

The Antifungal Activity and Mechanism of Dehydroabietic Acid Against Alternaria alternata Causing Poplar Leaf Spot

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Pathogens and Spore Suspensions
2.2. Reagents and Plant Samples
2.3. Effect of DHA on Mycelial Growth of Five Pathogenic Fungi
2.4. Effect of DHA on Spore Germination of A. alternata
2.5. In Vivo Control Effect
2.6. Morphological Observation of A. alternata Mycelium
2.7. Effect of DHA on Extracellular Proteins and Electrical Conductivity in A. alternata
2.8. Effect of DHA on MDA and H2O2 in A. alternata
2.9. Effect of DHA on the Enzyme Activities
2.10. Effect of DHA on the Gene Expression
2.11. Statistical Analysis
3. Results
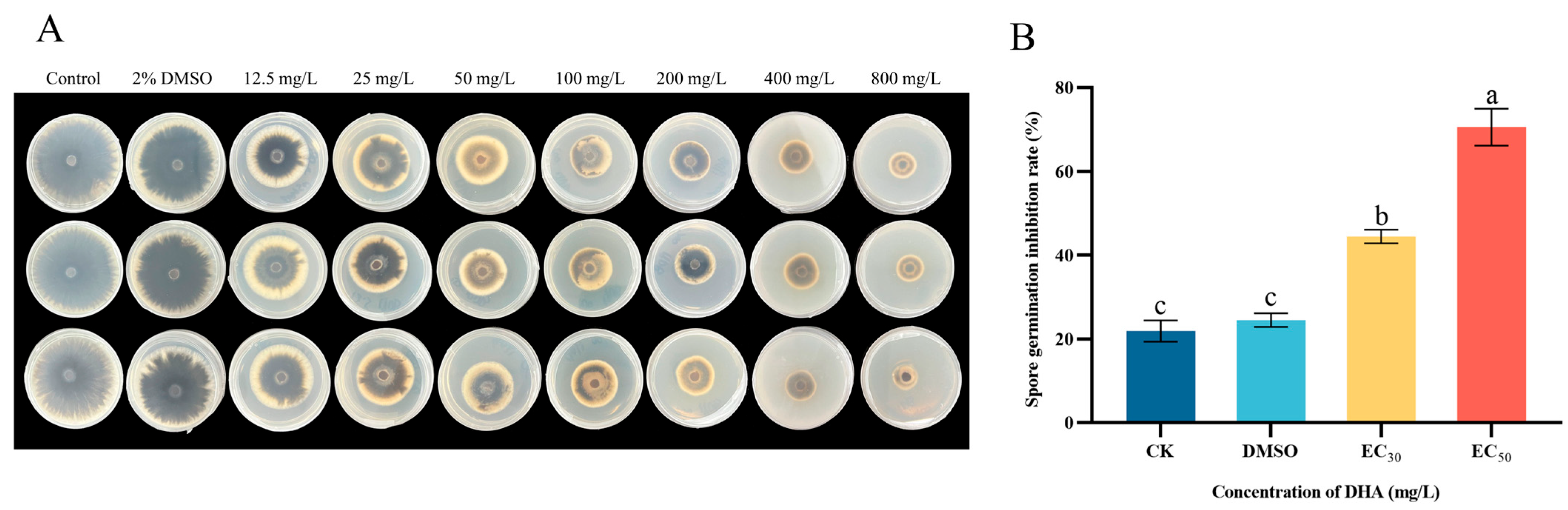
3.1. Effect of DHA on Mycelial Growth Inhibition of Five Pathogenic Fungi
3.2. Effect of DHA on Spore Germination of A. alternata
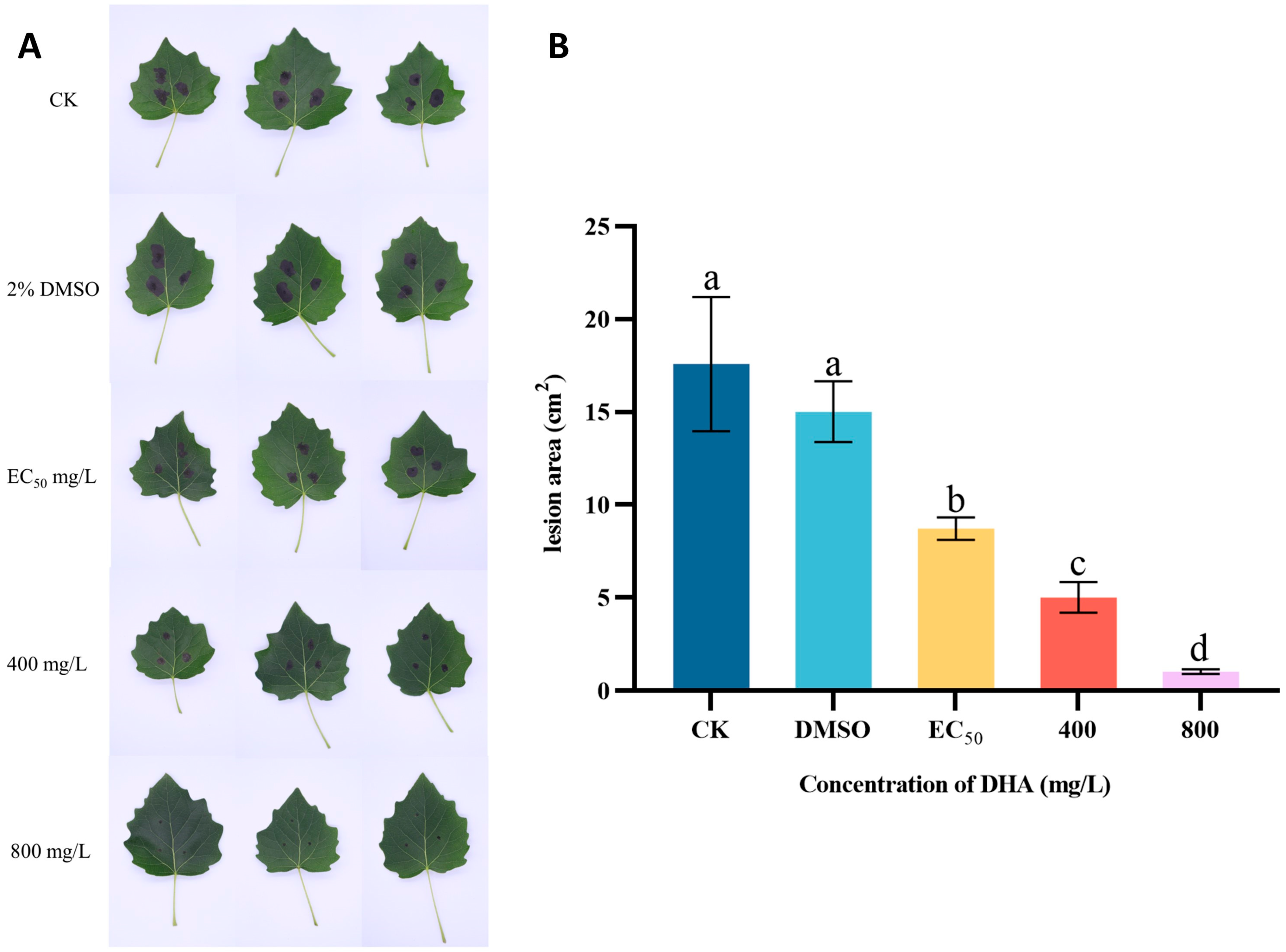
3.3. In Vivo Control Effect
3.4. Effect of DHA on Mycelial Morphology
3.5. Effect of DHA on Extracellular Proteins and Conductivity of A. alternata
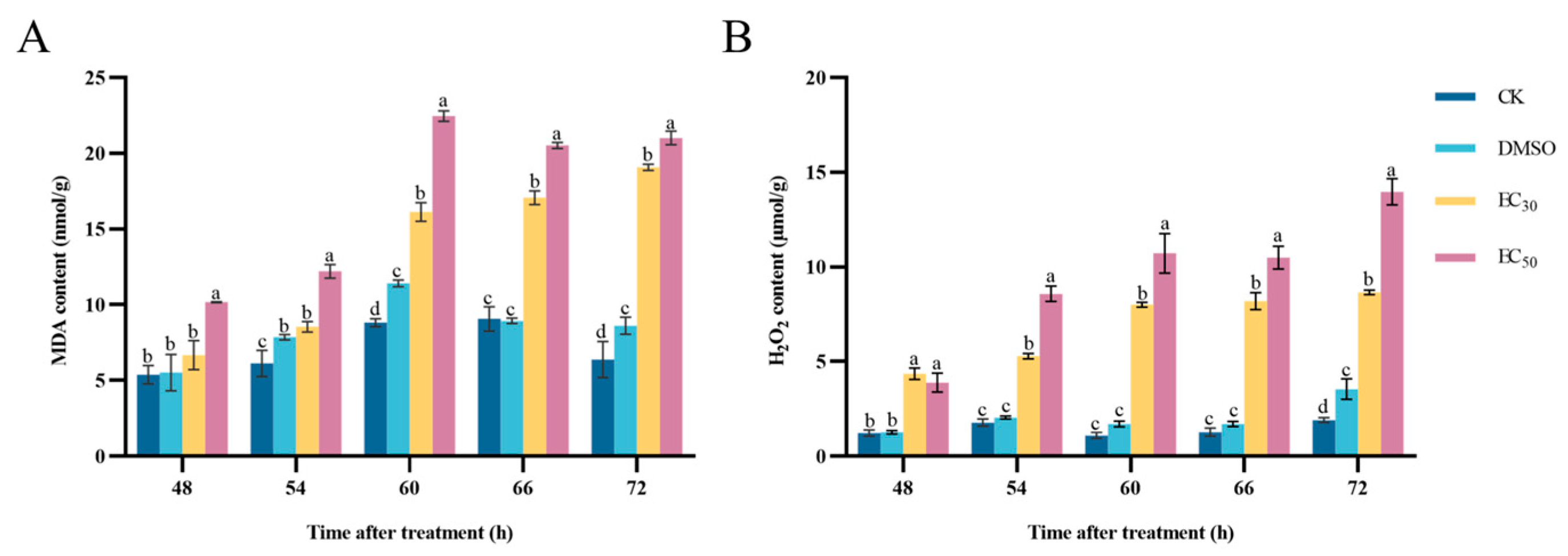
3.6. Effect of DHA on MDA and H2O2 Contents of A. alternata
3.7. Effect of DHA on Antioxidant Enzyme Activities of A. alternata
3.8. Effect of DHA on Cell-Wall-Degrading Enzymes Activities of A. alternata
3.9. Effect of DHA on the Gene Expression of Defense-Related Enzymes in A. alternata
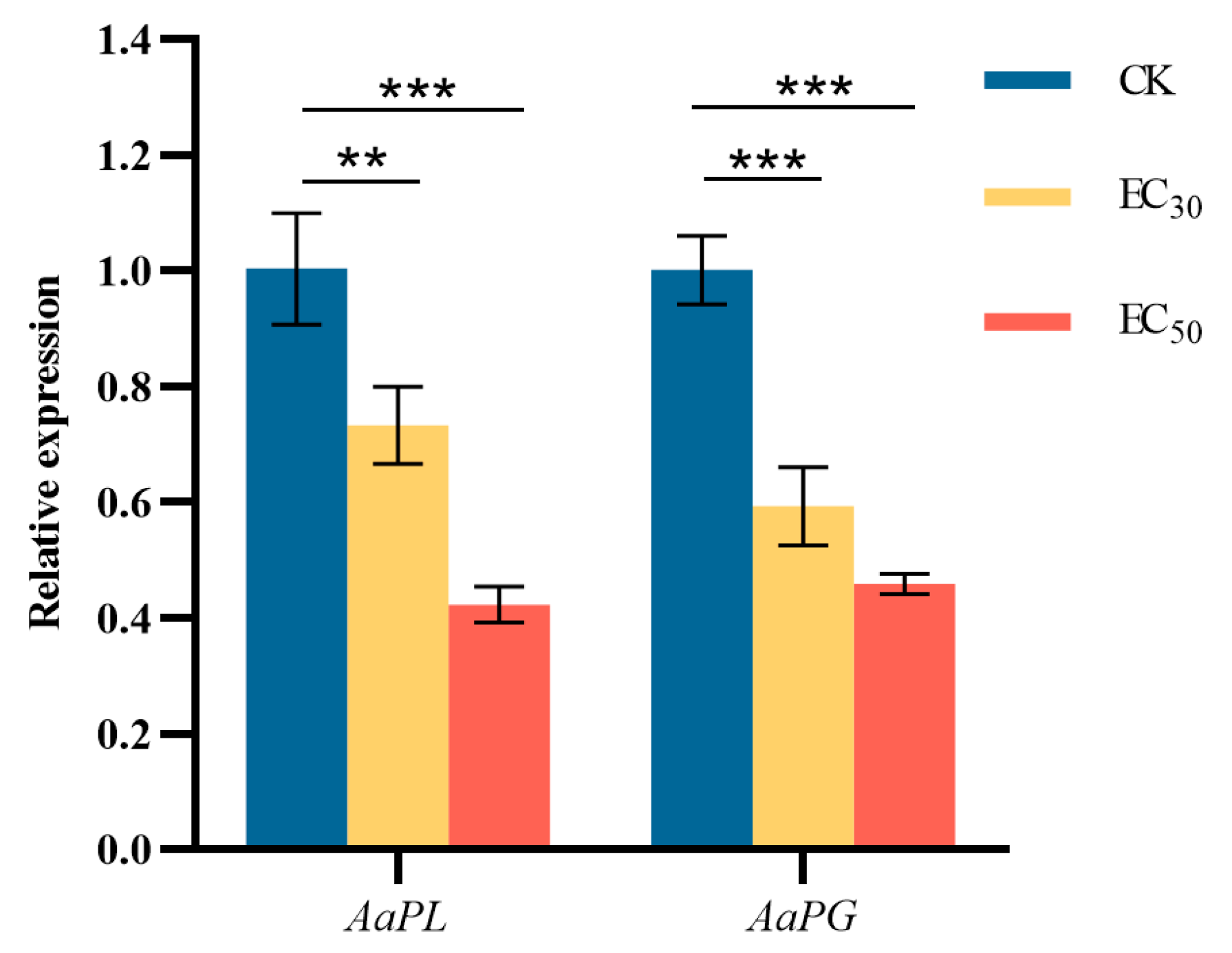
3.10. Effect of DHA on the Gene Expression of Pathogenicity-Related Enzyme in A. alternata
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DHA | Dehydroabietic acid |
| RT-qPCR | Real-time quantitative polymerase chain reaction |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| POD | Peroxidase |
| EG | Endoglucanase |
| PG | Polygalacturonase |
| PL | Pectin lyase |
| PDA | Potato dextrose agar |
| MDA | Malondialdehyde |
| H2O2 | Hydrogen peroxide |
| DMSO | Dimethyl sulfoxide |
| SEM | Scanning electron microscopy |
| RNA | Ribonucleic acid |
References
