Immunopathogenesis of Human Sporotrichosis: What We Already Know



Abstract
1. The Global Increase of Sporotrichosis Creates New Problems as Well as Knowledge Opportunities
2. Changes in the Epidemiology of Sporotrichosis Infection Alter Both, the Transmission and the Pathogenesis of the Mycosis
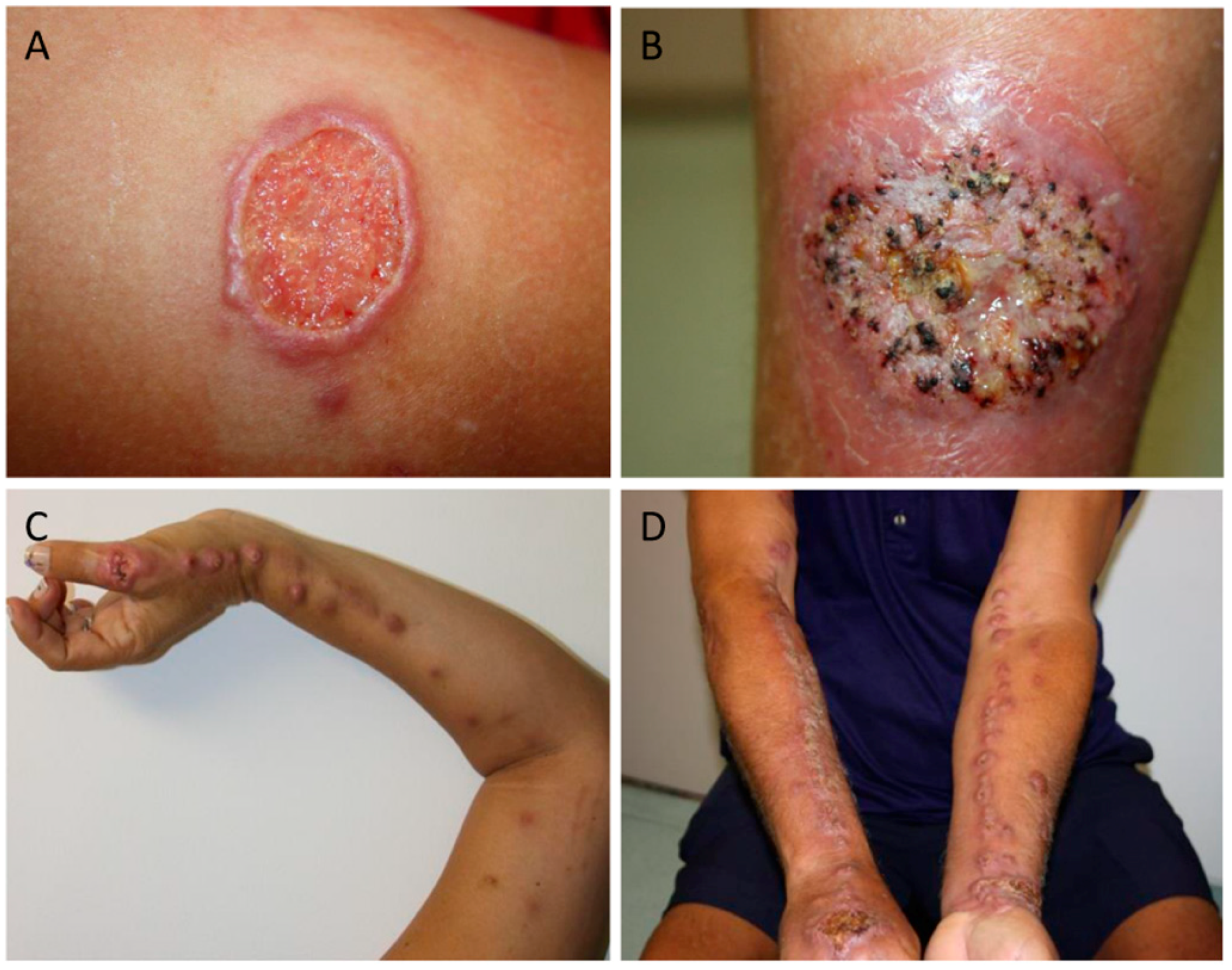
3. Clinical Presentation of the Human Sporotrichosis
4. Is the Etiological Agent a Source of Differences in Clinical Presentation?
5. Immune Response in Human Sporotrichosis
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gremião, I.D.F.; Miranda, L.H.M.; Reis, E.G.; Rodrigues, A.M.; Pereira, S.A. Zoonotic Epidemic of Sporotrichosis: Cat to Human Transmission. PLoS Pathog. 2017, 13, e1006077. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Bonifaz, A.; Gutierrez-Galhardo, M.C.; Mochizuki, T.; Li, S. Global epidemiology of sporotrichosis. Med. Mycol. 2015, 53, 3–14. [Google Scholar] [CrossRef] [PubMed]
- López-Romero, E.; Reyes-Montes, M.D.; Pérez-Torres, A.; Ruiz-Baca, E.; Villagómez-Castro, J.C.; Mora-Montes, H.M.; Flores-Carreón, A.; Toriello, C. Sporothrix schenckii complex and sporotrichosis, an emerging health problem. Future Microbiol. 2011, 6, 85–102. [Google Scholar] [CrossRef] [PubMed]
- Bhutia, P.Y.; Gurung, S.; Yegneswaran, P.P.; Pradhan, J.; Pradhan, U.; Peggy, T.; Pradhan, P.K.; Bhutia, C.D. A case series and review of sporotrichosis in Sikkim. J. Infect. Dev. Ctries. 2011, 5, 603–608. [Google Scholar] [PubMed]
- Dias, N.M.; Oliveira, M.M.E.; Santos, C.; Zancope-Oliveira, R.M.; Lima, N. Sporotrichosis caused by Sporothrix mexicana, Portugal. Emerg. Infect. Dis. 2011, 17, 1975–1976. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Yao, L.; Zhong, S.-X.; Tian, Y.-P.; Liu, Y.-Y.; Li, S.-S. Infant sporotrichosis in northeast China: A report of 15 cases. Int. J. Dermatol. 2011, 50, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Sivagnanam, S.; Bannan, A.M.; Chen, S.C.-A.; Ralph, A.P. Sporotrichosis (Sporothrix schenckii infection) in the New South Wales mid-north coast, 2000–2010. Med. J. Aust. 2012, 196, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Verma, G.K.; Singh, G.; Kanga, A.; Shanker, V.; Singh, D.; Gupta, P.; Mokta, K.; Sharma, V. Sporotrichosis in sub-himalayan India. PLoS Negl. Trop. Dis. 2012, 6, e1673. [Google Scholar] [CrossRef] [PubMed]
- Mata-Essayag, S.; Delgado, A.; Colella, M.T.; Landaeta-Nezer, M.E.; Rosello, A.; Perez de Salazar, C.; Olaizola, C.; Hartung, C.; Magaldi, S.; Velasquez, E. Epidemiology of sporotrichosis in Venezuela. Int. J. Dermatol. 2013, 52, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.M.E.; Almeida-Paes, R.; Gutierrez-Galhardo, M.C.; Zancope-Oliveira, R.M. Molecular identification of the Sporothrix schenckii complex. Rev. Iberoam. Micol. 2014, 31, 2–6. [Google Scholar] [CrossRef] [PubMed]
- De Lima Barros, M.B.; de Almeida Paes, R.; Schubach, A.O. Sporothrix schenckii and Sporotrichosis. Clin. Microbiol. Rev. 2011, 24, 633–654. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Bezerra, L.M.; Mora-Montes, H.M.; Zhang, Y.; Nino-Veja, G.; Rodrigues, A.M.; de Camargo, Z.P.; de Hoog, S. Sporotrichosis between 1898 and 2017: The evolution of knowledge on a changeable disease and on emerging etiological agents. Med. Mycol. 2018, 56, 126–143. [Google Scholar] [CrossRef] [PubMed]
- Sanchotene, K.O.; Madrid, I.M.; Klafke, G.B.; Bergamashi, M.; Della Terra, P.P.; Rodrigues, A.M.; de Camargo, Z.P.; Xavier, M.O. Sporothrix brasiliensis outbreaks and the rapid emergence of feline sporotrichosis. Mycoses 2015, 58, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Téllez, M.D.; Batista-Duharte, A.; Portuondo, D.; Quinello, C.; Bonne-Hernández, R.; Carlos, I.Z. Sporothrix schenckii complex biology: Environment and fungal pathogenicity. Microbiology 2014, 160, 2352–2365. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Gamboa, L.; Martínez-Hernandez, F.; Maravilla, P.; Arenas-Guzmán, R.; Flisser, A. Update of phylogenetic and genetic diversity of Sporothrix schenckii sensu lato. Med. Mycol. 2016, 54, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; de Hoog, G.S.; Zhang, Y.; de Camargo, Z.P. Emerging sporotrichosis is driven by clonal and recombinant Sporothrix species. Emerg. Microbes Infect. 2014, 3, e32. [Google Scholar] [CrossRef] [PubMed]
- Miranda, L.H.M.; Quintella, L.P.; Menezes, R.C.; dos Santos, I.B.; Oliveira, R.V.C.; Figueiredo, F.B.; Lopes-Bezerra, L.M.; Schubach, T.M. Evaluation of immunohistochemistry for the diagnosis of sporotrichosis in dogs. Vet. J. 2011, 190, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.A.; Menezes, R.C.; Gremião, I.D.F.; Silva, J.N.; de Honse, C.O.; Figueiredo, F.B.; da Silva, D.T.; Kitada, A.A.; dos Reis, E.G.; Schubach, T.M. Sensitivity of cytopathological examination in the diagnosis of feline sporotrichosis. J. Feline Med. Surg. 2011, 13, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Rees, R.K.; Swartzberg, J.E. Feline-transmitted sporotrichosis: A case study from California. Dermatol. Online J. 2011, 17, 2. [Google Scholar] [PubMed]
