


Identifying Key Pathogens and Effective Control Agents for Astragalus membranaceus var. mongholicus Root Rot
 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Occurrence of Root Rot in Astragalus membranaceus var. mongholicus
2.2. Sample Collection and Screening of Test Agents and Biocontrol Bacteria
2.3. Isolation and Identification of Pathogens
2.3.1. Isolation of Pathogens
2.3.2. Determination of the Pathogenicity of Pathogenic Fungi
2.3.3. Morphological Observation of Pathogenic Fungi
2.3.4. Molecular Identification of Pathogenic Fungi
2.4. Determination of the Sensitivity of Pathogenic Fungi to Fungicides
2.5. Determination of the Sensitivity of Pathogenic Fungi to Biocontrol Bacteria
2.5.1. Activation of Pathogenic Fungi and Probiotic Bacteria
2.5.2. Evaluation of the Efficacy of Biocontrol Bacteria Against Pathogenic Fungi
2.6. Data Analysis
3. Results
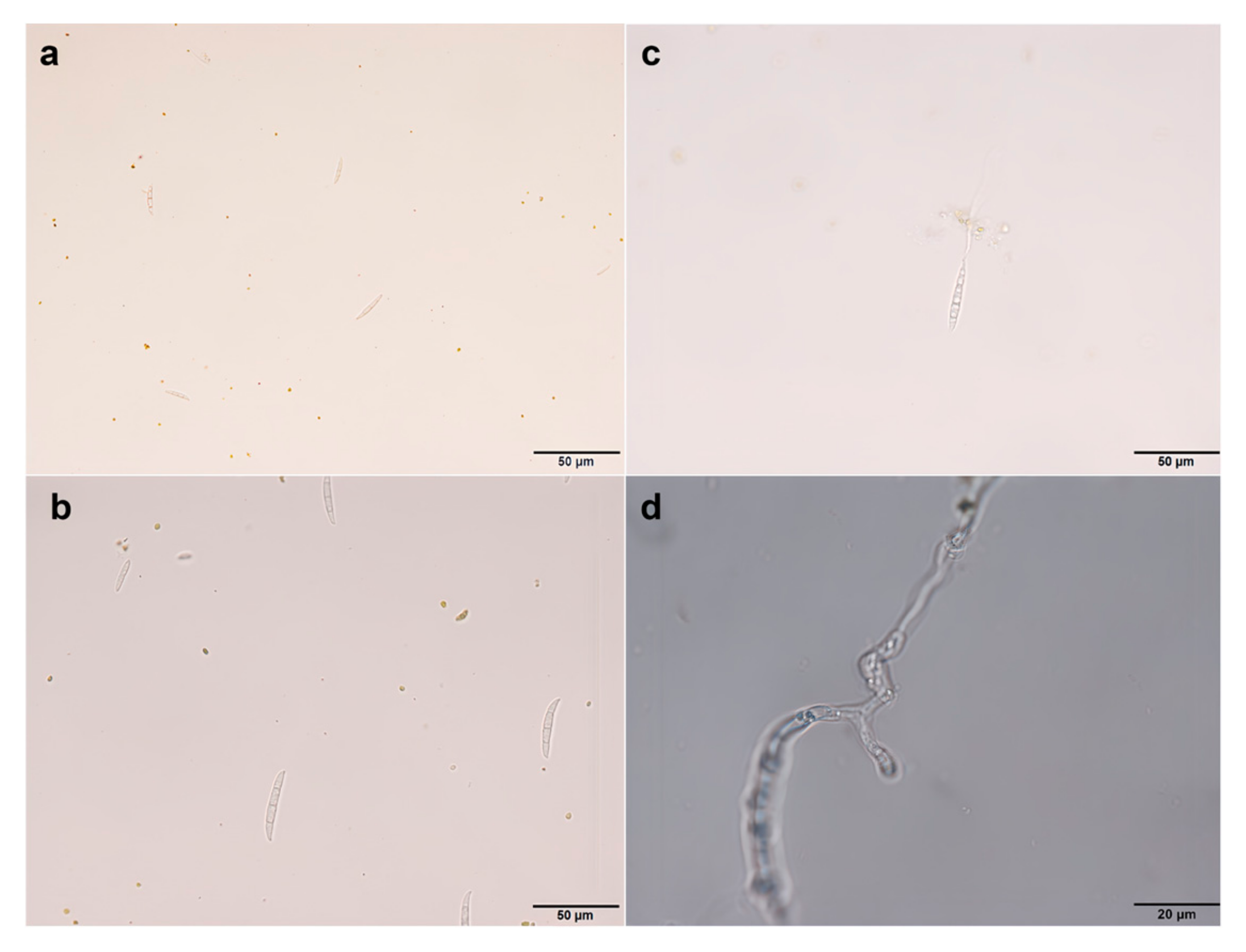
3.1. Morphological Observation and Pathogenicity Determination of Pathogenic Fungi
3.2. Phylogenetic Analysis
3.3. Toxicity of Fungicides to F. solani and F. oxysporum
3.4. Control Effect of Biocontrol Bacteria on F. solani and F. oxysporum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- National Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2020; pp. 283–284. [Google Scholar]
- Sumner, D. Etiology and control of root rot of summer squash in Georgia. Plant Dis. Rep. 1976, 60, 923–927. [Google Scholar]
- Gibert, S.; Edel-Hermann, V.; Gautheron, E.; Gautheron, N.; Sol, J.M.; Capelle, G.; Galland, R.; Bardon-Debats, A.; Lambert, C.; Steinberg, C. First Report of Fusarium avenaceum, Fusarium oxysporum, Fusarium redolens, and Fusarium solani Causing Root Rot in Pea in France. Plant Dis. 2022, 106, 1297. [Google Scholar] [CrossRef]
- Li, J.; Cheng, L. Fusarium solani Causing Root Rot Disease on Gastrodia elata in Shaxi, China. Plant Dis. 2021, 106, 320. [Google Scholar] [CrossRef]
- Zhao, X.; Yue, L.; Uwaremwe, C.; Liu, Y.; Tian, Y.; Zhao, H.; Zhou, Q.; Zhang, Y.; Wang, R. First Report of Root Rot Caused by the Fusarium oxysporum Species Complex on Codonopsis pilosula in China. Plant Dis. 2021, 105, 3742. [Google Scholar] [CrossRef]