- Xi, B.; Clothier, B.; Coleman, M.; Duan, J.; Hu, W.; Li, D.; Di, N.; Liu, Y.; Fu, J.; Li, J.; et al. Irrigation management in poplar (Populus spp.) plantations: A review. For. Ecol. Manag. 2021, 494, 119330. [Google Scholar]
- Liu, W. Research progress on physiologic and ecologic characteristics of popular. World For. Res. 2010, 23, 50–55. [Google Scholar]
- Zeng, D. Review of current situation of research and control on poplar diseases in China. For. Pest Dis. 2002, 21, 20–26. [Google Scholar]
- Xiang, C.; Zhu, H. Control tactics of poplar diseases in China. J. For. Res. 2000, 11, 252. [Google Scholar]
- Demers, M. Alternaria alternata as endophyte and pathogen. Microbiology 2022, 168, 001153. [Google Scholar]
- Harish, B.N.; Nagesha, S.N.; Ramesh, B.N.; Shyamalamma, S.; Nagaraj, M.S.; Girish, H.C.; Pradeep, C.; Shiva Kumar, K.S.; Tharun Kumar, K.S.; Pavan, S.N.; et al. Molecular characterization and antifungal activity of lipopeptides produced from Bacillus subtilis against plant fungal pathogen Alternaria alternata. BMC Microbiol. 2023, 23, 179. [Google Scholar]
- Thomma, B. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar]
- Feng, L.; Zhao, D.; Jiao, L.; Song, L.; Peng, R. Polygenic identification of Poplar Leaf Blight pathogen and its pathogenicity to different poplar varieties. Mol. Plant Breed. 2022, 20, 6097–6103. [Google Scholar]
- Yang, L.N.; He, M.H.; Ouyang, H.B.; Zhu, W.; Pan, Z.C.; Sui, Q.J.; Shang, L.P.; Zhan, J. Cross-resistance of the pathogenic fungus Alternaria alternata to fungicides with different modes of action. BMC Microbiol. 2019, 19, 205. [Google Scholar]
- Leskovac, A.; Petrovic, S. Pesticide use and degradation strategies: Food safety, challenges and perspectives. Foods 2023, 12, 2709. [Google Scholar] [CrossRef]
- Huang, Y.; Ma, H.; Yue, Y.; Zhou, T.; Zhu, Z.; Wang, C. Integrated transcriptomic and transgenic analyses reveal potential mechanisms of poplar resistance to Alternaria alternata infection. BMC Plant Biol. 2022, 22, 413. [Google Scholar]
- Liu, P.; Liu, X.; Saburi, T.; Kubota, S.; Huang, P.; Wada, Y. Thermal stability evaluation of resin acids and rosin modified resins. Acs Omega 2020, 5, 29102–29109. [Google Scholar] [PubMed]
