
Recombinant Ehrlichia canis GP19 Protein as a Promising Vaccine Prototype Providing a Protective Immune Response in a Mouse Model

 ,
,  , ,
, ,  and
and 
Abstract
Simple summary
Abstract
1. Introduction
2. Materials and Methods
2.1. E. canis Organism Cultivation and Stocks
2.2. Mice
2.3. Expression of Recombinant E. canis GP19 Proteins
2.4. Vaccine Immunizations and E. canis Challenge
2.5. Measurement of rGP19 Antibody Elicits in Mice
2.6. Assessment of Ehrlichial Load in Organs by Real-Time PCR
2.7. Relevance of rGP19 on Cytokine mRNA Expression by qPCR
2.8. Preparation of Mice Splenocyte and Flow Cytometry Analysis
2.9. Data Analysis
3. Results
3.1. Recombinant GP19 Protein (rGP19) Production
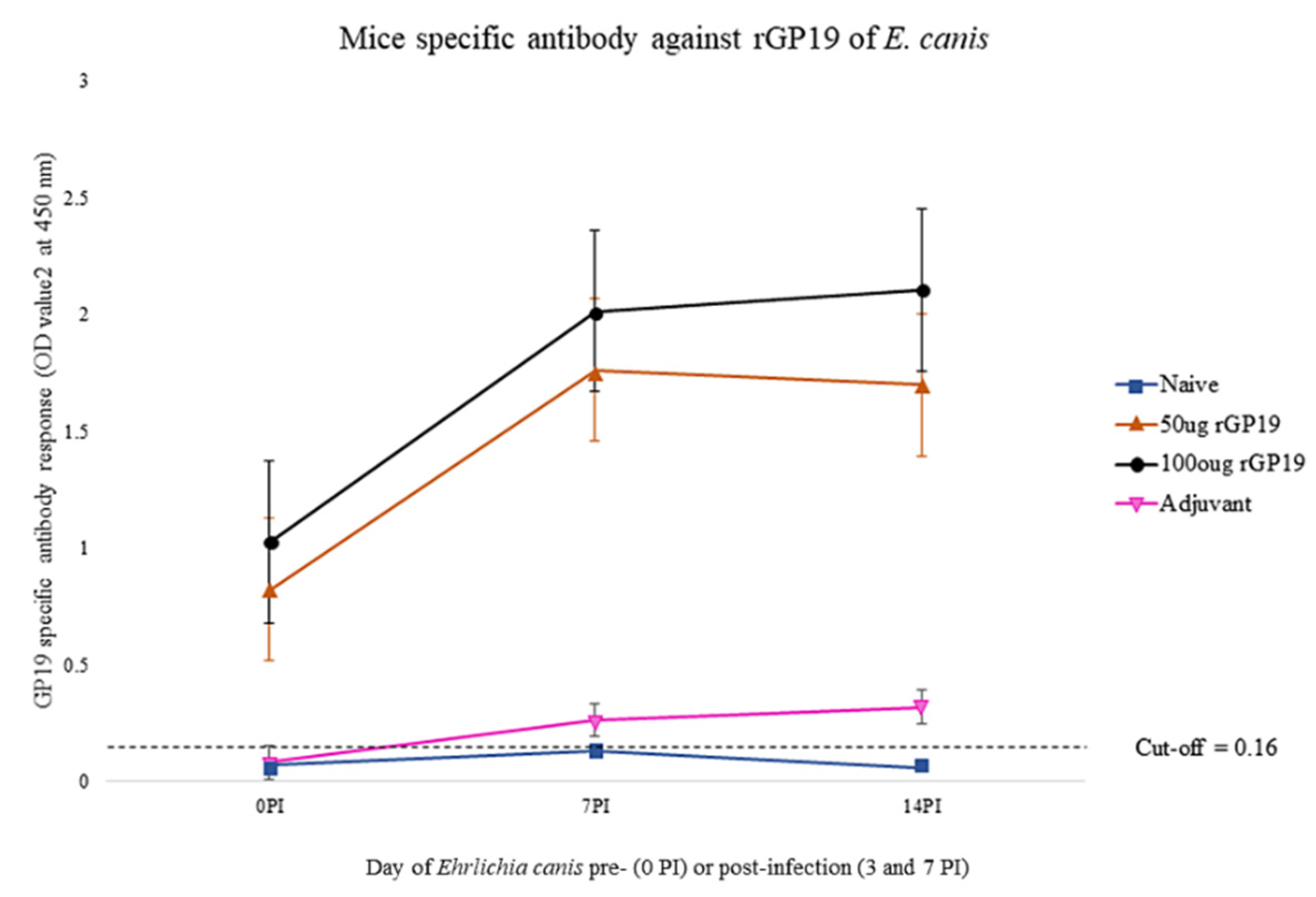
3.2. rGP19 Vaccine Prototypes Induce a Strong Antibody Response in Mice
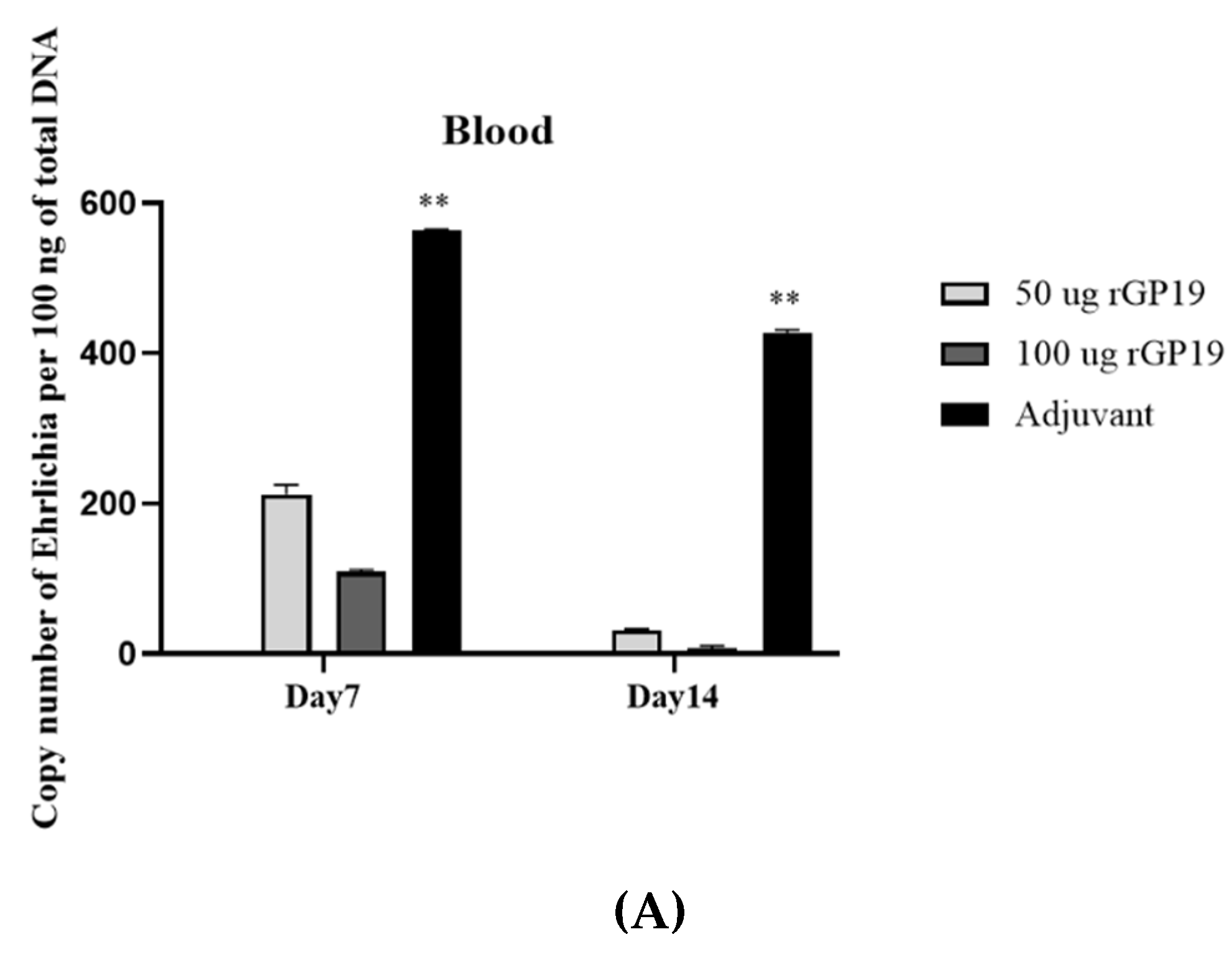
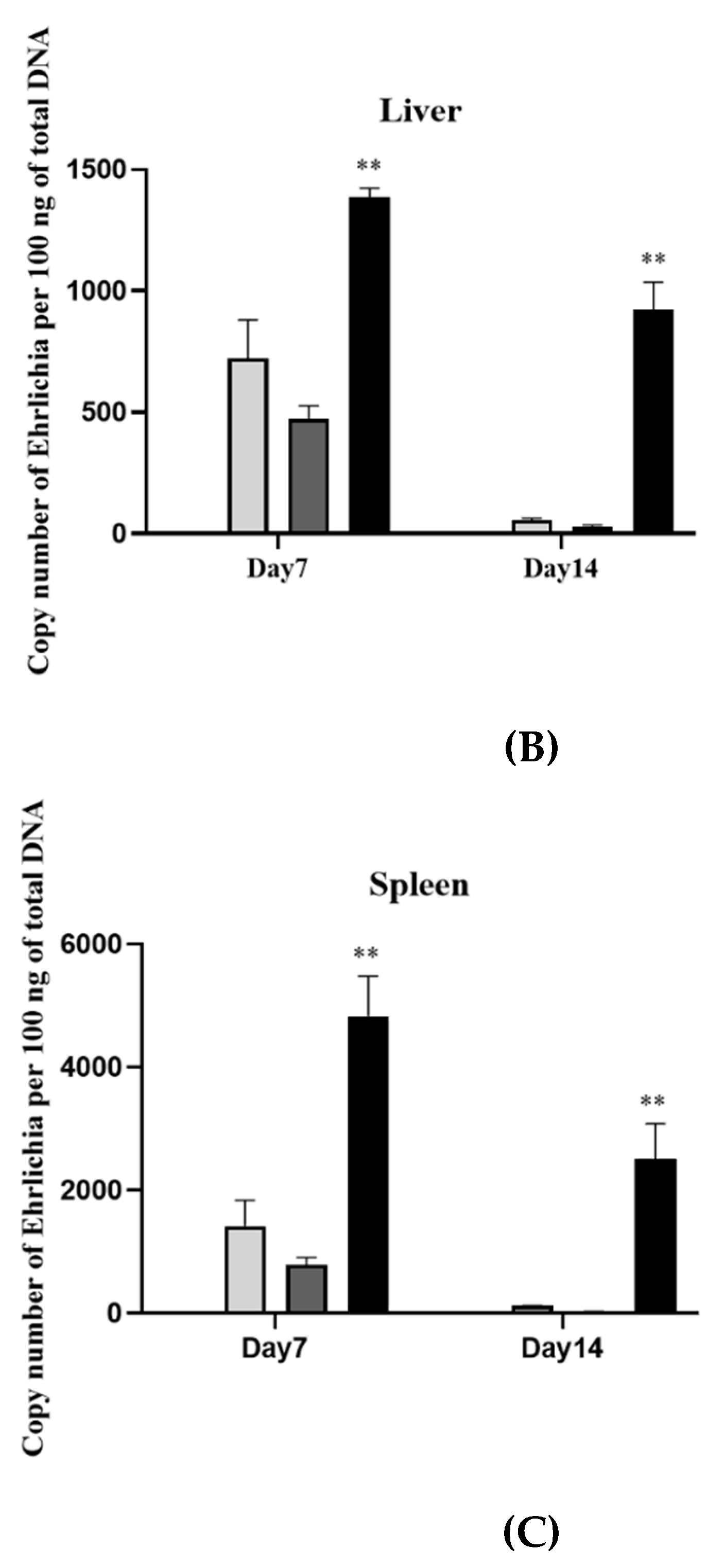
3.3. rGP19 Vaccine Prototypes Provide the Protection against the Pathogen
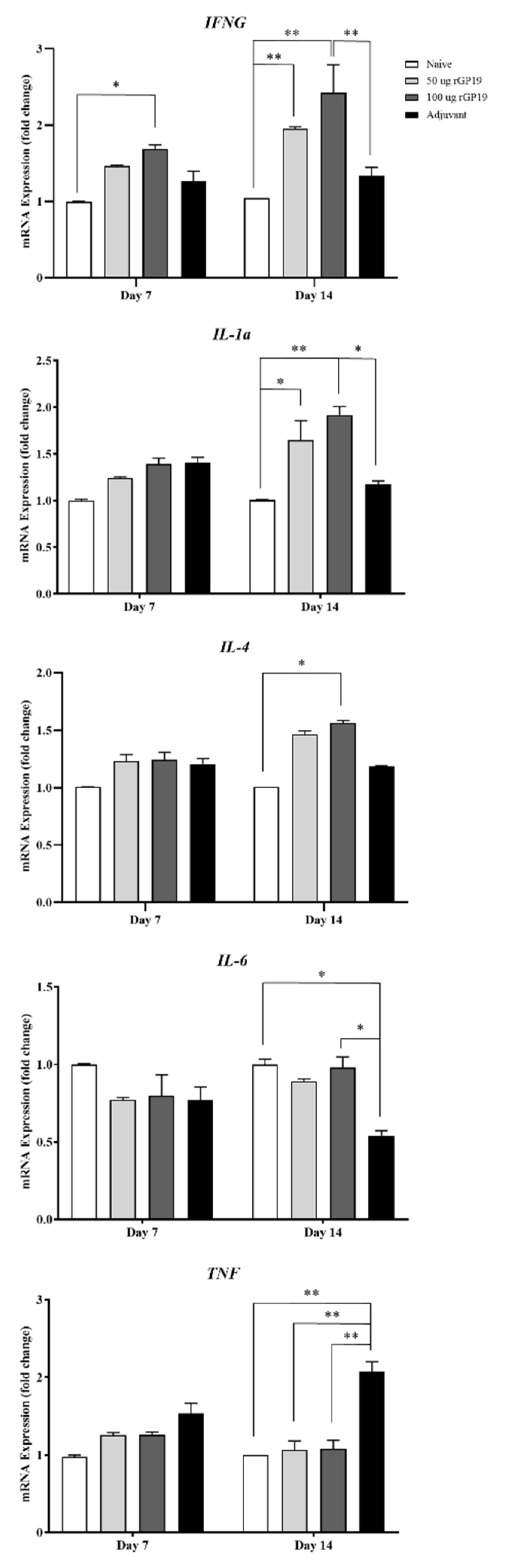
3.4. Relevance of rGP19 on Cytokine-Relative Gene Expression by qPCR
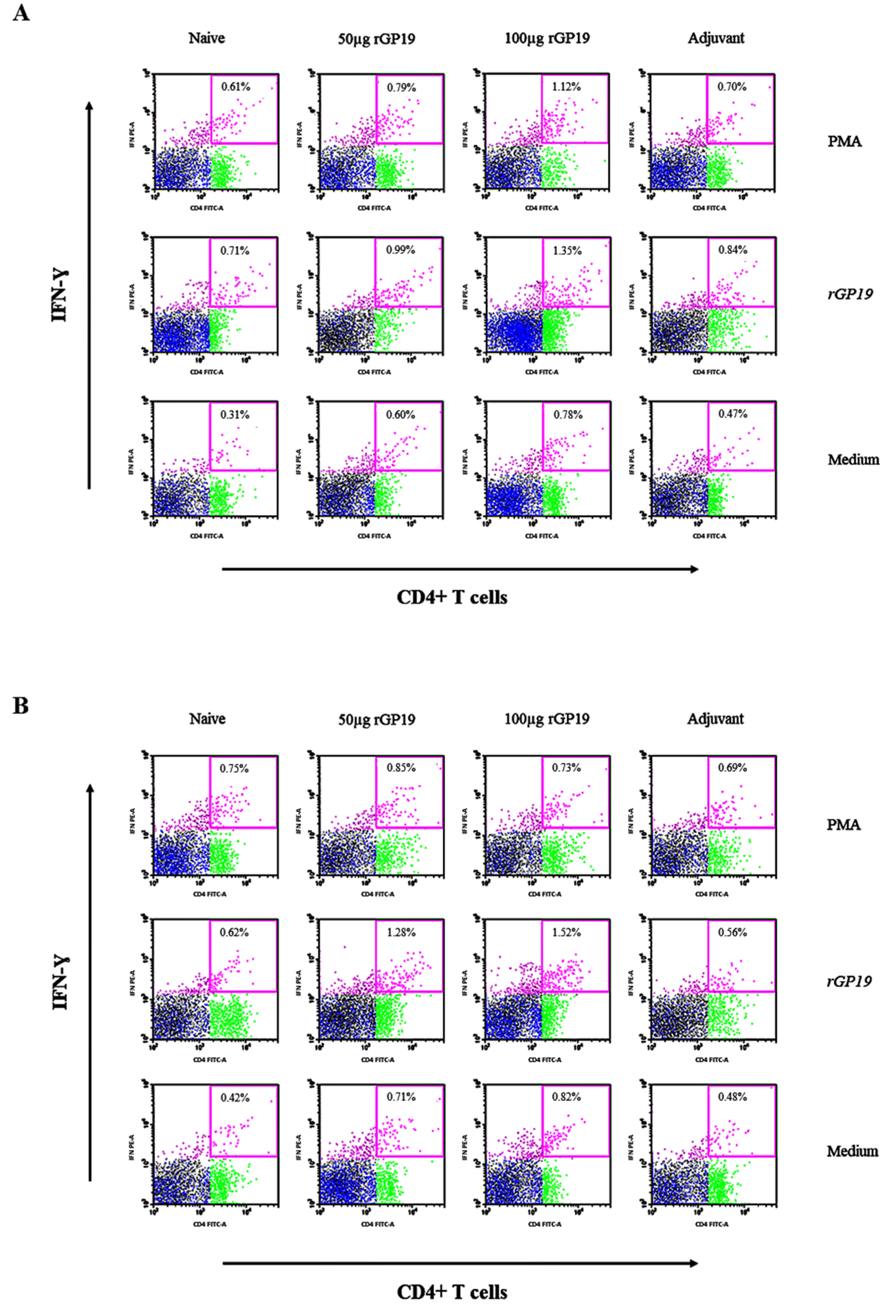
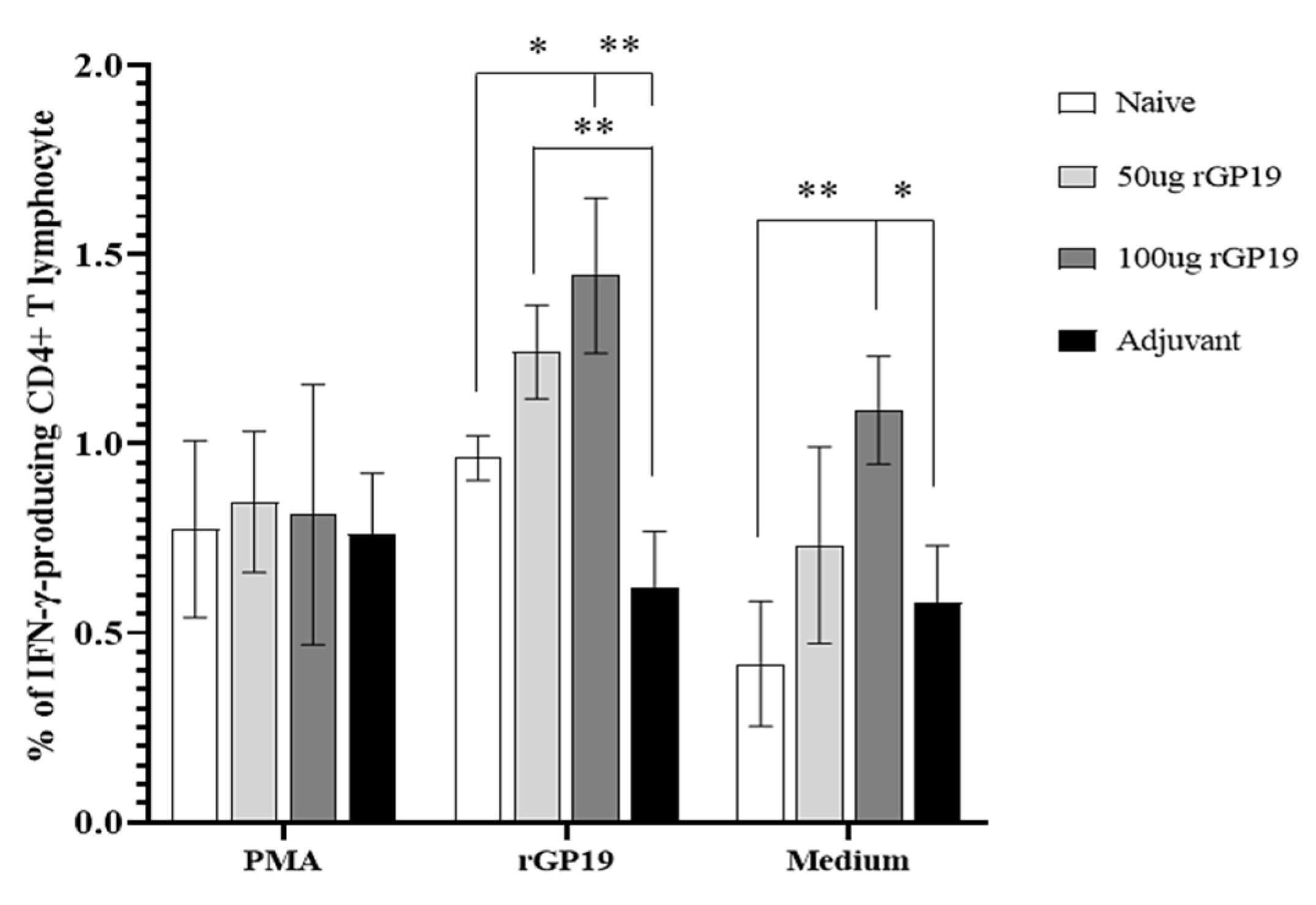
3.5. rGP19 Manipulated Specific Memory CD4+ Th1 Responses during E. canis Infection
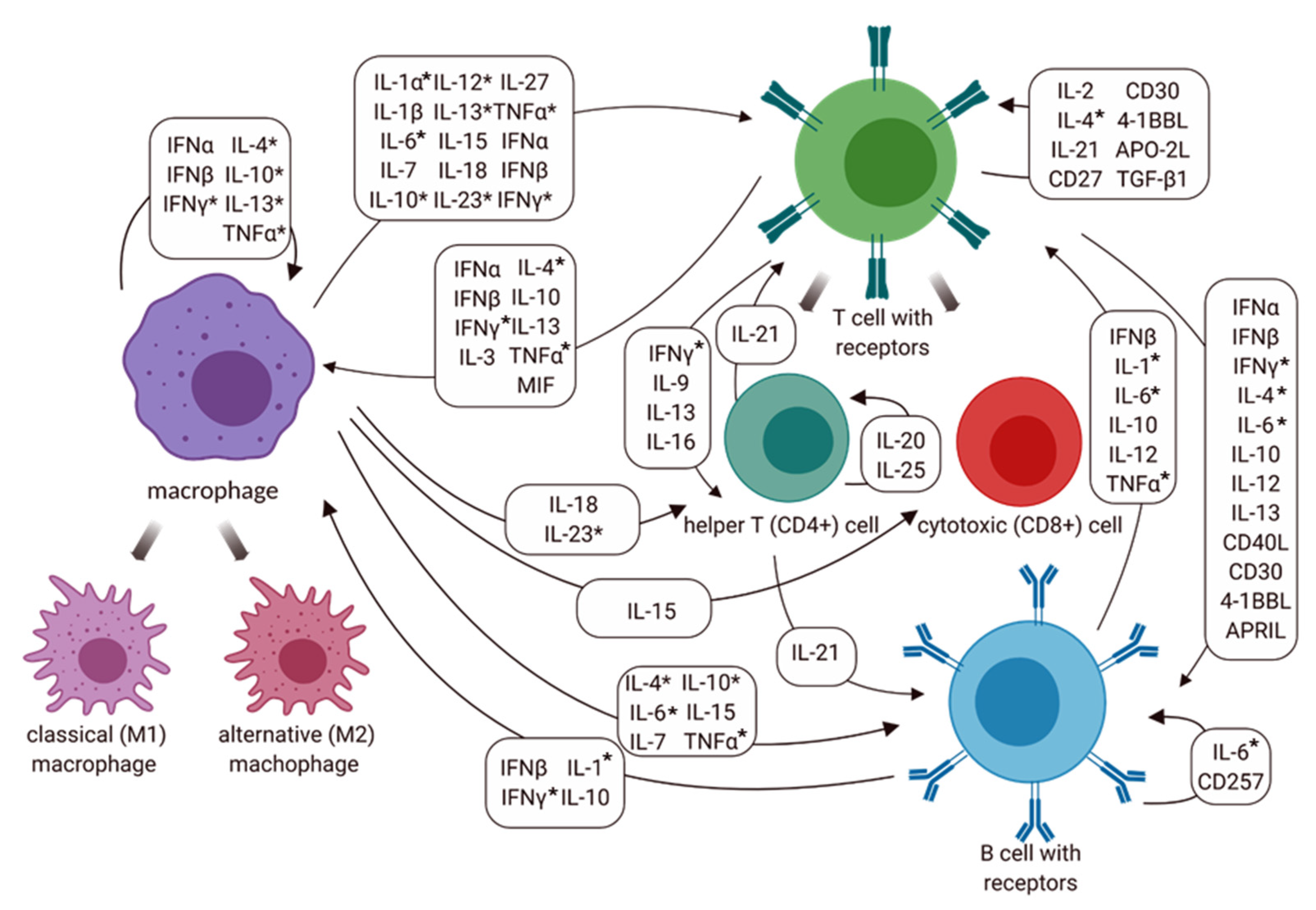
