori, S.K.; Schelenz, S.; Jeffery, K.; Johnson, E.M.; Borman, A.; Team, Candida auris Incident Management Team; Manuel, R.; Brown, C.S. Candida auris: A Review of the Literature. Clin. Microbiol. Rev. 2018, 31, e00029-17. [Google Scholar] [PubMed]
- Magobo, R.E.; Corcoran, C.; Seetharam, S.; Govender, N.P. Candida auris–Associated Candidemia, South Africa. Emerg. Infect. Dis. 2014, 20, 1250–1252. [Google Scholar] [CrossRef] [PubMed]
- Welsh, R.M.; Sexton, D.J.; Forsberg, K.; Vallabhaneni, S.; Litvintseva, A. Insights into the Unique Nature of the East Asian Clade of the Emerging Pathogenic Yeast Candida auris. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef] [PubMed]
- Yoshiura, K.; Kinoshita, A.; Ishida, T.; Ninokata, A.; Ishikawa, T.; Kaname, T.; Bannai, M.; Tokunaga, K.; Sonoda, S.; Komaki, R.; et al. A SNP in the ABCC11 gene is the determinant of human earwax type. Nat. Genet. 2006, 38, 324–330. [Google Scholar] [CrossRef]
- Lum, C.L.; Jeyanthi, S.; Prepageran, N.; Vadivelu, J.; Raman, R. Antibacterial and antifungal properties of human cerumen. J. Laryngol. Otol. 2009, 123, 375–378. [Google Scholar] [CrossRef]
- Steffen, W.; Broadgate, W.; Deutsch, L.; Gaffney, O.; Ludwig, C. The trajectory of the Anthropocene: The Great Acceleration. Anthr. Rev. 2015, 2, 81–98. [Google Scholar] [CrossRef]
- Berger, S.; El Chazli, Y.; Babu, A.F.; Coste, A.T. Azole Resistance in Aspergillus fumigatus: A Consequence of Antifungal Use in Agriculture? Front. Microbiol. 2017, 8, 789–795. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/RP (accessed on 6 June 2019).
- Meis, J.F.; Chowdhary, A.; Rhodes, J.L.; Fisher, M.C.; Verweij, P.E. Clinical implications of globally emerging azole resistance in Aspergillus fumigatus. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2016, 371, 20150460. [Google Scholar] [CrossRef]
- Lübbert, C.; Baars, C.; Dayakar, A.; Lippmann, N.; Rodloff, A.C.; Kinzig, M.; Sörgel, F. Environmental pollution with antimicrobial agents from bulk drug manufacturing industries in Hyderabad, South India, is associated with dissemination of extended-spectrum beta-lactamase and carbapenemase-producing pathogens. Infection 2017, 45, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Thomas, N.; Lucas, R.; Bunting, P.; Hardy, A.; Rosenqvist, A.; Simard, M. Distribution and drivers of global mangrove forest change, 1996–2010. PLoS ONE 2017, 12, e0179302. [Google Scholar] [CrossRef] [PubMed]
- Bostock, J.; McAndrew, B.; Richards, R.; Jauncey, K.; Telfer, T.; Lorenzen, K.; Little, D.; Ross, L.; Handisyde, N.; Gatward, I.; et al. Aquaculture: Global status and trends. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2897–2912. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C.; Godfrey, H.P.; Tomova, A.; Ivanova, L.; Dölz, H.; Millanao, A.; Buschmann, A.H. Antimicrobial use in aquaculture re-examined: Its relevance to antimicrobial resistance and to animal and human health: Aquacultural antimicrobial use and antimicrobial resistance. Environ. Microbiol. 2013, 15, 1917–1942. [Google Scholar] [CrossRef] [PubMed]
- Hai, N.V. The use of probiotics in aquaculture. J. Appl. Microbiol. 2015, 119, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Solache, M.A.; Casadevall, A. Global Warming Will Bring New Fungal Diseases for Mammals. mBio 2010, 1, e00061-10. [Google Scholar] [CrossRef] [PubMed]
- Altizer, S.; Ostfeld, R.S.; Johnson, P.T.J.; Kutz, S.; Harvell, C.D. Climate Change and Infectious Diseases: From Evidence to a Predictive Framework. Science 2013, 341, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef]
- O’Hanlon, S.J.; Rieux, A.; Farrer, R.A.; Rosa, G.M.; Waldman, B.; Bataille, A.; Kosch, T.A.; Murray, K.A.; Brankovics, B.; Fumagalli, M.; et al. Recent Asian origin of chytrid fungi causing global amphibian declines. Science 2018, 360, 621–627. [Google Scholar] [CrossRef]
- Leopardi, S.; Blake, D.; Puechmaille, S.J. White-Nose Syndrome fungus introduced from Europe to North America. Curr. Biol. 2015, 25, R217–R219. [Google Scholar] [CrossRef]
- Tsay, S.; Kallen, A.; Jackson, B.R.; Chiller, T.M.; Vallabhaneni, S. Approach to the Investigation and Management of Patients with Candida auris, an Emerging Multidrug-Resistant Yeast. Clin. Infect. Dis. 2018, 66, 306–311. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackson, B.R.; Chow, N.; Forsberg, K.; Litvintseva, A.P.; Lockhart, S.R.; Welsh, R.; Vallabhaneni, S.; Chiller, T. On the Origins of a Species: What Might Explain the Rise of Candida auris? J. Fungi 2019, 5, 58. https://doi.org/10.3390/jof5030058
Jackson BR, Chow N, Forsberg K, Litvintseva AP, Lockhart SR, Welsh R, Vallabhaneni S, Chiller T. On the Origins of a Species: What Might Explain the Rise of Candida auris? Journal of Fungi. 2019; 5(3):58. https://doi.org/10.3390/jof5030058
Chicago/Turabian StyleJackson, Brendan R., Nancy Chow, Kaitlin Forsberg, Anastasia P. Litvintseva, Shawn R. Lockhart, Rory Welsh, Snigdha Vallabhaneni, and Tom Chiller. 2019. "On the Origins of a Species: What Might Explain the Rise of Candida auris?" Journal of Fungi 5, no. 3: 58. https://doi.org/10.3390/jof5030058
APA StyleJackson, B. R., Chow, N., Forsberg, K., Litvintseva, A. P., Lockhart, S. R., Welsh, R., Vallabhaneni, S., & Chiller, T. (2019). On the Origins of a Species: What Might Explain the Rise of Candida auris? Journal of Fungi, 5(3), 58. https://doi.org/10.3390/jof5030058