
Genetic Diversity, Biofilm Formation, and Antibiotic Resistance of Pseudomonas aeruginosa Isolated from Cow, Camel, and Mare with Clinical Endometritis

 ,
,  ,
,  , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Samples
2.3. Bacterial Isolation
2.4. Molecular Conformation of the Isolates
2.5. Molecular Typing
2.6. Antimicrobial Susceptibility Testing
2.7. Phenotypic Detection of ESBLs and MBLs
2.8. Detection of Selected Resistance Genes
2.9. Biofilm Formation
2.10. Molecular Detection of Selected Virulence Factors and Quorum-Sensing (QS) Genes
2.11. Statistical Analysis
3. Results
3.1. Bacterial Isolation and Identification
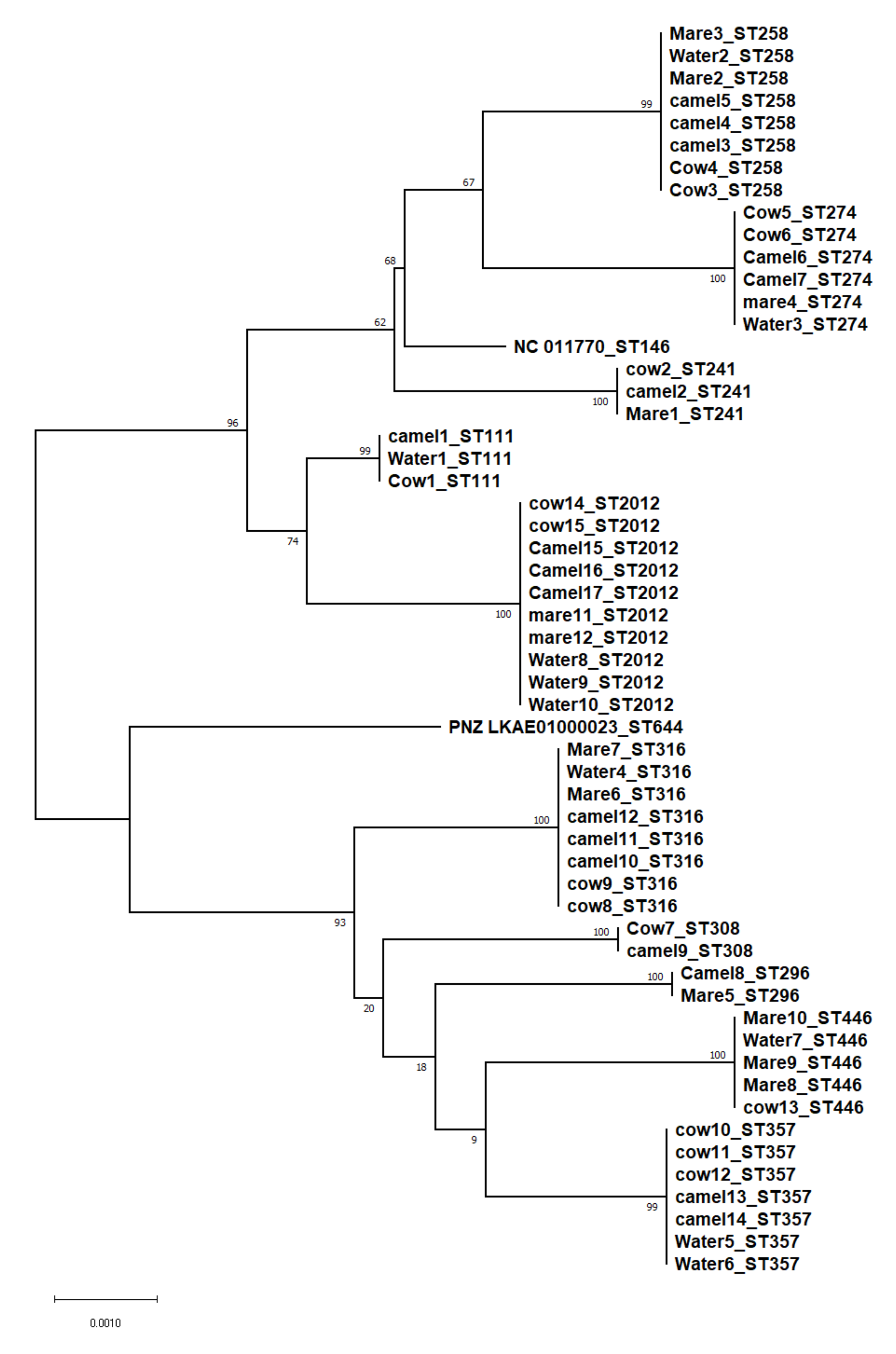
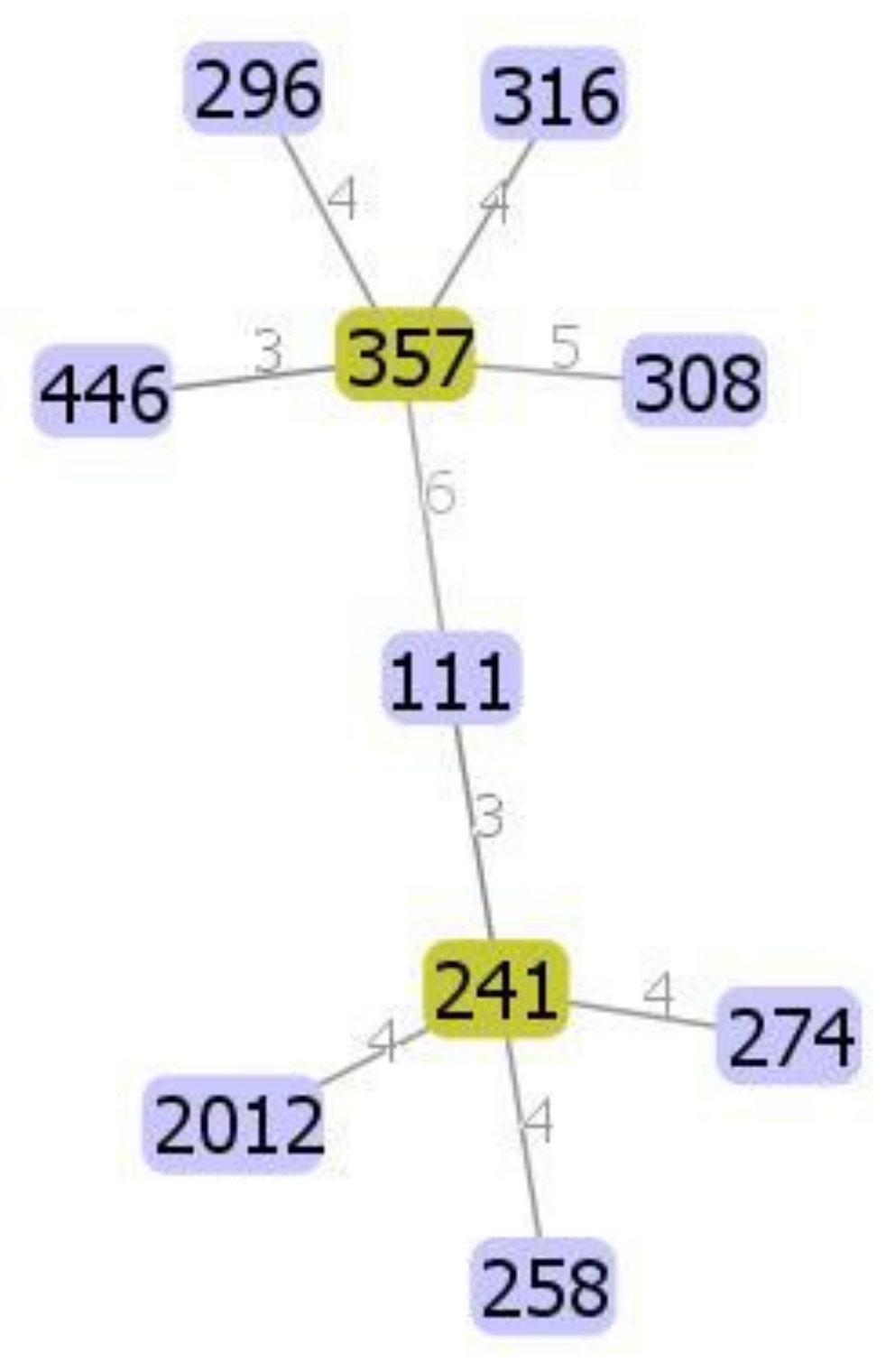
3.2. Molecular Typing
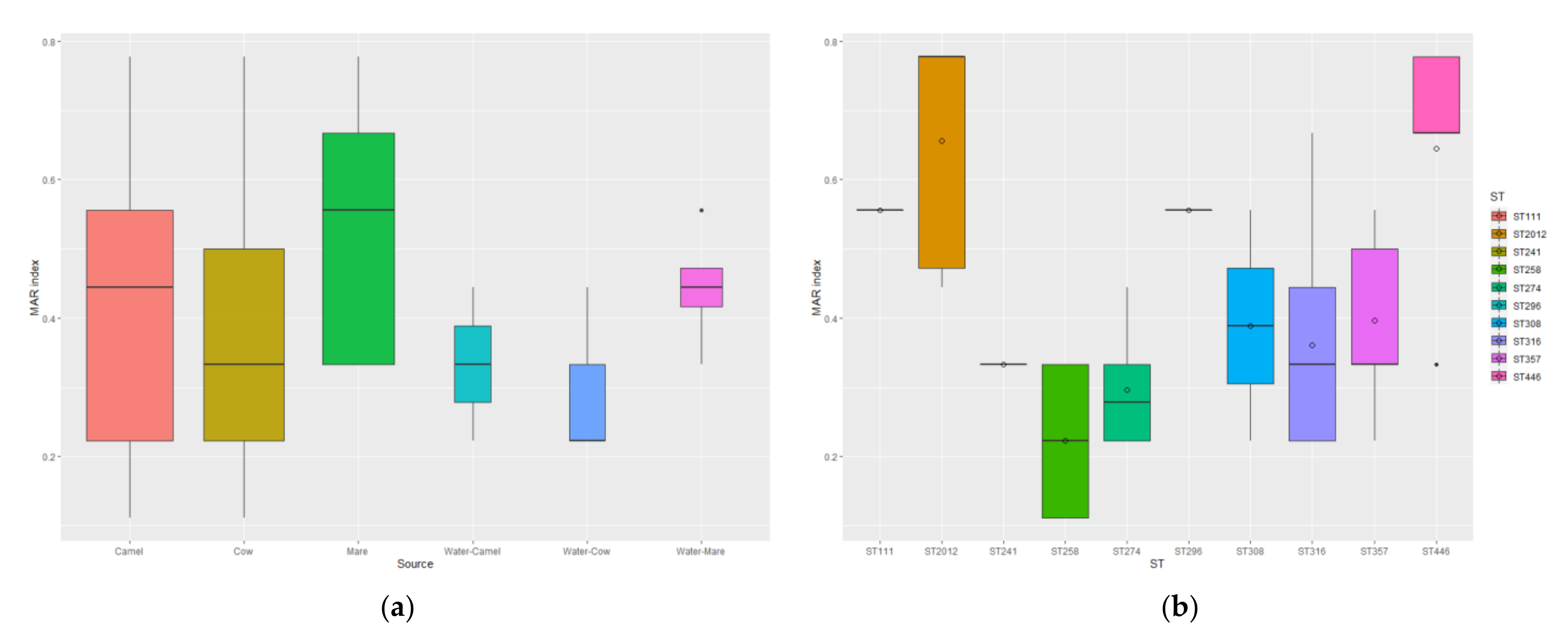
3.3. Antimicrobial Susceptibility
3.4. Phenotypic and Genotypic Detection of ESBLs and MBLs
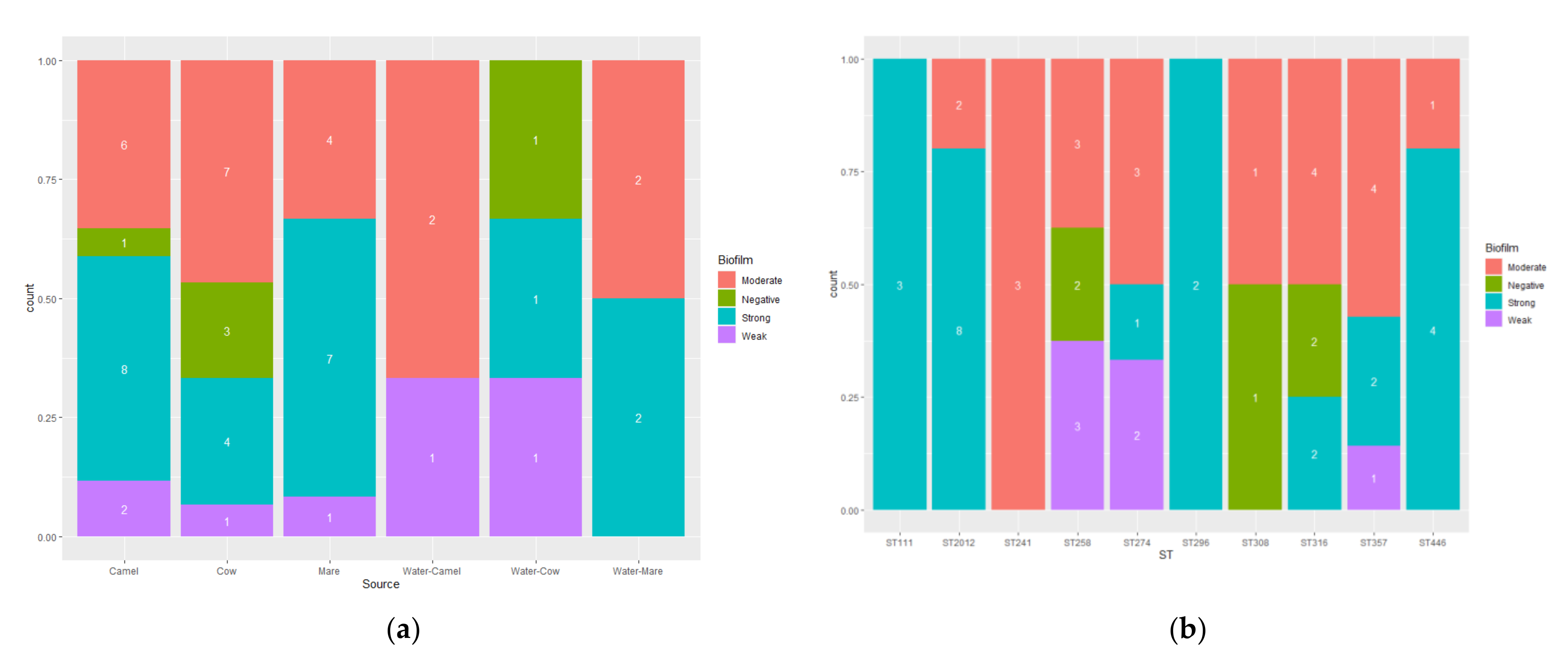
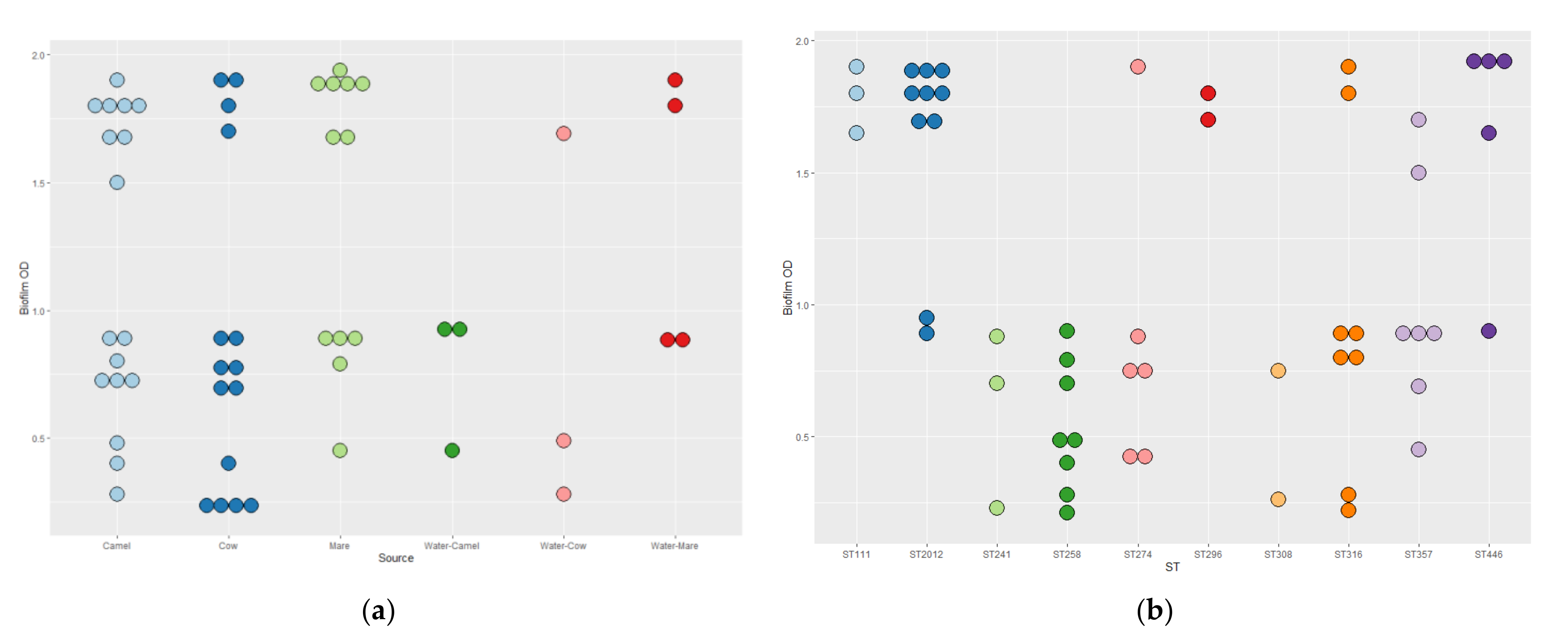
3.5. Biofilm Formation
3.6. Molecular Detection of Selected Virulence Factors and Quorum-Sensing (QS) Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cabot, G.; Zamorano, L.; Moya, B.; Juan, C.; Navas, A.; Blazquez, J.; Oliver, A. Evolution of Pseudomonas aeruginosa Antimicrobial Resistance and Fitness under Low and High Mutation Rates. Antimicrob. Agents Chemother. 2016, 60, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, P.; Davis, E.; Yu, F.; James, S.; Wildschutte, J.H.; Wiegmann, D.D.; Sherman, D.H.; McKay, R.M.; LiPuma, J.J.; Wildschutte, H. Environmental Pseudomonads Inhibit Cystic Fibrosis Patient-Derived Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2017, 83, e02701–e02716. [Google Scholar] [CrossRef] [PubMed]
- Arai, H. Regulation and Function of Versatile Aerobic and Anaerobic Respiratory Metabolism in Pseudomonas aeruginosa. Front. Microbiol. 2011, 2, 103. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, M.W.; Maxwell, J.A.; Vincent, T.S.; da Silva, J.; Olson, J.C. Comparison of the exoS gene and protein expression in soil and clinical isolates of Pseudomonas aeruginosa. Infect. Immun. 2001, 69, 2198–2210. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef]
- Davies, J.C. Pseudomonas aeruginosa in cystic fibrosis: Pathogenesis and persistence. Paediatr. Respir. Rev. 2002, 3, 128–134. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Sellera, F.P.; Moura, Q.; Carvalho, M.P.N.; Rosato, P.N.; Cerdeira, L.; Lincopan, N. Zooanthroponotic Transmission of Drug-Resistant Pseudomonas aeruginosa, Brazil. Emerg. Infect. Dis. 2018, 24, 1160–1162. [Google Scholar] [CrossRef]
- Getahun, A.M.; Hunderra, G.C.; Gebrezihar, T.G.; Boru, B.G.; Desta, N.T.; Ayana, T.D. Comparative study on lesions of reproductive disorders of cows and female dromedary camels slaughtered at Addis Ababa, Adama and Akaki abattoirs with bacterial isolation and characterization. BMC Vet. Res. 2021, 17, 134. [Google Scholar] [CrossRef]
- De Lima, F.S. Recent advances and future directions for uterine diseases diagnosis, pathogenesis, and management in dairy cows. Anim. Reprod. 2020, 17, e20200063. [Google Scholar] [CrossRef]
- Mahnani, A.; Sadeghi-Sefidmazgi, A.; Cabrera, V.E. Consequences and economics of metritis in Iranian Holstein dairy farms. J. Dairy Sci. 2015, 98, 6048–6057. [Google Scholar] [CrossRef]
- Benaissa, M.H.; Faye, B.; Kaidi, R. Reproductive abnormalities in female camel (Camelus dromedarius) in Algeria: Relationship with age, season, breed and body condition score. J. Camel Pract. Res. 2015, 22, 67–73. [Google Scholar] [CrossRef]
- Canisso, I.F.; Segabinazzi, L.; Fedorka, C.E. Persistent Breeding-Induced Endometritis in Mares—A Multifaceted Challenge: From Clinical Aspects to Immunopathogenesis and Pathobiology. Int. J. Mol. Sci. 2020, 21, 1432. [Google Scholar] [CrossRef] [PubMed]
- Rajala, P.; Gröhn, Y.J. Effects of dystocia, retained placenta, and metritis on milk yield in dairy cows. J. Dairy Sci. 1998, 81, 3172–3181. [Google Scholar] [CrossRef]
- Gröhn, Y.; Rajala-Schultz, P.; Allore, H.; De Lorenzo, M.; Hertl, J.; Galligan, D.J. Optimizing replacement of dairy cows: Modeling the effects of diseases. Prev. Vet. Med. 2003, 61, 27–43. [Google Scholar] [CrossRef]
- Paisley, L.G.; Mickelsen, W.D.; Anderson, P.B. Mechanisms and therapy for retained fetal membranes and uterine infections of cows: A review. Theriogenology 1986, 25, 353–381. [Google Scholar] [CrossRef]
- Morris, L.H.; McCue, P.M.; Aurich, C.J.R. Equine endometritis: A review of challenges and new approaches. Reproduction 2020, 160, R95–R110. [Google Scholar] [CrossRef]