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kawahara, M.; Suto, C.; Rikihisa, Y.; Yamamoto, S.; Tsuboi, Y. Characterization of ehrlichial organisms isolated from a wild mouse. J. Clin. Microbiol. 1993, 31, 89–96. [Google Scholar] [CrossRef]
- Perez, M.; Bodor, M.; Zhang, C.; Xiong, Q.; Rikihisa, Y. Human infection with Ehrlichia canis accompanied by clinical signs in Venezuela. Ann. N. Y. Acad. Sci. 2006, 1078, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Mahan, S.; Kelly, P.J.; Mahan, S.M. A preliminary study to evaluate the immune responses induced by immunization of dogs with inactivated Ehrlichia canis organisms. Onderstepoort J. Vet. Res. 2005, 72, 119–128. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rudoler, N.; Baneth, G.; Eyal, O.; van Straten, M.; Harrus, S. Evaluation of an attenuated strain of Ehrlichia canis as a vaccine for canine monocytic ehrlichiosis. Vaccine 2012, 31, 226–233. [Google Scholar] [CrossRef]
- Shams, H. Recent developments in veterinary vaccinology. Vet. J. 2005, 170, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Thirumalapura, N.R.; Crocquet-Valdes, P.A.; Saito, T.B.; Thomas, S.; McBride, J.W.; Walker, D.H. Recombinant Ehrlichia P29 protein induces a protective immune response in a mouse model of ehrlichiosis. Vaccine 2013, 31, 5960–5967. [Google Scholar] [CrossRef][Green Version]