- Ma, L.J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Zhao, Z.X.; Qin, X.; Lei, Z. Analysis of dominant pathogen community causing Astragalus membranaceus var. mongholicus root rot in Shanxi Province. J. Plant Prot. 2018, 45, 878–885. [Google Scholar]
- Ma, W.; Wang, B.; Ma, L.; Xie, J.; Kong, X. Inhibitory Effect of Chinese Medicine Extracts on Pathogen of Astragalus Root Rot. Bull. Bot. Res. 2010, 30, 634–636. [Google Scholar]
- Niu, S.Q.; Geng, H.; Han, C.H.; Yan, W.R.; Da, W.Y. Isolation and identification of pathogens causing Astragalus membranaceus root rot in Longxi of Gansu Province. J. Northwest Norm. Univ. 2016, 52, 75–78. [Google Scholar]
- Bai, L.; Wang, Y.; Li, Y.; Zhang, X.; Lu, Z.; Zhang, D.; Sun, F.; Zhao, X. Effects of Root Exudates from Different Croprotation on Root Rot Pathogen of the Astragalus Membranaceus (Fisch.) Bge. var. Mongholicus (Bge.) Hsiao; Inner Mongolia University: Hohhot, China, 2018. [Google Scholar]
- Luo, D.G.; Han, X.P.; Deng, C.G.; Zhang, G.Z.; Liu, X.Z.; Li, Z.M. Investigation and Pathogen Identification of Medicinal Astragalus Disease in Dingxi City. Gansu Agric. Sci. Technol. 2004, 1, 38–40. [Google Scholar]
- Chen, Y.; Zhu, L.; Guo, F.X.; Wu, Z.J. Isolation and identification of the pathogens causing Astragalus membranaceus var. mongholicus root rot in Weiyuan of Gansu Province. Acta Phytopathol. Sin. 2011, 41, 428–431. [Google Scholar]
- Ma, G.H.; Duan, X.M.; Xu, W.H.; Zhou, Y.D.; Ma, H.X.; Ma, W.L.; Qi, H.X. Identification and Laboratory Screening of Chemical Agents of Root Rot Pathogens of Astragalus membranaceus var. mongholicus. Acta Agrestia Sin. 2022, 30, 1122–1130. [Google Scholar]
- Zhou, M.; Bai, Q.R. Isolation and Identification of Root Rot Pathogen of Astragalus Membranaceus in Moqi, Inner Mongolia. J. Northeast Agric. Sci. 2021, 46, 52–55. [Google Scholar]
- Guan, Y.M.; Zhang, S.N.; Ma, Y.Y.; Zhang, Y.Y. Occurrence of Dactylonectria torresensis causing root rot on Astragalus membranaceus in northeastern China. Plant Dis. 2021, 105, 1219. [Google Scholar] [CrossRef]
- Qi, H.; Duan, X.; Wenhua, X.; Zhou, Y.; Ma, H.; Ma, W.; Ma, G. First report disease of Clonostachys rosea causing root rot on Astragalus membranaceus in China. Plant Dis. 2022, 106, 1752. [Google Scholar] [CrossRef]
