
Major Intrinsic Proteins in Fungi: A Special Emphasis on the XIP Subfamily

 , and
, and
Abstract
1. General Considerations on MIPs
2. MIP Fungal Diversity and Function
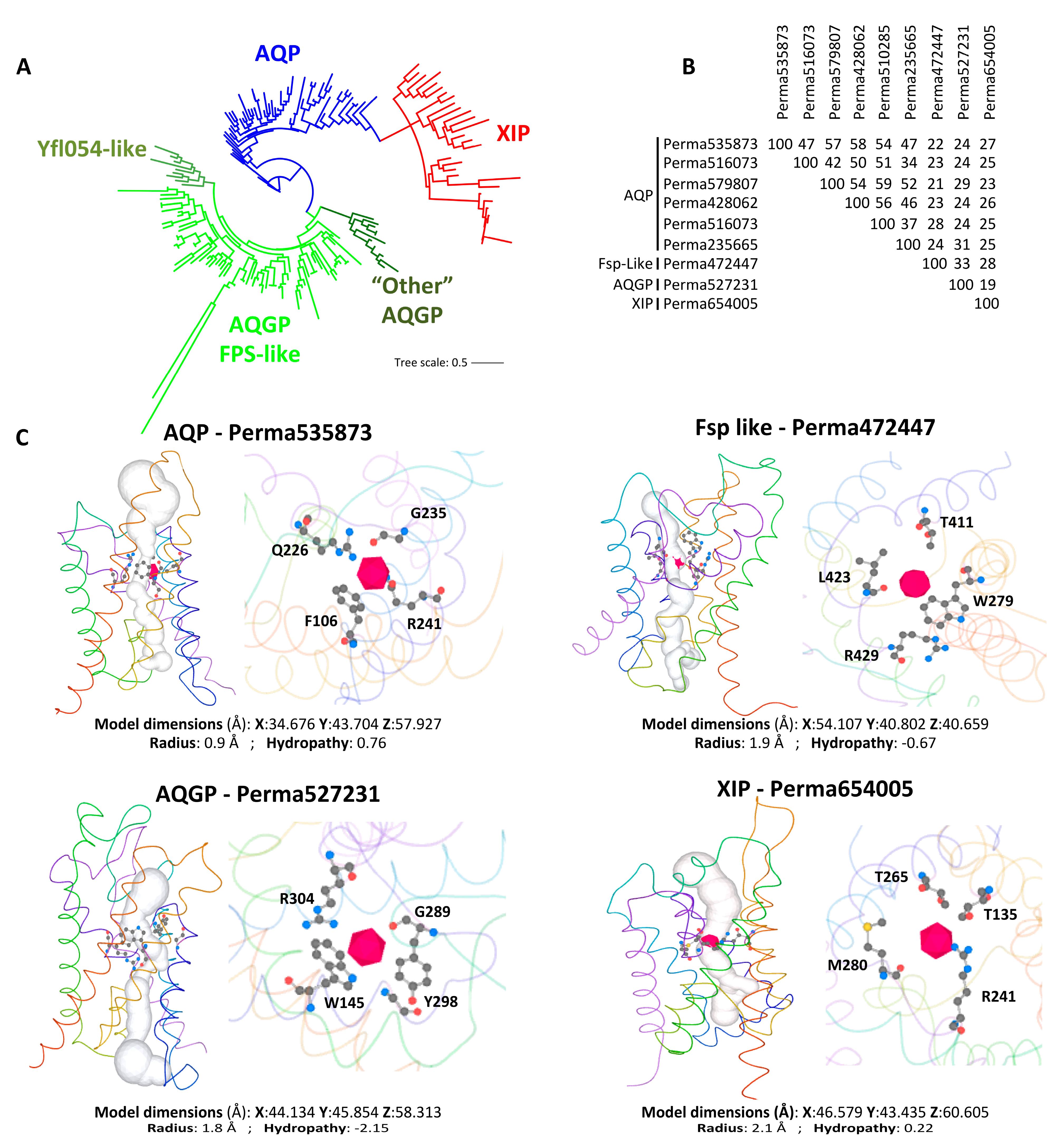
3. Case Study of Fungal XIPs—State of the Art
3.1. State of the Art
3.2. Diversity of the Fungal XIPs—Evolution and Topology
- (1)
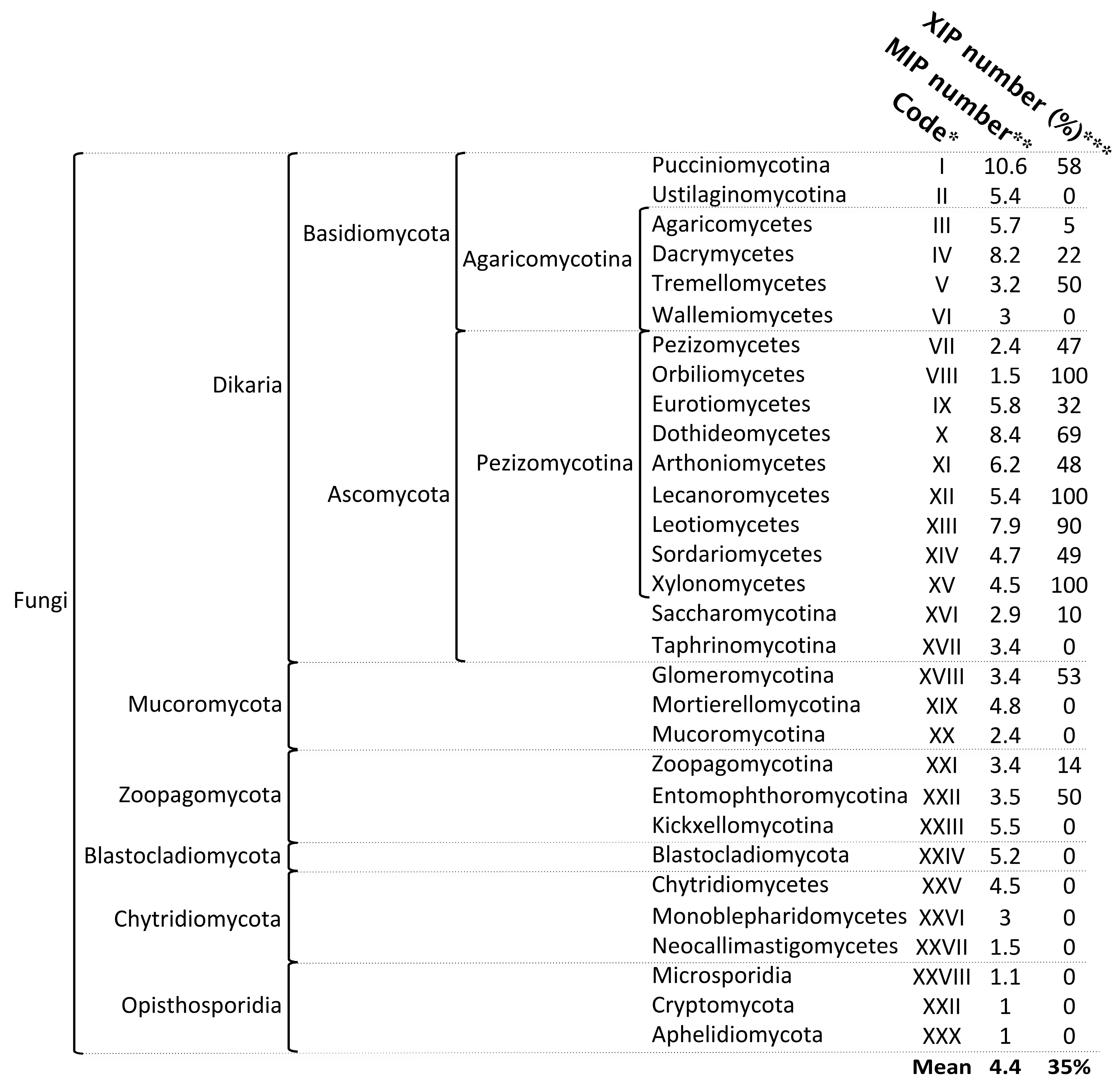
- According to currently available sequenced genomes, XIPs appear to be absent from several fungal lineages. Specifically, no XIP sequences were identified in the Kickxellomycotina (XXIII), Mucoromycotina (XX), Mortierellomycotina (XIX), Taphrinomycotina (XVII), Wallmiomycetes (VI), and Ustilaginomycotina (II) (Figure 2). Similarly, XIPs are entirely absent from all lineages within the Opisthosporidia subdivision (including Aphelidiomycota (XXX), Cryptomycota (XXIX), and Microsporidia (XXVIII)), as well as from all subdivisions of the Chytridiomycota (XXV to XXVII) and the Blastocladiomycotina (XXIV). These basal clades, generally regarded as the earliest-diverging lineages of the fungal kingdom, are predominantly composed of unicellular organisms and are characterized by the production of both motile and non-motile sporangiospores [62,63].
- (2)
- In their review of fungal AQPs, Nehls and Dietz [35] reported the presence of a putative XIP sequence in a species belonging to the phylum Microsporidia. Although the specific origin of this sequence remains unspecified, its mention is nonetheless significant as it highlights the limitations inherent to bioinformatic approaches commonly used to characterize specific subfamilies. Unless this observation stems from an annotation error or the inadvertent incorporation of foreign genetic material into public databases—an event we have not yet been able to confirm—our analyses suggest that Microsporidia do not possess any genuine XIP members (Figure S2 and Table S1). tBLASTn searches and keyword-based queries conducted in both the JGI and NCBI databases consistently retrieve aquaporin sequences that are not related to the XIP subfamily. Furthermore, the protein translations of these candidate sequences exhibit very low sequence identity with known XIPs (approximately 22%), and phylogenetic analyses—including XIP sequences from both closely and distantly related fungal species—do not support the hypothesis that Microsporidia harbor members of this aquaporin subfamily. This conclusion appears to extend to all members of the Chytridiomycetes and Opisthosporidia as well. Continued efforts to identify new species of basal fungi, particularly those inhabiting underexploited environments, combined with expanded genome sequencing initiatives, will be crucial for enriching and potentially refining our understanding of aquaporin diversity and evolution within these early-diverging fungal lineages.
- (3)