- McBride, J.W.; Corstvet, R.E.; Gaunt, S.D.; Boudreaux, C.; Guedry, T.; Walker, D.H. Kinetics of antibody response to Ehrlichia canis immunoreactive proteins. Infect. Immun. 2003, 71, 2516–2524. [Google Scholar] [CrossRef]
- Zhang, X.; Luo, T.; Keysary, A.; Baneth, G.; Miyashiro, S.; Strenger, C.; Waner, T.; McBride, J.W. Genetic and antigenic diversities of major immunoreactive proteins in globally distributed Ehrlichia canis strains. Clin. Vaccine Immunol. 2008, 15, 1080–1088. [Google Scholar] [CrossRef]
- Hsieh, Y.C.; Lee, C.C.; Tsang, C.L.; Chung, Y.T. Detection and characterization of four novel genotypes of Ehrlichia canis from dogs. Vet. Microbiol. 2010, 146, 70–75. [Google Scholar] [CrossRef]
- Nambooppha, B.; Rittipornlertrak, A.; Tattiyapong, M.; Tangtrongsup, S.; Tiwananthagorn, S.; Chung, Y.T.; Sthitmatee, N. Two different genogroups of Ehrlichia canis from dogs in Thailand using immunodominant protein genes. Infect. Genet. Evol. 2018, 63, 116–125. [Google Scholar] [CrossRef]
- McBride, J.W.; Doyle, C.K.; Zhang, X.; Cardenas, A.M.; Popov, V.L.; Nethery, K.A.; Woods, M.E. Identification of a glycosylated Ehrlichia canis 19-kilodalton major immunoreactive protein with a species-specific serine-rich glycopeptide epitope. Infect. Immun. 2007, 75, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Keysary, A.; Waner, T.; Strenger, C.; Harrus, S. Cultivation of Ehrlichia canis in a continuous BALB/C mouse macrophage cell culture line. J. Vet. Diagn. Investig. 