- Mshelia, G.; Abba, Y.; Voltaire, Y.; Akpojie, G.; Mohammed, H.; Aondona, D. Comparative uterine bacteriology and pathology of camels (Camelus dromedarius) and cows in north-eastern Nigeria. Comp. Clin. Pathol. 2013, 22, 1195–1200. [Google Scholar] [CrossRef]
- Tibary, A.; Abdelhaq, A.; Abdelmalek, S. Factors affecting reproductive performance of camels at the herd and individual level. In Proceedings of the Desertification Combat and Food Safety: The Added Value of Camel Producers, Ashkabad, Turkmenistan, 19–21 April 2005; pp. 97–114. [Google Scholar]
- Schauer, B.; Wald, R.; Urbantke, V.; Loncaric, I.; Baumgartner, M.J. Tracing Mastitis Pathogens—Epidemiological Investigations of a Pseudomonas aeruginosa Mastitis Outbreak in an Austrian Dairy Herd. Animals 2021, 11, 279. [Google Scholar] [CrossRef]
- Yeruham, I.; Elad, D.; Avidar, Y.; Goshen, T. A herd level analysis of urinary tract infection in dairy cattle. Vet. J. 2006, 171, 172–176. [Google Scholar] [CrossRef]
- Hall, J.L.; Holmes, M.A.; Baines, S.J. Prevalence and antimicrobial resistance of canine urinary tract pathogens. Vet. Rec. 2013, 173, 549. [Google Scholar] [CrossRef]
- Mekić, S.; Matanović, K.; Šeol, B.J.V.R. Antimicrobial susceptibility of Pseudomonas aeruginosa isolates from dogs with otitis externa. Vet. J. 2011, 169, 125. [Google Scholar] [CrossRef] [PubMed]
- Hillier, A.; Alcorn, J.R.; Cole, L.K.; Kowalski, J.J. Pyoderma caused by Pseudomonas aeruginosa infection in dogs: 20 cases. Vet. Dermatol. 2006, 17, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Speert, D.P. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and impact on treatment. Drug Resist. Update 2000, 3, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Collignon, P.; Conly, J.; Andremont, A.; World Health Organization Advisory Group, Bogotá Meeting on Integrated Surveillance of Antimicrobial Resistance (WHO-AGISAR). World Health Organization ranking of antimicrobials according to their importance in human medicine: A critical step for developing risk management strategies to control antimicrobial resistance from food animal production. Clin. Infect. Dis. 2016, 63, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Livermore, D.M.J. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare? Clin. Infect. Dis. 2002, 34, 634–640. [Google Scholar] [PubMed]
- Breidenstein, E.B.; de la Fuente-Núñez, C.; Hancock, R.E.J. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, R.A.; Szabo, D. Mechanisms of multidrug resistance in Acinetobacter species and Pseudomonas aeruginosa. Clin. Infect. Dis. 2006, 43 (Suppl. S2), S49–S56. [Google Scholar] [CrossRef]
- Weldhagen, G.F.; Poirel, L.; Nordmann, P. Ambler class A extended-spectrum beta-lactamases in Pseudomonas aeruginosa: Novel developments and clinical impact. Antimicrob. Agents Chemother. 2003, 47, 2385–2392. [Google Scholar] [CrossRef]
- Walsh, T.R.; Toleman, M.A.; Poirel, L.; Nordmann, P.J. Metallo-β-lactamases: The quiet before the storm? Clin. Microbiol. Rev. 2005, 18, 306–325. [Google Scholar] [CrossRef]
- Drenkard, E. Antimicrobial resistance of Pseudomonas aeruginosa biofilms. Microbes Infect. 2003, 5, 1213–1219. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Rasamiravaka, T.; Labtani, Q.; Duez, P.; El Jaziri, M. The formation of biofilms by Pseudomonas aeruginosa: A review of the natural and synthetic compounds interfering with control mechanisms. Biomed. Res. Int 2015, 2015, 759348. [Google Scholar] [CrossRef]
- De Kievit, T.R.; Iglewski, B.H. Bacterial quorum sensing in pathogenic relationships. Infect. Immun 2000, 68, 4839–4849. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Turner, K.E.; Kirienko, N.V.J. PqsA promotes pyoverdine production via biofilm formation. Pathogens 2018, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Storz, M.P.; Maurer, C.K.; Zimmer, C.; Wagner, N.; Brengel, C.; de Jong, J.C.; Lucas, S.; Müsken, M.; Häussler, S.; Steinbach, A.J. Validation of PqsD as an anti-biofilm target in Pseudomonas aeruginosa by development of small-molecule inhibitors. J. Am. Chem. Soc. 2012, 134, 16143–16146. [Google Scholar] [CrossRef] [PubMed]
- Curran, B.; Jonas, D.; Grundmann, H.; Pitt, T.; Dowson, C.G. Development of a multilocus sequence typing scheme for the opportunistic pathogen Pseudomonas aeruginosa. J. Clin. Microbiol. 2004, 42, 5644–5649. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Bassler, B.L.J. Quorum sensing signal–response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef]
