
Forecasting of Airborne Conidia Quantities and Potential Insect Associations of Cryphonectria parasitica, the Causal Agent of Chestnut Blight, in England

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Spore and Insect Traps
2.2. DNA Extraction from Sticky Rods
2.3. Insects Processing
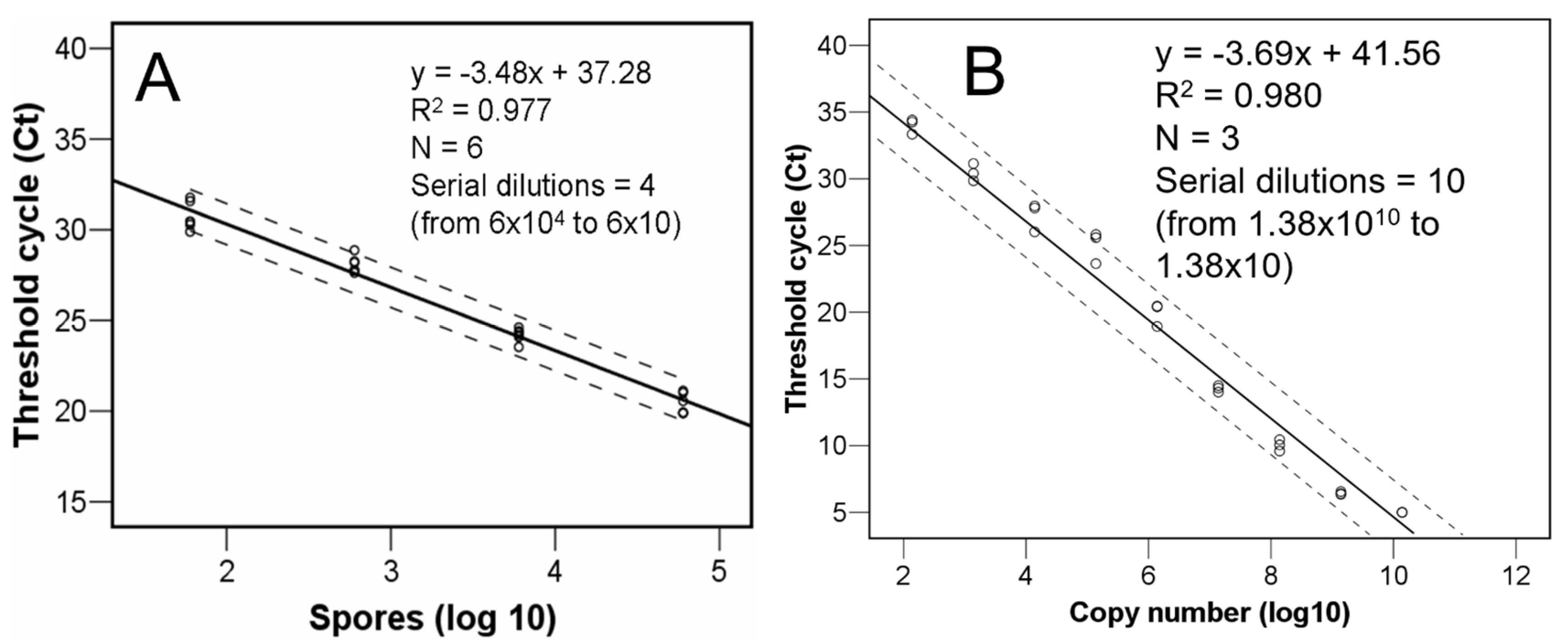
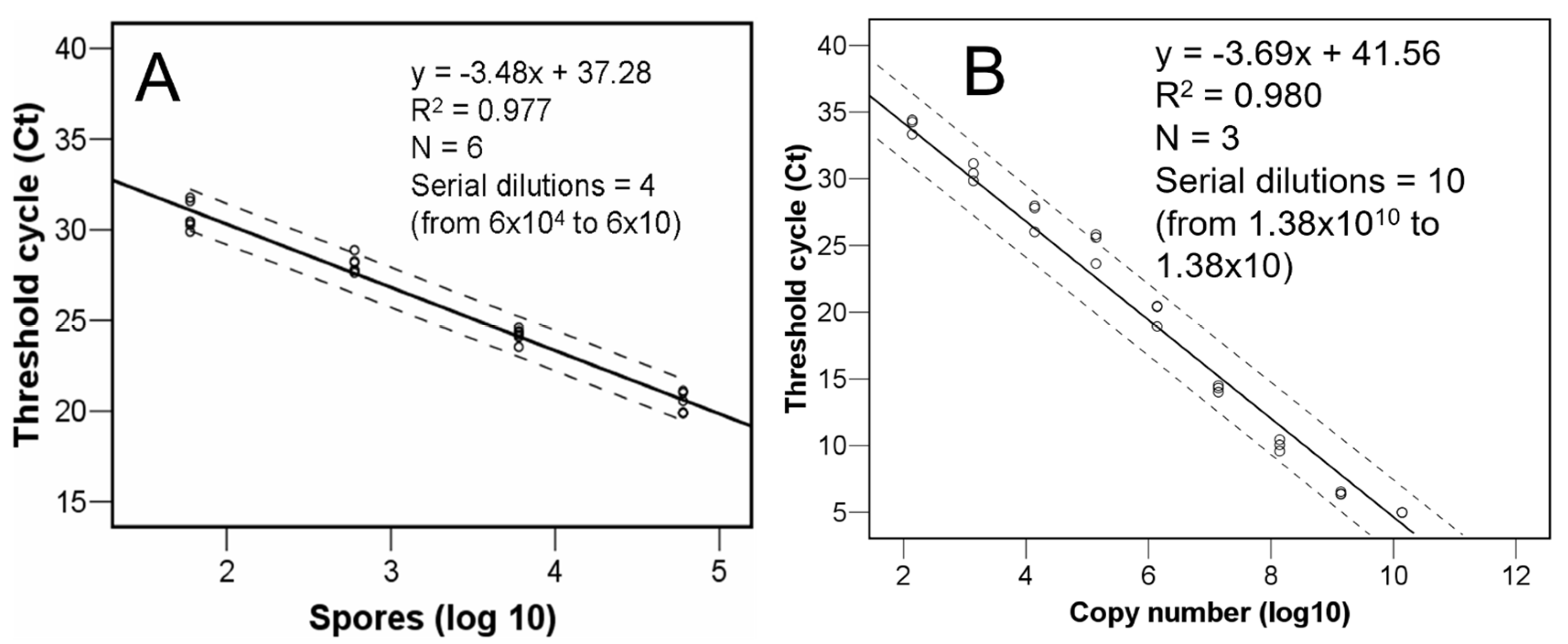
2.4. Real-Time PCR
2.5. Other Fungi Identification
2.6. Other Insects’ Identification
3. Results
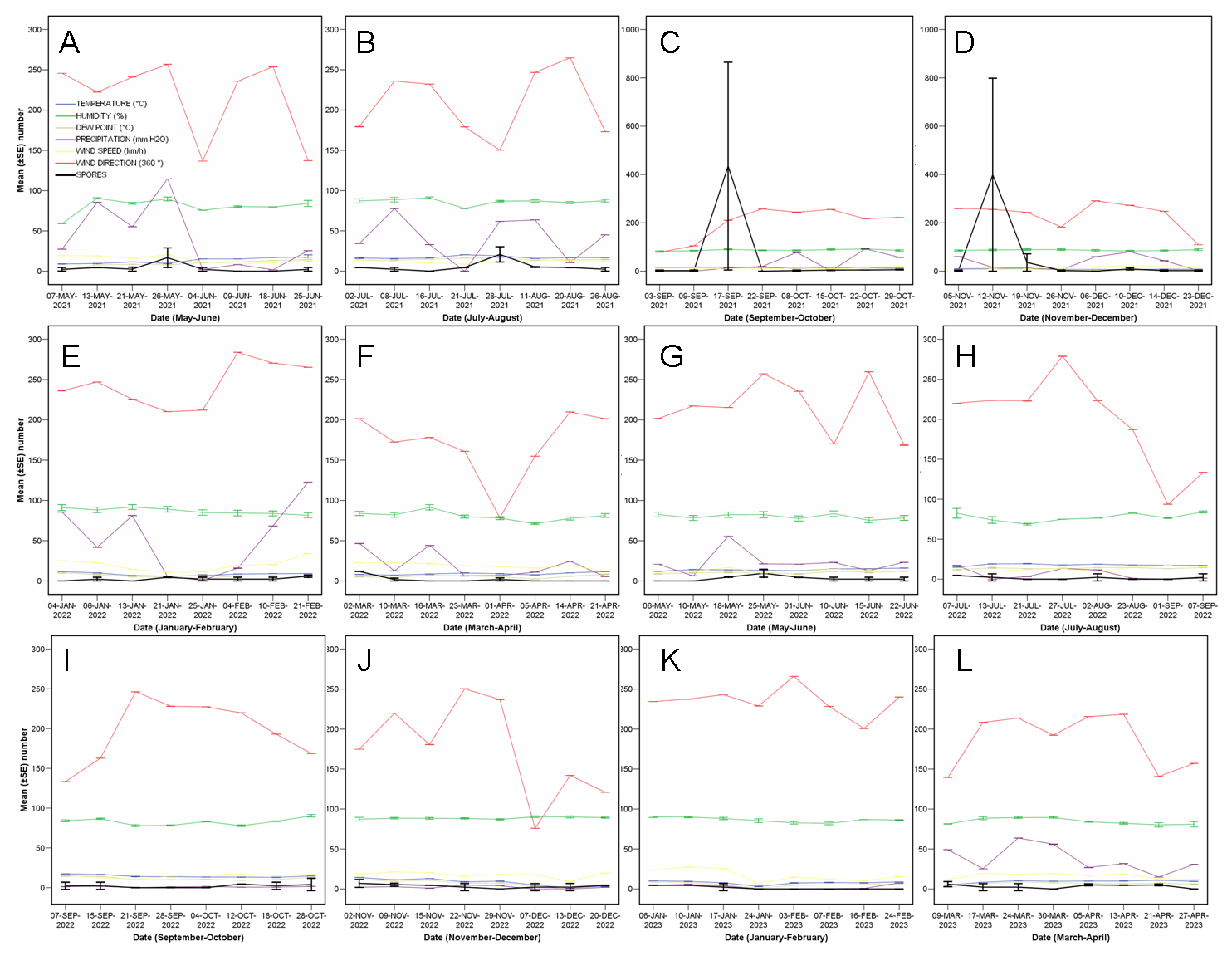
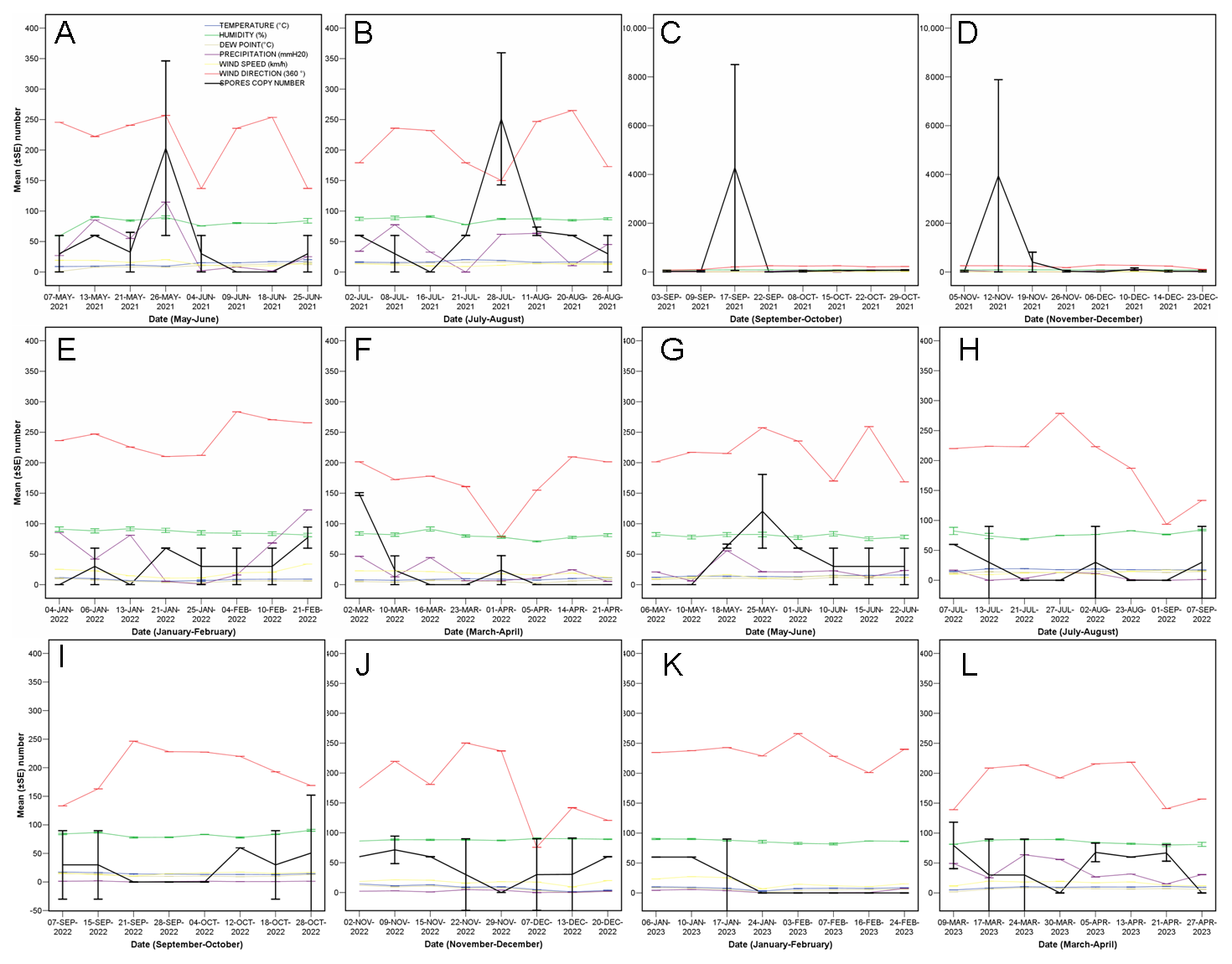
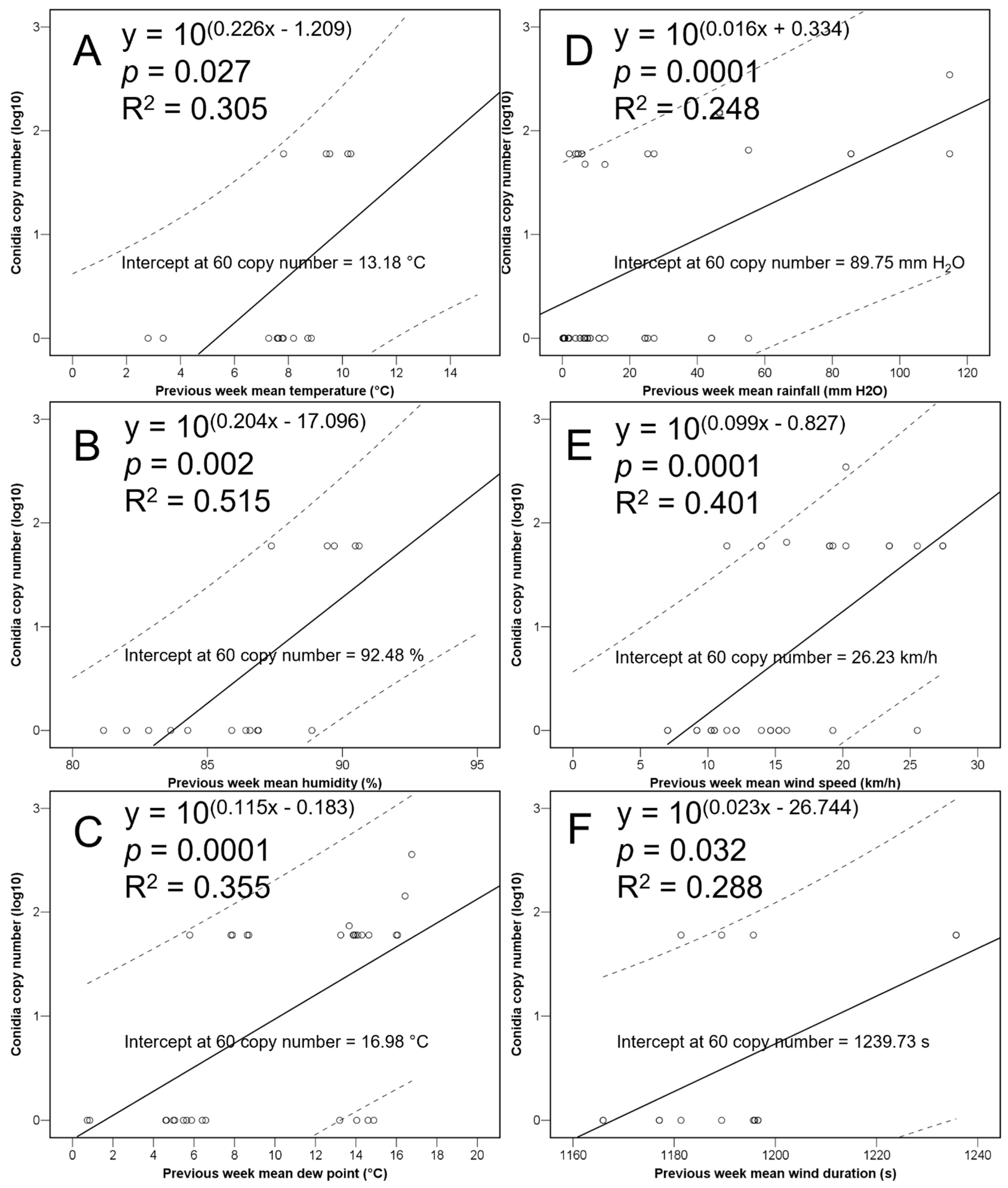
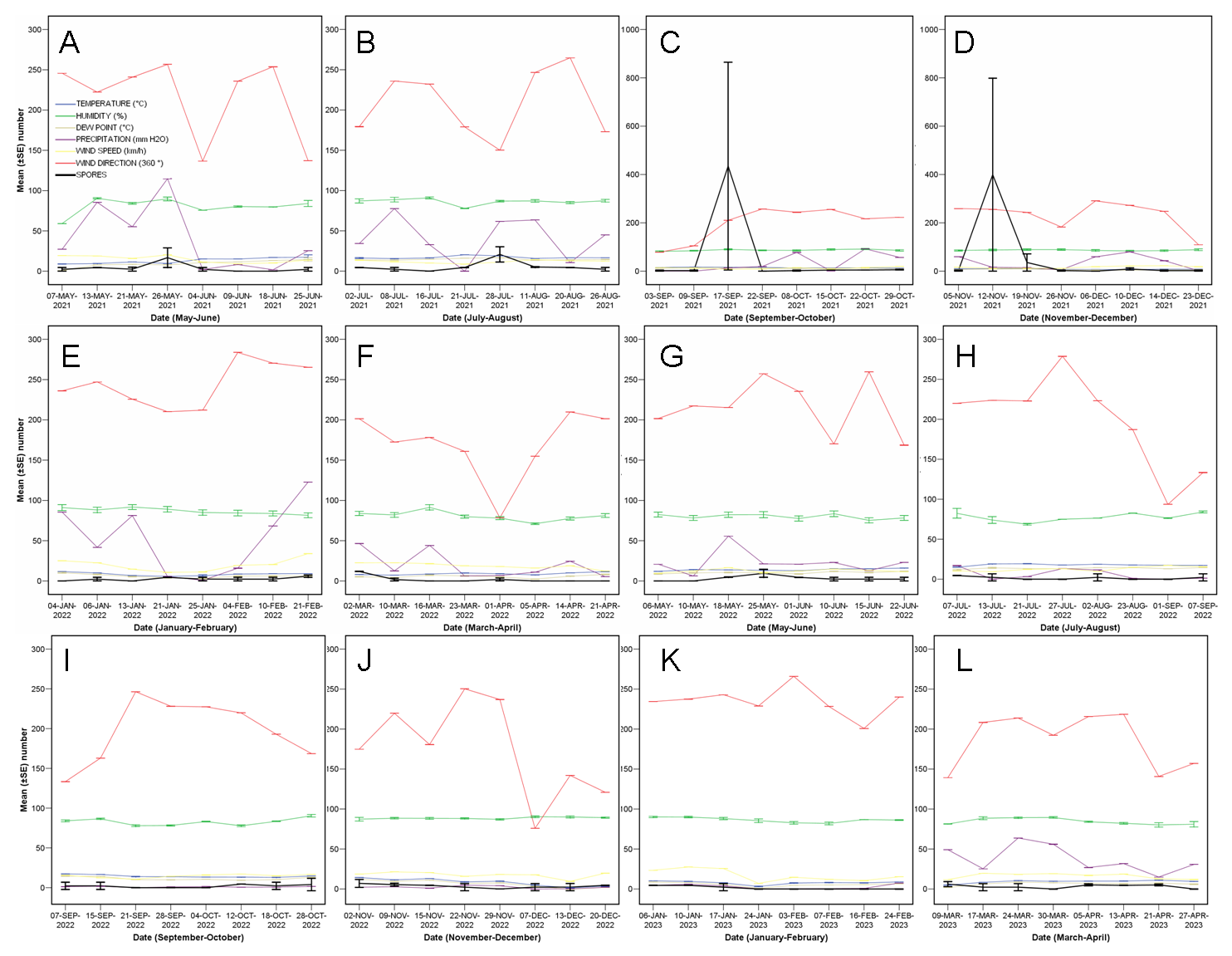
