
Thaldh3-Dependent GABA Metabolism Modulates Response of Trichoderma to Fusaric Acid-Induced Oxidative Stress

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
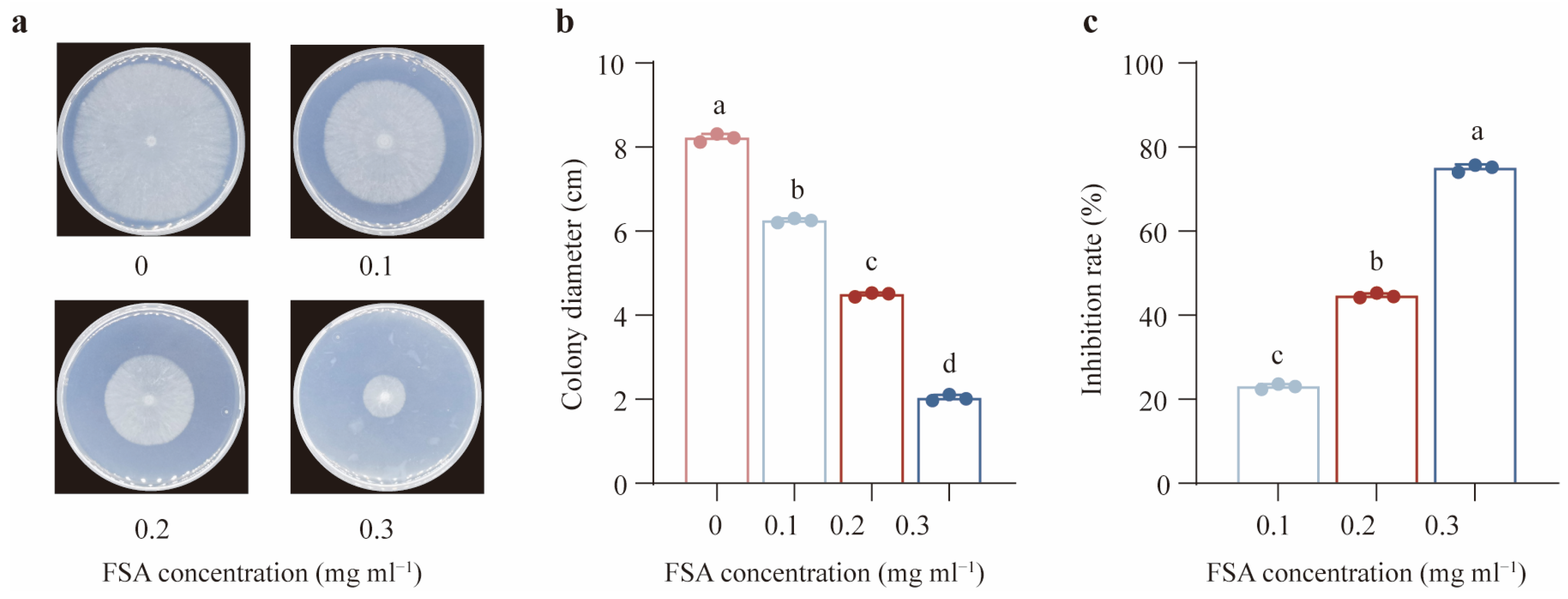
2.2. Inhibitory Effects of FSA on Th
2.3. RNA Sequencing and Transcriptome Analysis
2.4. Extraction of T. harzianum NJAU4742 Genomic DNA and Construction of Homologous Fragments
2.5. Generation of Targeted Gene Mutants in T. harzianum NJAU4742
2.6. Assay of Mycelium Inhibition Growth Rate and Intracellular Metabolites
2.7. Pot Experiments
- T1 (CK): Soil inoculated with Fol only, serving as the control.
- T2: Soil co-inoculated with Fol and Th-WT strains.
- T3: Soil co-inoculated with Fol and the ΔThaldh3 strains.
- T4: Soil co-inoculated with Fol and ΔThaldh3, with exogenous GABA added at 100 μg g−1 soil.