2001, 13, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Nambooppha, B.; Rittipornlertrak, A.; Muenthaisong, A.; Koonyosying, P.; Tangtrongsup, S.; Tiwananthagorn, S.; Chung, Y.T.; Sthitmatee, N. Effect of GP19 peptide hyperimmune antiserum on activated macrophage during Ehrlichia canis infection in canine macrophage-like cells. Animals 2021, 11, 2310. [Google Scholar] [CrossRef]
- Peleg, O.; Baneth, G.; Eyal, O.; Inbar, J.; Harrus, S. Multiplex real-time qPCR for the detection of Ehrlichia canis and Babesia canis vogeli. Vet. Parasitol. 2010, 173, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Crocquet-Valdes, P.A.; Thirumalapura, N.R.; Ismail, N.; Yu, X.; Saito, T.B.; Stevenson, H.L.; Pietzsch, C.A.; Thomas, S.; Walker, D.H. Immunization with Ehrlichia P28 outer membrane proteins confers protection in a mouse model of ehrlichiosis. Clin. Vaccine Immunol. 2011, 18, 2018–2025. [Google Scholar] [CrossRef]
- Thomas, S.; Thirumalapura, N.R.; Crocquet-Valdes, P.A.; Luxon, B.A.; Walker, D.H. Structure-based vaccines provide protection in a mouse model of ehrlichiosis. PLoS ONE 2011, 6, e27981. [Google Scholar] [CrossRef]
- Ohashi, N.; Zhi, N.; Zhang, Y.; Rikihisa, Y. Immunodominant major outer membrane proteins of Ehrlichia chaffeensis are encoded by a polymorphic multigene family. Infect. Immun. 1998, 66, 132–139. [Google Scholar] [CrossRef]