- Wright, C.L. Management of Water Quality for Beef Cattle. Vet. Clin. N. Am. Food Anim. Pract. 2007, 23, 91–103. [Google Scholar] [CrossRef]
- Maes, S.; Vackier, T.; Nguyen Huu, S.; Heyndrickx, M.; Steenackers, H.; Sampers, I.; Raes, K.; Verplaetse, A.; De Reu, K. Occurrence and characterisation of biofilms in drinking water systems of broiler houses. BMC Microbiol. 2019, 19, 77. [Google Scholar] [CrossRef]
- Mena, K.D.; Gerba, C.P. Risk assessment of Pseudomonas aeruginosa in water. Rev. Environ. Contam. Toxicol. 2009, 201, 71–115. [Google Scholar]
- Waage, A.; Vardund, T.; Lund, V.; Kapperud, G.J. Detection of low numbers of Salmonella in environmental water, sewage and food samples by a nested polymerase chain reaction assay. J. Appl. Microbiol. 1999, 87, 418–428. [Google Scholar] [CrossRef] [PubMed]
- McAllister, T.; Topp, E.J. Role of livestock in microbiological contamination of water: Commonly the blame, but not always the source. Anim. Front. 2012, 2, 17–27. [Google Scholar] [CrossRef]
- Elaichouni, A.; Verschraegen, G.; Claeys, G.; Devleeschouwer, M.; Godard, C.; Vaneechoutte, M. Pseudomonas aeruginosa serotype O12 outbreak studied by arbitrary primer PCR. J. Clin. Microbiol. 1994, 32, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Denamur, E.; Picard, B.; Goullet, P.; Bingen, E.; Lambert, N.; Elion, J. Complexity of Pseudomonas aeruginosa infection in cystic fibrosis: Combined results from esterase electrophoresis and rDNA restriction fragment length polymorphism analysis. Epidemiol. Infect. 1991, 106, 531–539. [Google Scholar] [CrossRef]
- Johnson, J.K.; Arduino, S.M.; Stine, O.C.; Johnson, J.A.; Harris, A.D. Multilocus sequence typing compared to pulsed-field gel electrophoresis for molecular typing of Pseudomonas aeruginosa. J. Clin. Microbiol. 2007, 45, 3707–3712. [Google Scholar] [CrossRef]
- Doumith, M.; Alhassinah, S.; Alswaji, A.; Alzayer, M.; Alrashidi, E.; Okdah, L.; Aljohani, S.; Balkhy, H.H.; Alghoribi, M.F.; NGHA AMR Surveillance Group; et al. Genomic Characterization of Carbapenem-Non-susceptible Pseudomonas aeruginosa Clinical Isolates From Saudi Arabia Revealed a Global Dissemination of GES-5-Producing ST235 and VIM-2-Producing ST233 Sub-Lineages. Front. Microbiol. 2022, 12, 765113. [Google Scholar] [CrossRef]
- Al-Agamy, M.H.; Shibl, A.M.; Tawfik, A.F.; Elkhizzi, N.A.; Livermore, D.M. Extended-spectrum and metallo-beta-lactamases among ceftazidime-resistant Pseudomonas aeruginosa in Riyadh, Saudi Arabia. J. Chemother. 2012, 24, 97–100. [Google Scholar] [CrossRef]
- Al-Agamy, M.H.; Jeannot, K.; El-Mahdy, T.S.; Samaha, H.A.; Shibl, A.M.; Plésiat, P.; Courvalin, P. Diversity of Molecular Mechanisms Conferring Carbapenem Resistance to Pseudomonas aeruginosa Isolates from Saudi Arabia. Can. J. Infect. Dis. Med. Microbiol. 2016, 2016, 4379686. [Google Scholar] [CrossRef]
- Balkhy, H.H.; Cunningham, G.; Chew, F.K.; Francis, C.; Al Nakhli, D.J.; Almuneef, M.A.; Memish, Z.A.J. Hospital-and community-acquired infections: A point prevalence and risk factors survey in a tertiary care center in Saudi Arabia. Int. J. Infect. Dis. 2006, 10, 326–333. [Google Scholar] [CrossRef]
- Yezli, S.; Shibl, A.M.; Livermore, D.M.; Memish, Z.A.J. Prevalence and antimicrobial resistance among Gram-negative pathogens in Saudi Arabia. J. Chemother. 2014, 26, 257–272. [Google Scholar] [CrossRef]
- Khan, M.A.; Faiz, A. Antimicrobial resistance patterns of Pseudomonas aeruginosa in tertiary care hospitals of Makkah and Jeddah. Ann. Saudi Med. 2016, 36, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Refaat, D.; Ali, A.; Saeed, E.M.; Al-Sobayil, F.; Al-Samri, A.; Elbehiry, A. Diagnostic evaluation of subclinical endometritis in dromedary camels. Anim. Reprod. Sci. 2020, 215, 106327. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, S.; Duffield, T.; Leslie, K.; Bateman, K.; Keefe, G.P.; Walton, J.; Johnson, W.J. Defining and diagnosing postpartum clinical endometritis and its impact on reproductive performance in dairy cows. J. Dairy Sci. 2002, 85, 2223–2236. [Google Scholar] [CrossRef]
- Hosu, M.C.; Vasaikar, S.D.; Okuthe, G.E.; Apalata, T.J. Detection of extended spectrum beta-lactamase genes in Pseudomonas aeruginosa isolated from patients in rural Eastern Cape Province, South Africa. Sci. Rep. 2021, 11, 7110. [Google Scholar] [CrossRef] [PubMed]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. CLSI Supplement M100, 31st ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Editorial: Assessing the antimicrobial susceptibility of bacteria obtained from animals. J. Antimicrob. Chemother. 2010, 65, 601–604. [Google Scholar] [CrossRef]