- Mu, X.R.; Ma, Y.Y.; Yang, Z.Z.; Ma, L.; Jiang, Y.B. Research advance on the control of root rot disease of medical plants. Pharm. Clin. Chin. Mater. Medica 2014, 5, 5–8+52. [Google Scholar]
- Fan, Q.; Jian, H. Toxicity Determination of Several Fungicides to Pathogens of Astragalus Root Rot. China Plant Prot. 2009, 29, 41–43. [Google Scholar]
- Zhu, L. Isolation and Identification the Pathogens Causing Astragalas mongholicus Root Rot and the Fungicides Screening. Doctor’s Thesis, Gansu Agricultural University, Lanzhou, China, 2010. [Google Scholar]
- Gao, F.; Zhao, X.-X.; Yan, H.; Lei, Z.-H.; Wang, M.-L.; Qin, X.-M. Screening and identification of antagonistic Bacillus against Astragalus membranaceus root rot and its effect on microorganism community in root zone soil. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Medica 2019, 44, 3942–3947. [Google Scholar]
- Xu, L.L.; Bai, X.L.; Li, Z.F. Screening of antagonistic bacteria and biocontrol mechanism of astragalus root rot. J. Northwest Norm. Univ. 2024, 60, 77–85. [Google Scholar] [CrossRef]
- Lu, Z.X.; Guo, W.N.; Liu, C. Isolation, identification and characterization of novel Bacillus subtilis. J. Vet. Med. Sci. 2018, 80, 427–433. [Google Scholar] [CrossRef]
- Fang, Z.D. Research Methods of Plant Disease, 3rd ed.; China Agricultural Publishing House: Beijing, China, 1998; pp. 6–13. [Google Scholar]
- Gams, W. Laboratory guide to the identification of the major species. Neth. J. Plant Pathol. 1978, 84, 84. [Google Scholar] [CrossRef]
- He, C.; Shi, J.; Shen, H.; Zhang, J.; Wang, D.; Zhang, J.; Wang, R.; Zhang, S.; Li, J. Research of different pesticides treatment on controlling the root rot disease of Astragalas mongholicus. Pratacultural Sci. 2013, 30, 1948–1952. [Google Scholar]
- Fan, L.; Huang, Z.; Jin, X.; Lin, H.; Liu, X.; Meng, J.; Zhao, S.; Zhou, J. Investigation of the impact of microbial agent treatment on controlling Astragalus membranaceus root rot disease. Chin. J. Pestic. Sci. 2025, 27, 160–170. [Google Scholar] [CrossRef]