2.8. Soil Sampling, DNA Extraction, and Real-Time Quantitative PCR Analysis
2.9. Statistical Analyses
3. Results
3.1. The Transcriptome Profiling of Th Exposed to FSA
3.2. FSA Induced Severe Intracellular Oxidative Stress in Th
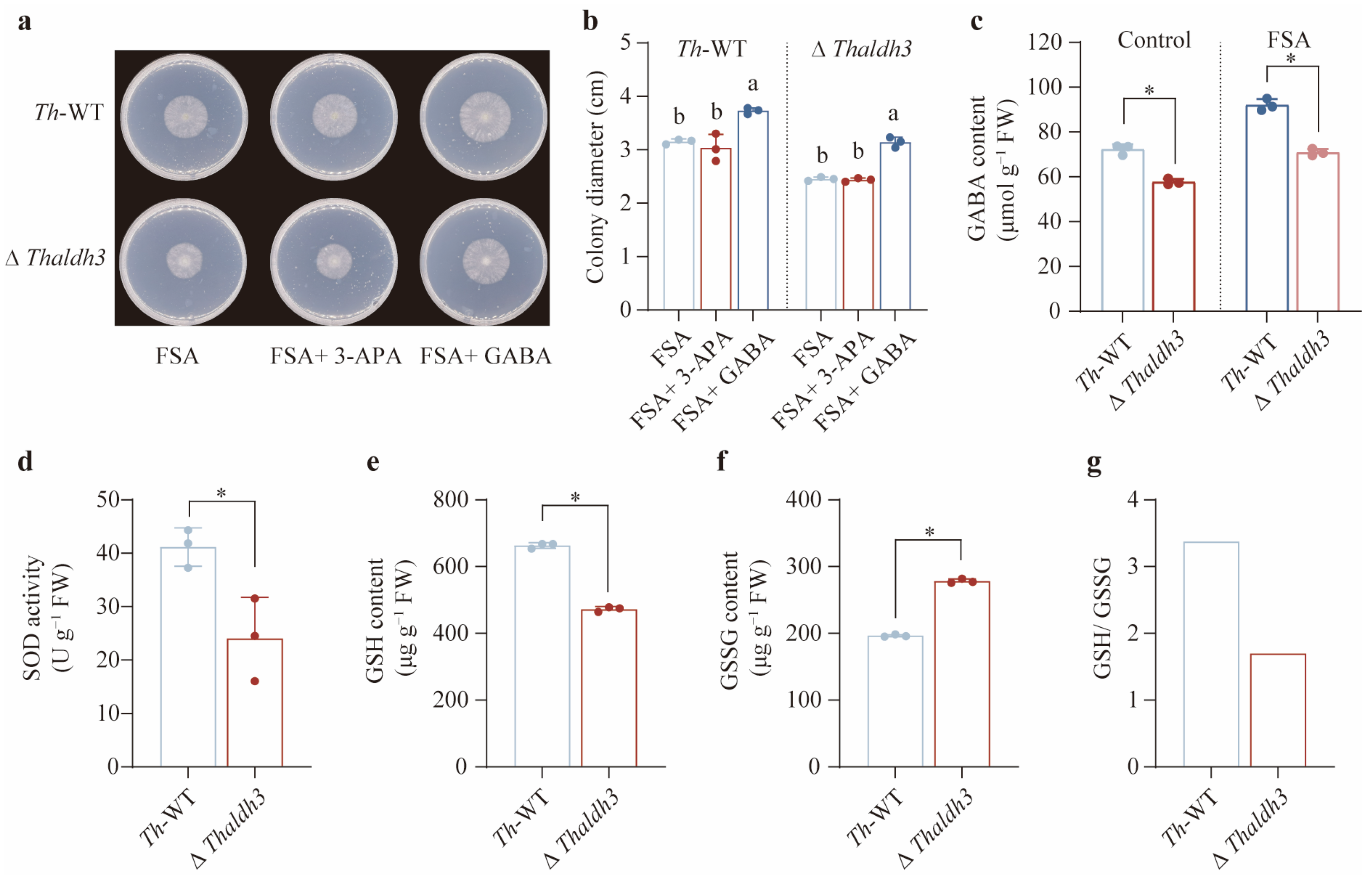
3.3. Thaldh3 Played a Critical Role in the Response to FSA Stress
3.4. GABA Was the Key Substance to Mitigate FSA-Triggered Oxidative Stress
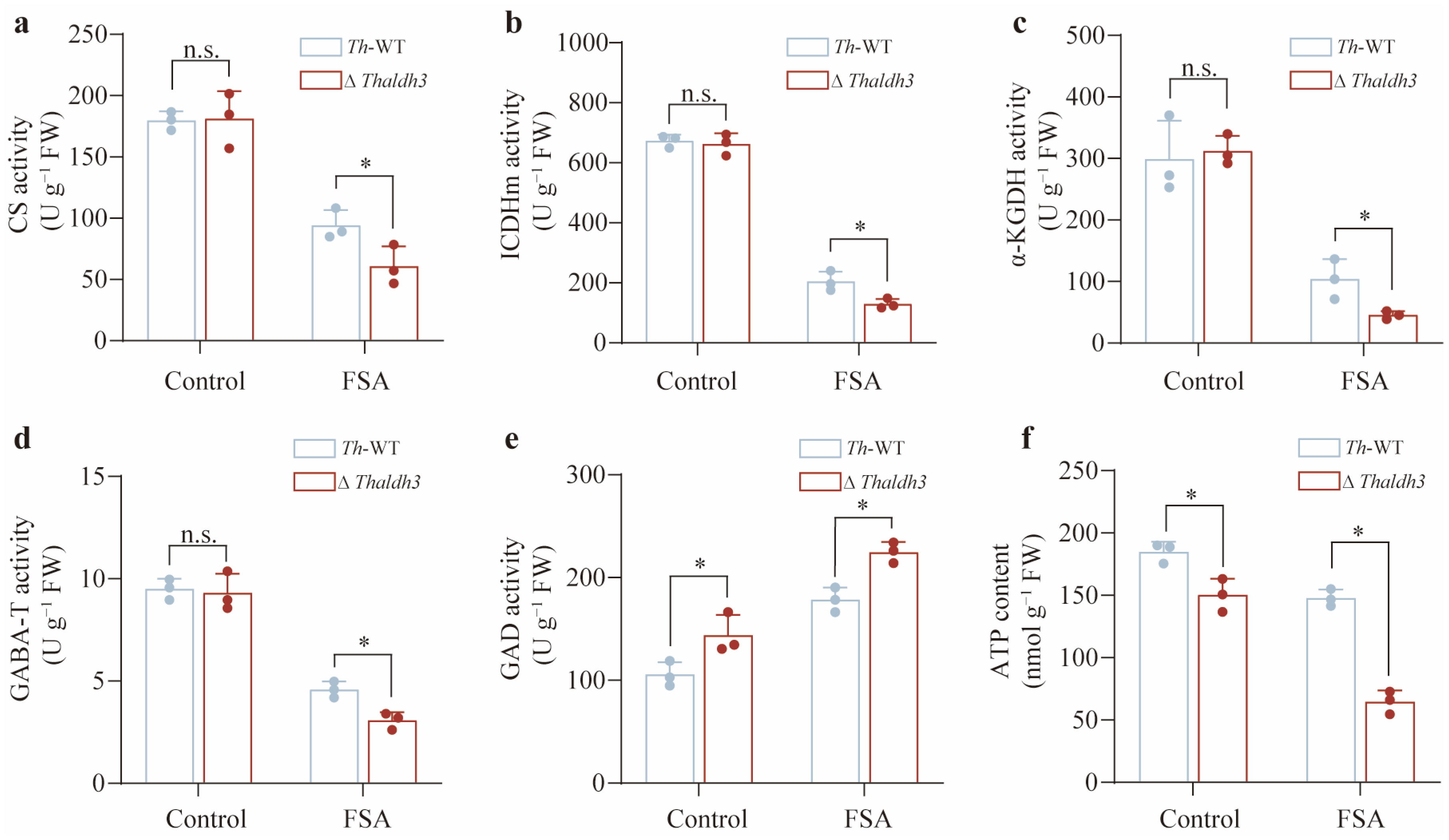
3.5. Thaldh3 Alleviated FSA Stress by Upregulating the Tricarboxylic Acid Cycle Rate
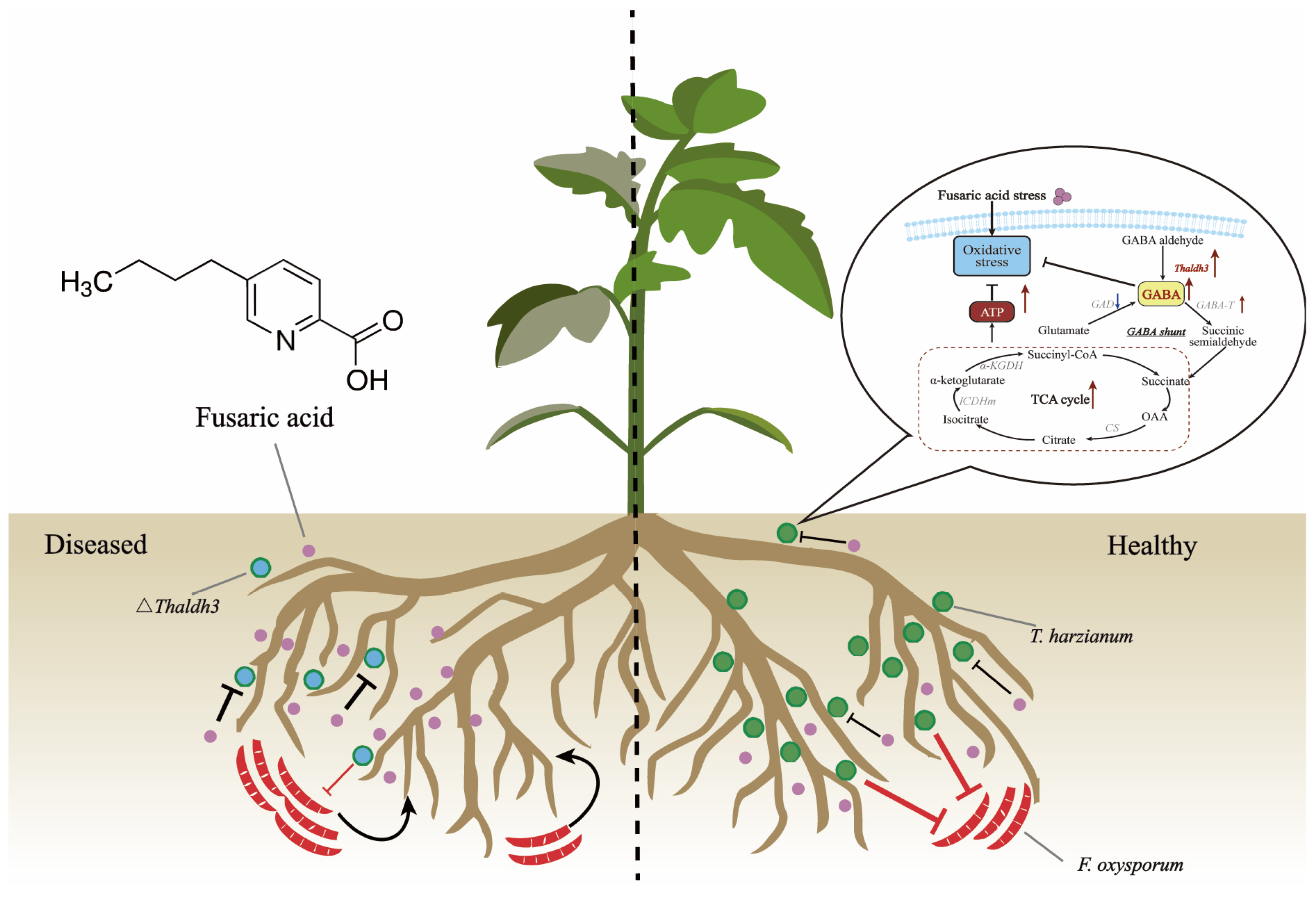
3.6. Enhancing Th Resistance to FSA Is Beneficial for Reducing the Occurrence of Plant Diseases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Munkvold, G.P. Fusarium species and their associated mycotoxins. Mycotoxigenic Fungi Methods Protoc. 2017, 1542, 51–106. [Google Scholar]
- Bakker, M.G.; Acharya, J.; Moorman, T.B.; Robertson, A.E.; Kaspar, T.C. The potential for cereal rye cover crops to host corn seedling pathogens. Phytopathology 2016, 106, 591–601. [Google Scholar] [CrossRef]
- Ma, L.-J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef]
- Kokkonen, M.; Ojala, L.; Parikka, P.; Jestoi, M. Mycotoxin production of selected Fusarium species at different culture conditions. Int. J. Food Microbiol. 2010, 143, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol. Plant-Microbe Interact. 1995, 8, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Marroquín-Cardona, A.; Johnson, N.; Phillips, T.; Hayes, A. Mycotoxins in a changing global environment–a review. Food Chem. Toxicol. 2014, 69, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public health impacts of foodborne mycotoxins. Annu. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef]
- Liu, S.; Li, J.; Zhang, Y.; Liu, N.; Viljoen, A.; Mostert, D.; Zuo, C.; Hu, C.; Bi, F.; Gao, H.; et al. Fusaric acid instigates the invasion of banana by Fusarium oxysporum f. sp. cubense TR 4. New Phytol. 2020, 225, 913–929. [Google Scholar] [CrossRef]