- Jarlier, V.; Nicolas, M.-H.; Fournier, G.; Philippon, A.J. Extended broad-spectrum β-lactamases conferring transferable resistance to newer β-lactam agents in Enterobacteriaceae: Hospital prevalence and susceptibility patterns. Clin. Infect. Dis. 1988, 10, 867–878. [Google Scholar] [CrossRef]
- Giakkoupi, P.; Vourli, S.; Polemis, M.; Kalapothaki, V.; Tzouvelekis, L.S.; Vatopoulos, A.C.J. Supplementation of growth media with Zn2+ facilitates detection of VIM-2-producing Pseudomonas aeruginosa. J. Clin. Microbiol. 2008, 46, 1568–1569. [Google Scholar] [CrossRef]
- Pitout, J.D.; Thomson, K.S.; Hanson, N.D.; Ehrhardt, A.F.; Moland, E.S.; Sanders, C.C. beta-Lactamases responsible for resistance to expanded-spectrum cephalosporins in Klebsiella pneumoniae, Escherichia coli, and Proteus mirabilis isolates recovered in South Africa. Antimicrob. Agents Chemother. 1998, 42, 1350–1354. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pitout, J.D.; Hossain, A.; Hanson, N.D. Phenotypic and molecular detection of CTX-M-beta-lactamases produced by Escherichia coli and Klebsiella spp. J. Clin. Microbiol. 2004, 42, 5715–5721. [Google Scholar] [CrossRef] [PubMed]
- Stepanovic, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukic, S.; Cirkovic, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Acta Pathol. Microbiol. Immunol. Scand. 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Golpayegani, A.; Nodehi, R.N.; Rezaei, F.; Alimohammadi, M.; Douraghi, M.J. Real-time polymerase chain reaction assays for rapid detection and virulence evaluation of the environmental Pseudomonas aeruginosa isolates. Mol. Biol. Rep. 2019, 46, 4049–4061. [Google Scholar] [CrossRef] [PubMed]
- Ghoneim, I.M.; Al-Ahmad, J.A.; Fayez, M.M.; El-Sabagh, I.M.; Humam, N.A.A.; Al-Eknah, M.M. Characterization of microbes associated with cervico-vaginal adhesion in the reproductive system of camels (Camelus dromedaries). Trop Anim. Health Prod. 2021, 53, 132. [Google Scholar] [CrossRef]
- Atherton, J.; Pitt, T.J. Types of Pseudomonas aeruginosa isolated from horses. Equine Vet. J. 1982, 14, 329–332. [Google Scholar] [CrossRef]
- Blanchard, T.L.; Kenney, R.M.; Timoney, P.J.J. Venereal disease. Vet. Clin. North Am. Equine Pract. 1992, 8, 191–203. [Google Scholar] [CrossRef]
- LeJeune, J.T.; Besser, T.E.; Merrill, N.L.; Rice, D.H.; Hancock, D.D. Livestock drinking water microbiology and the factors influencing the quality of drinking water offered to cattle. J. Dairy Sci. 2001, 84, 1856–1862. [Google Scholar] [CrossRef]
- Allen, J.L.; Begg, A.P.; Browning, G.F.J. Outbreak of equine endometritis caused by a genotypically identical strain of Pseudomonas aeruginosa. J. Vet.-Diagn. Investig. 2011, 23, 1236–1239. [Google Scholar] [CrossRef]
- Craun, G.F.; Brunkard, J.M.; Yoder, J.S.; Roberts, V.A.; Carpenter, J.; Wade, T.; Calderon, R.L.; Roberts, J.M.; Beach, M.J.; Roy, S.L. Causes of outbreaks associated with drinking water in the United States from 1971 to 2006. Clin. Microbiol. Rev. 2010, 23, 507–528. [Google Scholar] [CrossRef]
- Del Barrio-Tofiño, E.; López-Causapé, C.; Oliver, A. Pseudomonas aeruginosa epidemic high-risk clones and their association with horizontally-acquired β-lactamases: 2020 update. Int. J. Antimicrob. Agents 2020, 56, 106196. [Google Scholar] [CrossRef] [PubMed]
- Zowawi, H.M.; Syrmis, M.W.; Kidd, T.J.; Balkhy, H.H.; Walsh, T.R.; Al Johani, S.M.; Al Jindan, R.Y.; Alfaresi, M.; Ibrahim, E.; Al-Jardani, A.; et al. Identification of carbapenem-resistant Pseudomonas aeruginosa in selected hospitals of the Gulf Cooperation Council States: Dominance of high-risk clones in the region. J. Med. Microbiol. 2018, 67, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Al-Zahrani, I.A.; Al-Ahmadi, B.M. Dissemination of VIM-producing Pseudomonas aeruginosa associated with high-risk clone ST654 in a tertiary and quaternary hospital in Makkah, Saudi Arabia. J. Chemother. 2021, 33, 12–20. [Google Scholar] [CrossRef]
- Papagiannitsis, C.; Medvecky, M.; Chudejova, K.; Skalova, A.; Rotova, V.; Spanelova, P.; Jakubu, V.; Zemlickova, H.; Hrabak, J.J.B.; Czech Participants of the European Antimicrobial Resistance Surveillance Network. Molecular characterization of carbapenemase-producing Pseudomonas aeruginosa of Czech origin and evidence for clonal spread of extensively resistant sequence type 357 expressing IMP-7 metallo-β-lactamase. Antimicrob. Agents Chemother. 2017, 61, 75. [Google Scholar] [CrossRef] [PubMed]