- Borges, T.S.; Rossi, C.N.; Fedullo, J.D.L.; Taborda, C.P.; Taborda, J.P.; Larsson, C.E. Isolation of Sporothrix schenckii from the claws of domestic cats (indoor and outdoor) and in captivity in São Paulo (Brazil). Mycopathologia 2013, 176, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Miranda, L.H.M.; Conceição-Silva, F.; Quintella, L.P.; Kuraiem, B.P.; Pereira, S.A.; Schubach, T.M.P. Feline sporotrichosis: Histopathological profile of cutaneous lesions and their correlation with clinical presentation. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Bonifaz, A.; Tirado-Sánchez, A. Cutaneous Disseminated and Extracutaneous Sporotrichosis: Current Status of a Complex Disease. J. Fungi 2017, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.O.; Bernardes-Engemann, A.R.; Azulay-Abulafia, L.; Benvenuto, F.; de Neves, M.L.P.; Lopes-Bezerra, L.M. Sporotrichosis in pregnancy: Case reports of 5 patients in a zoonotic epidemic in Rio de Janeiro, Brazil. An. Bras. Dermatol. 2011, 86, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.P.; do Valle, A.C.F.; Freitas, D.F.S.; Reis, R.; Galhardo, M.C.G. Pregnancy during a sporotrichosis epidemic in Rio de Janeiro, Brazil. Int. J. Gynaecol. Obstet. 2012, 117, 294–295. [Google Scholar] [CrossRef] [PubMed]
- Freitas, D.F.S.; de Siqueira Hoagland, B.; do Valle, A.C.F.; Fraga, B.B.; de Barros, M.B.; de Oliveira Schubach, A.; de Almeida-Paes, R.; Cuzzi, T.; Rosalino, C.M.; Zancopé-Oliveira, R.M.; et al. Sporotrichosis in HIV-infected patients: Report of 21 cases of endemic sporotrichosis in Rio de Janeiro, Brazil. Med. Mycol. 2012, 50, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Hersh, A.M.; Naughton, G.; Mullins, K.; Fung, M.A.; Sharon, V.R. Disseminated cutaneous sporotrichosis. Dermatol. Online J. 2013, 19, 20401. [Google Scholar] [PubMed]
- Gewehr, P.; Jung, B.; Aquino, V.; Manfro, R.C.; Spuldaro, F.; Rosa, R.G.; Goldani, L.Z. Sporotrichosis in renal transplant patients. Can. J. Infect. Dis. Med. Microbiol. 2013, 24, e47–e49. [Google Scholar] [CrossRef] [PubMed]
- Orofino-Costa, R.; Unterstell, N.; Carlos Gripp, A.; de Macedo, P.M.; Brota, A.; Dias, E.; de Melo Teixeira, M.; Felipe, M.S.; Bernardes-Engemann, A.R.; Lopes-Bezerra, L.M. Pulmonary cavitation and skin lesions mimicking tuberculosis in a HIV negative patient caused by Sporothrix brasiliensis. Med. Mycol. Case Rep. 2013, 2, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Sharon, V.R.; Kim, J.; Sudhakar, S.; Fung, M.A.; Maniar, A. Disseminated cutaneous sporotrichosis. Lancet Infect. Dis. 2013, 13, 95. [Google Scholar] [CrossRef]
- Xavier, M.O.; Bittencourt, L.R.; da Silva, C.M.; Vieira, R.S.; Pereira, H.C.P. Atypical presentation of sporotrichosis: Report of three cases. Rev. Soc. Bras. Med. Trop. 2013, 46, 116–118. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho Aguinaga, F.; Trope, B.M.; Fernandes, N.C.; Engel, D.C.; Ramos-E-Silva, M. Sporotrichosis with bone involvement: An alert to an occupational disease. Case Rep. Dermatol. 2014, 6, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Pyla, V. Cancer-like lesions in a patient with sporotrichosis. Int. J. Dermatol. 2014, 53, e311–e312. [Google Scholar] [CrossRef] [PubMed]
- Miranda, L.H.M.; de Santiago, M.A.; Schubach, T.M.P.; Morgado, F.N.; Pereira, S.A.; de Oliveira, R.V.C.; Conceição-Silva, F. Severe feline sporotrichosis associated with an increased population of CD8low cells and a decrease in CD4+ cells. Med. Mycol. 2016, 54, 29–39. [Google Scholar] [PubMed]
- Oliveira, M.M.E.; Almeida-Paes, R.; Muniz, M.M.; Gutierrez-Galhardo, M.C.; Zancope-Oliveira, R.M. Phenotypic and molecular identification of Sporothrix isolates from an epidemic area of sporotrichosis in Brazil. Mycopathologia 2011, 172, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, G.F.; dos Santos, P.O.; Rodrigues, A.M.; Sasaki, A.A.; Burger, E.; de Camargo, Z.P. Characterization of virulence profile, protein secretion and immunogenicity of different Sporothrix schenckii sensu stricto isolates compared with S. globosa and S. brasiliensis species. Virulence 2013, 4, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Z.; Hou, B.; Wang, D.; Sun, T.; Li, F.; Wang, H.; Han, S. Rapid identification of Sporothrix schenckii in biopsy tissue by PCR. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 1491–1497. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.M.E.; Maifrede, S.B.; Ribeiro, M.A.; Zancope-Oliveira, R.M. Molecular identification of Sporothrix species involved in the first familial outbreak of sporotrichosis in the state of Espírito Santo, southeastern Brazil. Mem. Inst. Oswaldo Cruz 2013, 108, 936–938. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; de Hoog, G.S.; de Camargo, Z.P. Genotyping species of the Sporothrix schenckii complex by PCR-RFLP of calmodulin. Diagn. Microbiol. Infect. Dis. 2014, 78, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, T.; Matsuyama, T.; Mitsuyama, M. Characteristic infectivity of Sporothrix schenckii to mice depending on routes of infection and inherent fungal pathogenicity. Med. Mycol. 1998, 36, 21–27. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Morgado, F.N.; Schubach, A.O.; Barros, M.B.L.; Conceição-Silva, F. The in situ inflammatory profile of lymphocutaneous and fixed forms of human sporotrichosis. Med. Mycol. 2011, 49, 612–620. [Google Scholar] [PubMed]
- Guzman-Beltran, S.L.; Perez-Torres, A.; Coronel-Cruz, C.; Torres-Guerrero, H. Phagocytic receptors on macrophages distinguish between different Sporothrix schenckii morphotypes. Microbes Infect. 2012, 14, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, Q.; Sun, J.; Shen, Y.; Liu, W. Inflammatory response of human keratinocytes triggered by Sporothrix schenckii via Toll-like receptor 2 and 4. J. Dermatol. Sci. 2012, 66, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Romo-Lozano, Y.; Hernández-Hernández, F.; Salinas, E. Mast cell activation by conidia of Sporothrix schenckii: Role in the severity of infection. Scand. J. Immunol. 2012, 76, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Verdan, F.F.; Faleiros, J.C.; Ferreira, L.S.; Monnazzi, L.G.S.; Maia, D.C.G.; Tansine, A.; Placeres, M.C.; Carlos, I.Z.; Santos-Junior, R.R. Dendritic cell are able to differentially recognize Sporothrix schenckii antigens and promote Th1/Th17 response in vitro. Immunobiology 2012, 217, 788–794. [Google Scholar] [CrossRef] [PubMed]
- De Negrini, T.C.; Ferreira, L.S.; Alegranci, P.; Arthur, R.A.; Sundfeld, P.P.; Maia, D.C.G.; Spolidorio, L.C.; Carlos, I.Z. Role of TLR-2 and fungal surface antigens on innate immune response against Sporothrix schenckii. Immunol. Investig. 2013, 42, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Galhardo, M.C.G.; De Oliveira, R.M.Z.; Valle, A.C.F.D.; Paes, R.D.A.; Silvatavares, P.M.E.; Monzon, A.; Mellado, E.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. Molecular epidemiology and antifungal susceptibility patterns of Sporothrix schenckii isolates from a cat-transmitted epidemic of sporotrichosis in Rio de Janeiro, Brazil. Med. Mycol. 2008, 46, 141–151. [Google Scholar] [CrossRef] [PubMed]
- De Lima Barros, M.B.; Schubach, A.O.; de Vasconcellos Carvalhaes de Oliveira, R.; Martins, E.B.; Teixeira, J.L.; Wanke, B. Treatment of cutaneous sporotrichosis with itraconazole—Study of 645 patients. Clin. Infect. Dis. 2011, 52, e200–e206. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Silva, F.; Capilla, J.; Mayayo, E.; Guarro, J. Modest efficacy of voriconazole against murine infections by Sporothrix schenckii and lack of efficacy against Sporothrix brasiliensis. Mycoses 2014, 57, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Ottonelli Stopiglia, C.D.; Magagnin, C.M.; Castrillón, M.R.; Mendes, S.D.C.; Heidrich, D.; Valente, P.; Scroferneker, M.L. Antifungal susceptibilities and identification of species of the Sporothrix schenckii complex isolated in Brazil. Med. Mycol. 2014, 52, 56–64. [Google Scholar] [PubMed]
- Della Terra, P.P.; Rodrigues, A.M.; Fernandes, G.F.; Nishikaku, A.S.; Burger, E.; de Camargo, Z.P. Exploring virulence and immunogenicity in the emerging pathogen Sporothrix brasiliensis. PLoS Negl. Trop. Dis. 2017, 11, e0005903. [Google Scholar] [CrossRef] [PubMed]
- Brito, M.M.S.; Conceição-Silva, F.; Morgado, F.N.; Raibolt, P.S.; Schubach, A.; Schubach, T.P.; Schäffer, G.M.; Borba, C.M. Comparison of virulence of different Sporothrix schenckii clinical isolates using experimental murine model. Med. Mycol. 2007, 45, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Boyce, K.J.; Andrianopoulos, A. Fungal dimorphism: The switch from hyphae to yeast is a specialized morphogenetic adaptation allowing colonization of a host. FEMS Microbiol. Rev. 2015, 39, 797–811. [Google Scholar] [CrossRef] [PubMed]