- D’Alton, A.; Etherton, B. Effects of fusaric acid on tomato root hair membrane potentials and ATP levels. Plant Physiol. 1984, 74, 39–42. [Google Scholar] [CrossRef]
- Telles-Pupulin, A.R.; Diniz, S.; Bracht, A.; Ishii-Iwamoto, E. Effects of fusaric acid on respiration in maize root mitochondria. Biol. Plant. 1996, 38, 421–429. [Google Scholar] [CrossRef]
- Niehaus, E.-M.; Diaz-Sanchez, V.; Von Bargen, K.; Kleigrewe, K.; Humpf, H.-U.; Limón, M.C. Fusarins and fusaric acid in fusaria. In Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2014; pp. 239–262. [Google Scholar]
- Gxasheka, M.; Wang, J.; Gunya, B.; Mbanjwa, V.; Tyasi, T.L.; Dlamini, P.; Gao, J. In vitro effect of some commercial fungicides on mycelial growth of Fusarium species causing maize ear rot disease in China. Arch. Phytopathol. Plant Prot. 2021, 54, 557–569. [Google Scholar] [CrossRef]
- Chen, L.-H.; Cui, Y.-Q.; Yang, X.-M.; Zhao, D.-K.; Shen, Q.-R. An antifungal compound from Trichoderma harzianum SQR-T037 effectively controls Fusarium wilt of cucumber in continuously cropped soil. Australas. Plant Pathol. 2012, 41, 239–245. [Google Scholar] [CrossRef]
- Xu, S.; Liu, Y.-X.; Cernava, T.; Wang, H.; Zhou, Y.; Xia, T.; Cao, S.; Berg, G.; Shen, X.-X.; Wen, Z.; et al. Fusarium fruiting body microbiome member Pantoea agglomerans inhibits fungal pathogenesis by targeting lipid rafts. Nat. Microbiol. 2022, 7, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Fan, X.; Wang, Y.; Kusstatscher, P.; Duan, J.; Wu, S.; Chen, S.; Qiao, K.; Wang, Y.; Ma, B.; et al. Bacterial seed endophyte shapes disease resistance in rice. Nat. Plants 2021, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.; Wang, Z.; Liu, S.; Lv, N.; Deng, X.; Xiong, W.; Shen, Z.; Zhang, N.; Geisen, S.; Li, R.; et al. Additive fungal interactions drive biocontrol of Fusarium wilt disease. New Phytol. 2023, 238, 1198–1214. [Google Scholar] [CrossRef] [PubMed]