- The abundance of XIPs is markedly lower compared with that of the AQP and AQGP subfamilies. In the vast majority of fungal species, the XIP subfamily is represented by a single gene copy, a trend particularly evident in six clades: three within Agaricomycotina (II, IV, and V), as well as Orbiliomycetes (VIII), Arthoniomycetes (XI), and Saccharomycotina (XVI). This limited diversity among fungal XIPs is reminiscent of the low number of XIP members observed in plants, as previously reported by Verma et al. [23], and may reflect evolutionary constraints or functional specificity that limit the expansion of this subfamily.
- (4)
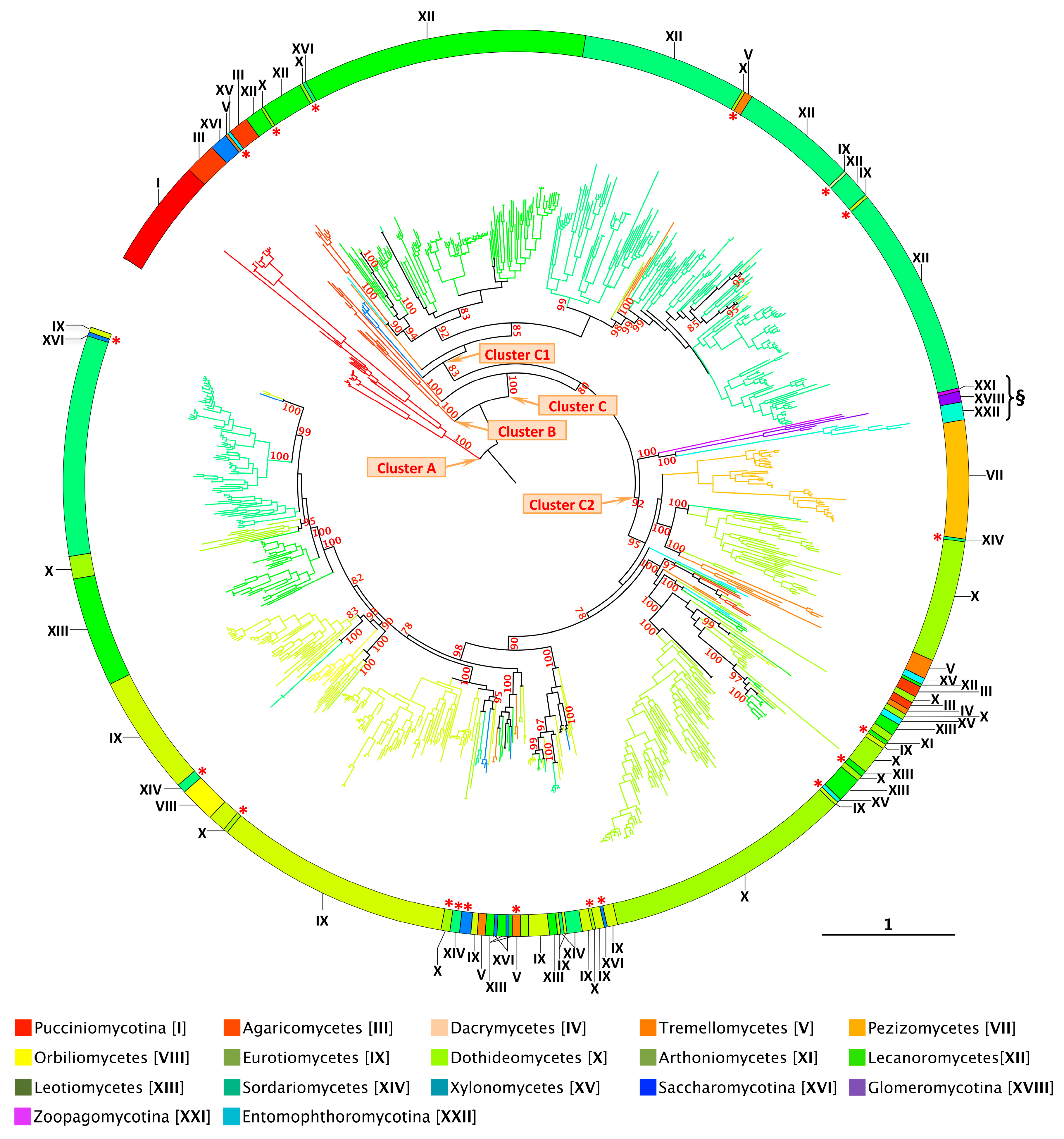
- A striking concordance emerges between the phylogenetic distribution of XIPs and the currently established fungal phylogeny, suggesting an evolutionary trajectory in which most XIP members have diversified in a lineage-specific manner across multiple distinct fungal clades (Figure 4 and Figure S3). This distribution pattern parallels the divergence of major fungal phyla, implying that XIPs may have undergone multiple and independent expansion events throughout fungal evolutionary history. Notably, such lineage-specific expansions of XIP genes in fungi closely mirror those documented in plants [19,54], further supporting the hypothesis of convergent evolutionary strategies among eukaryotes adapting to diverse environmental niches.
- (5)
- In contrast to AQPs and AQGPs members, XIPs display an uneven distribution across fungal clades, a pattern that persists even at finer taxonomic resolutions. For example, the class Sordariomycetes (XIV) contains the highest number of identified XIPs, totaling 271 sequences. However, these sequences are not uniformly distributed across all subclasses. XIP members have been identified in several orders belonging to four subclasses: Diaporthomycetidae, Sordariomycetidae, Xylariomycetidae, and Hypocreomycetidae (Figures S4 and S5). To date, no XIP sequences have been detected within the subclasses Savoryellomycetidae and Lulworthiomycetidae. This observation highlights the uneven retention of XIPs across fungal lineages and raises questions about the evolutionary and ecological factors underlying the differential presence of this subfamily among closely related taxonomic groups.
- (6)
- Fungal XIPs, based on their nucleotide sequences, are phylogenetically grouped into three well-supported clades (Figure 4 and Figure S3). The first (Cluster A) consists exclusively of members of the Pucciniomycotina (I); the second (Cluster B) comprises a subset of divergent sequences from the Agaricomycetes (III); and the third (Cluster C), which encompasses 96% of the retrieved members, forms the largest group. Notably, this major clade is sub-partitioned into two distinct branches, hereafter referred to as Clusters C1 and C2. This bipartite structure, described here for the first time, is consistently observed in fungal species that harbor multiple XIP paralogs. Despite a relatively low average sequence similarity across the four clusters, approximately 39%, all proteins within both clusters retain conserved molecular signatures characteristic of the XIP family [25,51]. This phylogenetic organization underscores an unexpected level of diversity and divergence within the fungal XIP subfamily. Importantly, the taxonomic distribution