
Five New Hypocrealean Species from Algae and Sediment in the Intertidal Zones of China



Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Fungal Isolation
2.2. Morphological Observation
2.3. DNA Extraction, PCR Amplification and Sequencing
2.4. Evolutionary Lineage Modeling
3. Results
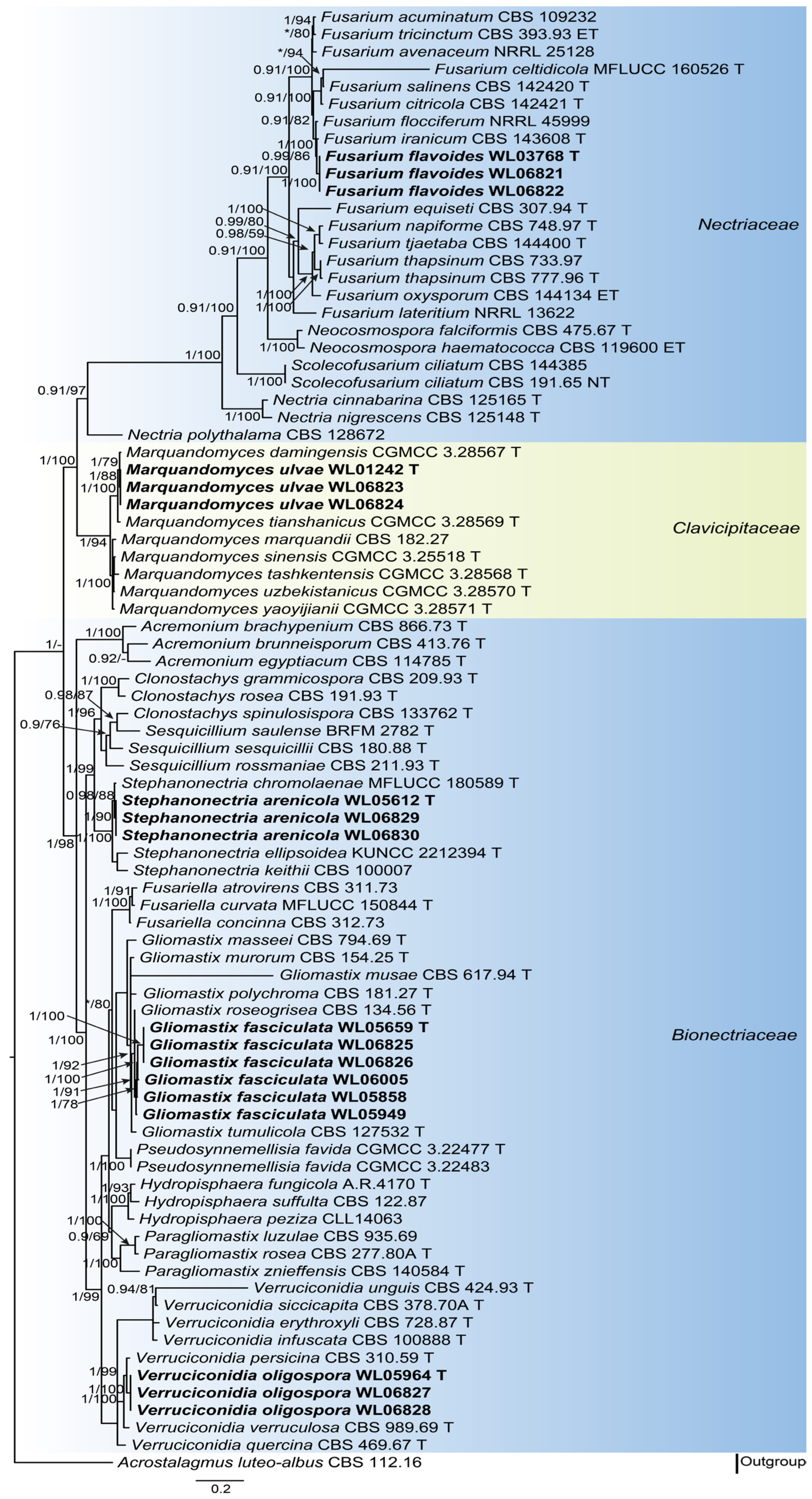
3.1. Phylogenetic Analyses
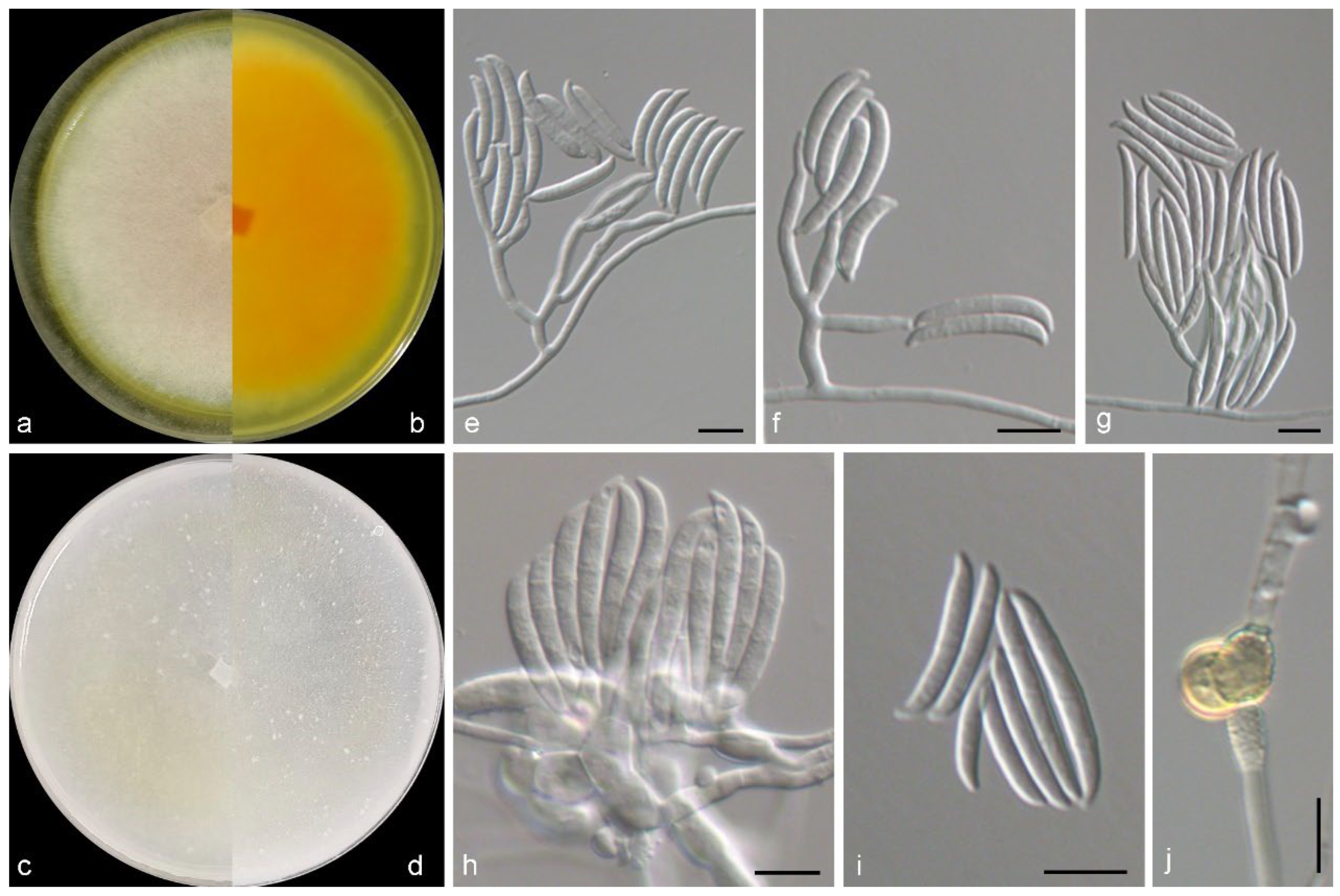
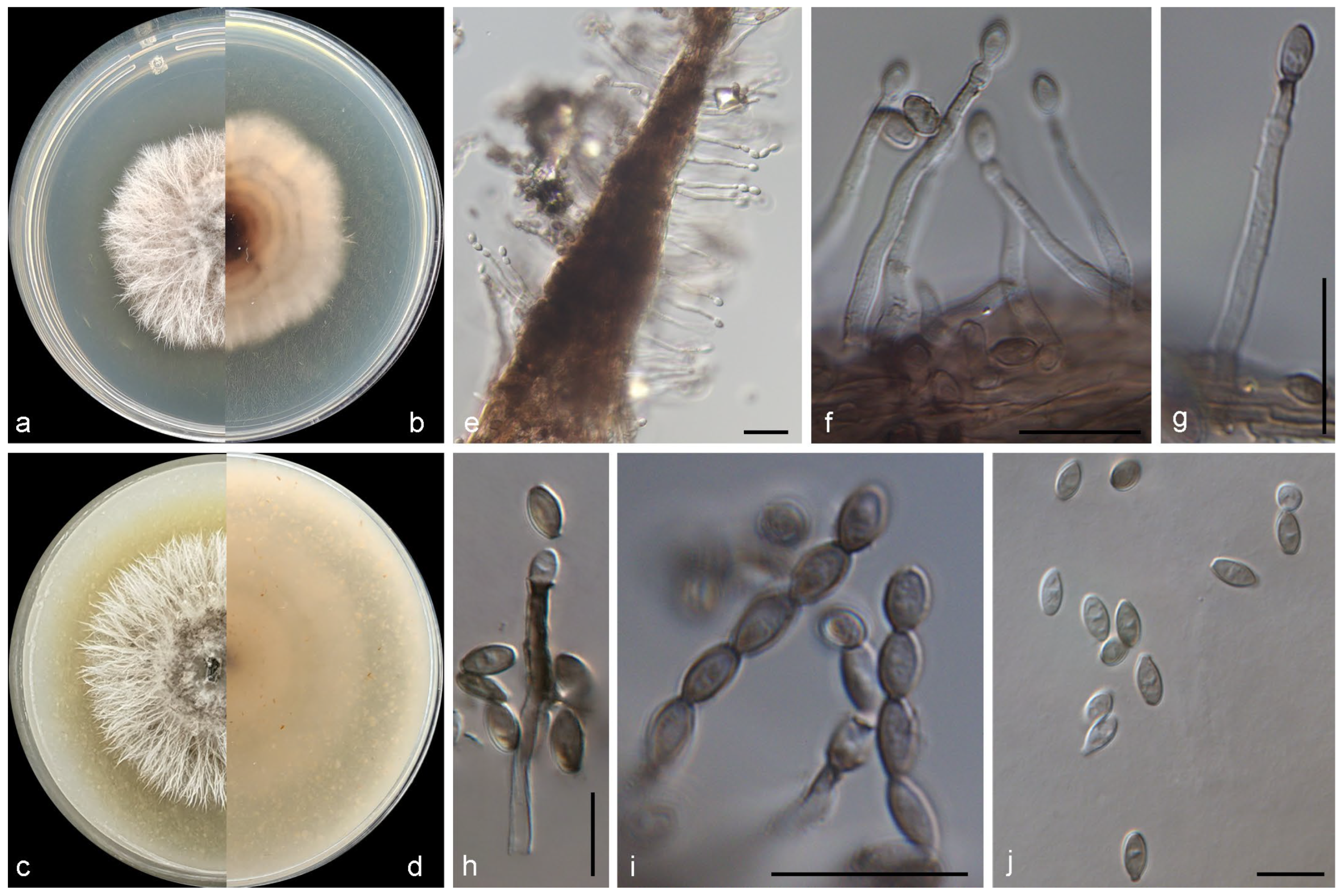
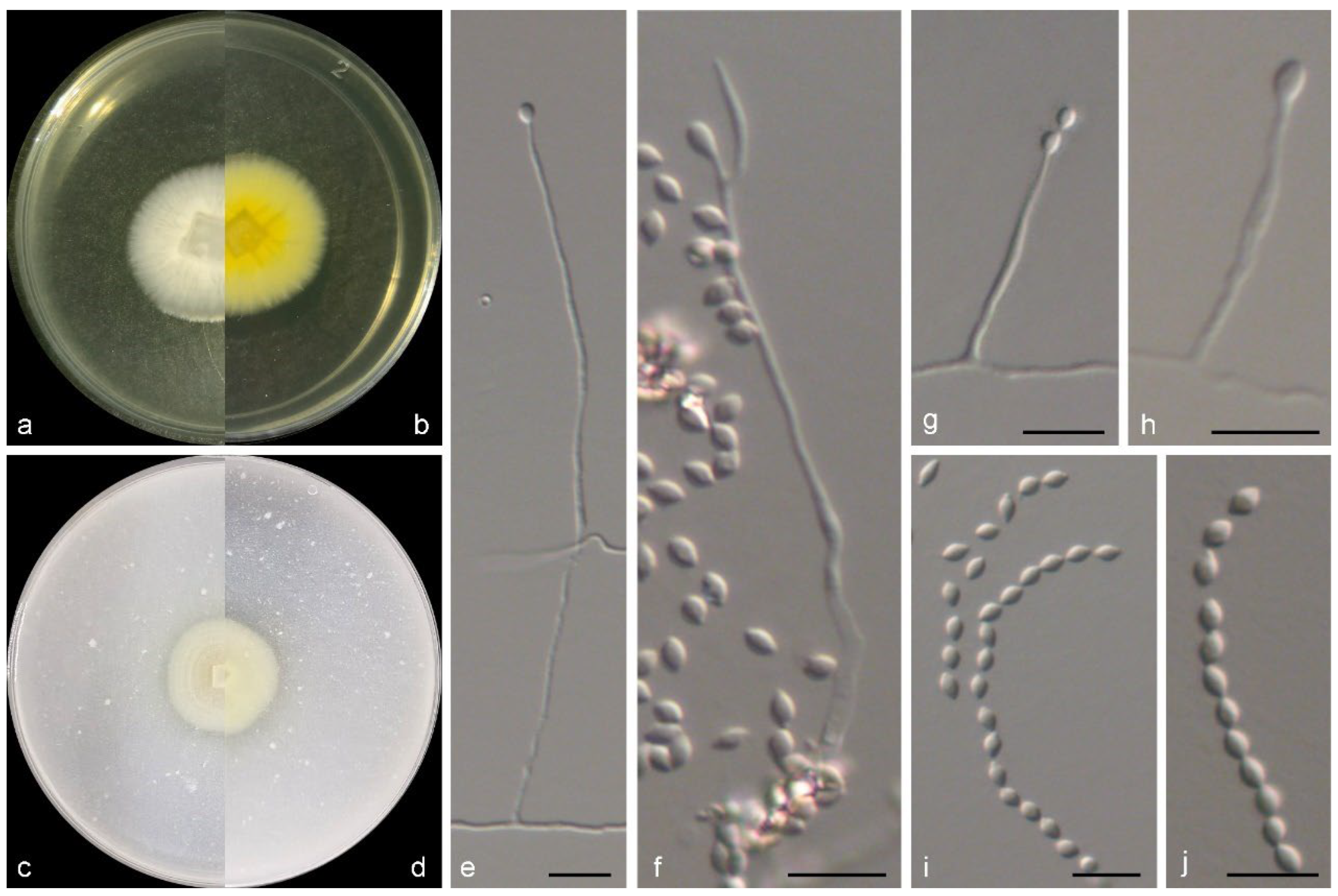
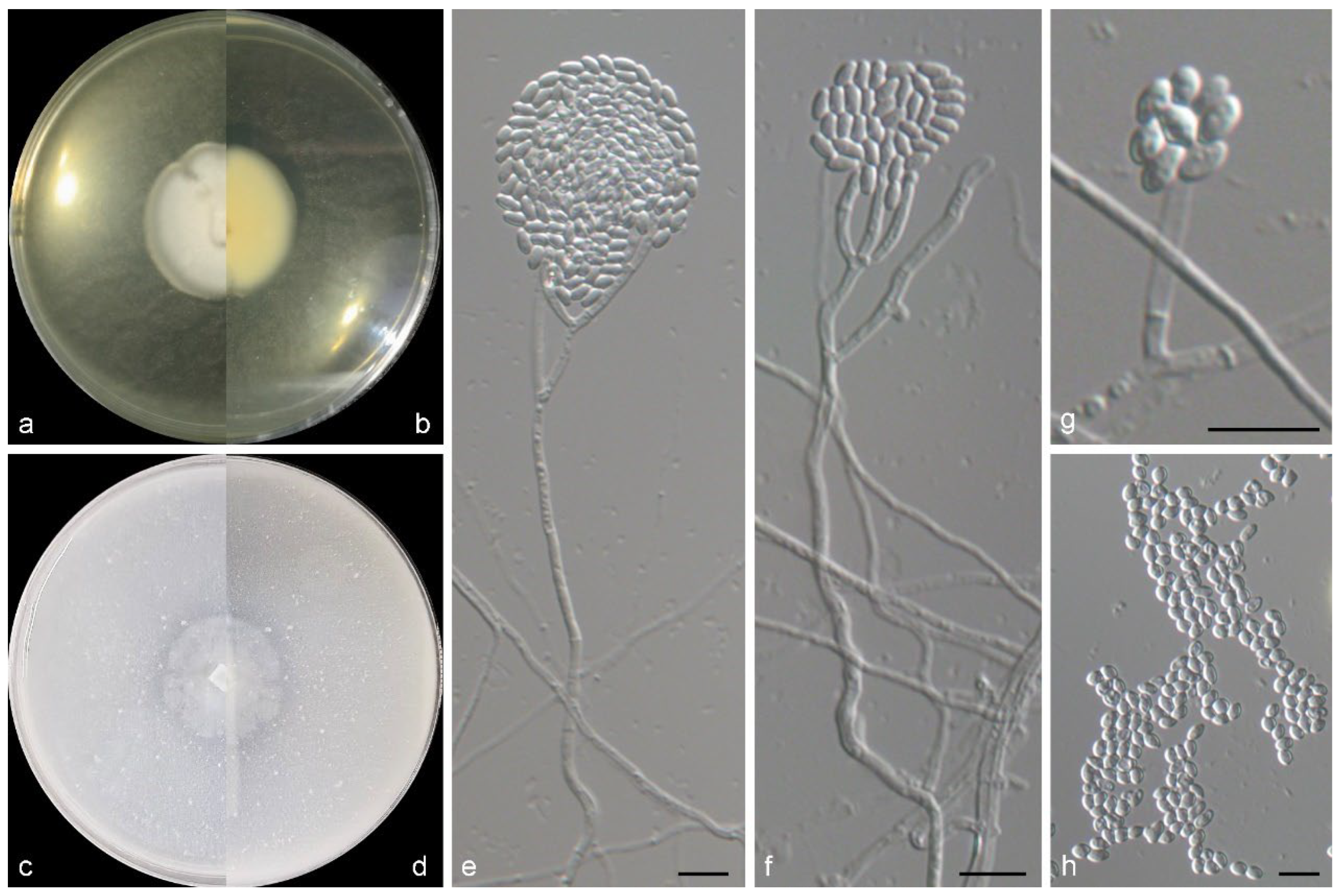
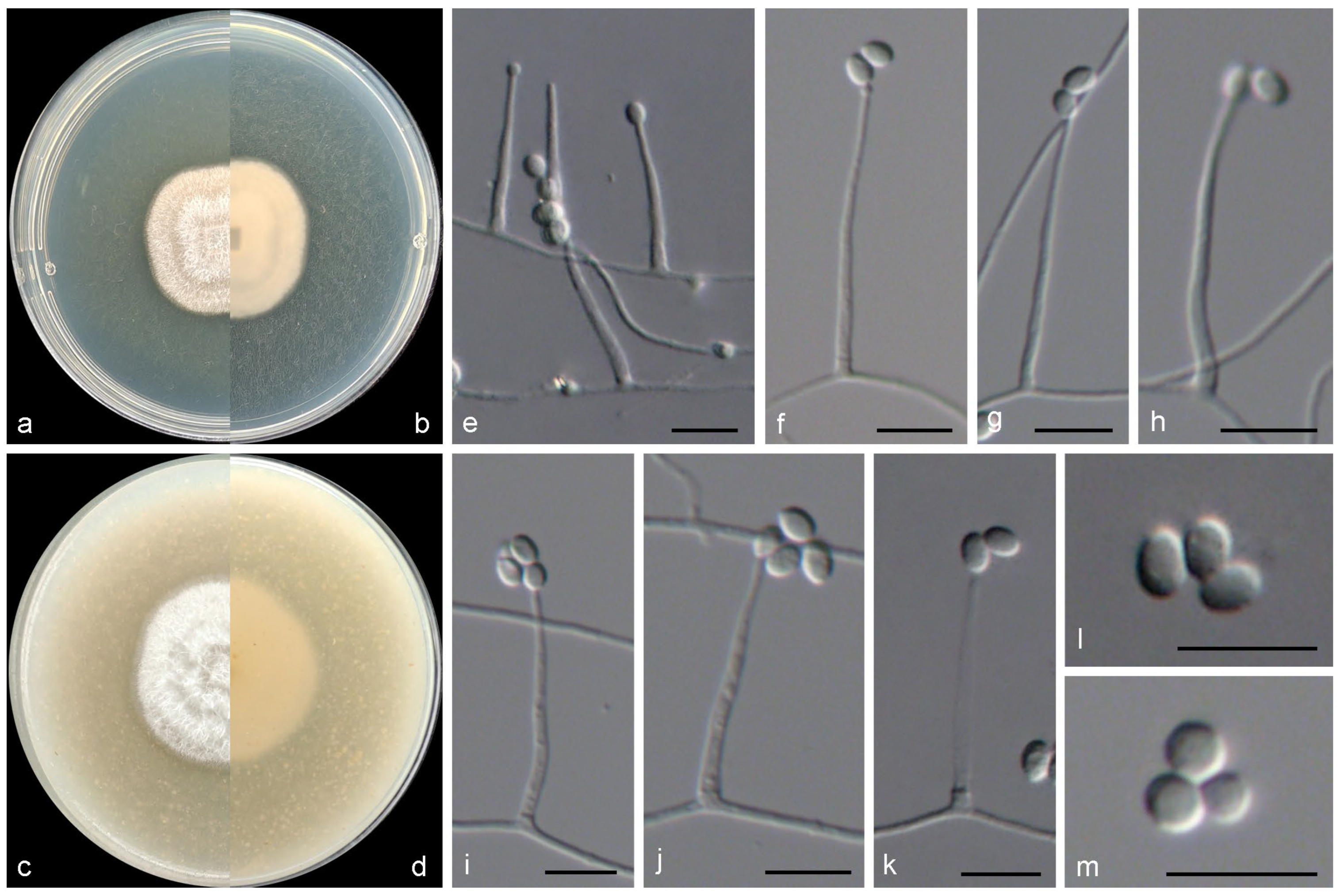
3.2. Taxonomy of the Novel Species
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Wijayawardene, N.; Hyde, K.; Dai, D.; Sánchez-García, M.; Goto, B.; Saxena, R.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Rogerson, C.T. The Hypocrealean Fungi (Ascomycetes, Hypocreales). Mycologia 1970, 62, 865–910. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Houbraken, J.; Varga, J.; Frisvad, J.C. Polyphasic taxonomy of the heat resistant ascomycete genus Byssochlamys and its Paecilomyces anamorphs. Persoonia 2009, 22, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Chaverri, P.; Salgado, C.; Hirooka, Y.; Rossman, A.Y.; Samules, G.J. Delimitation of Neonectria and Cylindrocarpon (Nectriaceae, Hypocreales, Ascomycota) and related genera with Cylindrocarpon-like anamorphs. Stud. Mycol. 2011, 68, 57–78. [Google Scholar] [CrossRef]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef]
- Hou, L.W.; Giraldo, A.; Groenewald, J.Z.; Rämä, T.; Summerbell, R.C.; Huang, G.Z.; Cai, L.; Crous, P.W. Redisposition of acremonium-like fungi in Hypocreales. Stud. Mycol. 2023, 105, 23–203. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.; Schroers, H.-J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef]
- Jones, E.B.G. Fifty years of marine mycology. Fungal Divers. 2011, 50, 73–112. [Google Scholar] [CrossRef]
- Morrison-Gardiner, S. Dominant fungi from Australian coral reefs. Fungal Divers. 2002, 9, 105–121. [Google Scholar]
- Zalar, P.; de Hoog, G.; Schroers, H.-J.; Crous, P.; Groenewald, J.; Gunde-Cimerman, N. Phylogeny and ecology of the ubiquitous saprobe Cladosporium sphaeros-permum, with descriptions of seven new species from hypersaline environments. Stud. Mycol. 2007, 58, 157–183. [Google Scholar] [CrossRef]
- Li, Q.; Wang, G. Diversity of fungal isolates from three Hawaiian marine sponges. Microb. Res. 2007, 164, 233–241. [Google Scholar] [CrossRef]
- Hong, J.-H.; Jang, S.; Heo, Y.M.; Min, M.; Lee, H.; Lee, Y.M.; Lee, H.; Kim, J.-J. Investigation of marine-derived fungal diversity and their exploitable biological activities. Mar. Drugs 2015, 13, 4137–4155. [Google Scholar] [CrossRef]
