
Virulence Spectra of Hungarian Pyrenophora teres f. teres Isolates Collected from Experimental Fields Show Continuous Variation without Specific Isolate × Barley Differential Interactions

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolate Collection and Isolation
2.2. DNA Extraction and PCR-Based Identification
2.3. Inoculum Production
2.4. Barley Genotypes
2.5. Inoculation
2.6. Scoring of the Infection Responses and Identification of the Pathotypes
2.7. Data Analysis
3. Results
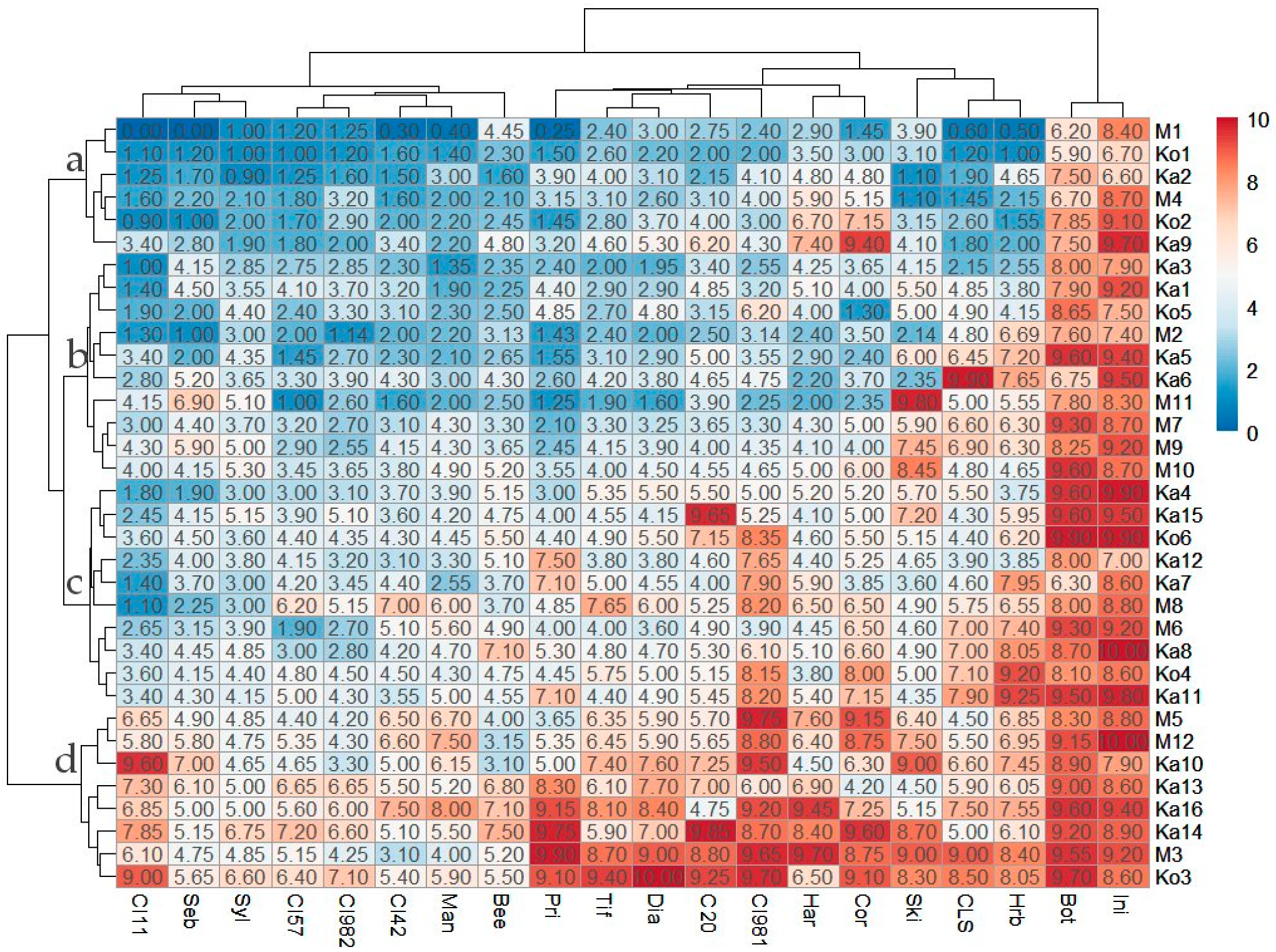
3.1. Virulence Quantification
3.2. Pathotypes
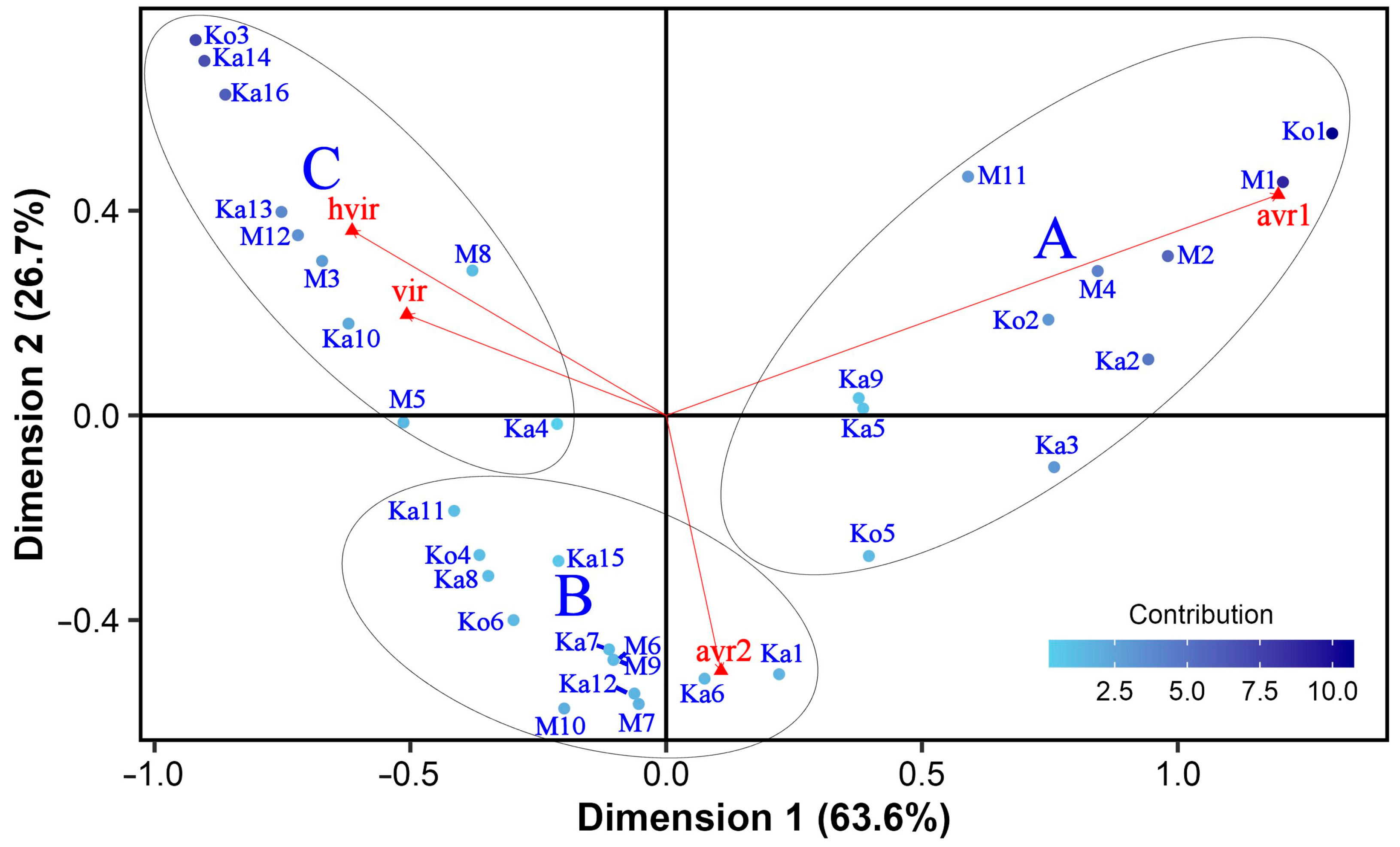
3.3. General Virulence and Specificity of the Isolates and Isolate × Differential Interactions
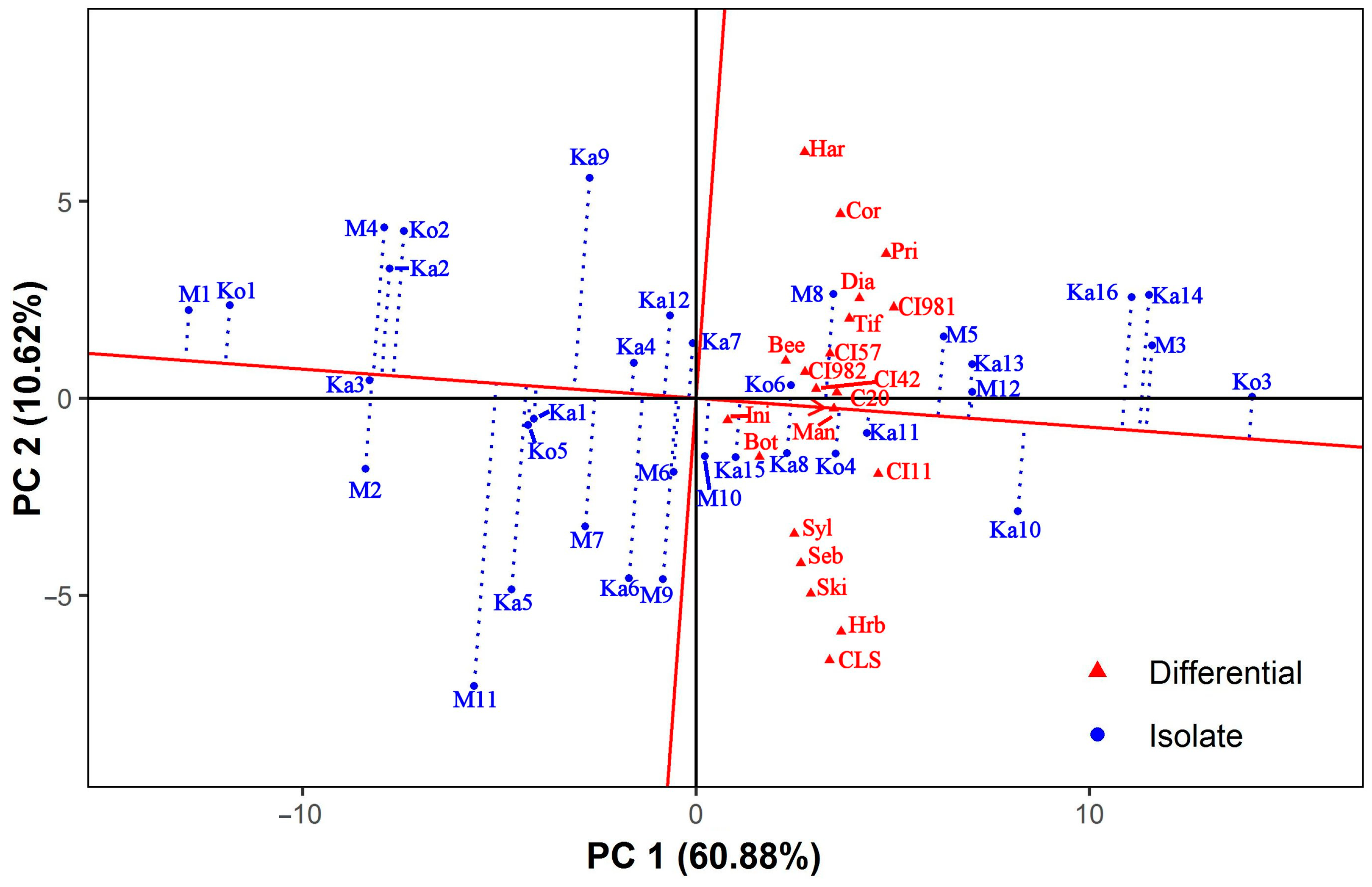
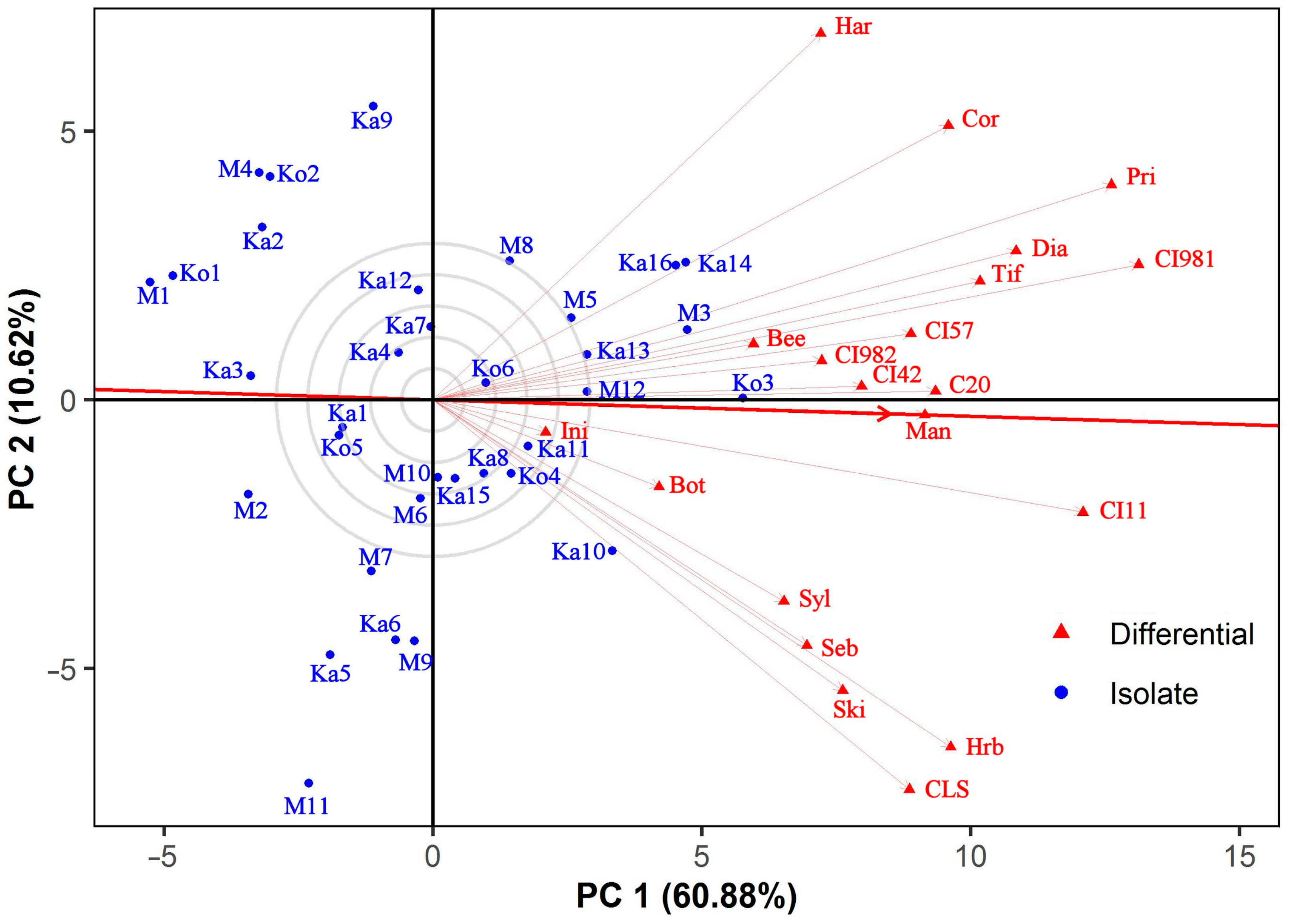
3.4. Discrimitiveness of the Barley Differentials
3.5. Classification of Ptt Isolates and Barley Differentials
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smedegård-Petersen, V. Pyrenophora teres f. maculata f. nov. and Pyrenophora teres f. teres on barley in Denmark; Royal Veterinary and Agriculture University: Copenhagen, Denmark, 1971. [Google Scholar]
- Van den Berg, C.G.J. Epidemiology of Pyrenophora teres and Its Effect on Grain Yield of Hordeum vulgare. Ph.D. Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 1988. Available online: https://harvest.usask.ca/items/77471020-1cbb-4891-8e70-cccc0810723c (accessed on 19 February 2024).