across the two clusters is markedly uneven. Cluster C2 contains the most divergent sequences, spanning a broad range of fungal phyla, from basal lineages (XVIII, XXI, and XXII) to more recently evolved groups such as the Dikarya. In contrast, Cluster C1 is restricted to the Dikarya and includes representatives from only a subset of its constituent phyla. Within the Dikarya, certain major phyla are represented exclusively in one cluster: for example, Pucciniomycotina, Pezizomycetes, and Lecanoromycetes in Cluster C2 and Dacrymycetes, Orbiliomycetes, and Leotiomycetes in Cluster C1. Intriguingly, some sequences (highlighted with an asterisk in Figure 4) do not cluster with their expected fungal clades but instead appear among distantly related fungal groups. This applies to several phyla (e.g., Eurotiomycetes (IX), Dothideomycetes (X), Sordariomycetes (XIV), and Leotiomycetes (XIII)); however, this pattern is particularly noticeable in Tremellomycetes (V), which fails to form a distinct cluster. These atypical phylogenetic placements suggest plausible horizontal gene transfer (HGT) between fungal species, a phenomenon previously reported for other fungal proteins [64,65]. HGT, coupled with the stable integration of the transferred genetic material, may confer adaptive advantages to the host, including enhanced environmental responsiveness and the acquisition of novel traits and functions [66,67]. This hypothesis warrants further dedicated investigation.
- (7)
- The amino acid length of fungal XIPs ranges from 274 to 362 residues, with an average of 326, corresponding to theoretical molecular weights between 29.17 and 38.82 kDa (mean: 34.97) (Table 1). The isoelectric points (pI) of these proteins span a range from 6.41 to 8.22, suggesting functional constraints potentially linked to regulatory mechanisms. These include conserved regulatory motifs in the C-terminal regions and in several electrostatic loops, which may be involved in post-translational modifications such as methylation, phosphorylation, and trafficking, as well as interactions with regulatory partners. Such modifications, known to vary among MIP families and subfamilies, underscore the multifunctional and dynamic regulation of solute permeability mediated by AQPs; this phenomenon is documented in plants and animals [68,69] but still insufficiently characterized in filamentous fungi. Additionally, XIPs exhibit favorable biochemical properties, including high intrinsic stability (instability index < 40), and thermal resilience, as indicated by an average aliphatic index of 95. These features are characteristic of canonical MIP protomers. In silico predictions of subcellular localization predominantly place XIPs in the plasma membranes; however, a marginal presence in other cellular compartments—such as the mitochondria, endoplasmic reticulum, and peroxisomes—cannot be excluded. In this context, the positive GRAVY (grand average of hydropathy) index of 0.389 further supports their hydrophobic nature, a fundamental trait for aquaporins that facilitates efficient transmembrane transport of solutes [70].