- Shaumi, A.; Cheng, U.-C.; Guo, S.Y.; Jones, E.B.G.; Chan, T.-Y.; Pang, K.-L. Diversity of fungi isolated from carapace and gut of the marine crab Portunus sanguinolentus in northern waters of Taiwan. Bot. Mar. 2023, 66, 301–307. [Google Scholar] [CrossRef]
- Li, M.; Raza, M.; Song, S.; Hou, L.; Zhang, Z.-F.; Gao, M.; Huang, J.-E.; Liu, F.; Cai, L. Application of culturomics in fungal isolation from mangrove sediments. Microbiome 2023, 11, 272. [Google Scholar] [CrossRef]
- Wang, X.; Ji, G.; Cun, J.; Xu, P.; Wang, X.; Ren, G.; Li, W. Screening of insecticidal and antifungal activities of the culturable fungi isolated from the intertidal zones of Qingdao, China. J. Fungi 2022, 8, 1240. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.M.; Ma, Y.Y.; Cai, L.; Tedersoo, L.; Bahram, M.; Burgaud, G.; Long, X.D.; Zhang, S.M.; Li, W. Seasonal dynamics of mycoplankton in the Yellow Sea reflect the combined effect of riverine inputs and hydrographic conditions. Mol. Ecol. 2021, 30, 3624–3637. [Google Scholar] [CrossRef]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Cheng, X.L.; Li, W.; Zhang, T.Y. A new species of Phaeoisaria from intertidal marine sediment collected in Weihai, China. Mycotaxon 2014, 127, 17–24. [Google Scholar] [CrossRef]
- Cheng, X.L.; Li, W.; Cai, L. Molecular phylogeny of Ascotricha, including two new marine algaeassociated species. Mycologia 2015, 107, 490–504. [Google Scholar] [CrossRef]
- Wang, M.-M.; Shenoy, B.D.; Li, W.; Cai, L. Molecular phylogeny of Neodevriesia, with two new species and several new combinations. Mycologia 2019, 109, 965–974. [Google Scholar] [CrossRef]
- Wang, M.-M.; Yang, S.-Y.; Li, Q.; Zheng, Y.; Ma, H.-H.; Tu, Y.-H.; Li, W.; Cai, L. Microascaceae from the marine environment, with descriptions of six new species. J. Fungi 2024, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978. [Google Scholar]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 2000, 147, 617–630. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to the Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Sun, B.L. Ancient and recent patterns of geographic speciation in the oyster mushroom Pleurotus revealed by phylogenetic analysis of ribosomal DNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 4599–4603. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-alpha sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Sung, G.-H.; Hywel-Jones, N.L.; Sung, J.-M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef]
- Wang, M.M.; Crous, P.W.; Sandoval-Denis, M.; Han, S.L.; Liu, F.; Liang, J.M.; Duan, W.J.; Cai, L. Fusarium and allied genera from China: Species diversity and distribution. Persoonia 2022, 48, 1–53. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY (are we there yet?): A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2008, 24, 581–583. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate Maximum-Likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Booth, C. The Genus Fusarium; Commonwealth Mycological Institute: Kew, Surrey, 1971. [Google Scholar]