- Liu, Z.; Ellwood, S.R.; Oliver, R.P.; Friesen, T.L. Pyrenophora teres: Profile of an increasingly damaging barley pathogen. Mol. Plant Pathol. 2011, 12, 1–19. [Google Scholar] [CrossRef]
- Khan, T.N. Relationship between net blotch (Drechslera teres) and losses in grain yield of barley in Western Australia. Aust. J. Agric. Res. 1987, 38, 671–679. [Google Scholar] [CrossRef]
- Steffenson, B.J.; Webster, R.K.; Jackson, L.F. Reduction in yield loss using incomplete resistance to Pyrenophora teres f. teres in barley. Plant Dis. 1991, 75, 96–100. [Google Scholar] [CrossRef]
- Mathre, D.E. Compendium of Barley Diseases, 2nd ed.; American Phytopathological Society: St. Paul, MN, USA, 1997; pp. 1–120. [Google Scholar]
- Murray, G.M.; Brennan, J.P. Estimating disease losses to the Australian barley industry. Australas. Plant Pathol. 2010, 39, 85–96. [Google Scholar] [CrossRef]
- Grewal, T.S.; Rossnagel, B.G.; Pozniak, C.J.; Scoles, G.J. Mapping quantitative trait loci associated with barley net blotch resistance. Theor. Appl. Genet. 2008, 116, 529–539. [Google Scholar] [CrossRef]
- Ho, K.M.; Tekauz, A.; Choo, T.M.; Martin, R.A. Genetic studies on net blotch resistance in a barley cross. Can. J. Plant Sci. 1996, 76, 715–719. [Google Scholar] [CrossRef]
- Steffenson, B.J.; Hayes, P.M.; Kleinhofs, A. Genetics of seedling and adult plant resistance to net blotch (Pyrenophora teres f. teres) and spot blotch (Cochliobolus sativus) in barley. Theor. Appl. Genet. 1996, 92, 552–558. [Google Scholar] [CrossRef]
- Afanasenko, O.S.; Makarova, I.G.; Zubkovich, A.A. The number of genes controlling resistance to Pyrenophora teres Drechs. strains in barley. Russ. J. Genet. 1999, 35, 274–283. [Google Scholar]
- Cakir, M.; Gupta, S.; Platz, G.J.; Ablett, G.A.; Loughman, R.; Embiri, L.C.; Poulsen, D.; Li, C.D.; Lance, R.C.M.; Galwey, N.W.; et al. Mapping and validation of the genes for resistance to Pyrenophora teres f. teres in barley (Hordeum vulgare L.). Aust. J. Agric. Res. 2003, 54, 1369–1377. [Google Scholar] [CrossRef]
- Manninen, O.M.; Jalli, M.; Kalendar, R.; Schulman, A.; Afanasenko, O.; Robinson, J. Mapping of major spot-type and net-type net blotch resistance genes in the Ethiopian barley line CI 9819. Genome 2006, 49, 1564–1571. [Google Scholar] [CrossRef]
- Lehmensiek, A.; Platz, G.J.; Mace, E.; Poulsen, D.; Sutherland, M.W. Mapping of adult plant resistance to net form of net blotch in three Australian barley populations. Aust. J. Agric. Res. 2007, 58, 1191–1197. [Google Scholar] [CrossRef]
- König, J.; Perovic, D.; Kopahnke, D.; Ordon, F. Mapping seedling resistance to net form of net blotch (Pyrenophora teres f. teres) in barley using detached leaf assay. Plant Breeding 2014, 133, 356–365. [Google Scholar] [CrossRef]
- Novakazi, F.; Afanasenko, O.S.; Anisimova, A.; Platz, G.J.; Snowdon, R.; Kovaleva, O.; Zubkovich, A.; Ordon, F. Genetic analysis of a worldwide barley collection for resistance to net form of net blotch disease (Pyrenophora teres f. teres). Theor. Appl. Genet. 2019, 132, 2633–2650. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, R.; Bryngelsson, T.; Jalli, M.; Gustafsson, M. Effect of growth stage on resistance to Drechslera teres f. teres in barley. J. Phytopathol. 1998, 146, 261–265. [Google Scholar] [CrossRef]
- Steffenson, B.J.; Webster, R.K. Quantitative resistance to Pyrenophora teres f. teres in barley. Phytopathology 1992, 82, 407–411. [Google Scholar] [CrossRef]
- Afanasenko, O.; Mironenko, N.; Filatova, O.; Kopahnke, D.; Krämer, I.; Ordon, F. Genetics of host-pathogen interactions in the Pyrenophora teres f. teres (net form)–barley (Hordeum vulgare) pathosystem. Eur. J. Plant Pathol. 2007, 117, 267–280. [Google Scholar] [CrossRef]
- Qamar, M.A.; Liu, Z.H.; Faris, J.D.; Chao, S.; Edwards, M.C.; Lai, Z.; Franckowiak, J.D.; Friesen, T.L. A region of barley chromosome 6H harbors multiple major genes associated with net type net blotch resistance. Theor. Appl. Genet. 2008, 117, 1261–1270. [Google Scholar] [CrossRef]
- Smedegård-Petersen, V. Isolation of two toxins produced by Pyrenophora teres and their significance in disease development of net-spot blotch of barley. Physiol. Plant Pathol. 1977, 10, 203–211. [Google Scholar] [CrossRef]
- Sarpeleh, A.; Wallwork, H.; Catcheside, D.E.; Tate, M.E.; Able, A.J. Proteinaceous metabolites from Pyrenophora teres contribute to symptom development of barley net blotch. Phytopathology 2007, 97, 907–915. [Google Scholar] [CrossRef]