- Sarria-Villa, R.A.; Gallo-Corredor, J.A.; Benitez-Benitez, R. Characterization and determination of the quality of rosins and turpentines extracted from Pinus oocarpa and Pinus patula resin. Heliyon 2021, 7, e07834. [Google Scholar] [PubMed]
- Wang, X.; Pang, F.; Li, F. Review on the structural modification and biological activities of dehydroabietic acid. Nat. Prod. Res. Dev. 2017, 29, 1621–1625. [Google Scholar]
- Savluchinske-Feio, S.; Nunes, L.; Pereira, P.T.; Silva, A.M.; Roseiro, J.C.; Gigante, B.; Marcelo Curto, M.J. Activity of dehydroabietic acid derivatives against wood contaminant fungi. J. Microbiol. Methods 2007, 70, 465–470. [Google Scholar]
- Kamaya, Y.; Tokita, N.; Suzuki, K. Effects of dehydroabietic acid and abietic acid on survival, reproduction, and growth of the crustacean Daphnia magna. Ecotoxicol. Environ. Saf. 2005, 61, 83–88. [Google Scholar]
- Minami, T.; Wada, S.; Tokuda, H.; Tanabe, G.; Muraoka, O.; Tanaka, R. Potential antitumor-promoting diterpenes from the cones of Pinus luchuensis. J. Nat. Prod. 2002, 65, 1921–1923. [Google Scholar]
- Kim, J.; Kang, Y.G.; Lee, J.Y.; Choi, D.H.; Cho, Y.U.; Shin, J.M.; Park, J.S.; Lee, J.H.; Kim, W.G.; Seo, D.B.; et al. The natural phytochemical dehydroabietic acid is an anti-aging reagent that mediates the direct activation of SIRT1. Mol. Cell. Endocrinol. 2015, 412, 216–225. [Google Scholar]
- Söderberg, T.A.; Gref, R.; Holm, S.; Elmros, T.; Hallmans, G. Antibacterial activity of rosin and resin acids in vitro. Scand. J. Plast. Reconstr. Surg. Hand Surg. 1990, 24, 199–205. [Google Scholar]
- Kusumoto, N.; Zhao, T.; Swedjemark, G.; Ashitani, T.; Takahashi, K.; Borg-Karlson, A.-K. Antifungal properties of terpenoids in Picea abies against Heterobasidion parviporum. For. Pathol. 2014, 44, 353–361. [Google Scholar]
- Fallarero, A.; Skogman, M.; Kujala, J.; Rajaratnam, M.; Moreira, V.M.; Yli-Kauhaluoma, J.; Vuorela, P. (+)-Dehydroabietic acid, an abietane-type diterpene, inhibits Staphylococcus aureus biofilms in vitro. Int. J. Mol. Sci. 2013, 14, 12054–12072. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Ma, L.; Mo, X.; Lei, L. Advances in structural modification of dehydroabietic phenanthrene ring and activity of derivatives. Yunnan Chem. Technol. 2022, 49, 1–5. (In Chinese) [Google Scholar]