- Marzano, M.; Gallo, A.; Altomare, C. Improvement of biocontrol efficacy of Trichoderma harzianum vs. Fusarium oxysporum f. sp. lycopersici through UV-induced tolerance to fusaric acid. Biol. Control 2013, 67, 397–408. [Google Scholar] [CrossRef]
- Karlovsky, P. Secondary metabolites in soil ecology. In Secondary Metabolites in Soil Ecology; Springer: Berlin/Heidelberg, Germany, 2008; pp. 1–19. [Google Scholar]
- Meca, G.; Soriano, J.; Gaspari, A.; Ritieni, A.; Moretti, A.; Mañes, J. Antifungal effects of the bioactive compounds enniatins A, A1, B, B1. Toxicon 2010, 56, 480–485. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, L. Beauvericin, a bioactive compound produced by fungi: A short review. Molecules 2012, 17, 2367–2377. [Google Scholar] [CrossRef]
- Woo, S.L.; Hermosa, R.; Lorito, M.; Monte, E. Trichoderma: A multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nat. Rev. Microbiol. 2023, 21, 312–326. [Google Scholar] [CrossRef]
- Zhang, J.; Miao, Y.; Rahimi, M.J.; Zhu, H.; Steindorff, A.; Schiessler, S.; Cai, F.; Pang, G.; Chenthamara, K.; Xu, Y.; et al. Guttation capsules containing hydrogen peroxide: An evolutionarily conserved NADPH oxidase gains a role in wars between related fungi. Environ. Microbiol. 2019, 21, 2644–2658. [Google Scholar] [CrossRef]
- Zhang, J.; Akcapinar, G.B.; Atanasova, L.; Rahimi, M.J.; Przylucka, A.; Yang, D.; Kubicek, C.P.; Zhang, R.; Shen, Q.; Druzhinina, I.S. The neutral metallopeptidase NMP1 of Trichoderma guizhouense is required for mycotrophy and self-defence. Environ. Microbiol. 2016, 18, 580–597. [Google Scholar] [CrossRef]
- Tripathi, P.; Singh, P.C.; Mishra, A.; Chauhan, P.S.; Dwivedi, S.; Bais, R.T.; Tripathi, R.D. Trichoderma: A potential bioremediator for environmental clean up. Clean Technol. Environ. Policy 2013, 15, 541–550. [Google Scholar] [CrossRef]
