
Establishment and Application of a Quantitative PCR Method for E248R Gene of African Swine Fever Virus

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Construct of Plasmid Containing the E248R Gene
2.2. Virus Strains and Viral Samples
2.3. Primers and Probe for the Real-Time PCR
2.4. Formulation of Standard Curve
2.5. Optimization of Reaction Conditions
2.6. Specificity, Sensitivity and Repeatability Analysis
2.7. Viral Rescue and Characteristic Analysis of Recombinant PRRSV
2.8. Detection of ASFV Gene in the Samples and Recombinant Viruses
3. Results
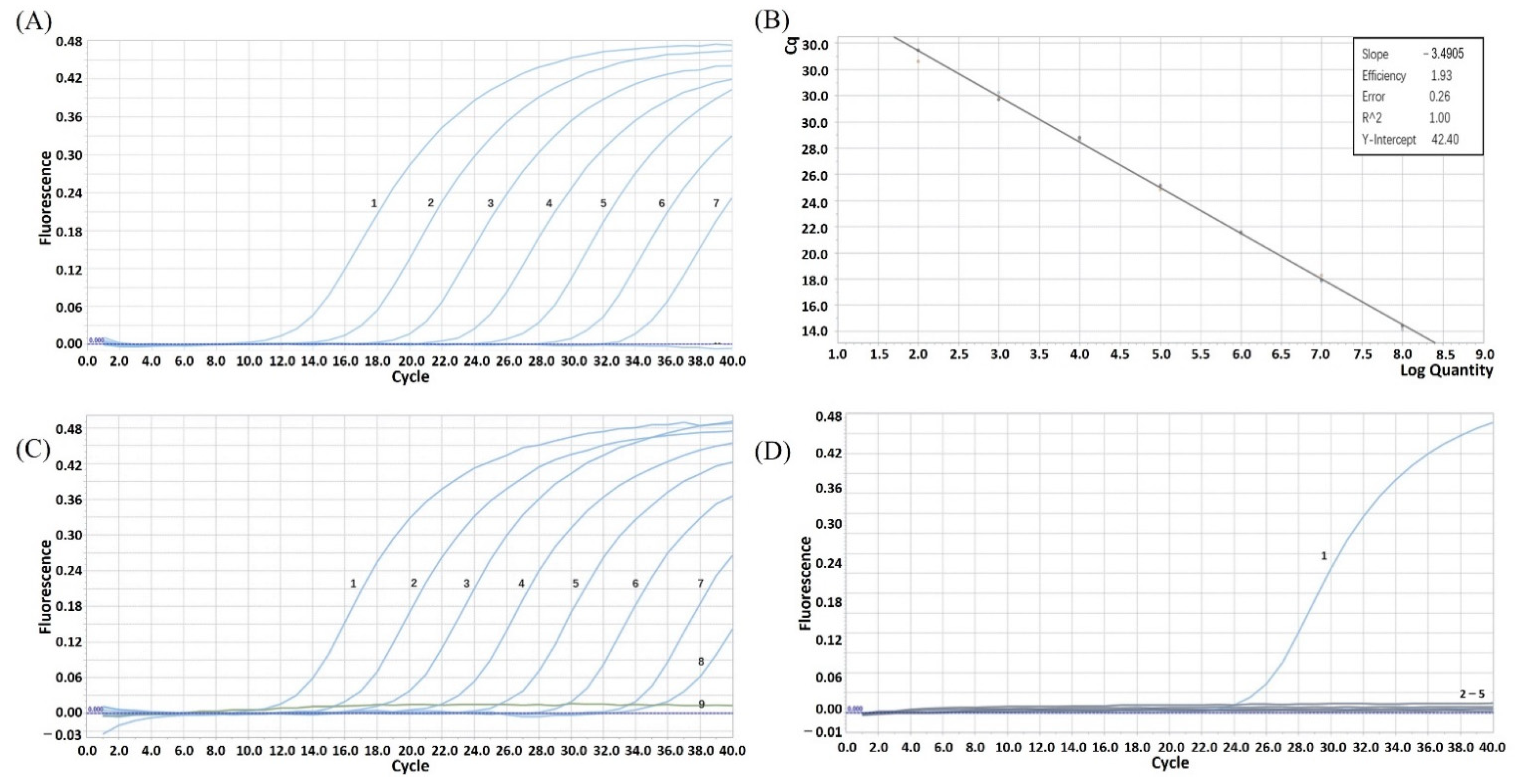
3.1. Establishment of Standard Curve
3.2. Optimization of Reaction Conditions
3.3. Analysis of Repeatability, Sensitivity and Specificity
3.4. Detection of the Serum Samples and Recombinant PRRSV-Expressing ASFV E248R
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dixon, L.K.; Sun, H.; Roberts, H. African swine fever. Antivir. Res. 2019, 165, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Galindo, I.; Alonso, C. African swine fever virus: A review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pikalo, J.; Zani, L.; Huhr, J.; Beer, M.; Blome, S. Pathogenesis of African swine fever in domestic pigs and European wild boar—Lessons learned from recent animal trials. Virus Res. 2019, 271, 197614. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vizcaino, J.M.; Mur, L.; Gomez-Villamandos, J.C.; Carrasco, L. An update on the epidemiology and pathology of African swine fever. J. Comp. Pathol. 2015, 152, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Boinas, F.S.; Wilson, A.J.; Hutchings, G.H.; Martins, C.; Dixon, L.J. The persistence of African swine fever virus in field-infected Ornithodoros erraticus during the ASF endemic period in Portugal. PLoS ONE 2011, 6, e20383. [Google Scholar] [CrossRef] [PubMed]
- Rowlands, R.J.; Michaud, V.; Heath, L.; Hutchings, G.; Oura, C.; Vosloo, W.; Dwarka, R.; Onashvili, T.; Albina, E.; Dixon, L.K. African swine fever virus isolate, Georgia, 2007. Emerg. Infect. Dis. 2008, 14, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, N.; Luo, Y.; Liu, Y.; Miao, F.; Chen, T.; Zhang, S.; Cao, P.; Li, X.; Tian, K.; et al. Emergence of African swine fever in China, 2018. Transbound. Emerg. Dis. 2018, 65, 1482–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, L.K.; Chapman, D.A.; Netherton, C.L.; Upton, C. African swine fever virus replication and genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Luo, Y.; Wang, Y.; Li, S.; Zhao, Z.; Bi, Y.; Sun, J.; Peng, R.; Song, H.; Zhu, D.; et al. Cryo-EM structure of the African swine fever virus. Cell Host Microbe 2019, 26, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Alejo, A.; Matamoros, T.; Guerra, M.; Andres, G. A proteomic atlas of the African swine fever virus particle. J. Virol. 