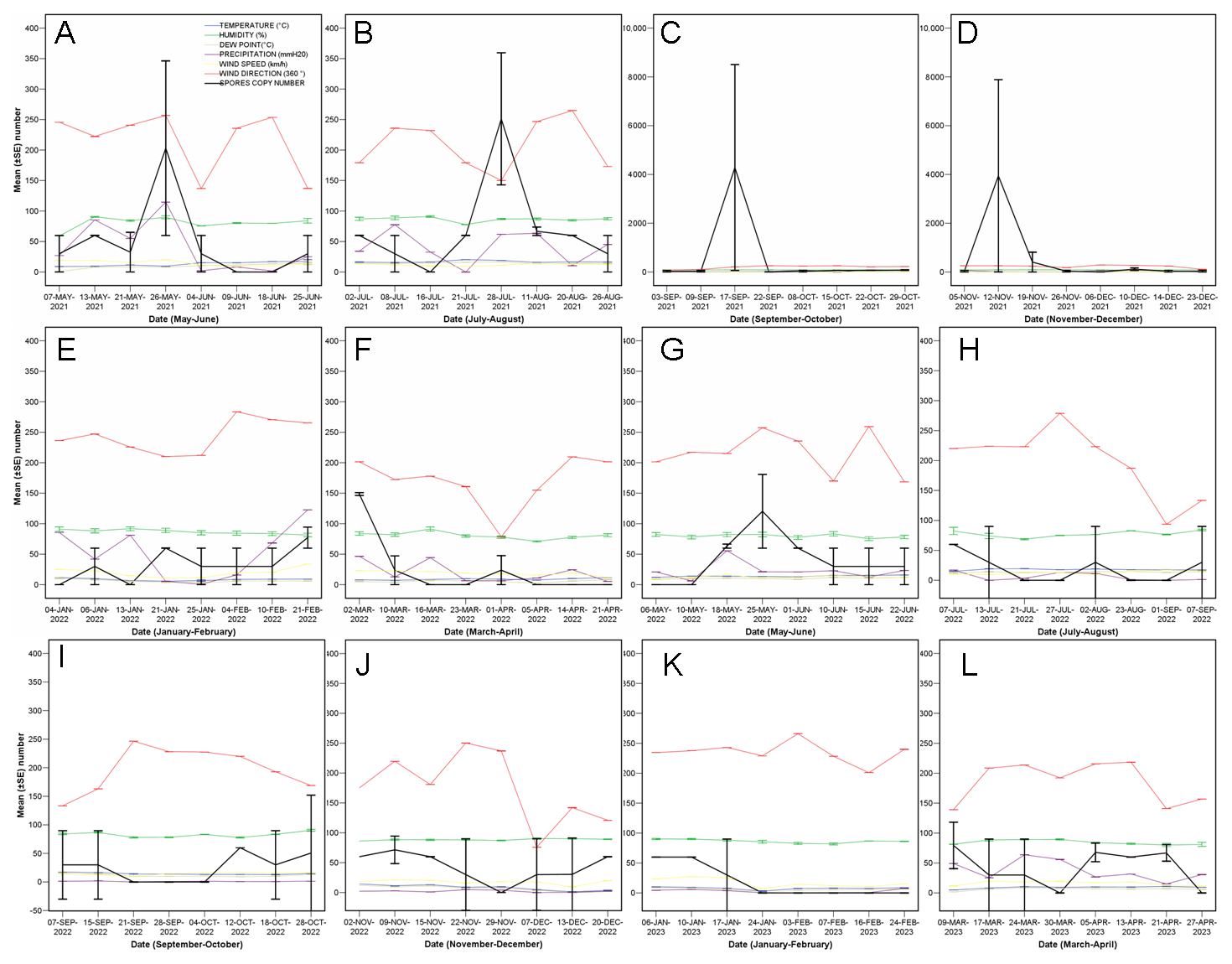
3.1. Quantity of C. parasitica Spores
3.2. Detection of C. parasitica from Insects
3.3. Other Fungi
3.4. Other Insects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.-C.; Dynek, J.N.; Hillman, B.I.; Milgroom, M.G. Diversity of viruses in Cryphonectria parasitica and C. nitschkei in Japan and China, and partial characterization of a new chrysovirus species. Mycol. Res. 2007, 111, 433–442. [Google Scholar] [CrossRef]
- Roane, M.K.; Griffin, G.J.; Elkins, J.R. Chestnut Blight, Other Endothia Diseases, and the Genus Endothia; APS Monograph Series 53; APS Press: St. Paul, MN, USA, 1986. [Google Scholar]
