
Perfluorooctanoic Acid (PFOA) Induces Redox Status Disruption in Swine Granulosa Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of Granulosa Cells
2.1.1. Granulosa Cell Viability
2.1.2. Granulosa Cell Redox Status
Granulosa Cell Superoxide (O2−) Production
Granulosa Cell Hydrogen Peroxide (H2O2) Production
Granulosa Cell Nitric Oxide (NO) Production
Non-Enzymatic Scavenging Activity
2.2. Statistical Analysis
3. Results
3.1. Effect of PFOA on Swine Granulosa Cells Viability
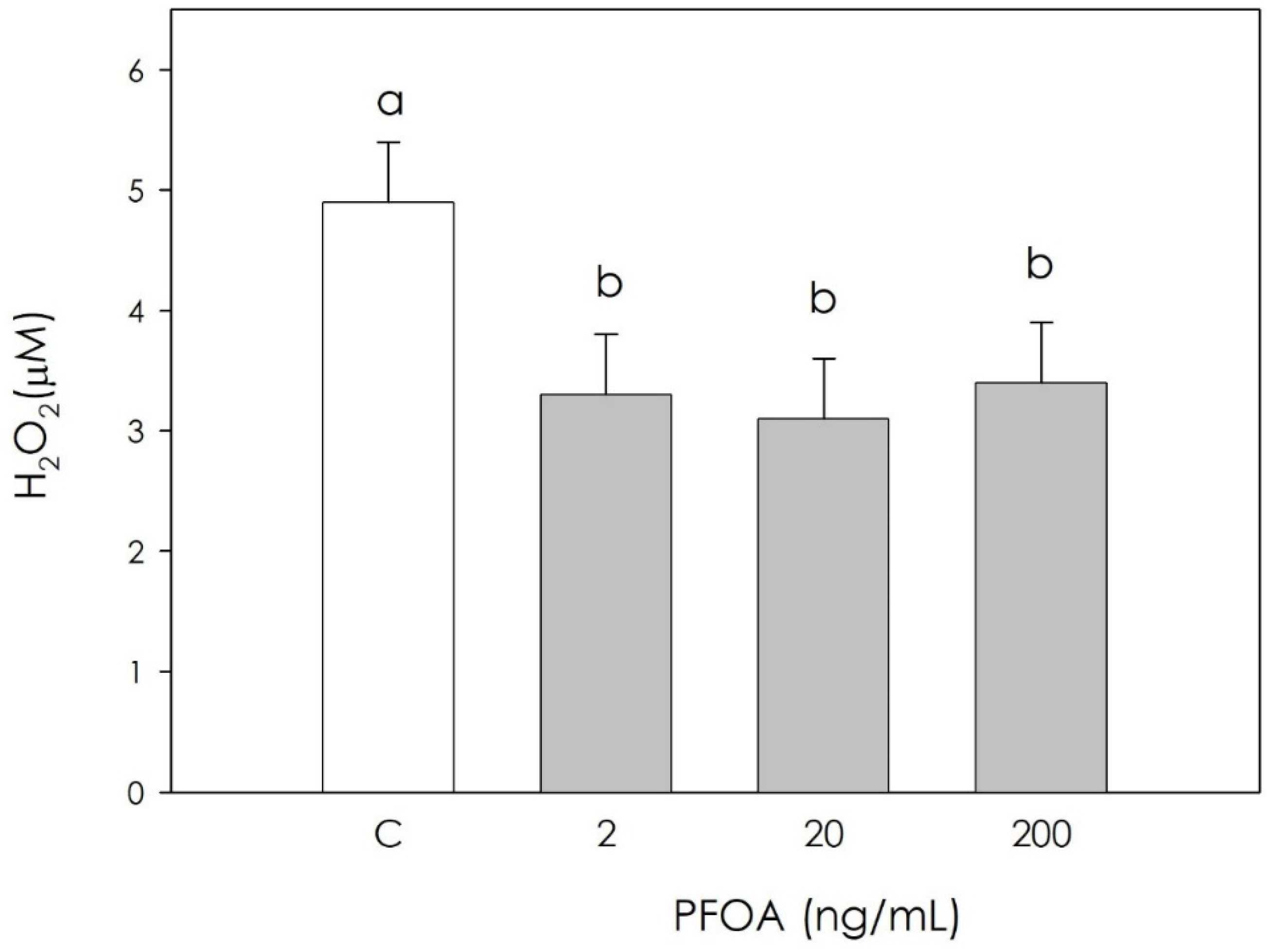
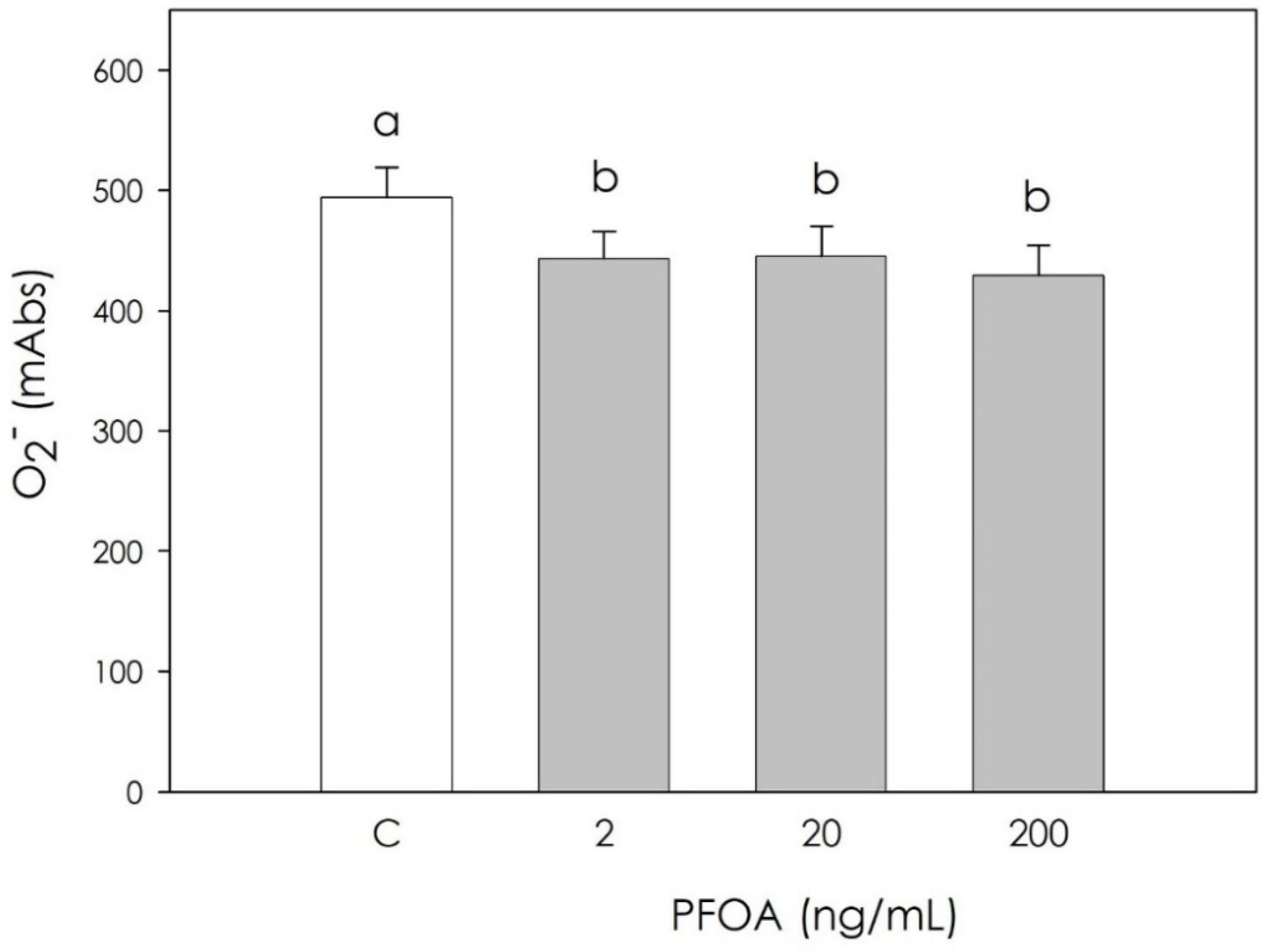
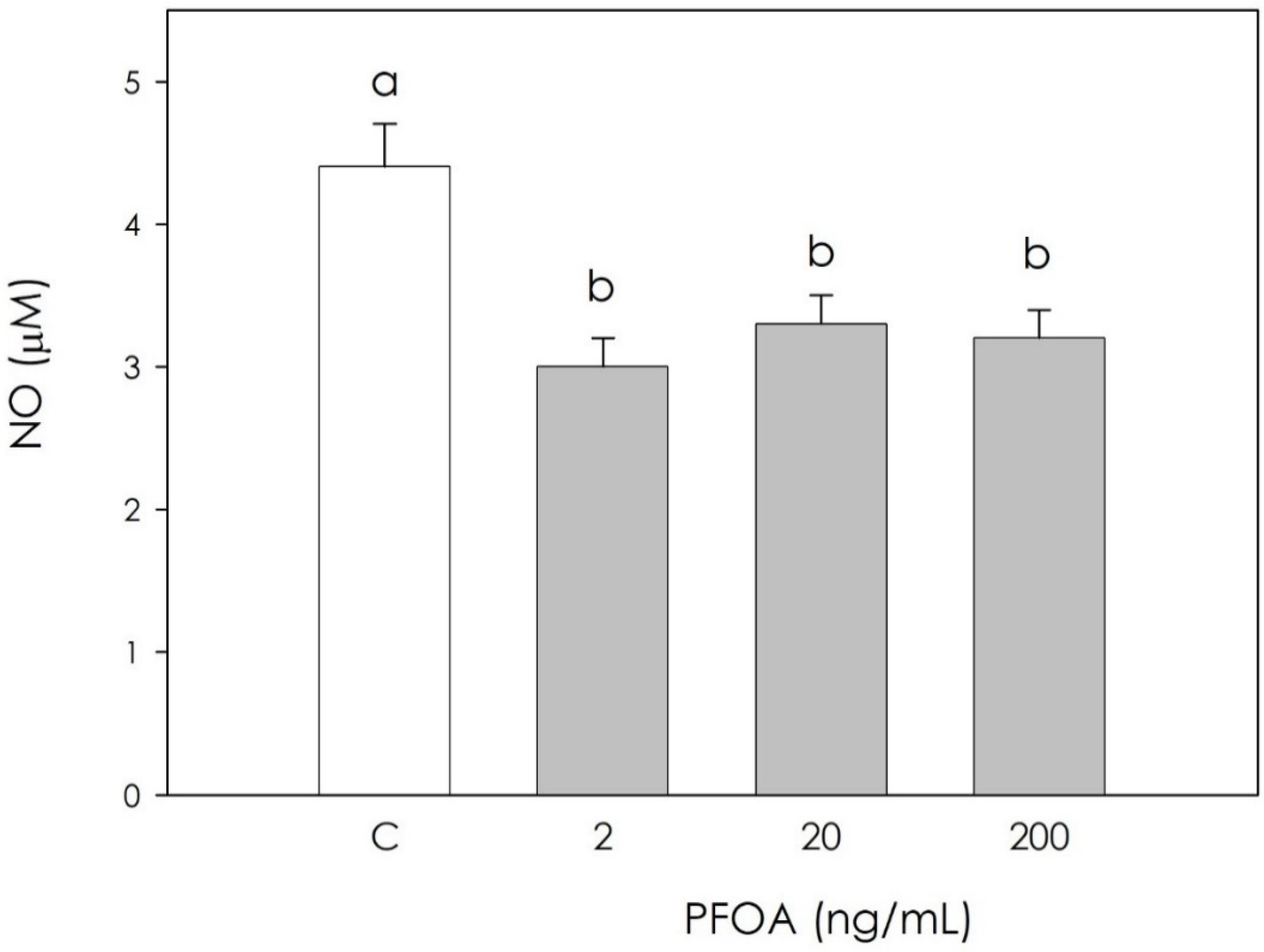
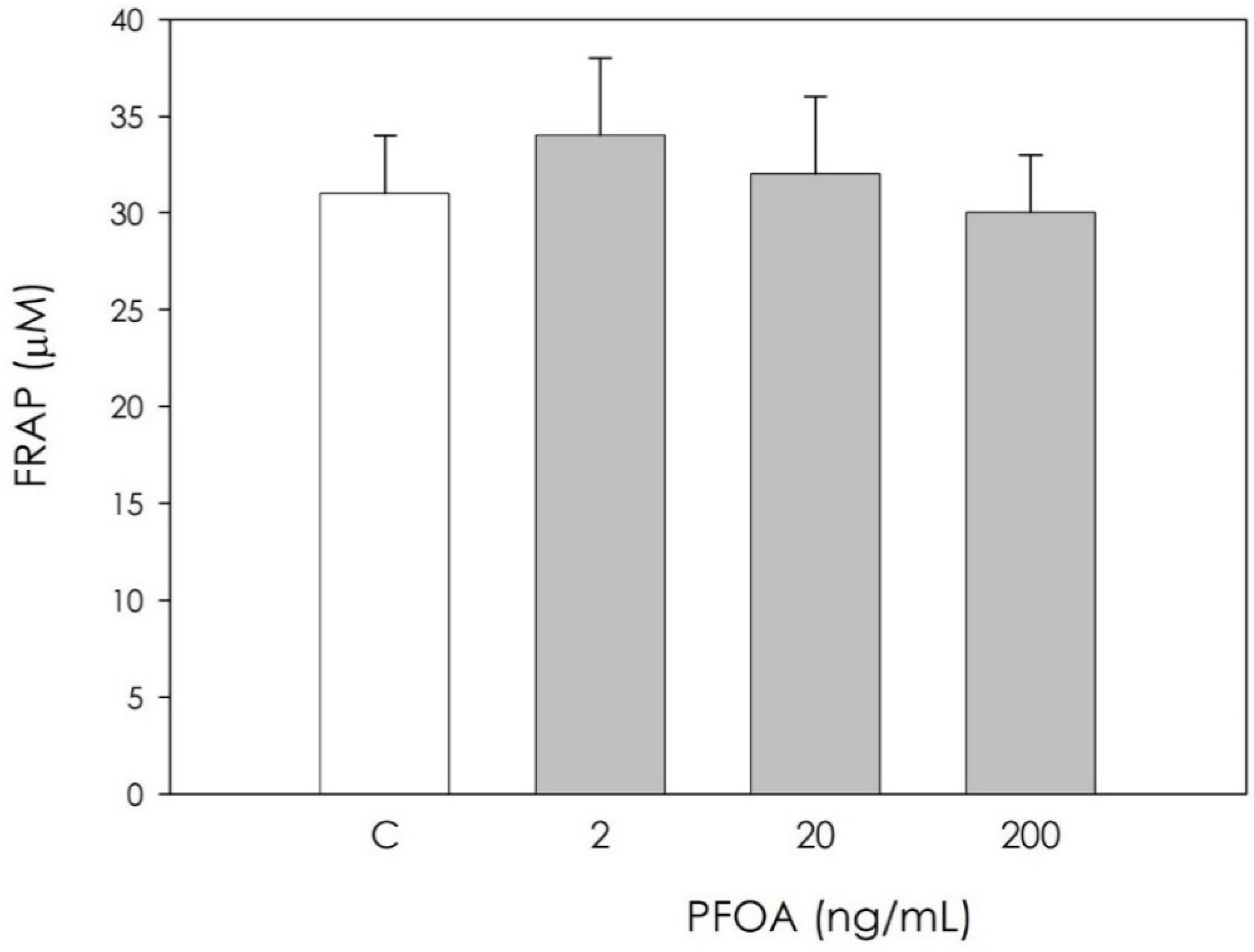
3.2. Effect of PFOA on Swine Granulosa Cells Redox Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evich, M.G.; Davis, M.J.B.; McCord, J.P.; Acrey, B.; Awkerman, J.A.; Knappe, D.R.U.; Lindstrom, A.B.; Speth, T.F.; Tebes-Stevens, C.; Strynar, M.J.; et al. Per-and polyfluoroalkyl substances in the environment. Science 2022, 375, eabg9065. [Google Scholar] [CrossRef]
- DeLuca, N.M.; Minucci, J.M.; Mullikin, A.; Slover, R.; Cohen Hubal, E.A. Human exposure pathways to poly- and perfluoroalkyl substances (PFAS) from indoor media: A systematic review. Environ. Int. 2022, 162, 107149. [Google Scholar] [CrossRef]
- Eichler, C.M.A.; Bi, C.; Wang, C.; Little, J.C. A modular mechanistic framework for estimating exposure to SVOCs: Next steps for modeling emission and partitioning of plasticizers and PFAS. J. Expo. Sci. Environ. Epidemiol. 2022, 32, 356–365. [Google Scholar] [CrossRef]