- Shelp, B.J.; Aghdam, M.S.; Flaherty, E.J. γ-Aminobutyrate (GABA) regulated plant defense: Mechanisms and opportunities. Plants 2021, 10, 1939. [Google Scholar] [CrossRef]
- Zhao, L.; Shu, Y.; Xiao, J.; Lin, R.; Godana, E.A.; Zhang, X.; Zhang, H. Transcriptome analysis reveals mechanisms involved in the enhanced antagonistic efficacy of Sporidiobolus pararoseus Y16 treated by γ-aminobutyric acid. Biol. Control 2022, 176, 105089. [Google Scholar] [CrossRef]
- Ji, H.M.; Mao, H.; Li, S.; Feng, T.; Zhang, Z.; Cheng, L.; Luo, S.; Borkovich, K.A.; Ouyang, S. Fol-milR1, a pathogenicity factor of Fusarium oxysporum, confers tomato wilt disease resistance by impairing host immune responses. New Phytol. 2021, 232, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, N.; Keller, N.P. Mycotoxins in conversation with bacteria and fungi. Front. Microbiol. 2019, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Jia, H.; Ran, L.; Wu, F.; Liu, J.; Schlaeppi, K.; Dini-Andreote, F.; Wei, Z.; Zhou, X. Fusaric acid mediates the assembly of disease-suppressive rhizosphere microbiota via induced shifts in plant root exudates. Nat. Commun. 2024, 15, 5125. [Google Scholar] [CrossRef]
- Ianiri, G.; Idnurm, A.; Wright, S.A.I.; Durán-Patrón, R.; Mannina, L.; Ferracane, R.; Ritieni, A.; Castoria, R. Searching for genes responsible for patulin degradation in a biocontrol yeast provides insight into the basis for resistance to this mycotoxin. Appl. Environ. Microbiol. 2013, 79, 3101–3115. [Google Scholar] [CrossRef]
- Magan, N.; Marcon, A.G.; Samsudin, N.I.P.; Rodríguez-Sixtos, A.; Garcia-Cela, E.; Verheecke-Vaessen, C.; Medina, A. Biological control agents for mycotoxin control: Are they resilient enough? In How Research Can Stimulate the Development of Commercial Biological Control Against Plant Diseases; Springer: Cham, Switzerland, 2020; pp. 295–309. [Google Scholar]