- (8)
- As expected for members of the MIP superfamily, the protomeric architecture of all fungal XIPs is highly conserved and adopts the canonical hourglass configuration. This structure comprises six transmembrane helices (TMH1 to TMH6), connected by five inter-helical loops (Loop A to E), with both the N- and C-terminal regions oriented toward the cytosolic side (Figure 3C and Table 1). Centrally located within the pore are two half-helices, derived from loops B and E, each capped by conserved “NPA” motifs, along with the ar/R selectivity filter. The latter is composed of specific hydrophobic amino acids whose side chains line the pore and form a narrow constriction at the extracellular entrance. Although this ar/R filter appears to be somewhat less hydrophobic than its counterparts in dicotyledonous plants and moss [51], it is likely optimized for the transmembrane passage of relatively large and/or hydrophobic solutes, such as glycerol, urea, ammonia (NH3), lactic and boric acids, polyols, and various metalloids or ions including arsenic (As), antimony (Sb), zinc (Zn2+), aluminum (Al3+), silicon (Si), and selenium (Se). Notably, there is considerable variability in the amino acid residues lining the channel both between and within fungal phyla, which may reflect adaptations to distinct physiological environments and suggest divergent substrate specificities of XIPs across fungal lineages.
3.3. Fungal XIPs: A Relatively Recent Chapter in the Evolution of Fungal AQP
3.4. Fungal XIP—Outstanding Questions
- -
- Do the expansion and relative conservation of the XIP subgroup within the Dikarya, together with its apparent correlation to the terrestrialization of the fungal kingdom, point to a specialized function for these proteins in transporting solutes essential to the full developmental cycle of these fungi?
- -
- Could the concomitant presence of XIP homologs in both fungal and green plant lineages be associated with their parallel evolutionary success of in terrestrial ecosystems? This prompts broader questions regarding the potential roles of XIPs as an evolutionary link between these two kingdoms, whose ecological interdependence is both diverse and deep. Despite their distinct phylogenetic classifications, remarkably, XIP proteins in plants and fungi share striking structural similarities, hinting at a plausible shared ancestral origin.
- -
- Do the two divergent XIP archetypes observed across fungal species reflect a degree of functional specialization, potentially involving distinct solute transport profiles and regulatory mechanisms, or are they associated with tissue-specific expression patterns in multicellular organisms, indicative of sub-functionalization?
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agre, P.; Sasaki, S.; Chrispeels, M.J. Aquaporins: A family of water channel proteins. Am. J. Phys. 1993, 265, F461. [Google Scholar] [CrossRef] [PubMed]
- Zardoya, R. Phylogeny and evolution of the major intrinsic protein family. Biol. Cell 2005, 97, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Gazzarrini, S.; Kang, M.; Epimashko, S.; Van Etten, J.L.; Dainty, J.; Thiel, G.; Moroni, A. Chlorella virus MT325 encodes water and potassium channels that interact synergistically. Proc. Natl. Acad. Sci. USA 2006, 103, 5355–5360. [Google Scholar] [CrossRef] [PubMed]
- Fujiyoshi, Y.; Mitsuoka, K.; de Groot, B.L.; Philippsen, A.; Grubmüller, H.; Agre, P.; Engel, A. Structure and function of water channels. Curr. Opin. Struct. Biol. 2002, 12, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Ozu, M.; Galizia, L.; Acuña, C.; Amodeo, G. Aquaporins: More Than Functional Monomers in a Tetrameric Arrangement. Cells 2018, 7, 209. [Google Scholar] [CrossRef] [PubMed]
- Luu, D.T.; Maurel, C. Aquaporins in a challenging environment: Molecular gears for adjusting plant water status. Plant Cell Environ. 2005, 28, 85–96. [Google Scholar] [CrossRef]
- Tajkhorshid, E.; Nollert, P.; Jensen, M.; Miercke, L.; O’Connell, J.; Stroud, R.M.; Schulten, K. Control of the selectivity of the aquaporin water channel family by global orientation tuning. Science 2002, 296, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Sui, H.; Han, B.G.; Lee, J.K.; Walian, P.; Jap, B.K. Structural basis of water specific transport through the AQP1 water channel. Nature 2001, 414, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Wallace, I.S.; Roberts, D.M. Homology modeling of representative subfamilies of Arabidopsis major intrinsic proteins. Classification based on the aromatic/arginine selectivity filter. Plant Physiol. 2004, 135, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Libson, A.; Miercke, L.J.; Weitzman, C.; Nollert, P.; Krucinski, J.; Stroud, R.M. Structure of a glycerol-conducting channel and the basis for its selectivity. Science 2000, 290, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Schulten, K.; Tajkhorshid, E. What makes an aquaporin a glycerol channel? A comparative study of AqpZ and GlpF. Structure 2005, 13, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Beitz, E. Aquaporins with selectivity for unconventional permeants. Cell. Mol. Life Sci. 2007, 64, 2413–2421. [Google Scholar] [CrossRef] [PubMed]