- Milinovic, J.; Lacorte, S.; Vidal, M.; Rigol, A. Sorption behaviour of perfluoroalkyl substances in soils. Scien. Total Environ. 2015, 511, 63–71. [Google Scholar] [CrossRef]
- Beale, D.J.; Nilsson, S.; Bose, U.; Bourne, N.; Stockwell, S.; Broadbent, J.A.; Gonzalez-Astudillo, V.; Braun, C.; Baddiley, B.; Limpus, D.; et al. Bioaccumulation and impact of maternal PFAS offloading on egg biochemistry from wild-caught freshwater turtles (Emydura macquarii macquarii). Sci. Total Environ. 2022, 817, 153019. [Google Scholar] [CrossRef]
- Fu, J.; Gao, Y.; Cui, L.; Wang, T.; Liang, Y.; Qu, G.; Yuan, B.; Wang, Y.; Zhang, A.; Jiang, G. Occurrence, temporal trends, and half-lives of perfluoroalkyl acids (PFAAs) in occupational workers in China. Sci. Rep. 2016, 6, 38039. [Google Scholar] [CrossRef] [Green Version]
- Vanden Heuvel, J.P.; Kuslikis, B.I.; Van Rafelghem, M.J.; Peterson, R.E. Tissue distribution, metabolism, and elimination of perfluorooctanoic acid in male and female rats. J. Biochem. Toxicol. 1991, 6, 83–92. [Google Scholar] [CrossRef]
- Fenton, S.E.; Reiner, J.L.; Nakayama, S.F.; Delinsky, A.D.; Stanko, J.P.; Hines, E.P.; White, S.S.; Lindstrom, A.B.; Strynar, M.J.; Petropoulou, S.E. Analysis of PFOA in dosed CD-1 mice. Part 2. Disposition of PFOA in tissues and fluids from pregnant and lactating mice and their pups. Reprod. Toxicol. 