2018, 92, e01293-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Zhao, D.; Wang, J.; Zhang, Y.; Wang, M.; Gao, Y.; Li, F.; Wang, J.; Bu, Z.; Rao, Z.; et al. Architecture of African swine fever virus and implications for viral assembly. Science 2019, 366, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, I.; Nogal, M.L.; Redrejo-Rodriguez, M.; Bustos, M.J.; Salas, M.L. The African swine fever virus virion membrane protein pE248R is required for virus infectivity and an early postentry event. J. Virol. 2009, 83, 12290–12300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazloum, A.; Zhukov, I.U.; Aronova, E.B.; Igolkin, A.S.; Vlasova, N.N. ASF virus replication features in the presence of recombinant proteins CD2v, pX69R and pE248R. Probl. Virol. 2019, 64, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernaez, B.; Alonso, C. Dynamin- and clathrin-dependent endocytosis in African swine fever virus entry. J. Virol. 2010, 84, 2100–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, C.; Borca, M.; Dixon, L.; Revilla, Y.; Rodriguez, F.; Escribano, J.M.; Ictv Report Consortium. ICTV virus taxonomy profile: Asfarviridae. J. Gen. Virol. 2018, 99, 613–614. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Jiang, Y.; Li, G.; Zhou, Y.; Yu, L.; Li, L.; Tong, W.; Zheng, H.; Zhang, Y.; Yu, H.; et al. Porcine reproductive and respiratory syndrome virus expressing E2 of classical swine fever virus protects pigs from a lethal challenge of highly-pathogenic PRRSV and CSFV. Vaccine 2018, 36, 3269–3277. [Google Scholar] [CrossRef] [PubMed]
- Gaudreault, N.N.; Richt, J.A. Subunit vaccine approaches for African swine fever virus. Vaccines 2019, 7, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.; Geng, R.; Lv, H.; Bao, C.; Shao, H.; Ye, J.; Qian, K.; Qin, A. Development of real-time PCR based on A137R gene for the detection of African swine fever virus. Front. Vet. Sci. 2021, 8, 753967. [Google Scholar] [CrossRef] [PubMed]
- King, D.P.; Reid, S.M.; Hutchings, G.H.; Grierson, S.S.; Wilkinson, P.J.; Dixon, L.K.; Bastos, A.D.; Drew, T.W. Development of a TaqMan PCR assay with internal amplification control for the detection of African swine fever virus. J. Virol. Methods 2003, 107, 53–61. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Name | Primers 5′-3′ | Position |