- Saboe, P.O.; Rapisarda, C.; Kaptan, S.; Hsiao, Y.S.; Summers, S.R.; De Zorzi, R.; Dukovski, D.; Yu, J.; de Groot, B.L.; Kumar, M.; et al. Role of Pore-Lining Residues in Defining the Rate of Water Conduction by Aquaporin-0. Biophys. J. 2017, 112, 953–965. [Google Scholar] [CrossRef] [PubMed]
- Pluhackova, K.; Schittny, V.; Bürkner, P.C.; Siligan, C.; Horner, A. Multiple pore lining residues modulate water permeability of GlpF. Protein Sci. 2022, 31, e4431. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.N.; Cerdà, J. Evolution and functional diversity of aquaporins. Biol. Bull. 2015, 229, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Johanson, U.; Karlsson, M.; Johansson, I.; Gustavsson, S.; Sjovall, S.; Fraysse, L.; Weig, A.R.; Kjellbom, P. The complete set of genes encoding major intrinsic proteins in Arabidopsis provides a framework for a new nomenclature for major intrinsic proteins in plants. Plant Physiol. 2001, 126, 358–1369. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C. Plant aquaporins: Novel functions and regulation properties. FEBS Lett. 2007, 581, 2227–2236. [Google Scholar] [CrossRef] [PubMed]
- Danielson, J.A.H.; Johanson, U. Unexpected complexity of the aquaporin gene family in the moss Physcomitrella patens. BMC Plant Biol. 2008, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Lopez, D.; Bronner, G.; Brunel, N.; Auguin, D.; Bourgerie, S.; Brignolas, F.; Carpin, S.; Tournaire-Roux, C.; Maurel, C.; Fumanal, B.; et al. Insights into Populus XIP aquaporins: Evolutionary expansion, protein functionality, and environmental regulation. J. Exp. Bot. 2012, 65, 2217–2230. [Google Scholar] [CrossRef] [PubMed]
- Anderberg, H.I.; Kjellbom, P.; Johanson, U. Annotation of Selaginella moellendorffii major intrinsic proteins and the evolution of the protein family in terrestrial plants. Front. Plant Sci. 2012, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Khabudaev, K.V.; Petrova, D.P.; Grachev, M.A.; Likhoshway, Y.V. A new subfamily LIP of the major intrinsic proteins. BMC Genom. 2014, 15, 173. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Irisarri, I.; Zardoya, R. Diversity and evolution of membrane intrinsic proteins. Biochim. Biophys. Acta 2014, 1840, 1468–1481. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.K.; Prabh, N.D.; Sankararamakrishnan, R. New subfamilies of major intrinsic proteins in fungi suggest novel transport properties in fungal channels: Implications for the host-fungal interactions. BMC Ecol. Evol. 2014, 14, 173. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Cooke, J.E.K.; Zwiazek, J.J. Phylogenetic analysis of fungal aquaporins provide insight into their possible role in water transport of mycorrhizal associations. Botany 2013, 91, 495–504. [Google Scholar] [CrossRef]
- Ben Amira, M.; Mom, R.; Lopez, D.; Chaar, H.; Khouaja, A.; Pujade-Renaud, V.; Fumanal, B.; Gousset-Dupont, A.; Bronner, G.; Label, P.; et al. MIP diversity from Trichoderma: Structural considerations and transcriptional modulation during mycoparasitic association with Fusarium solani olive trees. PLoS ONE 2018, 13, e0193760. [Google Scholar] [CrossRef] [PubMed]
- Purvis, A.; Hector, A. Getting the measure of biodiversity. Nature 2000, 405, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L.; Luecking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectrum. 2017, 5, FUNK-0052-2016. [Google Scholar] [CrossRef] [PubMed]
- James, T.Y.; Stajich, J.E.; Hittinger, C.T.; Rokas, A. Toward a Fully Resolved Fungal Tree of Life. Annu. Rev. Microbiol. 2020, 74, 291–313. [Google Scholar] [CrossRef] [PubMed]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Baldrian, P.; Větrovský, T.; Lepinay, C.; Kohout, P. High-throughput sequencing view on the magnitude of global fungal diversity. Fungal Divers. 2021, 114, 539–547. [Google Scholar] [CrossRef]
- Li, Y.; Steenwyk, J.L.; Chang, Y.; Wang, Y.; James, T.Y.; Stajich, J.E.; Spatafora, J.W.; Groenewald, M.; Dunn, C.W.; Hittinger, C.T.; et al. A genome-scale phylogeny of the kingdom Fungi. Curr. Biol. 2021, 31, 1653–1665. [Google Scholar] [CrossRef] [PubMed]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, N.; Filipsson, C.; Becit, E.; Brive, L.; Hohmann, S. Aquaporins in yeasts and filamentous fungi. Biol. Cell 2005, 97, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Dietz, S.; Bülow, J.V.; Beitz, E.; Nehls, U. The aquaporin gene family of the ectomycorrhizal fungus Laccaria bicolor: Lessons for symbiotic functions. New Phytol. 2011, 190, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Nehls, U.; Dietz, S. Fungal aquaporins: Cellular functions and ecophysiological perspectives. Appl. Microbiol. Biotechnol. 2014, 98, 8835–8851. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zwiazek, J.J. Fungal Aquaporins in Ectomycorrhizal Root Water Transport. Front. Plant Sci. 2020, 11, 302. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Kun, J.F.; Schultz, J.E.; Beitz, E. A single, bi-functional aquaglyceroporin in blood-stage Plasmodium falciparum malaria parasites. J. Biol. Chem. 2002, 277, 4874–4882. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic-Djuranovic, S.; Schultz, J.E.; Beitz, E. A single aquaporin gene encodes a water/glycerol/urea facilitator in Toxoplasma gondii with similarity to plant tonoplast intrinsic proteins. FEBS Lett. 2003, 555, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Laize, V.; Tacnet, F.; Ripoche, P.; Hohmann, S. Polymorphism of Saccharomyces cerevisiae aquaporins. Yeast 2000, 16, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Sidoux-Walter, F.; Pettersson, N.; Hohmann, S. The Saccharomyces cerevisiae aquaporin Aqy1 is involved in sporulation. Proc. Natl. Acad. Sci. USA 2004, 101, 17422–17427. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, N.; Hagstrom, J.; Bill, R.M.; Hohmann, S. Expression of heterologous aquaporins for functional analysis in Saccharomyces cerevisiae. Curr. Genet. 2006, 50, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Xu, H. Major Intrinsic Proteins of Laccaria Bicolor: Characterization, Transcript Profiling and Functions in Ectomycorrhizal Associations with Picea glauca. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 2015; p. 99. [Google Scholar]
- Navarro-Ródenas, A.; Ruíz-Lozano, J.M.; Kaldenhoff, R.; Morte, A. The aquaporin TcAQP1 of the desert truffle Terfezia claveryi is a membrane pore for water and CO2 transport. Mol. Plant-Microbe Interact. 2012, 25, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Ródenas, A.; Bárzana, G.; Nicolás, E.; Carra, A.; Schubert, A.; Morte, A. Expression analysis of aquaporins from desert truffle mycorrhizal symbiosis reveals a fine-tuned regulation under drought. Mol. Plant-Microbe Interact. 2013, 26, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Tisserant, E.; Brun, A.; Lege, V.; Martin, F.; Kohler, A. Laser microdissection and microarray analysis of Tuber melanosporum ectomycorrhizas reveal functional heterogeneity between mantle and Hartig net compartments. Environ. Microbiol. 2013, 15, 1853–1869. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Hu, Y.J.; Hao, Z.P.; Li, H.; Wang, Y.S.; Chen, B.D. First cloning and characterization of two functional aquaporin genes from an arbuscular mycorrhizal fungus Glomus intraradices. New Phytol. 2013, 197, 617–630. [Google Scholar] [CrossRef] [PubMed]
- An, B.; Li, B.; Li, H.; Zhang, Z.; Qin, G.; Tian, S. Aquaporin-8 regulates cellular development and reactive oxygen species production, a critical component of virulence in Botrytis cinerea. New Phytol. 2016, 209, 1668–1680. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.; Kohler, A.; Ohm, R.A.; Kuo, A.; Krützmann, J.; Morin, E.; Arend, M.; Barry, K.W.; Binder, M.; Choi, C.; et al. Ectomycorrhizal ecology is imprinted in the genome of the dominant symbiotic fungus Cenococcum geophilum. Nat. Commun. 2016, 7, 12662. [Google Scholar] [CrossRef] [PubMed]
- Ben Amira, M.; Faize, M.; Karlsson, M.; Dubey, M.; Frąc, M.; Panek, J.; Fumanal, B.; Gousset-Dupont, A.; Julien, J.-L.; Chaar, H.; et al. Fungal X-Intrinsic Protein aquaporin from Trichoderma atroviride: Structural and functional considerations. Biomolecules 2021, 11, 338. [Google Scholar] [CrossRef] [PubMed]
- Laothanachareon, T.; Asin-Garcia, E.; Volkers, R.J.M.; Tamayo-Ramos, J.A.; Martins dos Santos, V.A.P.; Schaap, P.J. Identification of Aspergillus niger Aquaporins Involved in Hydrogen Peroxide Signaling. J. Fungi 2023, 9, 499. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.B.; Sankararamakrishnan, R. Genome-wide analysis of major intrinsic proteins in the tree plant Populus trichocarpa: Characterization of XIP subfamily of aquaporins from evolutionary perspective. BMC Plant Biol. 2009, 9, 134. [Google Scholar] [CrossRef] [PubMed]
- Laur, J.; Hacke, U.G. Exploring Picea glauca aquaporins in the context of needle water uptake and xylem refilling. New Phytol. 2014, 203, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Diehn, T.A.; Pommerrenig, B.; Bernhardt, N.; Hartmann, A.; Bienert, G.P. Genome-wide identification of aquaporin encoding genes in Brassica oleracea and their phylogenetic sequence comparison to Brassica crops and Arabidopsis. Front. Plant Sci. 2015, 6, 166. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, J.; Yu, J.W.; Gaston, D.; Park, S.W. Molecular evolution and functional divergence of X-intrinsic protein genes in plants. Mol. Genet. Genom. 2015, 290, 443–460. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in Plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Bienert, M.D.; Jahn, T.P.; Boutry, M.; Chaumont, F. Solanaceae XIPs are plasma membrane aquaporins that facilitate the transport of many uncharged substrates. Plant J. 2011, 66, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Ali, Z.; Wang, C.B.; Xu, L.; Yi, J.X.; Xu, Z.L.; Liu, X.Q.; He, X.L.; Huang, Y.H.; Khan, I.A.; et al. Genome-wide sequence characterization and expression analysis of major intrinsic proteins in Soybean (Glycine max L.). PLoS ONE 2013, 8, e56312. [Google Scholar] [CrossRef] [PubMed]
- Reuscher, S.; Akiyama, M.; Mori, C.; Aoki, K.; Shibata, D.; Shiratake, K. Genome-wide identification and expression analysis of aquaporins in tomato. PLoS ONE 2013, 8, e79052R. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, J.; Yu, J.W.; Park, S.W. Genome-wide analysis and expression profiling of the Solanum tuberosum aquaporins. Plant Physiol. Biochem. 2013, 73, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Cao, H.; Wang, L.; Zhou, Y.; Hao, X.; Zeng, J.; Wang, X.; Yang, Y. Molecular cloning and expression analysis of tea plant aquaporin (AQP) gene family. Plant Physiol. Biochem. 2014, 83, 65–76. [Google Scholar] [CrossRef] [PubMed]