- Li, J.S.; Yager, E.; Reilly, M.; Freeman, C.; Reddy, G.R.; Reilly, A.A.; Chu, F.K.; Winslow, G.M. Outer membrane protein-specific monoclonal antibodies protect SCID mice from fatal infection by the obligate intracellular bacterial pathogen Ehrlichia chaffeensis. J. Immunol. 2001, 166, 1855–1862. [Google Scholar] [CrossRef]
- Nandi, B.; Hogle, K.; Vitko, N.; Winslow, G.M. CD4 T-cell epitopes associated with protective immunity induced following vaccination of mice with an ehrlichial variable outer membrane protein. Infect. Immun. 2007, 75, 5453–5459. [Google Scholar] [CrossRef]
- Barnewall, R.E.; Rikihisa, Y. Abrogation of gamma interferon-induced inhibition of Ehrlichia chaffeensis infection in human monocytes with iron-transferrin. Infect. Immun. 1994, 62, 4804–4810. [Google Scholar] [CrossRef]
- Tajima, T.; Wada, M. Inhibitory effect of interferon gamma on frequency of Ehrlichia canis-infected cells in vitro. Vet. Immunol. Immunopathol. 2013, 156, 200–204. [Google Scholar] [CrossRef]
- Bach, E.A.; Aguet, M.; Schreiber, R.D. The IFN gamma receptor: A paradigm for cytokine receptor signaling. Annu. Rev. Immunol. 1997, 15, 563–591. [Google Scholar] [CrossRef]
- Dhama, K.; Mahendran, M.; Chauhan, R.S.; Tomar, S. Cytokines: Their functional roles and prospective in veterinary practice—A review. Vet. Immunol. Immunopathol. 2008, 10, 79–89. [Google Scholar]