- Kainuma, A.; Momiyama, K.; Kimura, T.; Akiyama, K.; Inoue, K.; Naito, Y.; Kinoshita, M.; Shimizu, M.; Kato, H.; Shime, N.J.; et al. An outbreak of fluoroquinolone-resistant Pseudomonas aeruginosa ST357 harboring the exoU gene. J. Infect. Chemother. 2018, 24, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Pelegrin, A.C.; Saharman, Y.R.; Griffon, A.; Palmieri, M.; Mirande, C.; Karuniawati, A.; Sedono, R.; Aditianingsih, D.; Goessens, W.H.; van Belkum, A.J.M. High-risk international clones of carbapenem-nonsusceptible pseudomonas aeruginosa endemic to Indonesian intensive care units: Impact of a multifaceted infection control intervention analyzed at the genomic level. mBio 2019, 10, e02384-19. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi-Akiyama, T.; Tada, T.; Ohmagari, N.; Viet Hung, N.; Tharavichitkul, P.; Pokhrel, B.M.; Gniadkowski, M.; Shimojima, M.; Kirikae, T.J. Emergence and spread of epidemic multidrug-resistant Pseudomonas aeruginosa. Genome Biol. Evol. 2017, 9, 3238–3245. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Xu, S.; Tang, Z.; Li, Z.; Zhang, L. Use of antimicrobials in food animals and impact of transmission of antimicrobial resistance on humans. Biosaf. Health 2021, 3, 32–38. [Google Scholar] [CrossRef]
- Malinowski, E.; Lassa, H.; Markiewicz, H.; Kaptur, M.; Nadolny, M.; Niewitecki, W.; Ziętara, J.J.T. Sensitivity to antibiotics of Arcanobacterium pyogenes and Escherichia coli from the uteri of cows with metritis/endometritis. Veter J. 2011, 187, 234–238. [Google Scholar] [CrossRef]
- Fishman, N.J. Antimicrobial stewardship. Am. J. Infect. Control. 2006, 34, S55–S63. [Google Scholar] [CrossRef]
- Ozawa, T.; Kiku, Y.; Mizuno, M.; Inumaru, S.; Kushibiki, S.; Shingu, H.; Matsubara, T.; Takahashi, H.; Hayashi, T.J. Effect of intramammary infusion of rbGM-CSF on SCC and expression of polymorphonuclear neutrophil adhesion molecules in subclinical mastitis cows. Vet. Res. Commun. 2012, 36, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Eliasi, U.L.; Sebola, D.; Oguttu, J.W.; Qekwana, D.N. Antimicrobial resistance patterns of Pseudomonas aeruginosa isolated from canine clinical cases at a veterinary academic hospital in South Africa. J. S. Afr. Vet. Assoc. 2020, 91, e1–e6. [Google Scholar] [CrossRef] [PubMed]
- Gad, G.F.; el-Domany, R.A.; Ashour, H.M. Antimicrobial susceptibility profile of Pseudomonas aeruginosa isolates in Egypt. J. Urol. 2008, 180, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Yayan, J.; Ghebremedhin, B.; Rasche, K.J. Antibiotic resistance of Pseudomonas aeruginosa in pneumonia at a single university hospital center in Germany over a 10-year period. PLoS ONE 2015, 10, e0139836. [Google Scholar] [CrossRef]
- Nordmann, P.; Guibert, M.J. Extended-spectrum beta-lactamases in Pseudomonas aeruginosa. J. Antimicrob. Chemother. 1998, 42, 128–131. [Google Scholar] [CrossRef]
- Pechère, J.C.; Köhler, T. Patterns and modes of beta-lactam resistance in Pseudomonas aeruginosa. Clin. Microbiol. Infect. 1999, 5 (Suppl. S1), S15–S18. [Google Scholar] [CrossRef]
- Abdulhaq, N.; Nawaz, Z.; Zahoor, M.A.; Siddique, A.B. Association of biofilm formation with multi drug resistance in clinical isolates of Pseudomonas aeruginosa. EXCLI J. 2020, 19, 201–208. [Google Scholar] [CrossRef]
- Oliveira, L.; Medeiros, C.; Silva, I.; Monteiro, A.; Leite, C.; Carvalho, C.J. Susceptibilidade a antimicrobianos de bactérias isoladas de otite externa em cães. Arq. Bras. Med. Vet. Zootec. 2005, 57, 405–408. [Google Scholar] [CrossRef]
- Dégi, J.; Moțco, O.-A.; Dégi, D.M.; Suici, T.; Mareș, M.; Imre, K.; Cristina, R.T.J.A. Antibiotic susceptibility profile of Pseudomonas aeruginosa canine isolates from a multicentric study in Romania. Antibiotics 2021, 10, 846. [Google Scholar] [CrossRef]
- Haenni, M.; Bour, M.; Châtre, P.; Madec, J.-Y.; Plésiat, P.; Jeannot, K. Resistance of Animal Strains of Pseudomonas aeruginosa to Carbapenems. Front. Microbiol. 2017, 8, 1847. [Google Scholar] [CrossRef]
- Webb, H.E.; Bugarel, M.; den Bakker, H.C.; Nightingale, K.K.; Granier, S.A.; Scott, H.M.; Loneragan, G.H. Carbapenem-Resistant Bacteria Recovered from Faeces of Dairy Cattle in the High Plains Region of the USA. PLoS ONE 2016, 11, e0147363. [Google Scholar] [CrossRef] [PubMed]
- Isgren, C.J. Improving clinical outcomes via responsible antimicrobial use in horses. Equine Veter-Educ. 2021, 33, 653–658. [Google Scholar] [CrossRef]
- Elshafiee, E.A.; Nader, S.M.; Dorgham, S.M.; Hamza, D.A. Carbapenem-resistant Pseudomonas Aeruginosa Originating from Farm Animals and People in Egypt. J. Vet. Res. 2019, 63, 333–337. [Google Scholar] [CrossRef]