- de Paula Santos Martins, C.; Pedrosa, A.M.; Du, D.; Gonçalves, L.P.; Yu, Q.; Gmitter, F.G., Jr.; Costa, M.G.C.; Fang, D.D. Genome-Wide Characterization and Expression Analysis of Major Intrinsic Proteins during Abiotic and Biotic Stresses in Sweet Orange (Citrus sinensis L. Osb.). PLoS ONE 2015, 10, e0138786. [Google Scholar] [CrossRef] [PubMed]
- Berbee, M.L.; James, T.Y.; Strullu-Derrien, C. Early diverging fungi: Diversity and impact at the dawn of terrestrial life. Annu. Rev. Microbiol. 2017, 71, 41–60. [Google Scholar] [CrossRef] [PubMed]
- Voigt, K.; James, T.Y.; Kirk, P.M.; Santiago, A.L.C.M.d.A.; Waldman, B.; Griffith, G.W.; Fu, M.; Radek, R.; Strassert, J.F.H.; Wurzbacher, C.; et al. Early-diverging fungal phyla: Taxonomy, species concept, ecology, distribution, anthropogenic impact, and novel phylogenetic proposals. Fungal Divers. 2021, 109, 59–98. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, V.D.A.; Sukno, S.A.; Thon, M.R. Identification of horizontally transferred genes in the genus Colletotrichum reveals a steady tempo of bacterial to fungal gene transfer. BMC Genom. 2015, 16, 2. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, S.; Ge, W.; Zhao, L.; Hou, B.; Wang, K.; Lyu, Z.; Chen, L.; Xu, S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Bae, H. Horizontal Gene Transfer and Endophytes: An Implication for the Acquisition of Novel Traits. Plants 2020, 9, 305. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, S.; Cheewangkoon, R.; Zhao, R. Uneven distribution of prokaryote-derived horizontal gene transfer in fungi: A lifestyle-dependent phenomenon. mBio 2025, 16, e02855-24. [Google Scholar] [CrossRef] [PubMed]
- Chaumont, F.; Moshelion, M.; Daniels, M.J. Regulation of plant aquaporin activity. Biol. Cell 2005, 97, 749–764. [Google Scholar] [CrossRef] [PubMed]
- Day, R.E.; Kitchen, P.; Owen, D.S.; Bland, C.; Marshall, L.; Conner, A.C.; Bill, R.M.; Conner, M.T. Human aquaporins: Regulators of transcellular water flow. Biochim. Biophys. Acta 2014, 1840, 1492–1506. [Google Scholar] [CrossRef] [PubMed]
- De Groot, B.L.; Grubmüller, H. Water permeation across biological membranes: Mechanism and dynamics of aquaporin-1 and GlpF. Science 2001, 294, 2353–2357. [Google Scholar] [CrossRef] [PubMed]
- Hanson, K.; Nicolau, D.V.; Filipponi, L.; Wang, L.; Lee, A.P. Fungi use efficient algorithms for the exploration of microfluidic networks. Small 2006, 2, 1212–1220. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.G.; Kovács, G.M.; Krizsán, K. Complex multicellularity in fungi: Evolutionary convergence, single origin, or both? Biol. Rev. 2018, 93, 1778–1794. [Google Scholar] [CrossRef] [PubMed]
- László, G.N.; Varga, T.; Csernetics, Á.; Virágh, M. Fungi took a unique evolutionary route to multicellularity: Seven key challenges for fungal multicellular life. Fungal Biol. Rev. 2020, 34, 151–169. [Google Scholar] [CrossRef]
- Silberfeld, T.; Leigh, J.W.; Verbruggen, H.; Cruaud, C.; de Reviers, B.; Rousseau, F. A multi-locus time-calibrated phylogeny of the brown algae (Heterokonta, Ochrophyta, Phaeophyceae): Investigating the evolutionary nature of the “brown algal crown radiation”. Mol. Phylogenetics Evol. 2010, 56, 659–674. [Google Scholar] [CrossRef] [PubMed]
- Lutzoni, F.; Nowak, M.D.; Alfaro, M.E.; Reeb, V.; Miadlikowska, J.; Krug, M.; Arnold, A.E.; Lewis, L.A.; Swofford, D.L.; Hibbett, D.; et al. Contemporaneous radiations of fungi and plants linked to symbiosis. Nat. Commun. 2018, 9, 5451. [Google Scholar] [CrossRef] [PubMed]
- Irisarri, I.; Lorente-Martínez, H.; Strassert, J.F.H.; Zardoya, A.A.R.; San Mauro, D.; de Vries, J. Early Diversification of Membrane Intrinsic Proteins (MIPs) in Eukaryotes. Genome Biol. Evol. 2024, 16, evae164. [Google Scholar] [CrossRef] [PubMed]
- Burki, F.; Roger, A.J.; Brown, M.W.; Simpson, A.G.B. The new tree of eukaryotes. Trends Ecol. Evol. 2020, 35, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Strassert, J.F.H.; Irisarri, I.; Williams, T.A.; Burki, F. A molecular timescale for eukaryote evolution with implications for the origin of red algal- derived plastids. Nat. Commun. 2021, 12, 1879. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Mak, E.; Wu, B.; Beitz, E. Parasite aquaporins: Current developments in drug facilitation and resistance. Biochim. Biophys. Acta 2014, 1840, 1566–1573. [Google Scholar] [CrossRef] [PubMed]
- Hongsanan, S.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Samarakoon, M.C.; Jeewon, R.; Zhao, Q.; Al-Sadi, A.M.; Bahkali, A.H. An updated phylogeny of Sordariomycetes based on phylogenetic and molecular clock evidence. Fungal Divers. 2017, 84, 25–41. [Google Scholar] [CrossRef]
- Altschul, S.; Madden, T.; Schaffer, A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D. Gapped BLASt and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3401. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Sonnhammer, E.L.L.; von Heijne, G.; Krogh, A. A hidden Markov model for predicting transmembrane helices in protein sequences. In Proceedings of the Sixth International Conference on Intelligent Systems for Molecular Biology, Montreal, QC, Canada, 28 June–1 July 1998; AAAI Press: Menlo Park, CA, USA, 1998; pp. 175–182. [Google Scholar]