- Ianiri, G.; Idnurm, A.; Castoria, R. Transcriptomic responses of the basidiomycete yeast Sporobolomyces sp. to the mycotoxin patulin. BMC Genom. 2016, 17, 210. [Google Scholar] [CrossRef]
- Sharma, V.; Bhandari, P.; Singh, B.; Bhatacharya, A.; Shanmugam, V. Chitinase expression due to reduction in fusaric acid level in an antagonistic Trichoderma harzianum S17TH. Indian J. Microbiol. 2013, 53, 214–220. [Google Scholar] [CrossRef]
- Singh, V.K.; Upadhyay, R.S. Fusaric acid induced cell death changes in oxidative metabolism of Solanum lycopersicum L. Bot. Stud. 2014, 55, 66. [Google Scholar]
- Devnarain, N.; Tiloke, C.; Nagiah, S.; Chuturgoon, A.A. Fusaric acid induces oxidative stress and apoptosis in human cancerous oesophageal SNO cells. Toxicon 2017, 126, 4–11. [Google Scholar] [CrossRef]
- Jackson, B.; Brocker, C.; Thompson, D.C.; Black, W.; Vasiliou, K.; Nebert, D.W.; Vasiliou, V. Update on the aldehyde dehydrogenase gene (ALDH) superfamily. Hum. Genom. 2011, 5, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Shortall, K.; Durack, E.; Magner, E.; Soulimane, T. Study of ALDH from Thermus thermophilus—Expression, Purification and Characterisation of the Non-Substrate Specific, Thermophilic Enzyme Displaying Both Dehydrogenase and Esterase Activity. Cells 2021, 10, 3535. [Google Scholar] [CrossRef] [PubMed]
- Brocker, C.; Vasiliou, M.; Carpenter, S.; Carpenter, C.; Zhang, Y.; Wang, X.; Kotchoni, S.O.; Wood, A.J.; Kirch, H.-H.; Kopečný, D.; et al. Aldehyde dehydrogenase (ALDH) superfamily in plants: Gene nomenclature and comparative genomics. Planta 2013, 237, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Michaeli, S.; Fait, A.; Lagor, K.; Nunes-Nesi, A.; Grillich, N.; Yellin, A.; Bar, D.; Khan, M.; Fernie, A.R.; Turano, F.J.; et al. A mitochondrial GABA permease connects the GABA shunt and the TCA cycle, and is essential for normal carbon metabolism. Plant J. 2011, 67, 485–498. [Google Scholar] [CrossRef]