- Ferreira, V.L.; Borba, H.H.L.; Bonetti, A.F.; Leonart, L.P.; Pontarolo, R. Cytokines and Interferons: Types and Functions. Autoantibodies Cytokines 2018, 13, 65–87. [Google Scholar]
- Esteves, I.; Vachiéry, N.; Martinez, D.; Totté, P. Analysis of Ehrlichia ruminantium–specific T1/T2 responses during vaccination with a protective killed vaccine and challenge of goats. Parasite Immunol. 2004, 26, 95–103. [Google Scholar] [CrossRef]
- Riches, D.W.H.; Chan, E.D.; Winston, B.W. Chapter 7—TNF-α-induced regulation and signalling in macrophages. Immunobiology 1996, 195, 477–490. [Google Scholar] [CrossRef]
- Varma, T.K.; Toliver-Kinsky, T.E.; Lin, C.Y.; Koutrouvelis, A.P.; Nichols, J.E.; Sherwood, E.R. Cellular mechanisms that cause suppressed gamma interferon secretion in endotoxin-tolerant mice. Infect. Immun. 2001, 69, 5249–5263. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Control of adaptive immunity by the innate immune system. Nature. Immunol. 2015, 16, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Van Den Eeckhout, B.; Tavernier, J.; Gerlo, S. Interleukin-1 as innate mediator of T cell Immunity. Front. Immunol. 2011, 11, 621931. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number of Mice/Group | Immunogen Formulation | Challenge Exposure | Day of Euthanasia Post-Infection (PI) |
|---|---|---|---|---|