- Endimiani, A.; Hujer, K.M.; Hujer, A.M.; Bertschy, I.; Rossano, A.; Koch, C.; Gerber, V.; Francey, T.; Bonomo, R.A.; Perreten, V.J. Acinetobacter baumannii isolates from pets and horses in Switzerland: Molecular characterization and clinical data. J. Antimicrob. Chemother. 2011, 66, 2248–2254. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, B.W.; Nayak, R.; Boothe, D.M. Emergence of a New Delhi Metallo-β-Lactamase (NDM-1)-Encoding Gene in Clinical Escherichia coli Isolates Recovered from Companion Animals in the United States. Antimicrob. Agents Chemother. 2013, 57, 2902–2903. [Google Scholar] [CrossRef] [PubMed]
- Stolle, I.; Prenger-Berninghoff, E.; Stamm, I.; Scheufen, S.; Hassdenteufel, E.; Guenther, S.; Bethe, A.; Pfeifer, Y.; Ewers, C.J. Emergence of OXA-48 carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in dogs. J. Antimicrob. Chemother. 2013, 68, 2802–2808. [Google Scholar] [CrossRef]
- Boerlin, P.; Eugster, S.; Gaschen, F.; Straub, R.; Schawalder, P.J. Transmission of opportunistic pathogens in a veterinary teaching hospital. Vet. Microbiol. 2001, 82, 347–359. [Google Scholar] [CrossRef]
- Guardabassi, L.; Schwarz, S.; Lloyd, D.H. Pet animals as reservoirs of antimicrobial-resistant bacteria: Review. J. Antimicrob. Chemother. 2004, 54, 321–332. [Google Scholar] [CrossRef]
- Leite-Martins, L.; Meireles, D.; Bessa, L.J.; Mendes, Â.; de Matos, A.J.; Martins da Costa, P.J. Spread of multidrug-resistant Enterococcus faecalis within the household setting. Microb. Drug Resist. 2014, 20, 501–507. [Google Scholar] [CrossRef]
- Yao, H.; Wu, D.; Lei, L.; Shen, Z.; Wang, Y.; Liao, K.J. The detection of fosfomycin resistance genes in Enterobacteriaceae from pets and their owners. Vet. Microbiol. 2016, 193, 67–71. [Google Scholar] [CrossRef]
- Zhao, W.H.; Hu, Z.Q. Beta-lactamases identified in clinical isolates of Pseudomonas aeruginosa. Crit. Rev. Microbiol. 2010, 36, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Niu, H.; Chen, G.; Li, M.; Li, M.; Zhou, Y. Prevalence of ESBLs-producing Pseudomonas aeruginosa isolates from different wards in a Chinese teaching hospital. Int. J. Clin. Exp. Med. 2015, 8, 19400–19405. [Google Scholar] [PubMed]
- Elhariri, M.; Hamza, D.; Elhelw, R.; Dorgham, S.M. Extended-spectrum beta-lactamase-producing Pseudomonas aeruginosa in camel in Egypt: Potential human hazard. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Al-Agamy, M.H.; Shibl, A.M.; Tawfik, A.F. Prevalence and molecular characterization of extended-spectrum beta-lactamase-producing Klebsiella pneumoniae in Riyadh, Saudi Arabia. Ann. Saudi Med. 2009, 29, 253–257. [Google Scholar] [CrossRef][Green Version]
- Tawfik, A.F.; Alswailem, A.M.; Shibl, A.M.; Al-Agamy, M.H. Prevalence and genetic characteristics of TEM, SHV, and CTX-M in clinical Klebsiella pneumoniae isolates from Saudi Arabia. Microb. Drug Resist. 2011, 17, 383–388. [Google Scholar] [CrossRef]
- Hocquet, D.; Plésiat, P.; Dehecq, B.; Mariotte, P.; Talon, D.; Bertrand, X. Nationwide Investigation of Extended-Spectrum β-Lactamases, Metallo-β-Lactamases, and Extended-Spectrum Oxacillinases Produced by Ceftazidime-Resistant Pseudomonas aeruginosa Strains in France. Antimicrob. Agents Chemother. 2010, 54, 3512–3515. [Google Scholar] [CrossRef]
- Tawfik, A.F.; Shibl, A.M.; Aljohi, M.A.; Altammami, M.A.; Al-Agamy, M.H.J.B. Distribution of Ambler class A, B and D β-lactamases among Pseudomonas aeruginosa isolates. Ann Burn. Fire Disasters 2012, 38, 855–860. [Google Scholar] [CrossRef]
- Zafer, M.M.; Al-Agamy, M.H.; El-Mahallawy, H.A.; Amin, M.A.; El Din Ashour, S.J. Dissemination of VIM-2 producing Pseudomonas aeruginosa ST233 at tertiary care hospitals in Egypt. BMC Infect. Dis. 2015, 15, 122. [Google Scholar] [CrossRef]
- Al Bayssari, C.; Dabboussi, F.; Hamze, M.; Rolain, J.-M.J. Emergence of carbapenemase-producing Pseudomonas aeruginosa and Acinetobacter baumannii in livestock animals in Lebanon. J. Antimicrob. Chemother. 2015, 70, 950–951. [Google Scholar] [CrossRef]
- Sjölander, I.; Hansen, F.; Elmanama, A.; Khayyat, R.; Abu-Zant, A.; Hussein, A.; Taha, A.A.; Hammerum, A.M.; Ciofu, O.J. Detection of NDM-2-producing Acinetobacter baumannii and VIM-producing Pseudomonas aeruginosa in Palestine. J. Glob. Antimicrob. Resist. 2014, 2, 93–97. [Google Scholar] [CrossRef]