- Schubach, A.; de Barros, M.B.L.; Wanke, B. Epidemic sporotrichosis. Curr. Opin. Infect. Dis. 2008, 21, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Gremião, I.D.F.; Menezes, R.C.; Schubach, T.M.P.; Figueiredo, A.B.F.; Cavalcanti, M.C.H.; Pereira, S.A. Feline sporotrichosis: Epidemiological and clinical aspects. Med. Mycol. 2015, 53, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.B.L.; Schubach, A.O.; Schubach, T.M.P.; Wanke, B.; Lambert-Passos, S.R. An epidemic of sporotrichosis in Rio de Janeiro, Brazil: Epidemiological aspects of a series of cases. Epidemiol. Infect. 2008, 136, 1192–1196. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Bezerra, L.M.; Schubach, A.; Costa, R.O. Sporothrix schenckii and sporotrichosis. An. Acad. Bras. Cienc. 2006, 78, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Schubach, A.; Schubach, T.M.P.; de Barros, M.B.L.; Wanke, B. Cat-transmitted sporotrichosis, Rio de Janeiro, Brazil. Emerg. Infect. Dis. 2005, 11, 1952–1954. [Google Scholar] [CrossRef] [PubMed]
- De Barros, M.B.L.; de Schubach, A.O.; do Valle, A.C.F.; Gutierrez Galhardo, M.C.; Conceição-Silva, F.; Schubach, T.M.P.; Reis, R.S.; Wanke, B.; Marzochi, K.B.; Conceição, M.J. Cat-transmitted sporotrichosis epidemic in Rio de Janeiro, Brazil: Description of a series of cases. Clin. Infect. Dis. 2004, 38, 529–535. [Google Scholar] [CrossRef] [PubMed]
- De Lima Barros, M.B.; Schubach, T.M.; Galhardo, M.C.; de Oliviera Schubach, A.; Monteiro, P.C.; Reis, R.S.; Zancopé-Oliveira, R.M.; dos Santos Lazéra, M.; Cuzzi-Maya, T.; Blanco, T.C.; et al. Sporotrichosis: An emergent zoonosis in Rio de Janeiro. Mem. Inst. Oswaldo Cruz 2001, 96, 777–779. [Google Scholar] [CrossRef] [PubMed]
- Baum, G.L.; Donnerberg, R.L.; Stewart, D.; Mulligan, W.J.; Putnam, L.R. Pulmonary sporotrichosis. N. Engl. J. Med. 1969, 280, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.G.; Morgan, W.K.; Hornick, R.B.; Funk, A.M. Chronic pulmonary sporotrichosis: Report of a case, including morphologic and mycologic studies. Am. J. Clin. Pathol. 1970, 54, 401–409. [Google Scholar] [CrossRef] [PubMed]
- England, D.M.; Hochholzer, L. Primary pulmonary sporotrichosis. Report of eight cases with clinicopathologic review. Am. J. Surg. Pathol. 1985, 9, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Donabedian, H.; O’Donnell, E.; Olszewski, C.; MacArthur, R.D.; Budd, N. Disseminated cutaneous and meningeal sporotrichosis in an AIDS patient. Diagn. Microbiol. Infect. Dis. 1994, 18, 111–115. [Google Scholar] [CrossRef]
- Khabie, N.; Boyce, T.G.; Roberts, G.D.; Thompson, D.M. Laryngeal sporotrichosis causing stridor in a young child. Int. J. Pediatr. Otorhinolaryngol. 2003, 67, 819–823. [Google Scholar] [CrossRef]
- Silva-Vergara, M.L.; Maneira, F.R.Z.; De Oliveira, R.M.; Santos, C.T.B.; Etchebehere, R.M.; Adad, S.J. Multifocal sporotrichosis with meningeal involvement in a patient with AIDS. Med. Mycol. 2005, 43, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Appenzeller, S.; Amaral, T.N.; Amstalden, E.M.I.; Bertolo, M.B.; Neto, J.F.M.; Samara, A.M.; Fernandes, S.R. Sporothrix schenckii infection presented as monoarthritis: Report of two cases and review of the literature. Clin. Rheumatol. 2006, 25, 926–928. [Google Scholar] [CrossRef] [PubMed]
- Aung, A.K.; Spelman, D.W.; Thompson, P.J. Pulmonary Sporotrichosis: An Evolving Clinical Paradigm. Semin. Respir. Crit. Care Med. 2015, 36, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Freitas, D.F.S.; Santos, S.S.; Almeida-Paes, R.; de Oliveira, M.M.E.; do Valle, A.C.F.; Gutierrez-Galhardo, M.C.; Zancopé-Oliveira, R.M.; Nosanchuk, J.D. Increase in virulence of Sporothrix brasiliensis over five years in a patient with chronic disseminated sporotrichosis. Virulence 2015, 6, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Freitas, D.F.S.; Lima, M.A.; de Almeida-Paes, R.; Lamas, C.C.; do Valle, A.C.F.; Oliveira, M.M.E.; Zancopé-Oliveira, R.M.; Gutierrez-Galhardo, M.C. Sporotrichosis in the Central Nervous System Caused by Sporothrix brasiliensis. Clin. Infect. Dis. 2015, 61, 663–664. [Google Scholar] [CrossRef] [PubMed]
- Moreira, J.A.S.; Freitas, D.F.S.; Lamas, C.C. The impact of sporotrichosis in HIV-infected patients: A systematic review. Infection 2015, 43, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Biancardi, A.L.; Freitas, D.F.S.; Vitor, R.D.A.; Andrade, H.B.; de Oliveira, M.M.E.; do Valle, A.C.F.; Zancope-Oliveira, R.M.; Galhardo, M.C.; Curi, A.L. Multifocal choroiditis in disseminated sporotrichosis in patients with HIV/AIDS. Retin. Cases Brief Rep. 2017, 11, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Ramírez Soto, M.C. Differences in clinical ocular outcomes between exogenous and endogenous endophthalmitis caused by Sporothrix: A systematic review of published literature. Br. J. Ophthalmol. 2018, 102, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Paixão, A.G.; Galhardo, M.C.G.; Almeida-Paes, R.; Nunes, E.P.; Gonçalves, M.L.C.; Chequer, G.L.; Lamas, C.D.C. The difficult management of disseminated Sporothrix brasiliensis in a patient with advanced AIDS. AIDS Res. Ther. 2015, 12, 16. [Google Scholar] [CrossRef] [PubMed]