- Sarpeleh, A.; Wallwork, H.; Tate, M.E.; Catcheside, D.E.A.; Able, A.J. Initial characterization of phytotoxic proteins isolated from Pyrenophora teres. Physiol. Mol. Plant P 2008, 72, 73–79. [Google Scholar] [CrossRef]
- Friesen, T.L.; Faris, J.D.; Solomon, P.S.; Oliver, R.P. Host-specific toxins: Effectors of necrotrophic pathogenicity. Cell. Microbiol. 2008, 10, 1421–1428. [Google Scholar] [CrossRef]
- Stukenbrock, E.H.; McDonald, B.A. Population genetics of fungal and oomycete effectors involved in gene-for-gene interactions. Mol. Plant-Microbe Interact. 2009, 22, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Sarpeleh, A.; Tate, M.E.; Wallwork, H.; Catcheside, D.; Able, A.J. Characterisation of low molecular weight phytotoxins isolated from Pyrenophora teres. Physiol. Mol. Plant P 2009, 73, 154–162. [Google Scholar] [CrossRef]
- Pon, D.S. Physiologic specialization and variation in Helminthosporium teres. Phytopathology 1949, 39, 18. [Google Scholar]
- Khan, T.N.; Boyd, W.J.R. Physiologic specialization in Drechslera teres. Aust. J. Biol. Sci. 1969, 22, 1229–1235. [Google Scholar] [CrossRef]
- Steffenson, B.J.; Webster, R.K. Pathotype diversity of Pyrenophora teres f. teres on barley. Phytopathology 1992, 82, 170–177. [Google Scholar] [CrossRef]
- Douiyssi, A.; Rasmusson, D.; Roelfs, A. Responses of barley cultivars and lines to isolates of Pyrenophora teres. Plant Dis. 1998, 82, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Minařiková, V.; Polišenská, I. Analysis of populations of Pyrenophora teres on barley in the Czech Republic. Plant Protect. Sci. 1999, 35, 115–120. [Google Scholar] [CrossRef]
- Gupta, S.; Loughman, R. Current virulence of Pyrenophora teres on barley in Western Australia. Plant Dis. 2001, 85, 960–966. [Google Scholar] [CrossRef]
- Cromey, M.; Parkes, R. Pathogenic variation in Drechslera teres in New Zealand. N. Z. Plant Prot. 2003, 56, 251–256. [Google Scholar] [CrossRef]
- Arabi, M.I.E.; Al-Safadi, B.; Charbaji, T. Pathogenic variation among isolates of Pyrenophora teres, the causal agent of barley net blotch. J. Phytopathol. 2003, 151, 376–382. [Google Scholar] [CrossRef]
- Wu, H.-L.; Steffenson, B.J.; Zhong, S.; Li, Y.; Oleson, A.E. Genetic variation for virulence and RFLP markers in Pyrenophora teres. Can. J. Plant Pathol. 2003, 25, 82–90. [Google Scholar] [CrossRef]
- Tuohy, J.M.; Jalli, M.; Cooke, B.M.; O’Sullivan, E. Pathogenic variation in populations of Drechslera teres f. teres and D. teres f. maculata and differences in host cultivar responses. Eur. J. Plant Pathol. 2006, 116, 177–185. [Google Scholar] [CrossRef]
- Jebbouj, R.; El Yousfi, B. An integrated multivariate approach to net blotch of barley: Virulence quantification, pathotyping and a breeding strategy for disease resistance. Eur. J. Plant Pathol. 2010, 127, 521–544. [Google Scholar] [CrossRef]
- Boungab, K.; Belabid, L.; Fortas, Z.; Bayaa, B. Pathotype diversity among Algerian isolates of Pyrenophora teres f. teres. Phytopathol. Mediterr. 2012, 51, 577–586. [Google Scholar]
- Akhavan, A.; Turkington, T.K.; Askarian, H.; Tekauz, A.; Xi, K.; Tucker, J.R.; Kutcher, H.R.; Strelkov, S.E. Virulence of Pyrenophora teres populations in western Canada. Can. J. Plant Pathol. 2016, 38, 183–196. [Google Scholar] [CrossRef]
- Robinson, J.; Jalli, M. Diversity among Finnish net blotch isolates and resistance in barley. Euphytica 1996, 92, 81–87. [Google Scholar] [CrossRef]
- Fowler, R.A.; Platz, G.J.; Bell, K.L.; Fletcher, S.E.H.; Franckowiak, J.D.; Hickey, L.T. Pathogenic variation of Pyrenophora teres f. teres in Australia. Australas. Plant Path. 2017, 46, 115–128. [Google Scholar] [CrossRef]
- Oğuz, A.Ç.; Karakaya, A. Pathotypes of Pyrenophora teres on barley in Turkey. Phytopathol. Mediterr. 2017, 56, 224–234. [Google Scholar] [CrossRef]
- Gamba, F.M.; Šišić, A.; Finckh, M.R. Continuous variation and specific interactions in the Pyrenophora teres f. teres–barley pathosystem in Uruguay. J. Plant Dis. Prot. 2021, 128, 421–429. [Google Scholar] [CrossRef]
- Novakazi, F.; Göransson, M.; Stefánsson, T.S.; Hokka, M.; ·Jalli, M.; Hallsteinn Hallsson, J. Virulence of Icelandic Pyrenophora teres f. teres populations and resistance of Icelandic spring barley lines. J. Plant Pathol. 2022, 104, 205–213. [Google Scholar] [CrossRef]
- Afanasenko, O.S.; Hartleb, H.; Guseva, N.N. A set of differentials to characterize populations of Pyrenophora teres Drechs. for international use. J. Phytopathol. 1995, 143, 501–507. [Google Scholar] [CrossRef]