2009, 27, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Di Nisio, A.; Sabovic, I.; Valente, U.; Tescari, S.; Rocca, M.S.; Guidolin, D.; Dall’Acqua, S.; Acquasaliente, L.; Pozzi, N.; Plebani, M.; et al. Endocrine Disruption of Androgenic Activity by Perfluoroalkyl Substances: Clinical and Experimental Evidence. J. Clin. Endocrinol. Metab. 2019, 104, 1259–1271. [Google Scholar] [CrossRef] [Green Version]
- Basini, G.; Baioni, L.; Bussolati, S.; Grasselli, F.; Daquino, C.; Spatafora, C.; Tringali, C. Antiangiogenic properties of an unusual benzo[k,l]xanthene lignan derived from CAPE (caffeic acid phenethyl ester). Investig. New Drugs 2012, 30, 186–190. [Google Scholar] [CrossRef]
- Basini, G.; Bussolati, S.; Ciccimarra, R.; Grasselli, F. Melatonin potentially acts directly on swine ovary by modulating granulosa cell function and angiogenesis. Reprod. Fertil. Dev. 2017, 29, 2305–2312. [Google Scholar] [CrossRef]
- Basini, G.; Bussolati, S.; Andriani, L.; Grolli, S.; Ramoni, R.; Bertini, S.; Iemmi, T.; Menozzi, A.; Berni, P.; Grasselli, F. Nanoplastics impair in vitro swine granulosa cell functions. Domest. Anim. Endocrinol. 2021, 76, 106611. [Google Scholar] [CrossRef]
- Tumbleson, M.E.; Schook, L.B. Advances in Swine in Biomedical Research; Plenum Press: New York, NY, USA, 1996. [Google Scholar]
- Timóteo-Ferreira, F.; Abreu, D.; Mendes, S.; Matos, L.; Rodrigues, A.R.; Almeida, H.; Silva, E. Redox imbalance in age-related ovarian dysfunction and perspectives for its prevention. Ageing Res. Rev. 2021, 68, 101345. [Google Scholar] [CrossRef]