- Gerlach, W.; Nirenberg, H. The genus Fusarium—A pictorial atlas. In Mitteilungen aus der Biologischen Bundesanstalt für Land- und Forstwirtschaft Berlin-Dahlem; Kommissionsverlag P. Parey: Berlin, Germany, 1982; Volume 209, pp. 1–406. [Google Scholar]
- Sandoval-Denis, M.; Guarnaccia, V.; Polizzi, G.; Crous, P. Symptomatic Citrus trees reveal a new pathogenic lineage in Fusarium and two new Neocosmospora species. Persoonia 2018, 40, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Torbati, M.; Arzanlou, M.; Sandoval-Denis, M.; Crous, P.W. Multigene phylogeny reveals new fungicolous species in the Fusarium tricinctum species complex and novel hosts in the genus Fusarium from Iran. Mycol. Prog. 2019, 18, 119–133. [Google Scholar] [CrossRef]
- Corda, A.C.J. Deutschlands Flora, Abt. III. Die Pilze Dtschl. 1828, 2, 1–35. [Google Scholar]
- Torbati, M.; Arzanlou, M.; da Silva Santos, A.C. Fungicolous Fusarium Species: Ecology, Diversity, Isolation, and Identification. Curr. Microbiol. 2021, 78, 2850–2859. [Google Scholar] [CrossRef]
- Klittich, C.J.R.; Leslie, J.F.; Nelson, P.E.; Marasas, W.F.O. Fusarium thapsinum (Gibberella thapsina): A new species in section Liseola from sorghum. Mycologia 1997, 89, 643–652. [Google Scholar] [CrossRef]
- Matsushima, T. Icones Microfungorum a Matsushima Lectorum; The Nippon Printing & Publishing Co., Ltd.: Tokyo, Japan, 1975; pp. 1–209. [Google Scholar]
- de Hoog, G.S. Atlas of Clinical Fungi; Centraalbureau voor Schimmelcultures (CBS): Utrecht, The Netherlands, 2000; pp. 1–1126. [Google Scholar]
- Summerbell, R.C.; Gueidan, C.; Schroers, H.-J.; de Hoog, G.S.; Starink, M.; Arocha Rosete, Y.; Guarro, J.; Scott, J.A. Acremonium phylogenetic overview and revision of Gliomastix, Sarocladium, and Trichothecium. Stud. Mycol. 2011, 68, 139–162. [Google Scholar] [CrossRef]
- Mongkolsamrit, S.; Khonsanit, A.; Thanakitpipattana, D.; Tasanathai, K.; Noisripoom, W.; Lamlertthon, S.; Himaman, W.; Houbraken, J.; Samson, R.A.; Luangsa-ard, J. Revisiting Metarhizium and the description of new species from Thailand. Stud. Mycol. 2020, 95, 171–251. [Google Scholar] [CrossRef]
- Peng, L.-Y.; Wang, Y.-F.; Song, H.; Urinboev, I.; Zhuang, W.-Y.; Gafforov, Y.; Wang, X.-C. Five new species of Marquandomyces (Clavicipitaceae, Ascomycota) from Asia. J. Fungi 2025, 11, 180. [Google Scholar] [CrossRef] [PubMed]
- Schroers, H.-J.; Samuels, G.J.; Gams, W. Stephanonectria, a new genus of the Hypocreales (Bionectriaceae), and its sporodochial anamorph. Sydowia 1999, 51, 114–126. [Google Scholar]