|---|---|---|
| E248R-F | 5′-GGAGGCTCTACAAGCAAA-3′ | 166996–167013 |
| E248R-R | 5′-CATCACCGAATACGCCTA-3′ | 167105–167122 |
| E248R-P | 5′FAM-AATACGACCAACATTATCAGCAAT-3′BHQ1-3′ | 167025–167050 |
| System Components | Volume |
|---|---|
| Probe qPCR Mix (2×) | 10 μL |
| probe | 0.4 μL |
| Primer-F | 0.6 μL |
| Primer-R | 0.6 μL |
| Template | 1 μL |
| Nuclease-free water | 7.4 μL |
| Total | 20 μL |
| Tm (°C) | Ct | Average Value ± Standard Deviation |
|---|---|---|
| 56 | 24.95 | 24.927 ± 0.068 |
| 24.98 | ||
| 24.85 | ||
| 57 | 24.70 | 24.670 ± 0.030 |
| 24.67 | ||
| 24.64 | ||
| 58 | 24.47 | 24.767 ± 0.270 |
| 25.00 | ||
| 24.83 | ||
| 59 | 24.83 | 25.097 ± 0.326 |
| 25.00 | ||
| 25.46 | ||
| 60 | 24.22 | 25.25 ± 0.198 |
| 25.11 | ||
| 25.39 |
| Standard Plasmid | Ct | Average Value | Standard Deviation | CV% |
|---|---|---|---|---|
| 105 | 24.82 | 25.017 | 0.174 | 0.695 |
| 25.15 | ||||
| 25.08 | ||||
| 106 | 21.49 | 21.520 | 0.030 | 0.139 |
| 21.55 | ||||
| 21.52 | ||||
| 107 | 17.83 | 17.997 | 0.223 | 1.239 |
| 17.91 | ||||
| 18.25 |
| Standard Plasmid | Ct | Average Value | Standard Deviation | CV% |
|---|---|---|---|---|
| 105 | 24.82 | 24.303 | 0.480 | 1.977 |
| 24.22 | ||||
| 23.87 | ||||
| 106 | 21.49 | 21.700 | 0.279 | 1.286 |
| 21.86 | ||||
| 21.79 | ||||
| 107 | 17.83 | 17.540 | 0.252 | 1.439 |
| 17.42 | ||||
| 17.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Du, N.; Chen, J.; Zhang, K.; Tong, W.; Zheng, H.; Zhao, R.; Tong, G.; Gao, F. Establishment and Application of a Quantitative PCR Method for E248R Gene of African Swine Fever Virus. Vet. Sci. 2022, 9, 417. https://doi.org/10.3390/vetsci9080417
Li L, Du N, Chen J, Zhang K, Tong W, Zheng H, Zhao R, Tong G, Gao F. Establishment and Application of a Quantitative PCR Method for E248R Gene of African Swine Fever Virus. Veterinary Sciences. 2022; 9(8):417. https://doi.org/10.3390/vetsci9080417
Chicago/Turabian StyleLi, Liwei, Nannan Du, Jinxia Chen, Kuan Zhang, Wu Tong, Haihong Zheng, Ran Zhao, Guangzhi Tong, and Fei Gao. 2022. "Establishment and Application of a Quantitative PCR Method for E248R Gene of African Swine Fever Virus" Veterinary Sciences 9, no. 8: 417. https://doi.org/10.3390/vetsci9080417
APA StyleLi, L., Du, N., Chen, J., Zhang, K., Tong, W., Zheng, H., Zhao, R., Tong, G., & Gao, F. (2022). Establishment and Application of a Quantitative PCR Method for E248R Gene of African Swine Fever Virus. Veterinary Sciences, 9(8), 417. https://doi.org/10.3390/vetsci9080417