- Ejikeugwu, C.; Esimone, C.; Iroha, I.; Eze, P.; Ugwu, M.; Adikwu, M.J.; Microbiology, A. Genotypic and phenotypic characterization of MBL genes in Pseudomonas aeruginosa isolates from the non-hospital environment. J. Pure Appl. Microbiol. 2018, 12, 1877–1885. [Google Scholar] [CrossRef]
- Al-Orphaly, M.; Hadi, H.A.; Eltayeb, F.K.; Al-Hail, H.; Samuel, B.G.; Sultan, A.A.; Skariah, S.J.M. Epidemiology of multidrug-resistant Pseudomonas aeruginosa in the Middle East and North Africa Region. mSphere 2021, 6, e00202–e00221. [Google Scholar] [CrossRef]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S.J. Epidemiology and treatment of multidrug-resistant and extensively drug-resistant Pseudomonas aeruginosa infections. Clin. Microbiol. Rev. 2019, 32, e00019–e00031. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Martis, N.; Leroy, S.; Blanc, V. Colistin in multi-drug resistant Pseudomonas aeruginosa blood-stream infections: A narrative review for the clinician. J. Infect. 2014, 69, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Kasiakou, S.K. Colistin: The revival of polymyxins for the management of multidrug-resistant gram-negative bacterial infections. Clin. Infect. Dis. 2005, 40, 1333–1341. [Google Scholar] [CrossRef]
- Li, J.; Nation, R.L.; Turnidge, J.D.; Milne, R.W.; Coulthard, K.; Rayner, C.R.; Paterson, D.L. Colistin: The re-emerging antibiotic for multidrug-resistant Gram-negative bacterial infections. Lancet. Infect. Dis. 2006, 6, 589–601. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef]
- Pachori, P.; Gothalwal, R.; Gandhi, P. Emergence of antibiotic resistance Pseudomonas aeruginosa in intensive care unit; a critical review. Genes Dis. 2019, 6, 109–119. [Google Scholar] [CrossRef]
- Vasiljević, Z.; Jovčić, B.; Ćirković, I.; Đukić, S.J. An examination of potential differences in biofilm production among different genotypes of Pseudomonas aeruginosa. Arch. Biol. Sci. 2014, 66, 117–121. [Google Scholar] [CrossRef]
- Ciofu, O.; Tolker-Nielsen, T.J. Tolerance and resistance of Pseudomonas aeruginosa biofilms to antimicrobial agents—How P. aeruginosa can escape antibiotics. Front. Microbiol. 2019, 913, 2164. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Zhang, L. The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell 2015, 6, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, Y.C.; Furlaneto, I.P.; Maciel, A.H.P.; Quaresma, A.J.P.G.; de Matos, E.C.O.; Conceição, M.L.; Vieira, M.C.d.S.; Brabo, G.L.d.C.; Sarges, E.d.S.N.F.; Lima, L.N.G.C.J. High prevalence of atypical virulotype and genetically diverse background among Pseudomonas aeruginosa isolates from a referral hospital in the Brazilian Amazon. PLoS ONE 2020, 15, e0238741. [Google Scholar] [CrossRef] [PubMed]
- Senturk, S.; Ulusoy, S.; Bosgelmez-Tinaz, G.; Yagci, A.J.T. Quorum sensing and virulence of Pseudomonas aeruginosa during urinary tract infections. J. Infect. Dev. Ctries. 2012, 6, 501–507. [Google Scholar] [CrossRef]
- Osman, K.; Orabi, A.; Elbehiry, A.; Hanafy, M.H.; Ali, A.M.J.F.M. Pseudomonas species isolated from camel meat: Quorum sensing-dependent virulence, biofilm formation and antibiotic resistance. Future Microbiol. 2019, 14, 609–622. [Google Scholar] [CrossRef]
- Hassuna, N.A.; Mandour, S.A.; Mohamed, E.S.J.I.; Resistance, D. Virulence constitution of multi-drug-resistant Pseudomonas aeruginosa in Upper Egypt. Future Microbiol. 2020, 13, 587. [Google Scholar] [CrossRef]
- Ruiz-Roldán, L.; Bellés, A.; Bueno, J.; Azcona-Gutiérrez, J.M.; Rojo-Bezares, B.; Torres, C.; Castillo, F.J.; Sáenz, Y.; Seral, C.J. Pseudomonas aeruginosa isolates from Spanish children: Occurrence in faecal samples, antimicrobial resistance, virulence, and molecular typing. BioMed Res. Int. 2018, 2018, 8060178. [Google Scholar] [CrossRef]
- Morales-Espinosa, R.; Delgado, G.; Espinosa, L.F.; Isselo, D.; Méndez, J.L.; Rodriguez, C.; Miranda, G.; Cravioto, A. Fingerprint Analysis and Identification of Strains ST309 as a Potential High Risk Clone in a Pseudomonas aeruginosa Population Isolated from Children with Bacteremia in Mexico City. Front. Microbiol. 2017, 8, 313. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Type | Cow | Camel | Mare | Total |
|---|---|---|---|---|
| Uterine swabs | 21.4 (15/70) | 28.3 (17/60) | 24 (12/50) | 24.4 (44/180) |
| Drinking water | 10 (3/30) | 10 (3/30) | 13.3 (4/30) | 11.1 (10/90) |
| Sequence Type (ST) | N (%) | Uterine Swab | Drinking Water | ||||
|---|---|---|---|---|---|---|---|
| Cow | Camel | Mare | Cow | Camel | Mare | ||
| 111 | 3 (5.56) bc | 1 | 1 | 0 | 0 | 1 | 0 |
| 241 | 3 (5.56) bc | 1 | 1 | 1 | 0 | 0 | 0 |
| 258 | 8 (14.82) ab | 2 | 3 | 2 | 0 | 1 | 0 |