- He, S.C.; Wei, D.P.; Bhunjun, C.S.; Zhao, Q.; Jayawardena, R.S. A new species of Stephanonectria (Bionectriaceae) from southwestern China. Asian J. Mycol. 2023, 6, 98–106. [Google Scholar] [CrossRef]
- Perera, R.H.; Hyde, K.D.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Bundhun, D.; Camporesi, E.; Akulov, A.; Liu, J.K.; Liu, Z.Y. Profile of Bionectriaceae, Calcarisporiaceae, Hypocreaceae, Nectriaceae, Tilachlidiaceae, Ijuhyaceae fam. nov., Stromatonectriaceae fam. nov. and Xanthonectriaceae fam. nov. Fungal Divers. 2023, 118, 95–271. [Google Scholar] [CrossRef]
- Hyde, K.D.; Noorabadi, M.T.; Thiyagaraja, V.; He, M.Q.; Johnston, P.R.; Wijesinghe, S.N.; Armand, A.; Biketova, A.Y.; Chethana, K.W.T.; Erdoğdu, M.; et al. The 2024 Outline of Fungi and fungus-like taxa. Mycosphere 2024, 15, 5146–6239. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E.; Casper, H.H. Molecular, phylogenetic morphological and mycotoxin data support reidentification of the Quorn mycoprotein fungus as Fusarium venenatum. Fungal Genet. Biol. 1998, 23, 57–67. [Google Scholar] [CrossRef]
- Dimarogona, M.; Nikolaivits, E.; Kanelli, M.; Christakopoulos, P.; Sandgren, M.; Topakas, E. Structural and functional studies of a Fusarium oxysporum cutinase with polyethylene terephthalate modification potential. Biochim. Biophys. Acta-Gen. Subj. 2015, 11, 2308–2317. [Google Scholar] [CrossRef]
- Fernández-Remacha, D.; González-Riancho, C.; Osua, M.L.; Arce, A.G.; Montánchez, I.; García-Lobo, J.M.; Estrada-Tejedor, R.; Kaberdin, V.R. Analysis of laccase-like enzymes secreted by fungi isolated from a cave in northern Spain. MicrobiologyOpen 2022, 11, e1279. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Keyhani, N.O.; Xia, Y.X.; Xie, J.Q. The potential and limitations of entomopathogenic fungi as biocontrol agents for insect pest management. Entomol. Gen. 2024, 44, 797–811. [Google Scholar] [CrossRef]
- Huo, R.; Tu, Y.; Liu, C.; Zi, G.; Shi, Y.; Ren, J.; Cai, L.; Liu, L. New antifungal chlorinated orsellinic aldehydes from the deep-sea-derived fungus Acremonium sclerotigenum LW14. Mycology 2025, 1–10. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Pang, K.-L.; Abdel-Wahab, M.A.; Scholz, B.; Hyde, K.D.; Boekhout, T.; Ebel, R.; Rateb, M.E.; Henderson, L.; Sakayaroj, J.; et al. An online resource for marine fungi. Fungal Divers. 2019, 96, 347–433. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Devadatha, B.; Abdel-Wahab, M.A.; Dayarathne, M.C.; Zhang, S.-N.; Hyde, K.D.; Liu, J.-K.; Bahkali, A.H.; Sarma, V.V.; Tibell, S.; et al. Phylogeny of new marine Dothideomycetes and Sordariomycetes from mangroves and deep-sea sediments. Bot. Mar. 2020, 63, 155–181. [Google Scholar] [CrossRef]