- Gao, Y.; Xu, R.; Gu, S.; Chen, K.; Li, J.; He, X.; Shang, S.; Song, Z.; Song, J. Discovery of natural rosin derivatives containing oxime ester moieties as potential antifungal agents to control tomato gray mold caused by Botrytis cinerea. J. Agric. Food Chem. 2022, 70, 5551–5560. [Google Scholar] [PubMed]
- Li, W.; Yuan, S.; Li, Q.; Sang, W.; Cao, J.; Jiang, W. Methyl p-coumarate inhibits black spot rot on jujube fruit through membrane damage and oxidative stress against Alternaria alternata. Postharvest Biol. Technol. 2018, 145, 230–238. [Google Scholar]
- Li, R.Y.; Wu, X.M.; Yin, X.H.; Long, Y.H.; Li, M. Naturally produced citral can significantly inhibit normal physiology and induce cytotoxicity on Magnaporthe grisea. Pestic. Biochem. Physiol. 2015, 118, 19–25. [Google Scholar]
- Qiu, M.; Wu, C.; Ren, G.; Liang, X.; Wang, X.; Huang, J. Effect of chitosan and its derivatives as antifungal and preservative agents on postharvest green asparagus. Food Chem. 2014, 155, 105–111. [Google Scholar]
- Liu, B.; Han, J.; Zhang, H.; Li, Y.; An, Y.; Ji, S.; Liu, Z. The regulatory pathway of transcription factor MYB36 from Trichoderma asperellum Tas653 resistant to poplar leaf blight pathogen Alternaria alternata Aal004. Microbiol. Res. 2024, 282, 127637. [Google Scholar] [CrossRef]
- Li, Y.; Shao, X.; Xu, J.; Wei, Y.; Xu, F.; Wang, H. Tea tree oil exhibits antifungal activity against Botrytis cinerea by affecting mitochondria. Food Chem. 2017, 234, 62–67. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shukla, A.; Attri, K.; Kumar, M.; Kumar, P.; Suttee, A.; Singh, G.; Barnwal, R.P.; Singla, N. Global trends in pesticides: A looming threat and viable alternatives. Ecotoxicol. Environ. Saf. 2020, 201, 110812. [Google Scholar] [PubMed]
- Yoon, M.; Cha, B.; Kim, J. Recent trends in studies on botanical fungicides in agriculture. Plant Pathol. J. 2013, 29, 1–9. [Google Scholar] [PubMed]