- Yang, B.; Li, Y.; Xu, J.; Sha, Y. Screening, Identification, and Application Effects of Actinomycetes on Astragalus membranaceus Root Rot. Chin. J. Biol. Control. 2025, 41, 554–560. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, H.; Li, S.; Dong, Z.; Long, S.; Huang, X.; Guo, L.; Kang, C.; Liang, J.; Ma, X. The control effect of Trichoderma harzianum EMF910 on theroot rot pathogens of Astragalus membranaceus in Ningxia saline-alkali regions. Microbiology 2024, 51, 4162–4180. [Google Scholar] [CrossRef]
- Islam, T.; Danishuddin; Tamanna, N.T.; Matin, M.N.; Barai, H.R.; Haque, A. Resistance Mechanisms of Plant Pathogenic Fungi to Fungicide, Environmental Impacts of Fungicides, and Sustainable Solutions. Plants 2024, 13, 2737. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.L.; Liu, H.; Li, Z.J.; Wang, J.; Zhang, J.; Chen, H.B.; Yang, C.P.; Zhang, M.; Yang, L. Screening and Compounding of Biocontrol Bacillus and Fungicide and Their Control Effect on Branch Withered Disease on Koelreuteria paniculata. J. Sichuan Agric. Univ. 2021, 39, 55–62. [Google Scholar]
- Yang, S.Y.; Yang, F.; Duan, H.M.; Chen, S.; Zhou, H.; Zhang, J. Inhibitory Activity of the Combination of Biocontrol Metabolites and Fungicides Against Peanut Brown Spot Pathogen. J. Jianghan Univ. 2023, 51, 61–67. [Google Scholar]
- Mao, X.Q.; Peng, Z.R.; Qiu, H.P.; Zhang, Z.; Chai, R.Y.; Wang, J.Y.; Wang, Y.L.; Sun, G.C. Fermentation Conditions of Biocontrol Strain MT-06 and Efficacy of Mixed Fungicide against Wheat Fusarium Head Blight. J. Zhejiang Agric. Sci. 2013, 7, 821–824. [Google Scholar]
- Gao, Y.; Zhang, Y.; Cheng, X.; Zheng, Z.; Wu, X.; Dong, X.; Hu, Y.; Wang, X. Agricultural Jiaosu an eco-friendly and cost-effective control strategy for suppressing Fusarium root rot disease in Astragalus membranaceus. Front. Microbiol. 2022, 13, 823704. [Google Scholar] [CrossRef]
- Matthews, G. Chemical Pesticide Markets, Health Risks and Residues. Crop Prot. 2001, 20, 452. [Google Scholar] [CrossRef]
- Al-Hatmi, S.M.A.; Meis, F.J.; Hoog, D.S.G. Fusarium: Molecular Diversity and Intrinsic Drug Resistance. PLoS Pathog. 2016, 12, e1005464. [Google Scholar] [CrossRef]
- Busi, Z.; Dan, H.; Li, W. Advances in Fusarium Drug Resistance Research. J. Glob. Antimicrob. Resist. 2021, 24, 215–219. [Google Scholar] [CrossRef]