- Cecchi, G.; Cutroneo, L.; Piazza, S.D.; Capello, M.; Zotti, M. Culturable fungi from dredged and marine sediments from six ports studied in the framework of the SEDITERRA Project. J. Soils Sediments 2021, 21, 1563–1573. [Google Scholar] [CrossRef]
- Georgieva, M.L.; Bilanenko, E.N.; Georgiev, A.A.; Bubnova, E.N. Filamentous fungi in the sediments of the East Siberian and Laptev Seas. Microbiology 2024, 93, 364–368. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain Number and Status | Habitat/Host | Origin | GenBank Accession Number | |||
|---|---|---|---|---|---|---|---|
| ITS | LSU | tef1 | rpb2 | ||||
| Acremonium brachypenium | CBS 866.73 T | Cocos nucifera | Sri Lanka | OQ429443 | OQ055354 | OQ470740 | OQ453837 |
| A. brunneisporum | CBS 413.76 T | Colletotrichum dematium on pod of Albizzia lebbek | India | OQ429444 | OQ055355 | OQ470741 | OQ453838 |
| A. egyptiacum | CBS 114785 T | Ground | Egypt | OQ429456 | OQ055362 | OQ470749 | OQ453845 |
| Acrostalagmus luteo-albus | CBS 112.16 | -- | UK | LR026668 | LR025797 | LR026369 | LR026101 |
| Clonostachys grammicospora | CBS 209.93 T | Trunk of dead standing tree | French Guiana | OQ910625 | OQ910984 | OQ944637 | OQ927696 |
| C. spinulosispora | CBS 133762 T | Astrocarium sp. | French Guiana | MH634702 | KY006568 | -- | -- |
| C. vacuolata | CBS 191.93 T | Bark | Venezuela, Edo | OQ910868 | OQ911227 | OQ944876 | OQ927931 |
| Fusariella atrovirens | CBS 311.73 | Desert soil | Algeria | OQ429594 | OR052105 | OQ470904 | OQ453993 |
| Fu. concinna | CBS 312.73 | Desert soil | Algeria | OQ429595 | OQ055505 | OQ470905 | OQ453994 |
| Fu. curvata | MFLUCC 15-0844 T | Quercus sp. | Thailand | KX025152 | KX025154 | KX025155 | KX025156 |
| Fusarium acuminatum | NRRL 36147 = CBS 109232 | Man, bronchial secretion | -- | GQ505452 | GQ505452 | GQ505420 | GQ505484 |
| F. avenaceum | NRRL 25128 | Hymenoptera ichneumonidae | Poland | JF740894 | JF740894 | JF740751 | JF741079 |
| F. celtidicola | MFLUCC 16-0526 T | Dead branch of Celtis australis | Italy | MG873469 | MG873466 | MH576581 | ON759296 |
| F. citricola | CPC 27805 = CBS 142421 T | Crown of Citrus reticulata | Italy | LT746245 | LT746245 | LT746197 | LT746310 |
| F. equiseti | NRRL 26419 = BBA 68556 = CBS 307.94 T | Soil | Germany | NR_121457 | -- | GQ505599 | GQ505777 |
| F. flavoides | WL03768 = CGMCC 3.28711 T | Intertidal sediment | China, Shandong Province, Yantai city | PV020684 | PV020702 | PV050414 | PV023180 |
| WL06821 | Intertidal sediment | China, Shandong Province, Yantai city | PV020685 | PV020703 | PV050415 | PV023181 | |
| WL06822 | Intertidal sediment | China, Shandong Province, Yantai city | PV020686 | PV020704 | PV050416 | PV023182 | |
| F. flocciferum | NRRL 45999 | -- | United States CA | GQ505465 | GQ505465 | GQ505433 | GQ505497 |