- Afanasenko, O.S.; Jalli, M.; Pinnschmidt, H.O. Development of an international standard set of barley differential genotypes for Pyrenophora teres f. teres. Plant Pathol. 2009, 58, 665–676. [Google Scholar] [CrossRef]
- Sdrovich-Horváth, E.; Viola, J. Upcoming diseases of spring barley. Magy. Mezőgazdaság 1986, 41, 11. (In Hungarian) [Google Scholar]
- Palágyi, A.; Tomcsányi, A. Natural infections of winter barley by powdery mildew and net blotch from 2003 to 2005. Gyak. Agrofórum 2006, 17, 18–20. (In Hungarian) [Google Scholar]
- Tomcsányi, A.; Szeőke, K.; Tóth, Á. Protection of winter barley. Növényvédelem 2006, 42, 87–106. (In Hungarian) [Google Scholar]
- Tóth, B.; Csősz, M.; Kopahkne, D.; Varga, J. First report on Pyrenophora teres causing lesions of wheat leaves in Hungary. Plant Pathol. 2008, 57, 385. [Google Scholar] [CrossRef]
- Ficsor, A.; Bakonyi, J.; Csősz, L.; Tomcsányi, A.; Varga, J.; Tóth, B. Occurrence of barley pathogenic Pyrenophora species and their mating types in Hungary. Cereal Res. Commun. 2014, 42, 612–619. [Google Scholar] [CrossRef]
- Ficsor, A.; Tóth, B.; Varga, J.; Csősz, M.; Tomcsányi, A.; Mészáros, K.; Kótai, É.; Bakonyi, J. Variability of Pyrenophora teres f. teres in Hungary as revealed by mating type and RAPD analyses. J. Plant Pathol. 2014, 96, 515–523. [Google Scholar] [CrossRef]
- Martin, A.; Moolhuijzen, P.; Tao, Y.; McIlroy, J.; Ellwood, S.R.; Fowler, R.A.; Platz, G.J.; Kilian, A.; Snyman, L. Genomic regions associated with virulence in Pyrenophora teres f. teres identified by genome-wide association analysis and biparental mapping. Phytopathology 2020, 110, 881–891. [Google Scholar] [CrossRef]
- Poudel, B.; Ellwood, S.R.; Testa, A.C.; McLean, M.; Sutherland, M.W.; Martin, A. Rare Pyrenophora teres hybridization events revealed by development of sequence-specific PCR markers. Phytopathology 2017, 107, 878–884. [Google Scholar] [CrossRef]
- Tekauz, A. A numerical scale to classify reactions of barley to Pyrenophora teres. Can. J. Plant Pathol. 1985, 7, 181–183. [Google Scholar] [CrossRef]
- Tekauz, A. Characterization and distribution of pathogenic variation in Pyrenophora teres f. teres and P. teres f. maculata from western Canada. Can. J. Plant Pathol. 1990, 12, 141–148. [Google Scholar] [CrossRef]
- Liu, Z.H.; Zhong, S.; Stasko, A.K.; Edwards, M.C.; Friesen, T.L. Virulence profile and genetic structure of a North Dakota population of Pyrenophora teres f. teres, the causal agent of net form net blotch of barley. Phytopathology 2012, 102, 539–546. [Google Scholar] [CrossRef]
- R-Core-Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 18 December 2023).
- Posit-Team. RStudio: Integrated Development Environment for R, Posit Software; PBC: Boston, MA, USA, 2023; Available online: http://www.posit.co/ (accessed on 18 December 2023).
- Sanogo, S.; Yang, X.B. Overview of selected multivariate statistical methods and their use in phytopathological research. Phytopathology 2004, 94, 1004–1006. [Google Scholar] [CrossRef][Green Version]
- Kohler, U.; Luniak, M. Data inspection using biplots. Stata J. 2005, 5, 208–223. [Google Scholar] [CrossRef]
- Kim, S.-K.; McKay, D.; Taylor, S.; Tolin, D.; Olatunji, B.; Timpano, K.; Abramowitz, J. The structure of obsessive compulsive symptoms and beliefs: A correspondence and biplot analysis. J. Anxiety Disord. 2016, 38, 79–87. [Google Scholar] [CrossRef]
- Gower, J.C. Euclidean distance matrices. In Multidimensional Data Analysis; de Leeuw, J., Heiser, W.J., Meulman, J.J., Critchley, F., Eds.; DSWO Press: Leiden, The Netherlands, 1986; pp. 11–22. [Google Scholar]
- Spencer, N.H. (Ed.) Why do I want to do cluster analysis? In Essentials of Multivariate Data Analysis, 1st ed.; Chapman and Hall/CRC: New York, NY, USA, 2013; pp. 87–106. [Google Scholar]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Vaissie, P.; Monge, A.; Husson, F. Factoshiny: Perform Factorial Analysis from ‘FactoMineR’ with a Shiny Application. R Package Version 2.4. 2021. Available online: https://CRAN.R-project.org/package=Factoshiny (accessed on 18 December 2023).
- Yan, W.; Falk, D.E. Biplot analysis of host-by-pathogen data. Plant Dis. 2002, 86, 1396–1401. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Kang, M. GGE Biplot Analysis: A Graphical Tool for Breeders, Geneticists, and Agronomists, 1st ed.; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Dumble, S. GGEBiplots: GGE Biplots with ‘ggplot2’. R Package Version 0.1.3. 2017. Available online: https://CRAN.R-project.org/package=GGEBiplots (accessed on 18 December 2023).