- Gonzalez, M.A. Synthetic derivatives of aromatic abietane diterpenoids and their biological activities. Eur. J. Med. Chem. 2014, 87, 834–842. [Google Scholar] [CrossRef]
- Zhai, Z.; Lv, P.; Zhao, P.; Song, Z.; Shang, S.; Rao, X. Antibacterial activity of dehydroabietyl nitrogen derivatives. Biomass Chem. Eng. 2020, 54, 1–7. (In Chinese) [Google Scholar]
- Chen, N.; Duan, W.; Lin, G.; Liu, L.; Zhang, R.; Li, D. Synthesis and antifungal activity of dehydroabietic acid-based 1,3,4-thiadiazole-thiazolidinone compounds. Mol. Divers. 2016, 20, 897–905. [Google Scholar]
- Peng, Y.; Chen, B. Role of cell membrane homeostasis in the pathogenicity of pathogenic filamentous fungi. Virulence 2024, 15, 2299183. [Google Scholar]
- Yuan, T.; Hua, Y.; Zhang, D.; Yang, C.; Lai, Y.; Li, M.; Ding, S.; Li, S.; Chen, Y. Efficacy and antifungal mechanism of rosemary essential oil against Colletotrichum gloeosporioides. Forests 2024, 15, 377. [Google Scholar] [CrossRef]
- Xue, D.; Zou, Z.; Chen, B.; Wang, Y.; Wu, H.; Ye, X.; Li, X. Study on membrane injury mechanism of total alkaloids and berberine from Coptidis rhizoma on Aeromonas hydrophila. China J. Chin. Mater. Medica 2015, 40, 1787–1792. [Google Scholar]
- Zhou, H.; Tao, N.; Jia, L. Antifungal activity of citral, octanal and α-terpineol against Geotrichum citri-aurantii. Food Control 2014, 37, 277–283. [Google Scholar]
- Maness, P.C.; Smolinski, S.; Blake, D.M.; Huang, Z.; Wolfrum, E.J.; Jacoby, W.A. Bactericidal activity of photocatalytic TiO2 reaction: Toward an understanding of its killing mechanism. Appl. Environ. Microbiol. 1999, 65, 4094–4098. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Yang, J.; Zhang, G.C.; Zhang, X.; Wang, T.; Wu, D. Evaluation of the antifungal activity of Lysurus mokusin extract against Pestalotiopsis neglecta and GC-MS analysis of the active components. J. Plant Pathol. 2021, 103, 1295–1305. [Google Scholar] [CrossRef]
- Martinez-Finley, E.J.; Gavin, C.E.; Aschner, M.; Gunter, T.E. Manganese neurotoxicity and the role of reactive oxygen species. Free. Radic. Biol. Med. 2013, 62, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Serrano, M.; L’Haridon, F.; Tjamos, S.E.; Metraux, J.P. Reactive oxygen species and plant resistance to fungal pathogens. Phytochemistry 2015, 112, 54–62. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef]