- Anagnostakis, S.L. Chestnut blight: The classical problem of an introduced pathogen. Mycologia 1987, 79, 23–37. [Google Scholar] [CrossRef]
- Robin, C.; Heiniger, U. Chestnut blight in Europe: Diversity of Cryphonectria parasitica, hypovirulence and biocontrol. For. Snow Land. Res. 2001, 76, 361–367. [Google Scholar]
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the causal agent of chestnut blight: Invasion history, population biology and disease control. Mol. Plant Pathol. 2018, 19, 7–20. [Google Scholar] [CrossRef]
- Rodríguez-Molina, M.C.; García-García, M.B.; Osuna, M.D.; Gouveia, E.; Serrano-Pérez, P. Various population structures of Cryphonectria parasitica in Cáceres (Spain) determine the feasibility of the biological control of chestnut blight with hypovirulent strains. Agronomy 2023, 13, 1208–1221. [Google Scholar] [CrossRef]
- Hunter, G.; Wylder, B.; Jones, B.; Webber, J.F. First finding of Cryphonectria parasitica causing chestnut blight on Castanea sativa trees in England. New Dis. Rep. 2013, 27, 1. [Google Scholar] [CrossRef]
- Forestry Commission. Sweet Chestnut Blight (Cryphonectria parasitica). 2018. Available online: https://www.forestry.gov.uk/chestnutblight (accessed on 9 January 2024).