- Olivoto, T.; Lúcio, A.D. “metan: An R package for multi-environment trial analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Galindo-Villardón, M.P.G. Una alternativa de representación simultánea: HJ-Biplot. Qüestiió Quad. D’Estad. Investig. Oper. 1986, 10, 13–23. [Google Scholar]
- Hernández Suárez, M.; Molina Pérez, D.; Rodríguez-Rodríguez, E.M.; Díaz Romero, C.; Espinosa Borreguero, F.; Galindo-Villardón, P. The Compositional HJ-Biplot—A New Approach to Identifying the Links among Bioactive Compounds of Tomatoes. Int. J. Mol. Sci. 2016, 17, 1828. [Google Scholar] [CrossRef]
- Grünwald, N.J.; Goodwin, S.B.; Milgroom, M.G.; Fry, W.E. Analysis of genotypic diversity data for populations of microorganisms. Phytopathology 2003, 93, 738–746. [Google Scholar] [CrossRef]
- Koladia, V.M.; Faris, J.D.; Richards, J.K.; Brueggeman, R.; Chao, S.; Friesen, T.L. Genetic analysis of net form net blotch resistance in barley lines CIho 5791 and Tifang against a global collection of P. teres f. teres isolates. Theor. Appl. Genet. 2017, 130, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, R.; Bryngelsson, T.; Gustafsson, M. Virulence studies of Swedish net blotch isolates (Drechslera teres) and identification of resistant barley lines. Euphytica 1997, 94, 209–218. [Google Scholar] [CrossRef]
- Gamba, F.; Tekauz, A. Physiological specialization of a local population of Pyrenophora teres f. sp. teres. Agrociencia 2011, 15, 50–54. [Google Scholar] [CrossRef]
- McDonald, W.C. Heterothallism in Pyrenophora teres. Phytopathology 1963, 53, 771–773. [Google Scholar]
- Elliot, C.G. Reproduction in Fungi: Genetical and Physiological Aspects, 1st ed.; Chapman & Hall: London, UK, 1994. [Google Scholar]
- Platz, G.J.; Bell, K.L.; Rees, R.G.; Galea, V.J. Pathotype variation of the Australian net blotch population. In Barley Genetics VIII: Proceedings of the 8th International Barley Genetics Symposium, Adelaide, Australia, 22–27 October 2000; Adelaide University: Glen Osmond, Australia, 2000; pp. 160–162. [Google Scholar]
- Jalli, M. The Virulence of Finnish Pyrenophora teres f. teres Isolates and Its Implications for Resistance Breeding. Ph.D. Thesis, MTT Agrifood Research Finland, Jokioinen, Finland, 2010. Available online: http://www.mtt.fi/mtttiede/pdf/mtttiede9.pdf (accessed on 19 February 2024).
- Jánošová, M.; Kraic, J. Analysis of Pyrenophora teres (Died.) Drechs. population from Slovakia. In Diagnosis and Identification of Plant Pathogens. Developments in Plant Pathology; Dehne, H.W., Adam, G., Diekmann, M., Frahm, J., Mauler-Machnik, A., van Halteren, P., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 315–316. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | General Virulence 1 | Number of Virulent Infections | Pathotype | Virulence Class (Frequency Profile) 2 | HCPC Cluster 3 | Isolate Group 4 | |||
|---|---|---|---|---|---|---|---|---|---|
| Avirulent-1 | Avirulent-2 | Virulent | Highly Virulent | ||||||
| Karcag, Northern Great Plain of Hungary (Institute for Agricultural Research and Educational Farm of University of Debrecen) | |||||||||
| Ka1 | 4.16 | 4 | 1 | 3 | 13 | 2 | 2 | B | b |
| Ka2 | 3.07 | 2 | 2 | 10 | 8 | 1 | 1 | A | a |
| Ka3 | 3.23 | 2 | 2 | 8 | 10 | 0 | 2 | A | b |
| Ka4 | 4.74 | 11 | 3 | 2 | 7 | 9 | 2 | C | c |
| Ka5 | 4.05 | 6 | 4 | 6 | 8 | 4 | 2 | A | b |
| Ka6 | 4.63 | 5 | 5 | 2 | 13 | 2 | 3 | B | b |
| Ka7 | 4.79 | 7 | 6 | 1 | 12 | 4 | 3 | B | c |
| Ka8 | 5.55 | 10 | 7 | 0 | 10 | 7 | 3 | B | c |
| Ka9 | 4.39 | 6 | 8 | 6 | 8 | 3 | 3 | A | a |
| Ka10 | 6.54 | 15 | 9 | 0 | 5 | 9 | 6 | C | d |
| Ka11 | 5.88 | 11 | 10 | 0 | 9 | 6 | 5 | B | c |
| Ka12 | 4.67 | 6 | 11 | 1 | 13 | 3 | 3 | B | c |
| Ka13 | 6.47 | 18 | 12 | 0 | 2 | 14 | 4 | C | d |
| Ka14 | 7.44 | 20 | 13 | 0 | 0 | 10 | 10 | C | d |
| Ka15 | 5.33 | 9 | 14 | 1 | 10 | 6 | 3 | B | c |
| Ka16 | 7.33 | 19 | 15 | 0 | 1 | 8 | 11 | C | d |
| Mean of the Isolates from Karcag | 5.14 ± 2.29 | 9.44 ± 5.85 | |||||||
| Kompolt, Northern Hungary (Fleischmann Rudolf Research Institute, Eszterházy Károly University) | |||||||||
| Ko1 | 2.28 | 2 | 4 | 14 | 4 | 2 | 0 | A | a |
| Ko2 | 3.41 | 4 | 16 | 9 | 7 | 2 | 2 | A | a |
| Ko3 | 7.89 | 20 | 13 | 0 | 0 | 8 | 12 | C | d |
| Ko4 | 5.67 | 10 | 17 | 0 | 10 | 5 | 5 | B | c |
| Ko5 | 3.96 | 4 | 18 | 5 | 11 | 2 | 2 | A | b |
| Ko6 | 5.53 | 9 | 19 | 0 | 11 | 6 | 3 | B | c |
| Mean of the Isolates from Kompolt | 4.79 ± 2.58 | 8.17 ± 6.59 | |||||||
| Martonvásár, Central Transdanubia (Agricultural Institute, Centre for Agricultural Research) | |||||||||
| M1 | 2.17 | 2 | 2 | 13 | 5 | 1 | 1 | A | a |
| M2 | 3.09 | 3 | 20 | 11 | 6 | 2 | 1 | A | b |