- Na, M.K.; Jang, T.S.; Choi, J.Y.; Lee, S.H.; Bae, K.H. Protective Effect of Some Medicinal Plants on tert-Butyl Hydroperoxide-Induced Oxidative Stress in Human Keratinocytes. Nat. Prod. Sci. 2008, 14, 244–248. [Google Scholar]
- Taheri, P.; Kakooee, T. Reactive oxygen species accumulation and homeostasis are involved in plant immunity to an opportunistic fungal pathogen. J. Plant Physiol. 2017, 216, 152–163. [Google Scholar] [CrossRef]
- Al-Issawi, M.; Rihan, H.Z.; Al-Shmgani, H.; Fuller, M.P. Molybdenum application enhances antioxidant enzyme activity and COR15a protein expression under cold stress in wheat. J. Plant Interact. 2016, 11, 5–10. [Google Scholar] [CrossRef]
- Bao, H.; Li, K.; Zhong, M.; Hao, Z. Inhibitory effect of piperine on Botrytis cinerea and its mechanism. J. China Agric. Univ. 2022, 27, 117–124. [Google Scholar]
- Pan, J.; Hao, X.; Yao, H.; Ge, K.; Ma, L.; Ma, W. Matrine inhibits mycelia growth of Botryosphaeria dothidea by affecting membrane permeability. J. For. Res. 2019, 30, 1105–1113. [Google Scholar] [CrossRef]
- Wan, J.; He, M.; Hou, Q.; Zou, L.; Yang, Y.; Wei, Y.; Chen, X. Cell wall associated immunity in plants. Stress Biol. 2021, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Yamamoto, R.; Sakurai, N.; Nakano, Y.; Takeda, T. Fungal hemicellulose-degrading enzymes cause physical property changes concomitant with solubilization of cell wall polysaccharides. Planta 2015, 241, 359–370. [Google Scholar] [PubMed]
- Wang, D.; Kanyuka, K.; Papp-Rupar, M. Pectin: A critical component in cell-wall-mediated immunity. Trends Plant Sci. 2023, 28, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Noutoshi, Y. The function of the plant cell wall in plant-microbe interactions. Plant Physiol. Biochem. 2022, 192, 273–284. [Google Scholar] [CrossRef]