- Khan, N.; Maymon, M.; Hirsch, M.A. Combating Fusarium Infection Using Bacillus-Based Antimicrobials. Microorganisms 2017, 5, 75. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungicide (Common Name) | Chemical Formula | Fungicide Class | Initial Concentration (mg/L) | Series Dilution Concentration (mg/L) |
|---|---|---|---|---|
| Difenoconazole | C19H17Cl2N3O3 | Triazol | 1000 | 0.001, 0.01, 0.1, 1, 5, 10 |
| Chlorothalonil | C8Cl4N2 | Substitutive benzene | 1000 | 0.001, 0.01, 0.1, 1, 5, 10 |
| Bromothalonil | C6H6Br2N2 | Bromocyanoalkanes | 1000 | 0.001, 0.01, 0.1, 1, 5, 10 |
| Thiophanate-methyl | C12H14N4O4S2 | Substitutive benzene | 1000 | 0.001, 0.01, 0.1, 1, 5, 10 |
| Hymexazol | C4H5NO2 | Triadimefon | 1000 | 0.001, 0.01, 0.1, 1, 5, 10 |
| Tebuconazole | C16H22ClN3O | Triazol | 1000 | 0.001, 0.01, 0.1, 1, 5, 10 |
| Fludioxonil | C12H6F2N2O2 | Phenylpyrrole | 1000 | 0.001, 0.01, 0.1, 0.5, 1, 2 |
| Carbendazim | C9H9N3O2 | Benzimidazole | 1000 | 0.001, 0.01, 0.1, 1, 5, 10 |
| Fungicide | Isolate | Virulent Regression Equation | EC50/ (μg/mL) | Correlation Coefficient, r | Average EC50 Value/(μg/mL) |
|---|---|---|---|---|---|
| tebuconazole | CDF1 | Y = 0.5368 ∗ X + 4.505 | 8.36 | 0.98 | 1.74 |
| CDF2 | Y = 0.6164 ∗ X + 5.336 | 0.29 | 0.96 | ||
| CDF3 | Y = 0.6492 ∗ X + 4.855 | 1.67 | 0.99 | ||
| CDF4 | Y = 0.6911 ∗ X + 5.176 | 0.56 | 0.98 | ||
| CDF5 | Y = 0.7754 ∗ X + 5.486 | 0.24 | 0.98 | ||
| CDF6 | Y = 0.8742 ∗ X + 5.427 | 0.32 | 0.99 | ||
| CDF7 | Y = 0.7390 ∗ X + 5.099 | 0.73 | 0.89 | ||
| difenoconazole | CDF1 | Y = 0.5459 ∗ X + 4.343 | 15.98 | 0.99 | 11.55 |
| CDF2 | Y = 0.4562 ∗ X + 4.481 | 13.73 | 0.99 | ||
| CDF3 | Y = 0.3752 ∗ X + 4.761 | 4.34 | 0.96 | ||
| CDF4 | Y = 0.2438 ∗ X + 4.595 | 45.84 | 0.99 | ||
| CDF5 | Y = 0.5516 ∗ X + 5.362 | 0.22 | 0.99 | ||
| CDF6 | Y = 0.5149 ∗ X + 5.228 | 0.38 | 0.97 | ||
| CDF7 | Y = 0.6981 ∗ X + 5.288 | 0.39 | 0.99 | ||
| chlorothalonil | CDF1 | Y = 0.5787 ∗ X + 4.569 | 5.56 | 0.98 | 8.58 |
| CDF2 | Y = 0.4170 ∗ X + 4.414 | 25.43 | 0.97 | ||
| CDF3 | Y = 0.4801 ∗ X + 4.522 | 9.90 | 0.96 | ||
| CDF4 | Y = 0.4869 ∗ X + 4.515 | 9.91 | 0.98 | ||
| CDF5 | Y = 0.5135 ∗ X + 4.834 | 2.11 | 0.98 | ||