- Pérez-Sierra, A.; Romon-Ochoa, P.; Gorton, C.; Lewis, A.; Rees, H.; van der Linde, S.; Webber, J. High vegetative compatibility diversity of Cryphonectria parasitica infecting sweet chestnut (Castanea sativa) in Britain indicates multiple pathogen introductions. Plant Pathol. 2019, 68, 727–737. [Google Scholar] [CrossRef]
- Romon-Ochoa, P.; Kranjec Orlovic, J.; Gorton, C.; Lewis, A.; van der Linde, S.; Pérez-Sierra, A. New detections of chestnut blight in Britain during 2019–2020 reveal high Cryphonectria parasitica diversity and limited spread of the disease. Plant Pathol. 2021, 71, 793–804. [Google Scholar] [CrossRef]
- Romon-Ochoa, P.; Samal, P.; Gorton, C.; Lewis, A.; Chitty, R.; Eacock, A.; Krzywinska, E.; Crampton, M.; Pérez-Sierra, A.; Biddle, M.; et al. Cryphonectria parasitica detections in England, Jersey, and Guernsey during 2020–2023 reveal newly affected areas and infections by the CHV1 mycovirus. J. Fungi 2023, 9, 1036–1050. [Google Scholar] [CrossRef]
- Romon-Ochoa, P.; Forster, J.; Chitty, R.; Gorton, C.; Lewis, A.; Eacock, A.; Kupper, Q.; Rigling, D.; Pérez-Sierra, A. Canker development and biocontrol potential of CHV1 infected English isolates of Cryphonectria parasitica is dependent on the virus concentration and the compatibility of the fungal inoculums. Viruses 2022, 14, 2678–2694. [Google Scholar] [CrossRef] [PubMed]
- Romon-Ochoa, P.; Lewis, A.; Gorton, C.; van der Linde, S.; Pérez-Sierra, A. Effects of growth medium, temperature and mycelium age on CHV-1 accumulation and transmission. For. Ecol. Manag. 2023, 529, 120705–120717. [Google Scholar] [CrossRef]
- Romon-Ochoa, P.; Smith, O.; Lewis, A.; Kupper, Q.; Shamsi, W.; Rigling, D.; Pérez-Sierra, A.; Ward, L. Temperature effects on the Cryphonectria hypovirus 1 accumulation and recovery within its fungal host, the chestnut blight pathogen Cryphonectria parasitica. Viruses 2023, 15, 1260–1273. [Google Scholar] [CrossRef]
- Lione, G.; Brescia, F.; Giordano, L.; Gonthier, P. Effects of seasonality and climate on the propagule deposition patterns of the chestnut blight pathogen Cryphonectria parasitica in orchards of the Alpine district of North Western Italy. Agriculture 2022, 12, 644–668. [Google Scholar] [CrossRef]
- Romon-Ochoa, P.; Gorton, C.; Lewis, A.; van der Linde, S.; Webber, J.; Pérez-Sierra, A. Hypovirulent effect of the Cryphonectria Hypovirus 1 in British isolates of Cryphonectria parasitica. Pest Manag. Sci. 2020, 76, 1333–1343. [Google Scholar] [CrossRef]
- Morales-Rodriguez, C.; Pérez-Sierra, A.; Romon-Ochoa, P.; Inward, D.; Fabro, M.; Vannini, A. The fungal community associated with individuals of the ambrosia beetle Xylosandrus germanus captured in chestnut groves. In VII International Chestnut Symposium; International Society of Horticultural Science ISHS Press: Lugo, Spain, 2023. [Google Scholar]
- Pérez-Sierra, A.; van der Linde, S.; Romon-Ochoa, P.; Jones, B.; Gorton, C. First report of Cryphonectria parasitica on abandoned galls of Dryocosmus kuriphilus on sweet chestnut in the United Kingdom. New Dis. Rep. 2020, 41, 34. [Google Scholar] [CrossRef]
- Dvorak, M.; Rotkova, G.; Botella, L. Detection of airborne inoculum of Hymenoscyphus fraxineus and H. albidus during seasonal fluctuations associated with absence of apothecia. Forests 2015, 7, 1–13. [Google Scholar] [CrossRef]
- Chandelier, A.; Massot, M.; Fabreguettes, O. Early detection of Cryphonectria parasitica by real-time PCR. Eur. J. Plant Pathol. 2019, 153, 135–152. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Application; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Viviani, A.; Bernardi, R.; Cavallini, A.; Rossi, E. Genotypic characterization of Torymus sinensis (Hymenoptera: Torymidae) after its introduction in Tuscany (Italy) for the biological control of Dryocosmus kuriphilus (Hymenoptera: Cynipidae). J. Insect Sci. 2019, 19, 17–25. [Google Scholar] [CrossRef]
- Lewis, A.; Gorton, C.; Rees, H.; Webber, J.; Pérez-Sierra, A. First report of Gnomoniopsis smithogilvyi causing lesions and cankers of sweet chestnut in the United Kingdom. New Dis. Rep. 2017, 35, 20. [Google Scholar] [CrossRef]
- Srutka, P.; Pazoutova, S.; Kolarik, M. Daldinia decipiens and Entonaema cinnabarina as fungal symbionts of Xiphydria wood wasps. Mycol. Res. 2007, 111, 224–231. [Google Scholar] [CrossRef]