| M3 | 7.35 | 15 | 11 | 0 | 5 | 3 | 12 | C | d |
| M4 | 3.19 | 4 | 16 | 10 | 6 | 3 | 1 | A | a |
| M5 | 6.26 | 13 | 22 | 0 | 7 | 8 | 5 | C | d |
| M6 | 4.94 | 7 | 23 | 1 | 12 | 5 | 2 | B | c |
| M7 | 4.47 | 6 | 24 | 1 | 13 | 4 | 2 | B | b |
| M8 | 5.67 | 14 | 25 | 2 | 4 | 10 | 4 | C | c |
| M9 | 4.89 | 7 | 26 | 1 | 12 | 5 | 2 | B | b |
| M10 | 5.15 | 7 | 27 | 0 | 13 | 4 | 3 | B | b |
| M11 | 3.88 | 7 | 26 | 9 | 4 | 4 | 3 | A | b |
| M12 | 6.48 | 17 | 28 | 0 | 3 | 11 | 6 | C | d |
| Mean of the Isolates from Martonvásár | 4.79 ± 2.47 | 8.50 ± 4.98 | |||||||
| Overall Mean | 4.96 ± 2.41 | 8.88 ± 5.54 | |||||||
| PT | Isolate | Barley Differential | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Beecher | Canadian Lake Shore | Corvett | Diamond | Harbin | Manchurian | Prior | Skiff | Tifang | C-20019 | CI 5791 | CI 4207 | CI 9819 | CI 9825 | CI 11458 | Sylphid | Sebastian | Harrington | Botond | MV Initium | ||
| 1 | Ka1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| 2 | Ka2, Ka3, Ko1, M1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 3 | Ka4 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| 4 | Ka5 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 5 | Ka6 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 |
| 6 | Ka7 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| 7 | Ka8 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| 8 | Ka9 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| 9 | Ka10 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 1 |
| 10 | Ka11 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| 11 | Ka12 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 12 | Ka13 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 13 | Ka14, Ko3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 14 | Ka15 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 |
| 15 | Ka16 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 16 | Ko2, M4 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| 17 | Ko4 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 18 | Ko5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 19 | Ko6 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 20 | M2 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 21 | M3 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 |
| 22 | M5 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 |
| 23 | M6 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 24 | M7 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| 25 | M8 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 |
| 26 | M9, M11 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 |
| 27 | M10 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 |
| 28 | M12 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 |
| Total no. of PTs | 9 | 16 | 19 | 12 | 20 | 9 | 10 | 16 | 11 | 15 | 7 | 8 | 17 | 5 | 7 | 6 | 7 | 15 | 28 | 28 | |
| Isolate Group | Barley Differential | ||||
|---|---|---|---|---|---|
| Diamond | Harbin | Tifang | CI 9819 | CI 11458 | |
| IG ‘a’ | 1/0 | 0 | 0 | 0 | 0 |
| IG ‘b’ | 0 | 1/0 | 0 | 1/0 | 0 |
| IG ‘c’ | 1/0 | 1/0 | 1/0 | 1/0 | 0 |
| IG ‘d’ | 1 | 1 | 1 | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakonyi, J.; Seress, D.; Nagy, Z.Á.; Csorba, I.; Cséplő, M.; Marton, T.A.; Martin, A.; Mészáros, K. Virulence Spectra of Hungarian Pyrenophora teres f. teres Isolates Collected from Experimental Fields Show Continuous Variation without Specific Isolate × Barley Differential Interactions. J. Fungi 2024, 10, 184. https://doi.org/10.3390/jof10030184
Bakonyi J, Seress D, Nagy ZÁ, Csorba I, Cséplő M, Marton TA, Martin A, Mészáros K. Virulence Spectra of Hungarian Pyrenophora teres f. teres Isolates Collected from Experimental Fields Show Continuous Variation without Specific Isolate × Barley Differential Interactions. Journal of Fungi. 2024; 10(3):184. https://doi.org/10.3390/jof10030184
Chicago/Turabian StyleBakonyi, József, Diána Seress, Zoltán Á. Nagy, Ildikó Csorba, Mónika Cséplő, Tibor A. Marton, Anke Martin, and Klára Mészáros. 2024. "Virulence Spectra of Hungarian Pyrenophora teres f. teres Isolates Collected from Experimental Fields Show Continuous Variation without Specific Isolate × Barley Differential Interactions" Journal of Fungi 10, no. 3: 184. https://doi.org/10.3390/jof10030184
APA StyleBakonyi, J., Seress, D., Nagy, Z. Á., Csorba, I., Cséplő, M., Marton, T. A., Martin, A., & Mészáros, K. (2024). Virulence Spectra of Hungarian Pyrenophora teres f. teres Isolates Collected from Experimental Fields Show Continuous Variation without Specific Isolate × Barley Differential Interactions. Journal of Fungi, 10(3), 184. https://doi.org/10.3390/jof10030184