| 1 | 5 | Montanide (50 µL) * | ✓ | 7 PI |
| 2 | 5 | Montanide (50 µL) * | ✓ | 14 PI |
| 3 | 5 | 50 µg rGP19 protein (200 µL) with Montanide (50 µL) * | ✓ | 7 PI |
| 4 | 5 | 50 µg rGP19 protein (200 µL) with Montanide (50 µL) * | ✓ | 14 PI |
| 5 | 5 | 100 µg rGP19 protein (200 µL) with Montanide (50 µL) * | ✓ | 7 PI |
| 6 | 5 | 100 µg rGP19 protein (200 µL) with Montanide (50 µL) * | ✓ | 14 PI |
| 7 | 5 | No immunization (control) | 🗶 | 7 PI |
| 8 | 5 | No immunization (control) | 🗶 | 14 PI |
| Primer Name | Oligonucleotide Sequence (5′–3′) | Target Gene | Product Size |
|---|---|---|---|
| mIFNG-F mIFNG-R | 5′-TAT TGT CGC TTC TGG CTC CT-3′ 5′-AGA CTT ACG GCT GGC TTT GA-3′ | IFNG | 227 |
| mIL1a-F mIL1a-R | 5′-TCA AAG CCC AAA GGA AGC TA-3′ 5′-AGC TGA CTG CTC TGG GGA TA-3′ | IL-1a | 177 |
| mIL4-F mIL4-R | 5′-CTG GGC TTT CCT AGC TGA TG-3′ 5′-CTC TGT GGG GCA ATA CCT GT-3′ | IL-4 | 214 |
| mIL6-F mIL6-R | 5′-CAG AGG ATA CCA CTC CCA ACA-3′ 5′-TCC AGT TTG GTA GCA TCC ATC-3′ | IL-6 | 203 |
| mTNF-F mTNF-R | 5′-CAT GCG TCC AGC TGA CTA AA-3′ 5′-TCC CCT TCA TCT TCC TCC TT-3′ | TNF | 182 |
| mGAPDH-F mGAPDH-R | 5′-ACC CAG AAG ACT GTG GAT GG-3′ 5′-CAC ATT GGG GGT AGG AAC AC-3′ | GAPDH | 171 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nambooppha, B.; Rittipornlertrak, A.; Muenthaisong, A.; Koonyosying, P.; Chomjit, P.; Sangkakam, K.; Tangtrongsup, S.; Tiwananthagorn, S.; Sthitmatee, N. Recombinant