- Galhardo, M.C.G.; Silva, M.T.T.; Lima, M.A.; Nunes, E.P.; Schettini, L.E.C.; de Freitas, R.F.; de Paes, R.A.; de Neves, E.S.; do Valle, A.C. Sporothrix schenckii meningitis in AIDS during immune reconstitution syndrome. J. Neurol. Neurosurg. Psychiatry 2010, 81, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Lyra, M.R.; Nascimento, M.L.F.O.; Varon, A.G.; Pimentel, M.I.F.; de Antonio, L.F.; Saheki, M.N.; Bedoya-Pacheco, S.J.; Valle, A.C. Immune reconstitution inflammatory syndrome in HIV and sporotrichosis coinfection: Report of two cases and review of the literature. Rev. Soc. Bras. Med. Trop. 2014, 47, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Rojas, F.D.; Fernández, M.S.; Lucchelli, J.M.; Lombardi, D.; Malet, J.; Vetrisano, M.E.; Cattana, M.E.; Sosa, M.L.Á.; Giusiano, G. Cavitary Pulmonary Sporotrichosis: Case Report and Literature Review. Mycopathologia 2017, 182, 1119–1123. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez Galhardo, M.C.; de Oliveira Schubach, A.; de Lima Barros, M.B.; Moita Blanco, T.C.; Cuzzi-Maya, T.; Pacheco Schubach, T.M.; dos Santos Lazéra, M.; do Valle, A.C. Erythema nodosum associated with sporotrichosis. Int. J. Dermatol. 2002, 41, 114–116. [Google Scholar] [CrossRef] [PubMed]
- Papaiordanou, F.; da Silveira, B.R.L.; Abulafia, L.A. Hypersensitivity reaction to Sporothrix schenckii: Erythema nodosum associated with sporotrichosis. Rev. Soc. Bras. Med. Trop. 2015, 48, 504. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, L.M.V.; Pimentel, M.I.F.; Conceição-Silva, F.; Valete-Rosalino, C.M.; Lyra, M.R.; Salgueiro, M.M.; Saheki, M.N.; Madeira, M.F.; Mouta-Confort, E.; Antonio, L.F.; et al. Sporotrichoid leishmaniasis: A cross-sectional clinical, epidemiological and laboratory study in Rio de Janeiro State, Brazil. Rev. Inst. Med. Trop. São Paulo 2017, 59, e33. [Google Scholar] [CrossRef] [PubMed]
- Morgado, F.N.; de Carvalho, L.M.V.; Leite-Silva, J.; Seba, A.J.; Pimentel, M.I.F.; Fagundes, A.; Madeira, M.F.; Lyra, M.R.; Oliveira, M.M.; Schubach, A.O.; et al. Unbalanced inflammatory reaction could increase tissue destruction and worsen skin infectious diseases—A comparative study of leishmaniasis and sporotrichosis. Sci. Rep. 2018, 8, 2898. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Paes, R.; Oliveira, M.M.E.; Freitas, D.F.S.; do Valle, A.C.F.; Gutierrez-Galhardo, M.C.; Zancopé-Oliveira, R.M. Refractory sporotrichosis due to Sporothrix brasiliensis in humans appears to be unrelated to in vivo resistance. Med. Mycol. 2017, 55, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Paes, R.; Brito-Santos, F.; Figueiredo-Carvalho, M.H.G.; Machado, A.C.S.; Oliveira, M.M.E.; Pereira, S.A.; Gutierrez-Galhardo, M.C.; Zancopé-Oliveira, R.M. Minimal inhibitory concentration distributions and epidemiological cutoff values of five antifungal agents against Sporothrix brasiliensis. Mem. Inst. Oswaldo Cruz 2017, 112, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Chávez, M.J.; Pérez-García, L.A.; Niño-Vega, G.A.; Mora-Montes, H.M. Fungal Strategies to Evade the Host Immune Recognition. J. Fungi 2017, 3, 51. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Paes, R.; de Oliveira, L.C.; Oliveira, M.M.E.; Gutierrez-Galhardo, M.C.; Nosanchuk, J.D.; Zancopé-Oliveira, R.M. Phenotypic characteristics associated with virulence of clinical isolates from the Sporothrix complex. BioMed Res. Int. 2015, 2015, 212308. [Google Scholar] [CrossRef] [PubMed]
- Alba-Fierro, C.A.; Pérez-Torres, A.; López-Romero, E.; Cuéllar-Cruz, M.; Ruiz-Baca, E. Cell wall proteins of Sporothrix schenckii as immunoprotective agents. Rev. Iberoam. Micol. 2014, 31, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Álvarez, J.A.; Pérez-García, L.A.; Mellado-Mojica, E.; López, M.G.; Martínez-Duncker, I.; Lópes-Bezerra, L.M.; Mora-Montes, H.M. Sporothrix schenckii sensu stricto and Sporothrix brasiliensis Are Differentially Recognized by Human Peripheral Blood Mononuclear Cells. Front. Microbiol. 2017, 8, 843. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; de Hoog, S.; de Camargo, Z.P. Emergence of pathogenicity in the Sporothrix schenckii complex. Med. Mycol. 2013, 51, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; de Hoog, G.S.; de Cássia Pires, D.; Brihante, R.S.N.; da Costa Sidrim, J.J.; Gadelha, M.F.; Colombo, A.L.; de Camargo, Z.P. Genetic diversity and antifungal susceptibility profiles in causative agents of sporotrichosis. BMC Infect. Dis. 2014, 14, 219. [Google Scholar] [CrossRef] [PubMed]
- Camacho, E.; León-Navarro, I.; Rodríguez-Brito, S.; Mendoza, M.; Niño-Vega, G.A. Molecular epidemiology of human sporotrichosis in Venezuela reveals high frequency of Sporothrix globosa. BMC Infect. Dis. 2015, 15, 94. [Google Scholar] [CrossRef] [PubMed]