- Basini, G.; Tamanini, C. Interrelationship between nitric oxide and prostaglandins in bovine granulosa cells. Prostaglandins Other Lipid. Mediat. 2001, 66, 179–202. [Google Scholar] [CrossRef]
- Foxcroft, G.R.; Hunter, M.G. Basic physiology of follicular maturation in the pig. J. Reprod. Fertil. Suppl. 1985, 33, 1–19. [Google Scholar]
- Basini, G.; Falasconi, I.; Bussolati, S.; Grolli, S.; Di Lecce, R.; Grasselli, F. Swine granulosa cells show tipical endothelial cell characteristics. Reprod. Sci. 2016, 23, 630–637. [Google Scholar] [CrossRef]
- Basini, G.; Baioni, L.; Bussolati, S.; Grolli, S.; Kramer, L.H.; Wagner, G.F.; Grasselli, F. Expression and localization of stanniocalcin 1 in swine ovary. Gen. Comp. Endocrinol. 2010, 166, 404–408. [Google Scholar] [CrossRef]
- Basini, G.; Bianchi, F.; Bussolati, S.; Baioni, L.; Ramoni, R.; Grolli, S.; Conti, V.; Bianchi, F.; Grasselli, F. Atrazine disrupts steroidogenesis, VEGF and NO production in swine granulosa cells. Ecotoxicol. Environ. Saf. 2012, 85, 59–63. [Google Scholar] [CrossRef]
- Gogola, J.; Hoffmann, M.; Ptak, A. Persistent endocrine-disrupting chemicals found in human follicular fluid stimulate the proliferation of granulosa tumor spheroids via GPR30 and IGF1R but not via the classic estrogen receptors. Chemosphere 2019, 217, 100–110. [Google Scholar] [CrossRef]