- Hirokawa, T.; Boon Chieng, S.; Mitaku, S. SOSUI: Classification and secondary structure prediction system for membrane proteins. Bioinformatics 1998, 14, 378–379. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Berka, K.; Hanák, O.; Sehnal, D.; Banáš, P.; Navratilova, V.; Jaiswal, D.; Ionescu, C.-M.; Vařeková, R.S.; Koča, J.; Otyepka, M. MOLE online 2.0: Interactive web-based analysis of biomacromolecular channels. Nucleic Acids Res. 2012, 40, W222–W227. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.; Wong, T.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) XIP Physicochemical Properties *** | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Code * | Phyla | Cluster ** | Size (AA) | MW (kDa) | pI Moyen | Instability Index | Aliphatic Index | GRAVI | ||
| I | Pucciniomycotina | A | 275 ± 7 | 29.174 ± 1.292 | 7.36 ± 0.71 | 30.09 ± 2.24 | 99.81 ± 6.62 | 0.585 ± 0.071 | ||
| III | Agaricomycetes | B-C1-C2 | 334 ± 21 | 35.411 ± 1.962 | 7.54 ± 1.18 | 33.87 ± 6.83 | 95.46 ± 5.34 | 0.386 ± 0.093 | ||
| IV | Dacrymycetes | C2 | 322 ± 1 | 35.793 ± 0.334 | 8.22 ± 0.64 | 35.49 ± 1.85 | 95.78 ± 6.41 | 0.376 ± 0.057 | ||
| V | Tremellomycetes | C1-C2 | 326 ± 8 | 38.824 ± 1.018 | 8.22 ± 1.13 | 37.79 ± 6.68 | 95.12 ± 6.59 | 0.336 ± 0.105 | ||
| VII | Pezizomycetes | C2 | 316 ± 9 | 33.123 ± 1.256 | 7.71 ± 1.47 | 34.98 ± 4.79 | 98.73 ± 8.07 | 0.399 ± 0.111 | ||
| VIII | Orbiliomycetes | C2 | 351 ± 9 | 37.624 ± 0.964 | 8.18 ± 0.88 | 35.99 ± 2.96 | 89.01 ± 2.31 | 0.282 ± 0.061 | ||
| IX | Eurotiomycetes | C1-C2 | 326 ± 13 | 35.050 ± 1.344 | 7.53 ± 1.01 | 36.49 ± 4.59 | 96.09 ± 3.21 | 0.429 ± 0.087 | ||
| X | Dothideomycetes | C1-C2 | 323 ± 7 | 34.369 ± 0.914 | 7.29 ± 0.93 | 36.21 ± 4.94 | 99.11 ± 4.67 | 0.391 ± 0.086 | ||
| XI | Arthoniomycetes | C2 | 323 | 34.2391 | 6.41 | 40.04 | 93.62 | 0.355 | ||
| XII | Lecanoromycetes | C1 | 313 ± 14 | 33.297 ± 1.747 | 7.04 ± 1.15 | 33.42 ± 4.59 | 91.07 ± 3.82 | 0.361 ± 0.097 | ||
| XIII | Leotiomycetes | C2 | 319 ± 13 | 34.358 ± 1.569 | 7.92 ± 0.71 | 36.63 ± 5.13 | 93.28 ± 4.51 | 0.354 ± 0.075 | ||
| XIV | Sordariomycetes | C1-C2 | 332 ± 12 | 35.012 ± 1.141 | 7.58 ± 1.03 | 36.46 ± 4.41 | 89.55 ± 4.47 | 0.388 ± 0.097 | ||
| XV | Xylonomycetes | C1-C2 | 324 ± 2 | 34.687 ± 0.311 | 7.062 ± 1.01 | 34.46 ± 3.33 | 96.03 ± 2.57 | 0.374 ± 0.033 | ||
| XVI | Saccharomycotina | C1-C2 | 316 ± 5 | 33.998 ± 0.762 | 7.78 ± 0.28 | 34.12 ± 3.12 | 94.66 ± 4.99 | 0.373 ± 0.067 | ||
| XVIII | Glomeromycotina | C2 | 346 ± 22 | 38.362 ± 2.395 | 6.55 ± 0.73 | 39.80 ± 2.98 | 102.52 ± 8.08 | 0.297 ± 0.157 | ||
| XXI | Zoopagomycotina | C2 | 317 | 34.521 | 6.69 | 25.85 | 103.47 | 0.29 | ||
| XXII | Entomophthoromycotina | C2 | 334 ± 10 | 36.010 ± 0.981 | 6.36 ± 0.51 | 34.91 ± 4.66 | 93.83 ± 4.21 | 0.337 ± 0.072 | ||
| Mean | 326 | 34.973 | 7.24 | 35.34 | 95.46 | 0.356 | ||||
| SD | 12 | 1.537 | 0.60 | 3.46 | 4.53 | 0.044 | ||||
| (B) XIP Structure Singularities **** | ||||||||||
| Code * | Phyla | TMH | SubCL | NPA-LB | NPA-LE | Ar/r | ||||
| I | Pucciniomycotina | 6 | PM | (NS)P(IFLV) | (FY)P(NAT) | (VY)(VNCA)(VI)R | ||||
| III | Agaricomycetes | 6 | PM | N(PS)(LM) | (FN)P(NAGS) | (NTS)(VGTN)(SLGC)R | ||||
| IV | Dacrymycetes | 6 | PM | NP(MT) | NPA | N(ST)SR | ||||
| V | Tremellomycetes | 6 | PM(Mito) | NP(LIMF) | NP(ATGS) | (NT)(TVFM)(QS)R | ||||
| VII | Pezizomycetes | 6 | PM(Pero) | N(PS)(LM) | NP(ASTCG) | (IAV)(STE)SR | ||||
| VIII | Orbiliomycetes | 6 | PM | NPL | NP(AGT) | (SA)(AI)(AS)R | ||||
| IX | Eurotiomycetes | 6 | PM | (NPS)P(TLM) | NPA | (NAQ)(SAT)(SCGA)R | ||||
| X | Dothideomycetes | 6 | PM | NP(LFTSA) | N(PS)A | (NTQ)(STL)(SGA)R | ||||
| XI | Arthoniomycetes | 6 | PM | NPM | NPA | QICR | ||||
| XII | Lecanoromycetes | 6 | PM | NP(MTL) | (NH)P(AT) | (NS)(STG)(SGA)R | ||||
| XIII | Leotiomycetes | 6 | PM | NP(VTILM) | NP(ASG) | (NS)(ASTL)(GNS)R | ||||
| XIV | Sordariomycetes | 6 | PM | NP(LVMTSA) | (NMH)P(ASTG) | (NCST)(VSTAV)(QSGN)R | ||||
| XV | Xylonomycetes | 6 | PM | N(PSQ)(TFL) | (NF)P(ASG) | (LTN)(AGT)SR | ||||
| XVI | Saccharomycotina | 6 | PM | NP(LTI) | NP(GA) | (NGI)(IS)SR | ||||
| XVIII | Glomeromycotina | 6 | PM(Mito) | N(PS)(FL) | NP(AV) | H(VA)SK | ||||
| XXI | Zoopagomycotina | 6 | PM | NPS | NPA | HISR | ||||
| XXII | Entomophthoromycotina | 6 | PM | N(PT)(MLCI) | NP(AG) | (VT)IGK | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venisse, J.-S.; Bronner, G.; Saadaoui, M.; Roeckel-Drevet, P.; Faize, M.; Fumanal, B. Major Intrinsic Proteins in Fungi: A Special Emphasis on the XIP Subfamily. J. Fungi 2025, 11, 543. https://doi.org/10.3390/jof11070543
Venisse J-S, Bronner G, Saadaoui M, Roeckel-Drevet P, Faize M, Fumanal B. Major Intrinsic Proteins in Fungi: A Special Emphasis on the XIP Subfamily. Journal of Fungi. 2025; 11(7):543. https://doi.org/10.3390/jof11070543
Chicago/Turabian StyleVenisse, Jean-Stéphane, Gisèle Bronner, Mouadh Saadaoui, Patricia Roeckel-Drevet, Mohamed Faize, and Boris Fumanal. 2025. "Major Intrinsic Proteins in Fungi: A Special Emphasis on the XIP Subfamily" Journal of Fungi 11, no. 7: 543. https://doi.org/10.3390/jof11070543
APA StyleVenisse, J.-S., Bronner, G., Saadaoui, M., Roeckel-Drevet, P., Faize, M., & Fumanal, B. (2025). Major Intrinsic Proteins in Fungi: A Special Emphasis on the XIP Subfamily. Journal of Fungi, 11(7), 543. https://doi.org/10.3390/jof11070543