- Fischman Gompertz, O.; Rodrigues, A.M.; Fernandes, G.F.; Bentubo, H.D.L.; de Camargo, Z.P.; Petri, V. Atypical Clinical Presentation of Sporotrichosis Caused by Sporothrix globosa Resistant to Itraconazole. Am. J. Trop. Med. Hyg. 2016, 94, 1218–1222. [Google Scholar] [CrossRef] [PubMed]
- Alba-Fierro, C.A.; Pérez-Torres, A.; Toriello, C.; Romo-Lozano, Y.; López-Romero, E.; Ruiz-Baca, E. Molecular Components of the Sporothrix schenckii Complex that Induce Immune Response. Curr. Microbiol. 2016, 73, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Carlos, I.Z.; Sassá, M.F.; da Graça Sgarbi, D.B.; Placeres, M.C.P.; Maia, D.C.G. Current research on the immune response to experimental sporotrichosis. Mycopathologia 2009, 168, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gaze, S.T.; Dutra, W.O.; Lessa, M.; Lessa, H.; Guimarães, L.H.; Jesus, A.R.; de Carvalho, L.P.; Machado, P.; Carvalho, E.M.; Gollob, K.J. Mucosal leishmaniasis patients display an activated inflammatory T-cell phenotype associated with a nonbalanced monocyte population. Scand. J. Immunol. 2006, 63, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, V.; Rivera, A. Cytokines and the regulation of fungus-specific CD4 T cell differentiation. Cytokine 2012, 58, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.S.; Gonçalves, A.C.; Portuondo, D.L.; Maia, D.C.G.; Placeres, M.C.P.; Batista-Duharte, A.; Carlos, I.Z. Optimal clearance of Sporothrix schenckii requires an intact Th17 response in a mouse model of systemic infection. Immunobiology 2015, 220, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Quintella, L.P.; Passos, S.R.L.; Francesconi do Vale, A.C.; Galhardo, M.C.G.; Barros, M.B.D.L.; Cuzzi, T.; Dos Santos Reis, R.; de Figueiredo Carvalho, M.H.; Zappa, M.B.; De Oliveira Schubach, A. Histopathology of cutaneous sporotrichosis in Rio de Janeiro: A series of 119 consecutive cases. J. Cutan. Pathol. 2011, 38, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Underhill, D.M.; Pearlman, E. Immune Interactions with Pathogenic and Commensal Fungi: A Two-Way Street. Immunity 2015, 43, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Bär, E.; Whitney, P.G.; Moor, K.; e Sousa, C.R.; LeibundGut-Landmann, S. IL-17 regulates systemic fungal immunity by controlling the functional competence of NK cells. Immunity 2014, 40, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.C.; Maia, D.C.G.; Ferreira, L.S.; Monnazzi, L.G.S.; Alegranci, P.; Placeres, M.C.P.; Batista-Duharte, A.; Carlos, I.Z. Involvement of major components from Sporothrix schenckii cell wall in the caspase-1 activation, nitric oxide and cytokines production during experimental sporotrichosis. Mycopathologia 2015, 179, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D. Dectin-1: A signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Saijo, S.; Iwakura, Y. Dectin-1 and Dectin-2 in innate immunity against fungi. Int. Immunol. 2011, 23, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, C.; Majer, O.; Frohner, I.E.; Tierney, L.; Kuchler, K. Fungal attacks on mammalian hosts: Pathogen elimination requires sensing and tasting. Curr. Opin. Microbiol. 2010, 13, 401–408. [Google Scholar] [CrossRef] [PubMed]
- LeibundGut-Landmann, S.; Wüthrich, M.; Hohl, T.M. Immunity to fungi. Curr. Opin. Immunol. 2012, 24, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Jellmayer, J.A.; Ferreira, L.S.; Manente, F.A.; Gonçalves, A.C.; Polesi, M.C.; Batista-Duharte, A.; Carlos, I.Z. Dectin-1 expression by macrophages and related antifungal mechanisms in a murine model of Sporothrix schenckii sensu stricto systemic infection. Microb. Pathog. 2017, 110, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Koga, T.; Duan, H.; Furue, M. Immunohistochemical detection of interferon-gamma-producing cells in granuloma formation of sporotrichosis. Med. Mycol. 2002, 40, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Koga, T.; Duan, H.; Urabe, K.; Furue, M. Immunohistochemical localization of activated and mature CD83+ dendritic cells in granulomas of sporotrichosis. Eur. J. Dermatol. 2001, 11, 527–529. [Google Scholar] [PubMed]
- Morgado, F.N.; Schubach, A.O.; Pimentel, M.I.; Lyra, M.R.; Vasconcellos, É.C.F.; Valete-Rosalino, C.M.; Conceição-Silva, F. Is There Any Difference between the In Situ and Systemic IL-10 and IFN-γ Production when Clinical Forms of Cutaneous Sporotrichosis Are Compared? PLoS ONE 2016, 11, e0162764. [Google Scholar] [CrossRef] [PubMed]
- Maia, D.C.G.; Sassá, M.F.; Placeres, M.C.P.; Carlos, I.Z. Influence of Th1/Th2 cytokines and nitric oxide in murine systemic infection induced by Sporothrix schenckii. Mycopathologia 2006, 161, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Almeida, S.R. Therapeutic monoclonal antibody for sporotrichosis. Front. Microbiol. 2012, 3, 409. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.L.; Garcia, M.E. Immune response to fungal infections. Vet. Immunol. Immunopathol. 2008, 125, 47–70. [Google Scholar] [CrossRef] [PubMed]