- Gogola, J.; Hoffmann, M.; Ptak, A. Persistent endocrine-disrupting chemicals found in human follicular fluid stimulate IGF1 secretion by adult ovarian granulosa cell tumor spheroids and thereby increase proliferation of non-cancer ovarian granulosa cells. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2020, 65, 104769. [Google Scholar] [CrossRef]
- Basini, G.; Simona, B.; Santini, S.E.; Grasselli, F. Reactive oxygen species and anti-oxidant defences in swine follicular fluids. Reprod. Fertil. Dev. 2008, 20, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.L.; Yallampalli, C. Interaction between nitric oxide and prostaglandin E2 pathways in pregnant rat uteri. Am. J. Physiol. Endocrinol. Metab. 1996, 270, E471–E476. [Google Scholar] [CrossRef]
- Basini, G.; Santini, S.E.; Bussolati, S.; Grasselli, F. The phytoestrogen quercetin impairs steroidogenesis and angiogenesis in swine granulosa cells in vitro. J. Biomed. Biotechnol. 2009, 2009, 419891. [Google Scholar]
- Chambers, W.S.; Hopkins, J.G.; Richards, S.M. A Review of Per-and Polyfluorinated Alkyl Substance Impairment of Reproduction. Front. Toxicol. 2021, 3, 732436. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, K.; Marynowicz, W.; Gogola-Mruk, J.; Jakubowska, K.; Tworzydło, W.; Opydo-Chanek, M.; Ptak, A. A mixture of persistent organic pollutants detected in human follicular fluid increases progesterone secretion and mitochondrial activity in human granulosa HGrC1 cells. Reprod. Toxicol. 2021, 104, 114–124. [Google Scholar] [CrossRef]