| CDF6 | Y = 0.4439 ∗ X + 4.844 | 2.25 | 0.95 | ||
| CDF7 | Y = 0.3768 ∗ X + 4.739 | 4.93 | 0.96 | ||
| bromothalonil | CDF1 | Y = 0.3942 ∗ X + 3.960 | 434.77 | 0.97 | 368.73 |
| CDF2 | Y = 0.4215 ∗ X + 3.696 | 1240.83 | 0.98 | ||
| CDF3 | Y = 0.4000 ∗ X + 3.835 | 817.52 | 0.96 | ||
| CDF4 | Y = 0.5585 ∗ X + 4.067 | 46.83 | 0.93 | ||
| CDF5 | Y = 0.4883 ∗ X + 4.478 | 12.31 | 0.98 | ||
| CDF6 | Y = 0.4701 ∗ X + 4.381 | 20.74 | 0.96 | ||
| CDF7 | Y = 0.3285 ∗ X + 4.701 | 8.13 | 0.96 | ||
| hymexazol | CDF1 | Y = 0.2996 ∗ X + 4.116 | 892.48 | 0.99 | 959.33 |
| CDF2 | Y = 0.3008 ∗ X + 4.187 | 504.42 | 0.98 | ||
| CDF3 | Y = 0.3277 ∗ X + 4.076 | 660.16 | 0.98 | ||
| CDF4 | Y = 0.3298 ∗ X + 3.925 | 1817.82 | 0.97 | ||
| CDF5 | Y = 0.3508 ∗ X + 3.917 | 1222.44 | 0.98 | ||
| CDF6 | Y = 0.3316 ∗ X + 3.972 | 1259.28 | 0.97 | ||
| CDF7 | Y = 0.4431 ∗ X + 3.868 | 358.70 | 0.99 | ||
| thiophanate- methyl | CDF1 | Y = 0.3701 ∗ X + 4.473 | 26.54 | 0.94 | 36.82 |
| CDF2 | Y = 0.4092 ∗ X + 4.504 | 16.30 | 0.98 | ||
| CDF3 | Y = 0.4116 ∗ X + 4.587 | 10.08 | 0.96 | ||
| CDF4 | Y = 0.3127 ∗ X + 4.539 | 29.80 | 0.99 | ||
| CDF5 | Y = 0.5373 ∗ X + 4.457 | 10.25 | 0.99 | ||
| CDF6 | Y = 0.3421 ∗ X + 4.521 | 25.13 | 0.94 | ||
| CDF7 | Y = 0.2606 ∗ X + 4.441 | 139.65 | 0.97 | ||
| fludioxonil | CDF1 | Y = 0.3546 ∗ X + 3.716 | 4178.12 | 0.90 | 2872.67 |
| CDF2 | Y = 0.2119 ∗ X + 4.458 | 361.25 | 0.88 | ||
| CDF3 | Y = 0.3290 ∗ X + 3.847 | 3195.65 | 0.91 | ||
| CDF4 | Y = 0.2238 ∗ X + 4.210 | 3387.95 | 0.85 | ||
| CDF5 | Y = 0.3708 ∗ X + 3.646 | 4482.95 | 0.91 | ||
| CDF6 | Y = 0.3646 ∗ X + 3.702 | 3631.33 | 0.91 | ||
| CDF7 | Y = 0.3765 ∗ X + 3.893 | 871.44 | 0.91 | ||
| carbendazim | CDF1 | Y = 0.9079 ∗ X + 5.303 | 0.46 | 0.96 | 0.44 |
| CDF2 | Y = 0.7022 ∗ X + 5.254 | 0.43 | 0.98 | ||
| CDF3 | Y = 0.8832 ∗ X + 5.053 | 0.87 | 0.96 | ||
| CDF4 | Y = 0.6784 ∗ X + 5.196 | 0.51 | 0.97 | ||
| CDF5 | Y = 1.336 ∗ X + 5.778 | 0.26 | 0.93 | ||
| CDF6 | Y = 1.356 ∗ X + 5.775 | 0.27 | 0.92 | ||
| CDF7 | Y = 1.177 ∗ X + 5.658 | 0.28 | 0.85 |
| Treatment | Inhibition Rate of Fungal Colony Growth (%) | ||||||
|---|---|---|---|---|---|---|---|
| CDF1 | CDF2 | CDF3 | CDF4 | CDF5 | CDF6 | CDF7 | |