- De Franco, D.L.; Nascimento, R.C.; Ferreira, K.S.; Almeida, S.R. Antibodies against Sporothrix schenckii Enhance TNF-α Production and Killing by Macrophages. Scand. J. Immunol. 2012, 75, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Maia, D.C.G.; Gonçalves, A.C.; Ferreira, L.S.; Manente, F.A.; Portuondo, D.L.; Vellosa, J.C.R.; Polesi, M.C.; Batista-Duharte, A.; Carlos, I.Z. Response of Cytokines and Hydrogen Peroxide to Sporothrix schenckii Exoantigen in Systemic Experimental Infection. Mycopathologia 2016, 181, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.C.; Ferreira, L.S.; Manente, F.A.; de Faria, C.M.Q.G.; Polesi, M.C.; de Andrade, C.R.; Zamboni, D.S.; Carlos, I.Z. The NLRP3 inflammasome contributes to host protection during Sporothrix schenckii infection. Immunology 2017, 151, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Da Daniel Rosa, W.; Gezuele, E.; Calegari, L.; Goñi, F. Asteroid body in sporotrichosis. Yeast viability and biological significance within the host immune response. Med. Mycol. 2008, 46, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; Kubitschek-Barreira, P.H.; Fernandes, G.F.; de Almeida, S.R.; Lopes-Bezerra, L.M.; de Camargo, Z.P. Immunoproteomic analysis reveals a convergent humoral response signature in the Sporothrix schenckii complex. J. Proteom. 2015, 115, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Portuondo, D.L.; Batista-Duharte, A.; Ferreira, L.S.; Martínez, D.T.; Polesi, M.C.; Duarte, R.A.; Marcos, C.M.; de Almeida, A.M.; Carlos, I.Z. A cell wall protein-based vaccine candidate induce protective immune response against Sporothrix schenckii infection. Immunobiology 2016, 221, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Portuondo, D.L.; Batista-Duharte, A.; Ferreira, L.S.; de Andrade, C.R.; Quinello, C.; Téllez-Martínez, D.; de Aguiar Loesch, M.L.; Carlos, I.Z. Comparative efficacy and toxicity of two vaccine candidates against Sporothrix schenckii using either MontanideTM Pet Gel A or aluminum hydroxide adjuvants in mice. Vaccine 2017, 35, 4430–4436. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Jiang, R.; Wang, Y.; Zhu, M.; Zhang, X.; Dong, S.; Shi, H.; Wang, L. Recombinant Phage Elicits Protective Immune Response against Systemic S. globosa Infection in Mouse Model. Sci. Rep. 2017, 7, 42024. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, J.R.F.; Jannuzzi, G.P.; Kaihami, G.H.; Breda, L.C.D.; Ferreira, K.S.; de Almeida, S.R. An immunoproteomic approach revealing peptides from Sporothrix brasiliensis that induce a cellular immune response in subcutaneous sporotrichosis. Sci. Rep. 2018, 8, 4192. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.S.; Coelho, A.L.; Lopes Bezerra, L.M.; Barja-Fidalgo, C. Virulence of Sporothrix schenckii conidia and yeast cells, and their susceptibility to nitric oxide. Immunology 2000, 101, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.S.S.; Neto, E.H.; Brito, M.M.S.; Silva, J.S.; Cunha, F.Q.; Barja-Fidalgo, C. Detrimental role of endogenous nitric oxide in host defence against Sporothrix schenckii. Immunology 2008, 123, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Alegranci, P.; de Abreu Ribeiro, L.C.; Ferreira, L.S.; de Negrini, T.C.; Maia, D.C.G.; Tansini, A.; Gonçalves, A.C.; Placeres, M.C.; Carlos, I.Z. The predominance of alternatively activated macrophages following challenge with cell wall peptide-polysaccharide after prior infection with Sporothrix schenckii. Mycopathologia 2013, 176, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Madrid, I.M.; Xavier, M.O.; Mattei, A.S.; Fernandes, C.G.; Guim, T.N.; Santin, R.; Schuch, L.F.; de Nobre, M.O.; Araújo Meireles, M.C. Role of melanin in the pathogenesis of cutaneous sporotrichosis. Microbes Infect. 2010, 12, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.F.; Bonfitto, M.; da Silva Junior, F.I.M.; de Ameida, M.T.G.; da Silva, R.D.C. Sporotrichosis in a liver transplant patient: A case report and literature review. Med. Mycol. Case Rep. 2017, 17, 25–27. [Google Scholar] [CrossRef] [PubMed]
- Lederer, H.T.; Sullivan, E.; Crum-Cianflone, N.F. Sporotrichosis as an unusual case of osteomyelitis: A case report and review of the literature. Med. Mycol. Case Rep. 2016, 11, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.P.; Lau, Y.-L. Cellular and Molecular Defects Underlying Invasive Fungal Infections-Revelations from Endemic Mycoses. Front. Immunol. 2017, 8, 735. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Paes, R.; Figueiredo-Carvalho, M.H.G.; Brito-Santos, F.; Almeida-Silva, F.; Oliveira, M.M.E.; Zancopé-Oliveira, R.M. Melanins Protect Sporothrix brasiliensis and Sporothrix schenckii from the Antifungal Effects of Terbinafine. PLoS ONE 2016, 11, e0152796. [Google Scholar] [CrossRef] [PubMed]