| F. iranicum | CBS 143608 = CPC 30860 T | Agaricus bisporus | Orumieh-Salmas, Iran | LT970821 | LT970821 | LT970785 | LT970757 |
| F. lateritium | NRRL 13622 | Ulmus sp. | USA | -- | -- | AY707173 | JX171571 |
| F. napiforme | CBS 748.97 T | Pennisetum typhoides | Namibia | KR071645 | KU604071 | AF160266 | KU604233 |
| F. oxysporum | CBS 144134 ET | Solanum tuberosum | Germany | -- | -- | MH485044 | MH484953 |
| F. salinens | CBS 142420 = CPC 26973 T | Citrus sinensis twigs | Italy | LT746241 | LT746241 | LT746193 | LT746306 |
| F. thapsinum | ATCC 200522 = CBS 777.96 = FRC M-6564 T | Stalk of Sorghum sp. | USA | MH862618 | MH874241 | MW928844 | MW928833 |
| CBS 733.97 = NRRL 22045 | -- | -- | KR071690 | KU604054 | AF160270 | JX171600 | |
| F. tricinctum | CBS 393.93 ET | Winter wheat | Germany | HM068317 | HM068317 | AB674263 | JX171629 |
| Gliomastix fasciculata | WL05659 = CGMCC 3.28712 T | Intertidal sediment | China, Shandong Province, Weihai city | PV020693 | PV020711 | PV050422 | -- |
| WL05858 | Intertidal sediment | China, Shandong Province, Qingdao city | PV020690 | PV020708 | -- | -- | |
| WL05949 | Intertidal sediment | China, Liaoning Province, Dalian city | PV020691 | PV020709 | PV050420 | PV023186 | |
| WL06005 | Intertidal sediment | China, Liaoning Province, Huludao city | PV020692 | PV020710 | PV050421 | PV023187 | |
| WL06825 | Intertidal sediment | China, Shandong Province, Weihai city | PV020694 | PV020712 | PV050423 | -- | |
| WL06826 | Intertidal sediment | China, Shandong Province, Weihai city | PV020695 | PV020713 | PV050424 | -- | |
| G. masseei | CBS 794.69 T | Dung of rabbit | Italy | OQ429601 | OQ055510 | OQ470911 | OQ454000 |
| G. musae | CBS 617.94 T | Musa sp. | Colombia | OQ429616 | OQ055523 | OQ470926 | OQ454015 |
| G. murorum | CBS 154.25 T | Malus sylvestris | -- | OQ429613 | HQ232063 | OQ470923 | OQ454012 |
| G. polychroma | CBS 181.27 T | Hevea brasiliensis | Indonesia | OQ429629 | OQ055528 | OQ470931 | OQ454020 |
| G. roseogrisea | CBS 134.56 T | Grassland soil | India | OQ429639 | OQ055545 | OQ470948 | OQ454037 |
| G. tumulicola | CBS 127532 T | White salt-like masses on teh central part of teh painting | Japan | OQ429641 | OQ055547 | OQ470950 | OQ454039 |
| Hydropisphaera fungicola | A.R. 4170 T | Ulocladium atrum | USA | OQ429666 | OR052107 | OQ470973 | OQ454063 |
| H. peziza | CLL 14063 | Dead wood | France | OQ429667 | OQ055572 | OQ470974 | OQ454064 |
| H. suffulta | CBS 122.87 | Cocos nucifera | Indonesia | OQ429672 | OQ055577 | OQ470979 | OQ454068 |
| Marquandomyces damingensis | JJJ73-30 = CGMCC 3.28567 T | Soil | China, Hebei Province | PQ484187 | PQ484201 | PQ469018 | -- |
| M. marquandii | CBS 182.27 | Soil | USA | MH854923 | MH866418 | EF468793 | EF468942 |