| KRS002 | 37.70 ± 2.18 | 33.06 ± 1.31 | 40.49 ± 2.22 | 21.17 ± 3.23 | 38.59 ± 1.24 | 29.01 ± 3.03 | 41.91 ± 1.43 |
| KRS003 | 12.12 ± 1.82 | 1.74 ± 0.99 | 10.67 ± 0.80 | 6.78 ± 0.93 | 4.55 ± 0.75 | 4.03 ± 0.40 | 2.63 ± 0.34 |
| KRS004 | 17.59 ± 0.47 | 21.78 ± 0.99 | 8.01 ± 0.65 | 10.52 ± 0.94 | 9.14 ± 1.13 | 4.48 ± 0.69 | 10.91 ± 1.21 |
| KRS006 | 55.51 ± 3.34 | 44.83 ± 0.27 | 49.42 ± 0.25 | 47.60 ± 0.44 | 42.57 ± 0.49 | 45.70 ± 0.74 | 46.39 ± 1.30 |
| KRS008 | 23.79 ± 2.62 | 16.96 ± 2.95 | 10.29 ± 3.33 | 13.58 ± 4.78 | 6.79 ± 2.24 | 7.25 ± 0.552 | 11.18 ± 0.59 |
| KRS009 | 20.46 ± 0.50 | 21.47 ± 0.68 | 23.92 ± 0.30 | 7.28 ± 0.47 | 5.17 ± 0.19 | 2.83 ± 0.18 | 4.76 ± 0.20 |
| KRS013 | 24.49 ± 3.45 | 10.43 ± 0.51 | 15.21 ± 1.29 | 9.68 ± 0.40 | 5.89 ± 1.43 | 5.84 ± 0.28 | 10.20 ± 1.05 |
| KRS014 | 26.58 ± 3.06 | 17.75 ± 2.56 | 17.11 ± 2.57 | 8.50 ± 2.66 | 14.09 ± 4.23 | 19.03 ± 5.16 | 5.80 ± 2.11 |
| KRS034 | 24.80 ± 1.67 | 21.88 ± 3.79 | 19.33 ± 1.09 | 19.02 ± 4.53 | 14.77 ± 3.28 | 15.44 ± 3.25 | 31.73 ± 5.31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Xia, B.; Wang, C.; Xiao, O.; Wang, T.; Zhao, H.; Dai, X.; Chen, J.; Wang, Y.; Kong, Z. Identifying Key Pathogens and Effective Control Agents for Astragalus membranaceus var. mongholicus Root Rot. J. Fungi 2025, 11, 544. https://doi.org/10.3390/jof11070544
Zhang B, Xia B, Wang C, Xiao O, Wang T, Zhao H, Dai X, Chen J, Wang Y, Kong Z. Identifying Key Pathogens and Effective Control Agents for Astragalus membranaceus var. mongholicus Root Rot. Journal of Fungi. 2025; 11(7):544. https://doi.org/10.3390/jof11070544
Chicago/Turabian StyleZhang, Bo, Bingyan Xia, Chunyan Wang, Ouli Xiao, Tielin Wang, Haoran Zhao, Xiaofeng Dai, Jieyin Chen, Yonggang Wang, and Zhiqiang Kong. 2025. "Identifying Key Pathogens and Effective Control Agents for Astragalus membranaceus var. mongholicus Root Rot" Journal of Fungi 11, no. 7: 544. https://doi.org/10.3390/jof11070544
APA StyleZhang, B., Xia, B., Wang, C., Xiao, O., Wang, T., Zhao, H., Dai, X., Chen, J., Wang, Y., & Kong, Z. (2025). Identifying Key Pathogens and Effective Control Agents for Astragalus membranaceus var. mongholicus Root Rot. Journal of Fungi, 11(7), 544. https://doi.org/10.3390/jof11070544