- Batista-Duharte, A.; Téllez-Martínez, D.; Aparecida Jellmayer, J.; Leandro Portuondo Fuentes, D.; Campos Polesi, M.; Martins Baviera, A.; Zeppone Carlos, I. Repeated Exposition to Mercury (II) Chloride Enhances Susceptibility to S. schenckii sensu stricto Infection in Mice. J. Fungi 2018, 4, 64. [Google Scholar] [CrossRef] [PubMed]
- De Lima Barros, M.B.; Schubach, A.; Francesconi-do-Valle, A.C.; Gutierrez-Galhardo, M.C.; Schubach, T.M.P.; Conceição-Silva, F.; de Matos Salgueiro, M.; Mouta-Confort, E.; Reis, R.S.; de Fátima Madeira, M.; et al. Positive Montenegro skin test among patients with sporotrichosis in Rio De Janeiro. Acta Trop. 2005, 93, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.G.; Abraham, R.M.; Mihova, D.; Xu, X.; Frank, D.M.; Rosenbach, M.; Kim, E.J. Acute onset of leg nodules in a sporotrichoid pattern—Quiz case. Diagnosis: Primary cutaneous diffuse large B-cell lymphoma, leg type (PCLBCL-LT). Arch. Dermatol. 2012, 148, 1199–1200. [Google Scholar] [CrossRef] [PubMed]
- Lauermann, F.; Lyra, M.; Gaudio, R. Sporotrichosis mimicking keratoacanthoma. Am. J. Trop. Med. Hyg. 2012, 86, 741. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Hashimoto, Y.; Nishi, K.; Takahashi, H.; Takeda, K.; Mizumoto, T.; Iizuka, H. Cutaneous tuberculosis simulating lymphocutaneous sporotrichosis. Australas. J. Dermatol. 2012, 53, 316–317. [Google Scholar] [CrossRef] [PubMed]
- Quintella, L.P.; Passos, S.R.L.; de Miranda, L.H.M.; Cuzzi, T.; Barros, M.D.; Francesconi-do-Vale, A.C.; Galhardo, M.C.; Madeira, M.D.; de Figueiredo Carvalho, M.H.; Schubach, A.D. Proposal of a histopathological predictive rule for the differential diagnosis between American tegumentary leishmaniasis and sporotrichosis skin lesions. Br. J. Dermatol. 2012, 167, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Aung, A.K.; Teh, B.M.; McGrath, C.; Thompson, P.J. Pulmonary sporotrichosis: Case series and systematic analysis of literature on clinico-radiological patterns and management outcomes. Med. Mycol. 2013, 51, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Di Luca, D.G.; De Andrade, P.J.S.; Sales, A.M.; De Menezes, V.M.; Galhardo, M.C.G.; Pimentel, M.I.F.; Lyra, M.R.; Nery, J.A. Superposition of leprosy and other neglected tropical diseases in the state of Rio de Janeiro: A case series report. Lepr. Rev. 2013, 84, 302–307. [Google Scholar] [PubMed]
- Kawtar, I.; Salim, G.; Mariame, M.; Fatimazahra, M.; Imane, T.; Salma, B.; Imane, E.H.; Mohamed, E. Sporotrichoid chromomycosis. Dermatol. Online J. 2013, 19, 20394. [Google Scholar] [PubMed]
- Zhang, Y.; Pyla, V. Nasal sporotrichosis in children. Int. J. Dermatol. 2014, 53, e342–e343. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Clinical Presentation | Main Features | Some Reports |
|---|---|---|
| Fixed | Single nodule, ulcer, or verrucous lesion without signs of lymphangitis | [11,12,47,53,55,56,57,58,59] |
| Lymphocutaneous | Multiple nodules, ulcers, and signs of lymphangitis following the path of lymphatic draining | [11,12,47,53,55,56,57,58,59] |
| Mucosal | Ulcers, granulomatous infiltration, serous-purulent discharge, crusts in conjunctiva, nasal and/or oral mucosa | [11,12,47,53,55,56,57] |
| Cutaneous disseminated in immunocompetent or immunocompromised patients | Multiple nodules and/or ulcers in different non-contiguous parts of the body | [11,12,47,53,55,56,57,58,59] |
| Extracutaneous in immunocompetent or immunocompromised patients | Osteoarticular, pulmonary, meningitis, exogenous and endogenous endophthalmitis etc. | [22,24,30,57,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76] |
| Reactional cases | Erythema multiforme, erythema nodosum | [30,77,78] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conceição-Silva, F.; Morgado, F.N. Immunopathogenesis of Human Sporotrichosis: What We Already Know. J. Fungi 2018, 4, 89. https://doi.org/10.3390/jof4030089
Conceição-Silva F, Morgado FN. Immunopathogenesis of Human Sporotrichosis: What We Already Know. Journal of Fungi. 2018; 4(3):89. https://doi.org/10.3390/jof4030089
Chicago/Turabian StyleConceição-Silva, Fatima, and Fernanda Nazaré Morgado. 2018. "Immunopathogenesis of Human Sporotrichosis: What We Already Know" Journal of Fungi 4, no. 3: 89. https://doi.org/10.3390/jof4030089
APA StyleConceição-Silva, F., & Morgado, F. N. (2018). Immunopathogenesis of Human Sporotrichosis: What We Already Know. Journal of Fungi, 4(3), 89. https://doi.org/10.3390/jof4030089