- Smith, M.A.; Bertrand, C.; Crosby, K.; Eveleigh, E.S.; Fernandez-Triana, J.; Fisher, B.L.; Gibbs, J.; Hajibabaei, M.; Hallwachs, W.; Hind, K.; et al. Wolbachia and DNA Barcoding Insects: Patterns, Potential, and Problems. PLoS ONE 2012, 7, e36514. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, M.; Drapela, K.; Jankovsky, L. Dothistroma septosporum: Spore production and weather conditions. For. Syst. 2012, 2, 323–328. [Google Scholar] [CrossRef]
- Guerin, L.; Froidefond, G.; Xu, X.M. Seasonal patterns of dispersal of ascospores of Cryphonectria parasitica (chestnut blight). Plant Pathol. 2001, 50, 717–724. [Google Scholar] [CrossRef]
- Heald, F.D.; Gardner, M.W.; Studhalter, R.A. Air and wind dissemination of ascospores of chestnut-blight fungus. J. Agric. Res. 1915, 3, 493–526. [Google Scholar]
- Bryner, S.F.; Rigling, D. Temperature-dependent genotype-by-genotype interaction between a pathogenic fungus and its hyperparasitic virus. Am. Nat. 2011, 177, 65–74. [Google Scholar] [CrossRef]
- Guerin, L.; Robin, C. Seasonal effect on infection and development of lesions caused by Cryphonectria parasitica in Castanea sativa. For. Pathol. 2003, 33, 223–235. [Google Scholar] [CrossRef]
- Rankin, W.H. Field studies on Endothia canker of chestnut in New York State. Phytopathology 1914, 4, 233–261. [Google Scholar]
- Prospero, S.; Rigling, D. Chestnut blight. In Infectious Forest Diseases; Gonthier, P., Nicolotti, P., Eds.; CAB International: Wallingford, UK, 2013; pp. 318–339. [Google Scholar]
- Li, D.W.; Kendrick, B. A year-round study on functional relationships of airborne fungi with meteorological factors. Int. J. Biometeor. 1995, 39, 74–80. [Google Scholar] [CrossRef]
- Lione, G.; Giordano, L.; Sillo, F.; Brescia, F.; Gonthier, P. Temporal and spatial propagule deposition patterns of the emerging fungal pathogen of chestnut Gnomoniopsis castaneae in orchards of north-western Italy. Plant Pathol. 2021, 70, 2016–2033. [Google Scholar] [CrossRef]
- Lione, G.; Giordano, L.; Turina, M.; Gonthier, P. Hail-induced infections of the chestnut blight pathogen Cryphonectria parasitica depend on wound size and may lead to severe diebacks. Phytopathology 2020, 110, 1280–1293. [Google Scholar] [CrossRef]
- Griffin, G.J.; Wendt, R.A.; Elkins, J.R. Association of hypovirulent Endothia parasitica with American chestnut in forest clear-cuts and with mites. Phytopathology 1984, 74, 804. [Google Scholar]
- Griffin, G.J.; Eisenback, J.D.; Yancey, M.M.; Templeton, J. Aphelenchoides hylurgi as a carrier of white hypovirulent Cryphonectria parasitica and its possible role in hypovirulence spread on blight-controlled American chestnut trees. J. Nematol. 2009, 41, 267–273. [Google Scholar]
- Russin, J.S.; Shain, L.; Nordin, G.L. Insects as carriers of virulent and cytoplasmic hypovirulent isolates of the chestnut blight fungus. J. Econ. Entomol. 1984, 77, 838–846. [Google Scholar] [CrossRef]
- Diamandis, S.; Perlerou, C. The role of Spulerina simploniella in the spread of the chestnut blight. For. Pathol. 2005, 35, 315–320. [Google Scholar] [CrossRef]
- Anagnostakis, S. Carpenter ants as carriers of Endothia parasitica. In Proc. USDA American Chestnut Symposium Cooperative Meeting; Smith, H.C., McDonald, W.L., Morgantown, W.V., Eds.; US Department of Agriculture: Washington, DC, USA, 1982; pp. 111–113. [Google Scholar]
- Craighead, F.C. Insects in their relation to the chestnut bark disease. Science 1916, 43, 133–135. [Google Scholar] [CrossRef]
- Wendt, R.; Weidhaas, J.; Griffin, G.J.; Elkins, J.R. Association of Endothia parasitica with mites isolated from cankers on American chestnut trees. Plant Dis. 1983, 67, 737–738. [Google Scholar] [CrossRef]
- Nannelli, R.; Turchetti, T. Mites as carriers of hypovirulent strains of the chestnut blight fungus (Cryphonectria parasitica). Redia 1999, 82, 89–99. [Google Scholar]
- Pakaluk, J.; Anagnostakis, S.L. Acoptus suturalis Le Conte (Coleoptera: Curculionidae: Zygopinae) a potential vector of the chestnut blight fungus, Cryphonectria parasitica (Murrill) Barr, in the Eastern United States. Proc. Entomol. Soc. Wash. 1997, 99, 583–584. [Google Scholar]