Ehrlichia canis GP19 Protein as a Promising Vaccine Prototype Providing a Protective Immune Response in a Mouse Model. Vet. Sci. 2022, 9, 386. https://doi.org/10.3390/vetsci9080386
Nambooppha B, Rittipornlertrak A, Muenthaisong A, Koonyosying P, Chomjit P, Sangkakam K, Tangtrongsup S, Tiwananthagorn S, Sthitmatee N. Recombinant Ehrlichia canis GP19 Protein as a Promising Vaccine Prototype Providing a Protective Immune Response in a Mouse Model. Veterinary Sciences. 2022; 9(8):386. https://doi.org/10.3390/vetsci9080386
Chicago/Turabian StyleNambooppha, Boondarika, Amarin Rittipornlertrak, Anucha Muenthaisong, Pongpisid Koonyosying, Paweena Chomjit, Kanokwan Sangkakam, Sahatchai Tangtrongsup, Saruda Tiwananthagorn, and Nattawooti Sthitmatee. 2022. "Recombinant Ehrlichia canis GP19 Protein as a Promising Vaccine Prototype Providing a Protective Immune Response in a Mouse Model" Veterinary Sciences 9, no. 8: 386. https://doi.org/10.3390/vetsci9080386
APA StyleNambooppha, B., Rittipornlertrak, A., Muenthaisong, A., Koonyosying, P., Chomjit, P., Sangkakam, K., Tangtrongsup, S., Tiwananthagorn, S., & Sthitmatee, N. (2022). Recombinant Ehrlichia canis GP19 Protein as a Promising Vaccine Prototype Providing a Protective Immune Response in a Mouse Model. Veterinary Sciences, 9(8), 386. https://doi.org/10.3390/vetsci9080386