- Sun, S.; Guo, H.; Wang, J.; Dai, J. Hepatotoxicity of perfluorooctanoic acid and two emerging alternatives based on a 3D spheroid model. Environ. Pollut. 2019, 246, 955–962. [Google Scholar] [CrossRef]
- Souders, C.L.; Sanchez, C.L.; Malphurs, W.; Aristizabal-Henao, J.J.; Bowden, J.A.; Martyniuk, C.J. Metabolic profiling in human SH-SY5Y neuronal cells exposed to perfluorooctanoic acid (PFOA). Neurotoxicology 2021, 85, 160–172. [Google Scholar] [CrossRef]
- López-Arellano, P.; López-Arellano, K.; Luna, J.; Flores, D.; Jiménez-Salazar, J.; Gavia, G.; Teteltitla, M.; Rodríguez, J.J.; Domínguez, A.; Casas, E.; et al. Perfluorooctanoic acid disrupts gap junction intercellular communication and induces reactive oxygen species formation and apoptosis in mouse ovaries. Environ. Toxicol. 2019, 34, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhou, L.; Xu, J.; Zhang, L.; Li, M.; Xie, X.; Xie, Y.; Luo, D.; Zhang, D.; Yu, X.; et al. Maternal exposure to perfluorooctanoic acid inhibits luteal function via oxidative stress and apoptosis in pregnant mice. Reprod. Toxicol. 2017, 69, 159–166. [Google Scholar] [CrossRef]
- Teteltitla, M.; Ducolomb, Y.; Souza, V.; Dominguez, A.; Rodriguez-Mercado, J.; Flores, D.; Bonilla, E.; Casas, E.; Altamirano, M.; Lopez, A.; et al. Effects of perfluorooctanoic acid in oxidative stress generation, DNA damage in cumulus cells, and its impact on in vitro maturation of porcine oocytes. Environ. Toxicol. 2022, 37, 1394–1403. [Google Scholar]