- Nannelli, R.; Turchetti, T.; Maresi, G. Corticolous mites (acari) as potential vectors of Cryphonectria parasitica (Murr.) Barr hypovirulent strains. Int. J. Acarol. 1998, 24, 237–244. [Google Scholar] [CrossRef]
- Simoni, S.; Nannelli, R.; Robersi, P.F.; Turchetti, T.; Bouneb, M. Thyreophagus corticalis as a vector of hypovirulence of Cryphonectria parasitica in chestnut stands. Exp. Appl. Acarol. 2014, 62, 363–375. [Google Scholar] [CrossRef]
- Bouneb, M.; Turchetti, T.; Nannelli, R.; Robersi, P.F.; Paoli, F.; Danti, R.; Simoni, S. Occurrence and transmission of mycovirus Cryphonectria hypovirus 1 from dejecta of Thyreophagus corticalis (Acari, Acaridae). Fun. Biol. 2016, 120, 351–357. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
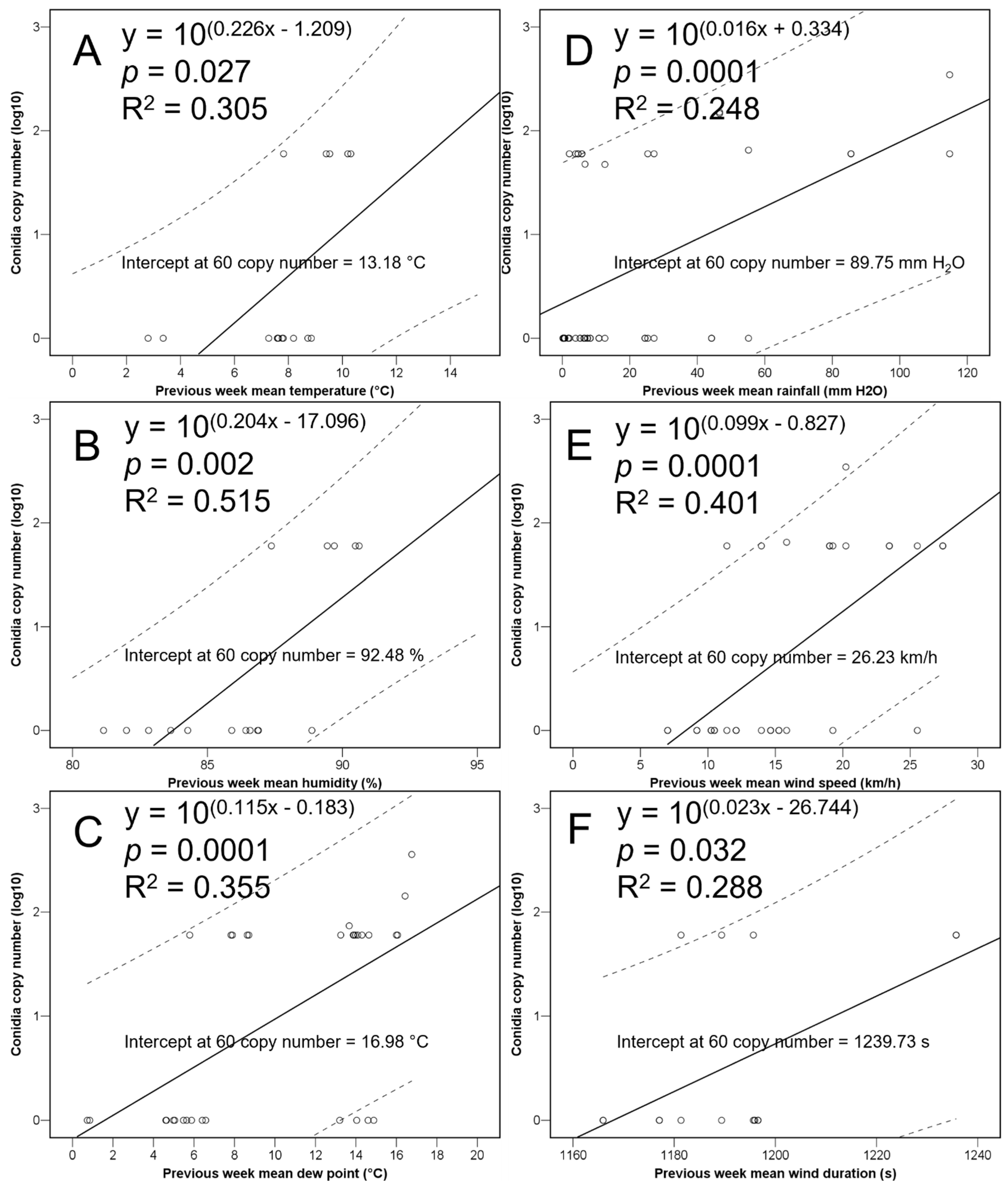{kind=link}
| Based on Spores Serial Dilutions | Based on Fragment Copy Number Dilutions | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Humidity (%) | Dew Point (°C) | Rain (mm H2O) | Wind Speed (km/h) | Wind Direction (360°) | Wind Duration (s) | Temperature (°C) | Humidity (%) | Dew Point (°C) | Rain (mm H2O) | Wind Speed (km/h) | Wind Direction (360°) | Wind Duration (s) | ||
| May–Jun 2021 | Pearson Correlation | −0.423 | 0.386 | −0.181 | 0.629 | 0.493 | 0.185 | 0.183 | −0.437 | 0.391 | −0.191 | 0.640 | 0.507 | 0.180 | 0.183 |
| Sig. (2-tailed) | 0.103 | 0.140 | 0.502 | 0.009 | 0.053 | 0.492 | 0.497 | 0.091 | 0.134 | 0.478 | 0.008 | 0.045 | 0.505 | 0.498 | |
| Jul–Aug 2021 | Pearson Correlation | 0.434 | 0.008 | 0.616 | 0.213 | −0.036 | −0.462 | 0.296 | 0.438 | −0.003 | 0.615 | 0.206 | −0.025 | −0.462 | 0.306 |
| Sig. (2-tailed) | 0.093 | 0.977 | 0.011 | 0.429 | 0.895 | 0.072 | 0.265 | 0.089 | 0.992 | 0.011 | 0.445 | 0.926 | 0.072 | 0.249 | |
| Sep–Oct 2021 | Pearson Correlation | 0.278 | 0.084 | 0.321 | −0.142 | −0.292 | 0.048 | 0.123 | 0.276 | 0.084 | 0.319 | −0.141 | −0.290 | 0.049 | 0.121 |
| Sig. (2-tailed) | 0.297 | 0.758 | 0.226 | 0.599 | 0.273 | 0.859 | 0.650 | 0.300 | 0.757 | 0.228 | 0.602 | 0.275 | 0.857 | 0.656 | |
| Nov–Dic 2021 | Pearson Correlation | 0.383 | 0.283 | 0.425 | −0.192 | −0.313 | 0.115 | −0.011 | 0.386 | 0.288 | 0.430 | −0.192 | −0.314 | 0.116 | −0.011 |
| Sig. (2-tailed) | 0.143 | 0.288 | 0.101 | 0.477 | 0.239 | 0.671 | 0.968 | 0.139 | 0.279 | 0.097 | 0.475 | 0.236 | 0.669 | 0.969 | |
| Jan–Feb 2022 | Pearson Correlation | −0.247 | −0.312 | −0.268 | 0.014 | 0.212 | 0.118 | 0.474 | −0.252 | −0.312 | −0.271 | 0.006 | 0.205 | 0.115 | 0.468 |
| Sig. (2-tailed) | 0.356 | 0.239 | 0.316 | 0.960 | 0.430 | 0.664 | 0.064 | 0.347 | 0.239 | 0.309 | 0.983 | 0.447 | 0.672 | 0.067 | |
| Mar–Apr 2022 | Pearson Correlation | −0.343 | 0.183 | −0.132 | 0.564 | 0.470 | 0.168 | −0.137 | −0.348 | 0.182 | −0.136 | 0.561 | 0.474 | 0.168 | −0.137 |
| Sig. (2-tailed) | 0.193 | 0.497 | 0.627 | 0.023 | 0.066 | 0.534 | 0.614 | 0.187 | 0.500 | 0.616 | 0.024 | 0.064 | 0.535 | 0.612 | |
| May–Jun 2022 | Pearson Correlation | −0.121 | 0.153 | 0.012 | 0.269 | −0.012 | 0.391 | −0.423 | −0.118 | 0.166 | 0.023 | 0.284 | 0.004 | 0.389 | −0.420 |
| Sig. (2-tailed) | 0.656 | 0.571 | 0.964 | 0.313 | 0.964 | 0.134 | 0.103 | 0.662 | 0.539 | 0.932 | 0.286 | 0.987 | 0.136 | 0.106 | |
| Jul–Aug 2022 | Pearson Correlation | −0.400 | 0.390 | −0.112 | 0.362 | −0.430 | 0.077 | 0.537 | −0.400 | 0.390 | −0.112 | 0.362 | −0.430 | 0.077 | 0.537 |
| Sig. (2-tailed) | 0.125 | 0.136 | 0.680 | 0.168 | 0.096 | 0.778 | 0.032 | 0.125 | 0.136 | 0.680 | 0.168 | 0.096 | 0.778 | 0.032 | |
| Sep–Oct 2022 | Pearson Correlation | 0.103 | 0.300 | 0.196 | 0.162 | 0.384 | −0.323 | 0.0001 | 0.104 | 0.292 | 0.193 | 0.159 | 0.383 | −0.323 | −0.002 |
| Sig. (2-tailed) | 0.704 | 0.259 | 0.467 | 0.548 | 0.142 | 0.222 | 0.999 | 0.702 | 0.273 | 0.473 | 0.556 | 0.143 | 0.222 | 0.995 | |
| Nov–Dec 2022 | Pearson Correlation | 0.363 | 0.096 | 0.376 | −0.128 | 0.345 | −0.091 | 0.224 | 0.221 | 0.137 | 0.227 | −0.164 | 0.377 | −0.114 | 0.467 |
| Sig. (2-tailed) | 0.167 | 0.723 | 0.151 | 0.637 | 0.191 | 0.737 | 0.403 | 0.449 | 0.625 | 0.416 | 0.559 | 0.166 | 0.687 | 0.079 | |
| Jan–Feb 2023 | Pearson Correlation | 0.552 | 0.718 | 0.654 | 0.508 | 0.810 | 0.099 | 0.321 | 0.552 | 0.718 | 0.654 | 0.508 | 0.810 | 0.099 | 0.321 |
| Sig. (2-tailed) | 0.027 | 0.002 | 0.006 | 0.045 | 0.0001 | 0.716 | 0.225 | 0.027 | 0.002 | 0.006 | 0.045 | 0.0001 | 0.716 | 0.225 | |