| 274 | 6 (11.11) abc | 2 | 2 | 1 | 1 | 0 | 0 |
| 296 | 2 (3.70) c | 0 | 1 | 1 | 0 | 0 | 0 |
| 308 | 2 (3.70) c | 1 | 1 | 0 | 0 | 0 | 0 |
| 316 | 8 (14.82) ab | 2 | 3 | 2 | 0 | 0 | 1 |
| 357 | 7 (12.96) abc | 3 | 2 | 0 | 1 | 0 | 0 |
| 446 | 5 (9.26) abc | 1 | 0 | 3 | 0 | 0 | 1 |
| 2012 | 10 (18.52) a | 2 | 3 | 2 | 0 | 1 | 2 |
| Antimicrobials | Breakpoints | N (%) | No. of Resistant P. aeruginosa Isolates (%) | |||||
|---|---|---|---|---|---|---|---|---|
| Uterine Swab | Drinking Water | |||||||
| Cow | Camel | Mare | Cow | Camel | Mare | |||
| Piperacillin | S ≤ 16 R ≥ 128 | 42 (77.78) a | 11 (26.1) | 15 (35.7) | 10 (23.8) | 3 (7.1) | 1 (2.3) | 2 (4.7) |
| Piperacillin/Tazobactam | S ≤ 16 R ≥ 128 | 32 (59.26) b | 6 (18.7) | 10 (31.2) | 10 (31.2) | 1 (3.1) | 2 (6.2) | 3 (9.3) |
| Ceftazidime | S ≤ 8 R ≥ 32 | 21 (38.89) c | 6 (28.5) | 6 (28.5) | 7 (33.3) | 1 (4.7) | 0 (0) | 1 (4.7) |
| Aztreonam | S ≤ 8 R ≥ 32 | 21 (38.89) c | 6 (28.5) | 6 (28.5) | 7 (33.3) | 1 (4.7) | 0 (0) | 1 (4.7) |
| Imipenem | S ≤ 2 R ≥ 8 | 8 (14.82) d | 2 (25) | 3 (37.5) | 3 (37.5) | 0 (0) | 0 (0) | 0 (0) |
| Amikacin | S ≤ 16 R ≥ 64 | 27 (50) bc | 8 (29.6) | 9 (33.3) | 4 (14.8) | 2 (7.4) | 1 (3.7) | 3 (11.1) |
| Gentamicin | S ≤ 4 R ≥ 16 | 27 (50) bc | 8 (29.6) | 9 (33.3) | 4 (14.8) | 2 (7.4) | 1 (3.7) | 3 (11.1) |
| Ciprofloxacin | S ≤ 0.5 R ≥ 2 | 32 (59.25) b | 10 (31.2) | 10 (31.2) | 10 (31.2) | 1 (3.1) | 1 (3.1) | 0 (0) |
| Colistin | S ≤ 0.001 R ≥ 4 | 0 (0) e | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Resistance Pattern | MDR | No. | Uterine Swab | Drinking Water | ||||
|---|---|---|---|---|---|---|---|---|
| Cow | Camel | Mare | Cow | Camel | Mare | |||
| CIP | 1 | 1 | 0 | 0 | 0 | 0 | 0 | |
| GEN AMK | 3 | 0 | 0 | 2 | 0 | 0 | 1 | |
| GEN CIP AMK | 7 | 3 | 2 | 0 | 1 | 1 | 0 | |
| PIP | 2 | 0 | 2 | 0 | 0 | 0 | 0 | |
| PIP CIP | 6 | 3 | 3 | 0 | 0 | 0 | 0 | |
| PIP GEN AMK | 2 | 2 | 0 | 0 | 0 | 0 | 0 | |
| PIP TZP | 2 | 0 | 0 | 0 | 1 | 0 | 1 | |
| PIP TZP CIP | 3 | 0 | 0 | 3 | 0 | 0 | 0 | |
| PIP TZP GEN AMK | 4 | 0 | 2 | 0 | 0 | 0 | 2 | |
| PIP TZP GEN CIP AMK | MDR | 2 | 0 | 2 | 0 | 0 | 0 | 0 |
| PIP TZP CAZ ATM | MDR | 3 | 1 | 0 | 0 | 1 | 1 | 0 |
| PIP TZP CAZ ATM CIP | MDR | 3 | 0 | 0 | 3 | 0 | 0 | 0 |
| PIP TZP CAZ ATM GEN CIP AMK | MDR | 8 | 3 | 3 | 2 | 0 | 0 | 0 |
| PIP TZP CAZ ATM IPM | MDR | 5 | 2 | 3 | 0 | 0 | 0 | 0 |
| PIP TZP CAZ ATM IPM CIP | MDR | 3 | 0 | 0 | 2 | 0 | 1 | 0 |
| Profile | No. | Uterine Swab | Drinking Water | ||||
|---|---|---|---|---|---|---|---|
| Cow | Camel | Mare | Cow | Camel | Mare | ||
| bla TEM | 4 | 1 | 2 | 1 | 0 | 0 | 0 |
| bla SHV | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| bla SHV-blaCTX-M | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| bla TEM-blaCTX-M | 3 | 1 | 0 | 2 | 0 | 0 | 0 |
| bla TEM-blaCTX M-blaVIM | 4 | 0 | 2 | 2 | 0 | 0 | 0 |
| bla TEM-blaSHV-blaCTX-M | 4 | 1 | 1 | 1 | 0 | 0 | 1 |
| bla TEM-blaSHV-blaVIM | 3 | 2 | 0 | 1 | 0 | 0 | 0 |
| bla TEM-blaVIM | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoud, S.F.; Fayez, M.; Swelum, A.A.; Alswat, A.S.; Alkafafy, M.; Alzahrani, O.M.; Alsunaini, S.J.; Almuslem, A.; Al Amer, A.S.; Yusuf, S. Genetic Diversity, Biofilm Formation, and Antibiotic Resistance of Pseudomonas aeruginosa Isolated from Cow, Camel, and Mare with Clinical Endometritis. Vet. Sci. 2022, 9, 239. https://doi.org/10.3390/vetsci9050239
Mahmoud SF, Fayez M, Swelum AA, Alswat AS, Alkafafy M, Alzahrani OM, Alsunaini SJ, Almuslem A, Al Amer AS, Yusuf S. Genetic Diversity, Biofilm Formation, and Antibiotic Resistance of Pseudomonas aeruginosa Isolated from Cow, Camel, and Mare with Clinical Endometritis. Veterinary Sciences. 2022; 9(5):239. https://doi.org/10.3390/vetsci9050239
Chicago/Turabian StyleMahmoud, Samy F., Mahmoud Fayez, Ayman A. Swelum, Amal S. Alswat, Mohamed Alkafafy, Othman M. Alzahrani, Saleem J. Alsunaini, Ahmed Almuslem, Abdulaziz S. Al Amer, and Shaymaa Yusuf. 2022. "Genetic Diversity, Biofilm Formation, and Antibiotic Resistance of Pseudomonas aeruginosa Isolated from Cow, Camel, and Mare with Clinical Endometritis" Veterinary Sciences 9, no. 5: 239. https://doi.org/10.3390/vetsci9050239
APA StyleMahmoud, S. F., Fayez, M., Swelum, A. A., Alswat, A. S., Alkafafy, M., Alzahrani, O. M., Alsunaini, S. J., Almuslem, A., Al Amer, A. S., & Yusuf, S. (2022). Genetic Diversity, Biofilm Formation, and Antibiotic Resistance of Pseudomonas aeruginosa Isolated from Cow, Camel, and Mare with Clinical Endometritis. Veterinary Sciences, 9(5), 239. https://doi.org/10.3390/vetsci9050239