- Yang, M.; Yang, Y.; Zheng, L.; Li, M.; Zhou, E. Component analysis, activity measurement and pathogenic effect of cell wall degrading enzymes from rice sheath blight pathogen Rhizoctonia solani. Chin. J. Rice Sci. 2012, 26, 600–606. [Google Scholar]
- Kim, B.; Nguyen, M.V.; Park, J.; Kim, Y.S.; Han, J.W.; Lee, J.-Y.; Jeon, J.; Son, H.; Choi, G.J.; Kim, H. The antifungal activity and mechanism of Brevibacillus brevis on plant pathogenic fungus. J. Plant Prot. 2016, 43, 600–607. [Google Scholar]
- Li, N.; Wu, Y.X.; Zhang, Y.D.; Wang, S.R.; Zhang, G.C.; Yang, J. Phytic acid is a new substitutable plant-derived antifungal agent for the seedling blight of Pinus sylvestris var. mongolica caused by Fusarium oxysporum. Pestic. Biochem. Physiol. 2023, 191, 105341. [Google Scholar]
- Zhang, J.; Zhu, W.; Goodwin, P.H.; Lin, Q.; Xia, M.; Xu, W.; Sun, R.; Liang, J.; Wu, C.; Li, H.; et al. Response of Fusarium pseudograminearum to biocontrol agent Bacillus velezensis YB-185 by phenotypic and transcriptome analysis. J. Fungi 2022, 8, 763. [Google Scholar] [CrossRef]
- Zhang, Z.; Bai, Y.; Liu, Y.; Tang, M.; Saibihai, Y.; Feng, Z. Gene expression and activity change analysis of cell wall degrading enzyme in postharvest melon infected by Alternaria alternata. Sci. Technol. Food Ind. 2024, 45, 291–299. [Google Scholar]
- Yang, J.; Ji, J.Y.; Zhang, B.W.; Chen, Y.Z.; Wang, S.R.; Zhang, G.C.; Zhang, J. Transcriptome and cell wall degrading enzyme-related gene analysis of Pestalotiopsis neglecta in response to sodium pheophorbide a. Pestic. Biochem. Physiol. 2020, 169, 104639. [Google Scholar] [CrossRef]
- Paccanaro, M.C.; Sella, L.; Castiglioni, C.; Giacomello, F.; Martínez-Rocha, A.L.; D’Ovidio, R.; Schäfer, W.; Favaron, F. Synergistic effect of different plant cell wall-degrading enzymes is important for virulence of Fusarium graminearum. Mol. Plant Microbe Interact. 2017, 30, 886–895. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Gene ID | Encoded Protein | Forward Primer (5′−3′) | Reverse Primer (5′−3′) |
|---|---|---|---|---|
| AaSOD | 29119212 | Cu/Zn superoxide dismutase | TCCTCGCAACCGCAAAGTCT | CTTGAGCCCTCCCGTCTCAC |
| AaPOD | 29116333 | Peroxidase | AGGAAGGACCAGACAAGCAC | ATCTTCCTGCTGCTGGTCTG |