| Mar–Apr 2023 | Pearson Correlation | −0.247 | −0.403 | −0.372 | −0.268 | −0.244 | −0.142 | −0.228 | −0.240 | −0.401 | −0.365 | −0.272 | −0.239 | −0.135 | −0.234 |
| Sig. (2-tailed) | 0.357 | 0.121 | 0.156 | 0.316 | 0.362 | 0.601 | 0.395 | 0.372 | 0.123 | 0.164 | 0.308 | 0.373 | 0.618 | 0.383 | |
| Date | Ambrosia Beetles | Bark Beetles | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WEST SUSSEX | ||||||||||||||||||||||||
| Xylosandrus germanus | Anisandrus dispar | Xyleborinus saxesenii | Xyleborus dryographus | Trypodendron domesticum | Trypodendron signatum | Dryocoetes villosus | Hylesinus wachtli | |||||||||||||||||
| Gosden5 | Gosden6 | Hollist | Gosden5 | Gosden6 | Hollist | Gosden5 | Gosden6 | Hollist | Gosden5 | Gosden6 | Hollist | Gosden5 | Gosden6 | Hollist | Gosden5 | Gosden6 | Hollist | Gosden5 | Gosden6 | Hollist | Gosden5 | Gosden6 | Hollist | |
| 22/06/20 | 100 | 60 | 118 | 1 | 11 | 1 | 2 | 1 | 4 | 2 | ||||||||||||||
| 17/07/20 | 52 | 57 | 96 | 3 | 10 | 3 | ||||||||||||||||||
| DEVON | ||||||||||||||||||||||||
| Xylosandrus germanus | Anisandrus dispar | Xyleborinus saxesenii | Xyleborus dryographus | Trypodendron domesticum | Trypodendron signatum | Dryocoetes villosus | Hylesinus wachtli | |||||||||||||||||
| upper | lower | coppice | upper | lower | coppice | upper | lower | coppice | upper | lower | coppice | upper | lower | coppice | upper | lower | coppice | upper | lower | coppice | upper | lower | coppice | |
| 21/05/21 | 1 | 2 | 1 | 3 | 2 | |||||||||||||||||||
| 07/06/21 | 39 | 80 | 8 | 2 | 10 | 17 | 3 | 12 | 5 | 3 | 2 | 1 | 1 | 7 | 1 | 2 | ||||||||
| 14/06/21 | 1 | 94 | 60 | 18 | 7 | 28 | 5 | 2 | 3 | 15 | 3 | 5 | 1 | 12 | 2 | 1 | ||||||||
| 02/07/21 | 1 | 15 | 2 | 1 | 1 | 10 | 1 | 1 | 6 | 2 | 6 | |||||||||||||
| 16/07/21 | 7 | 4 | 3 | 2 | 2 | 3 | ||||||||||||||||||
| 31/07/21 | 1 | 11 | 3 | 2 | 1 | 1 | 1 | |||||||||||||||||
| LONDON | ||||||||||||||||||||||||
| Xylosandrus germanus | Anisandrus dispar | Xyleborinus saxesenii | Xyleborus dryographus | Trypodendron domesticum | Trypodendron signatum | Dryocoetes villosus | Hylesinus wachtli | |||||||||||||||||
| avenue | copse | woodland | avenue | copse | woodland | avenue | copse | woodland | avenue | copse | woodland | avenue | copse | woodland | avenue | copse | woodland | avenue | copse | woodland | avenue | copse | woodland | |
| 07/06/21 | 5 | 8 | 34 | 26 | 34 | |||||||||||||||||||
| 14/06/21 | 2 | 7 | 23 | 1 | 18 | |||||||||||||||||||
| 29/06/21 | 2 | 6 | 1 | 8 | ||||||||||||||||||||
| 16/07/21 | 1 | |||||||||||||||||||||||
| Processed for Cryphonectria by real-time and negative. | ||||||||||||||||||||||||
| Processed for Cryphonectria by real-time and positive. | ||||||||||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romon-Ochoa, P.; Samal, P.; Pace, T.; Newman, T.; Oram, M.; Baxter, N.; Manning, J.A.S.; Biddle, M.; Barnard, K.; Inward, D.; et al. Forecasting of Airborne Conidia Quantities and Potential Insect Associations of Cryphonectria parasitica, the Causal Agent of Chestnut Blight, in England. J. Fungi 2024, 10, 181. https://doi.org/10.3390/jof10030181
Romon-Ochoa P, Samal P, Pace T, Newman T, Oram M, Baxter N, Manning JAS, Biddle M, Barnard K, Inward D, et al. Forecasting of Airborne Conidia Quantities and Potential Insect Associations of Cryphonectria parasitica, the Causal Agent of Chestnut Blight, in England. Journal of Fungi. 2024; 10(3):181. https://doi.org/10.3390/jof10030181
Chicago/Turabian StyleRomon-Ochoa, Pedro, Pankajini Samal, Tom Pace, Tim Newman, Mark Oram, Nicholas Baxter, John A. S. Manning, Mick Biddle, Kerry Barnard, Daegan Inward, and et al. 2024. "Forecasting of Airborne Conidia Quantities and Potential Insect Associations of Cryphonectria parasitica, the Causal Agent of Chestnut Blight, in England" Journal of Fungi 10, no. 3: 181. https://doi.org/10.3390/jof10030181
APA StyleRomon-Ochoa, P., Samal, P., Pace, T., Newman, T., Oram, M., Baxter, N., Manning, J. A. S., Biddle, M., Barnard, K., Inward, D., Taylor, P., Hendry, S., Pérez-Sierra, A., & Ward, L. (2024). Forecasting of Airborne Conidia Quantities and Potential Insect Associations of Cryphonectria parasitica, the Causal Agent of Chestnut Blight, in England. Journal of Fungi, 10(3), 181. https://doi.org/10.3390/jof10030181