| M. sinensis | CGMCC 3.25518 T | Soil | China, Guizhou Province, Guiyang city | OR680543 | OR680610 | OR858937 | OR842958 |
| M. tashkentensis | UZ13-25 = CGMCC 3.28568 T | Soil | Uzbekistan, Tashkent | PQ484189 | PQ484203 | PQ469020 | -- |
| M. tianshanicus | UZ14-25 = CGMCC 3.28569 T | Soil | Uzbekistan, Tashkent | PQ484190 | PQ484204 | PQ469021 | -- |
| M. ulvae | WL01242 = CGMCC 3.28716 T | Ulva spinulosa | China, Guangdong Province, Shenzhen city | PV020687 | PV020705 | PV050417 | PV023183 |
| WL06823 | Ulva spinulosa | China, Guangdong Province, Shenzhen city | PV020688 | PV020706 | PV050418 | PV023184 | |
| WL06824 | Ulva spinulosa | China, Guangdong Province, Shenzhen city | PV020689 | PV020707 | PV050419 | PV023185 | |
| M. uzbekistanicus | UZ11-45 = CGMCC 3.28570 T | Soil | Uzbekistan, Tashkent | PQ484192 | PQ484206 | PQ469023 | -- |
| M. yaoyijianii | HLJ55-10 = CGMCC 3.28571 T | Soil | China, Heilongjiang Province | PQ484193 | PQ484207 | PQ469024 | -- |
| Nectria cinnabarina | CBS 125165 T | Aesculus sp. | France | HM484548 | HM484562 | HM484527 | KM232402 |
| N. nigrescens | CBS 125148 T | Wood | USA | HM484707 | HM484720 | HM484672 | KM232403 |
| N. polythalama | CBS 128672 | Decaying twig | New Zealand | OQ429722 | OQ055623 | OQ471033 | OQ454123 |
| Neocosmospora falciformis | CBS 475.67 = IMI 268681 T | Human mycetoma | Puerto Rico | MG189935 | MG189915 | LT906669 | LT960558 |
| Ne. haematococca | CBS 119600 ET | Dying tree | Sri Lanka | KM231797 | KM231664 | KM231926 | LT960561 |
| Paragliomastix luzulae | CBS 935.69 | Fagus sylvatica | Germany | OQ429774 | OQ055672 | OQ471102 | OQ454185 |
| P. rosea | CBS 277.80A T | -- | India | OQ429775 | OQ055673 | OQ471103 | OQ454186 |
| P. znieffensis | CBS 140584 T | Cyathea sp. | Martinique | OQ429776 | KU198185 | OQ471104 | OQ454187 |
| Pseudosynnemellisia favida | CGMCC 3.22477 = LC15930 T | Sediment | China, Guangdong Province, Shenzhen city | OQ798973 | OQ758164 | OQ809058 | OQ809024 |
| CGMCC 3.22483 = LC15931 | Sediment | China, Guangdong Province, Shenzhen city | OQ798974 | OQ758165 | OQ809059 | OQ809025 | |
| Scolecofusarium ciliatum | CBS 144385 | Leaf of Fagus sylvatica | Belgium | KJ125591 | KJ126479 | MW834297 | KP835472 |
| CBS 191.65 NT | Fagus sylvatica, branch canker | Germany | MW827636 | MW827677 | MW834296 | MW834035 | |
| Sesquicillium rossmaniae | CBS 211.93 T | Twigs of recently dead tree | French Guiana, Piste de Saint-Elie | OQ911299 | OQ911360 | OQ944534 | OQ914852 |
| Se. saulense | BRFM 2782 T | Dead bark of Bauhinia sp. | French Guana, Saül | MK635054 | -- | -- | -- |
| Se. sesquicillii | CBS 180.88 T | Twigs and lichen | Guyana, Cuyuni-Mazaruni Region | OQ911300 | MH873818 | OQ944535 | -- |