- Li, L.; Dou, N.; Zhang, H.; Wu, C. The versatile GABA in plants. Plant Signal. Behav. 2021, 16, 1862565. [Google Scholar] [CrossRef]
- Deng, H.; Chen, J.; Gao, R.; Liao, X.; Cai, Y. Adaptive Responses to Oxidative Stress in the Filamentous Fungal Shiraia bambusicola. Molecules 2016, 21, 1118. [Google Scholar] [CrossRef]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Wang, P.; Lopes, L.D.; Lopez-Guerrero, M.G.; van Dijk, K.; Alvarez, S.; Riethoven, J.-J.; Schachtman, D.P.; Xu, G. Natural variation in root exudation of GABA and DIMBOA impacts the maize root endosphere and rhizosphere microbiomes. J. Exp. Bot. 2022, 73, 5052–5066. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, L.; Shi, X.; Li, T.; Liu, Y.; Wang, T.; Lin, B.; Liu, D.; Shen, Q. Thaldh3-Dependent GABA Metabolism Modulates Response of Trichoderma to Fusaric Acid-Induced Oxidative Stress. J. Fungi 2025, 11, 542. https://doi.org/10.3390/jof11070542
Cao L, Shi X, Li T, Liu Y, Wang T, Lin B, Liu D, Shen Q. Thaldh3-Dependent GABA Metabolism Modulates Response of Trichoderma to Fusaric Acid-Induced Oxidative Stress. Journal of Fungi. 2025; 11(7):542. https://doi.org/10.3390/jof11070542
Chicago/Turabian StyleCao, Linhua, Xiaoteng Shi, Tuo Li, Yang Liu, Tuokai Wang, Bozheng Lin, Dongyang Liu, and Qirong Shen. 2025. "Thaldh3-Dependent GABA Metabolism Modulates Response of Trichoderma to Fusaric Acid-Induced Oxidative Stress" Journal of Fungi 11, no. 7: 542. https://doi.org/10.3390/jof11070542
APA StyleCao, L., Shi, X., Li, T., Liu, Y., Wang, T., Lin, B., Liu, D., & Shen, Q. (2025). Thaldh3-Dependent GABA Metabolism Modulates Response of Trichoderma to Fusaric Acid-Induced Oxidative Stress. Journal of Fungi, 11(7), 542. https://doi.org/10.3390/jof11070542