| AaCAT | 29114347 | Catalase | ATGGTGCGAGTTCAGATCCT | CGTCAGTCGACGCAGCATTG |
| AaPL | 29114697 | Pectin lyase | GGATGTTTGCATGAACTACT | CTATCTGCAGACGGGAAGGG |
| AaPG | 29119730 | Polygalacturonase | ATGGTTGCCTTTGCACTCAC | TGCAAGAAGCCTTGCCCTTG |
| AaBenA | 29108751 | β-tubulin | GTTGAGAACTCAGACGAGACCTTCTGCATTG | CTCCTTCAACCGGCAGTTGTACCAAG |
| Phytopathogenic Fungi | Toxicity Equation | X2 | R2 | EC30 (mg/L) | 95% Confidence Interval (mg/L) | EC50 (mg/L) | 95% Confidence Interval (mg/L) |
|---|---|---|---|---|---|---|---|
| A. alternata | y = 0.7x − 1.23 | 1.903 | 0.991 | 9.966 | 6.672–13.691 | 56.015 | 45.542–67.604 |
| P. neglecta | y = 0.94x − 2.41 | 4.227 | 0.988 | 101.918 | 87.669–117.725 | 492.687 | 306.352–441.426 |
| F. oxysporum | y = 0.69x − 1.92 | 13.724 | 0.935 | 106.78 | 87.701–129.194 | 595.547 | 456.929–827.609 |
| B. cinerea | y = 0.64x − 1.18 | 4.062 | 0.979 | 10.727 | 6.969–15.009 | 70.664 | 57.146–86.211 |
| V. mali | y = 0.76x − 1.42 | 20.814 | 0.924 | 14.763 | 10.686–19.21 | 72.319 | 60.488–85.681 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-Z.; Zhang, Y.-D.; Chen, C.; Sa, Q.-E.; Yang, J.; Zhang, G.-C. The Antifungal Activity and Mechanism of Dehydroabietic Acid Against Alternaria alternata Causing Poplar Leaf Spot. J. Fungi 2025, 11, 265. https://doi.org/10.3390/jof11040265
Chen Y-Z, Zhang Y-D, Chen C, Sa Q-E, Yang J, Zhang G-C. The Antifungal Activity and Mechanism of Dehydroabietic Acid Against Alternaria alternata Causing Poplar Leaf Spot. Journal of Fungi. 2025; 11(4):265. https://doi.org/10.3390/jof11040265
Chicago/Turabian StyleChen, Yun-Ze, Yun-Di Zhang, Cheng Chen, Qiu-Er Sa, Jing Yang, and Guo-Cai Zhang. 2025. "The Antifungal Activity and Mechanism of Dehydroabietic Acid Against Alternaria alternata Causing Poplar Leaf Spot" Journal of Fungi 11, no. 4: 265. https://doi.org/10.3390/jof11040265
APA StyleChen, Y.-Z., Zhang, Y.-D., Chen, C., Sa, Q.-E., Yang, J., & Zhang, G.-C. (2025). The Antifungal Activity and Mechanism of Dehydroabietic Acid Against Alternaria alternata Causing Poplar Leaf Spot. Journal of Fungi, 11(4), 265. https://doi.org/10.3390/jof11040265