| Stephanonectria arenicola | WL05612 = CGMCC 3.28726 T | Intertidal sediment | China, Shandong Province, Weihai city | PV020699 | PV020717 | PV050428 | -- |
| WL06829 | Intertidal sediment | China, Shandong Province, Weihai city | PV020700 | PV020718 | PV050429 | -- | |
| WL06830 | Intertidal sediment | China, Shandong Province, Weihai city | PV020701 | PV020719 | PV050430 | -- | |
| S. chromolaenae | MFLUCC 18-0589 T | Dead stem of Chromolaena odorata | Thailand, Chiang Mai Province | NR_189387 | NG_241983 | -- | -- |
| S. ellipsoidea | KUNCC 22-12394 T | Dried fruit of a woody plant | China, Yunnan Province, Kunming City | OP205363 | OP205375 | -- | -- |
| S. keithii | CBS 100007 | Beilschmiedia tawa | New Zealand | OQ429871 | OQ430120 | OQ471202 | OQ454269 |
| Verruciconidia erythroxyli | CBS 728.87 T | Erythroxylum areolatum | Cuba | OQ429910 | OQ430161 | OQ471240 | OQ454307 |
| V. infuscata | CBS 100888 T | Air | Japan | OQ429911 | OQ430162 | OQ471241 | OQ454308 |
| V. oligospora | WL05964 = CGMCC 3.28727 T | Intertidal sediment | China, Liaoning Province, Dalian city | PV020696 | PV020714 | PV050425 | PV023188 |
| WL06827 | Intertidal sediment | China, Liaoning Province, Dalian city | PV020697 | PV020715 | PV050426 | PV023189 | |
| WL06828 | Intertidal sediment | China, Liaoning Province, Dalian city | PV020698 | PV020716 | PV050427 | PV023190 | |
| V. persicina | CBS 310.59 T | Coastal sand under Ammophila arenaria | France | OQ429921 | OQ430172 | OQ471251 | OQ454318 |
| V. quercina | CBS 469.67 T | Quercus sp. | -- | OQ429925 | OQ430176 | OQ471255 | OQ454322 |
| V. siccicapita | CBS 378.70A T | Soil | Thailand | OQ429928 | OQ430179 | OQ471258 | OQ454325 |
| V. unguis | CBS 424.93 T | Nail of man | Netherlands | OQ429929 | OQ430180 | OQ471259 | OQ454326 |
| V. verruculosa | CBS 989.69 T | Agricultural soil | Netherlands | OQ429933 | OQ430184 | OQ471263 | OQ454330 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.-M.; Mo, W.-Y.; Sun, M.-Y.; Tu, Y.-H.; Li, W. Five New Hypocrealean Species from Algae and Sediment in the Intertidal Zones of China. J. Fungi 2025, 11, 476. https://doi.org/10.3390/jof11070476
Wang M-M, Mo W-Y, Sun M-Y, Tu Y-H, Li W. Five New Hypocrealean Species from Algae and Sediment in the Intertidal Zones of China. Journal of Fungi. 2025; 11(7):476. https://doi.org/10.3390/jof11070476
Chicago/Turabian StyleWang, Meng-Meng, Wang-Ying Mo, Meng-Yi Sun, Ye-Hui Tu, and Wei Li. 2025. "Five New Hypocrealean Species from Algae and Sediment in the Intertidal Zones of China" Journal of Fungi 11, no. 7: 476. https://doi.org/10.3390/jof11070476
APA StyleWang, M.-M., Mo, W.-Y., Sun, M.-Y., Tu, Y.-H., & Li, W. (2025). Five New Hypocrealean Species from Algae and Sediment in the Intertidal Zones of China. Journal of Fungi, 11(7), 476. https://doi.org/10.3390/jof11070476