- Xu, M.; Cui, Z.; Zhao, L.; Hu, S.; Zong, W.; Liu, R. Characterizing the binding interactions of PFOA and PFOS with catalase at the molecular level. Chemosphere 2018, 203, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Suh, K.S.; Choi, E.M.; Kim, Y.J.; Hong, S.M.; Park, S.Y.; Rhee, S.Y.; Oh, S.; Kim, S.W.; Pak, Y.K.; Choe, W.; et al. Perfluorooctanoic acid induces oxidative damage and mitochondrial dysfunction in pancreatic β-cells. Mol. Med. Rep. 2017, 15, 3871–3878. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basini, G.; Bussolati, S.; Torcianti, V.; Grasselli, F. Perfluorooctanoic Acid (PFOA) Induces Redox Status Disruption in Swine Granulosa Cells. Vet. Sci. 2022, 9, 254. https://doi.org/10.3390/vetsci9060254
Basini G, Bussolati S, Torcianti V, Grasselli F. Perfluorooctanoic Acid (PFOA) Induces Redox Status Disruption in Swine Granulosa Cells. Veterinary Sciences. 2022; 9(6):254. https://doi.org/10.3390/vetsci9060254
Chicago/Turabian StyleBasini, Giuseppina, Simona Bussolati, Veronica Torcianti, and Francesca Grasselli. 2022. "Perfluorooctanoic Acid (PFOA) Induces Redox Status Disruption in Swine Granulosa Cells" Veterinary Sciences 9, no. 6: 254. https://doi.org/10.3390/vetsci9060254
APA StyleBasini, G., Bussolati, S., Torcianti, V., & Grasselli, F. (2022). Perfluorooctanoic Acid (PFOA) Induces Redox Status Disruption in Swine Granulosa Cells. Veterinary Sciences, 9(6), 254. https://doi.org/10.3390/vetsci9060254